

Abstract
Our group has proposed that in contrast to chronic stress that can have harmful effects, the short-term (fight-or-flight) stress response (lasting for minutes to hours) is nature’s fundamental survival mechanism that enhances protection and performance under conditions involving threat/challenge/opportunity. Short-term stress enhances innate/primary, adaptive/secondary, vaccine-induced, and anti-tumor immune responses, and post-surgical recovery. Mechanisms and mediators include stress hormones, dendritic cell, neutrophil, macrophage, and lymphocyte trafficking/function and local/systemic chemokine and cytokine production. Short-term stress may also enhance mental/cognitive and physical performance through effects on brain, musculo-skeletal, and cardiovascular function, reappraisal of threat/anxiety, and training-induced stress-optimization. Therefore, short-term stress psychology/physiology could be harnessed to enhance immuno-protection, as well as mental and physical performance. This review aims to provide a conceptual framework and targets for further investigation of mechanisms and conditions under which the protective/adaptive aspects of short-term stress/exercise can be optimized/harnessed, and for developing pharmacological/biobehavioral interventions to enhance health/healing, and mental/cognitive/physical performance.
Keywords: eustress/distress, psycho-neuro-endocrine-immunology, wound healing, infection/autoimmune disease/cancer, sleep, stress reduction, stress optimization, amateur/elite athletes, performers/entertainers, armed forces/special operations forces
1. Introduction
Chronic or long-term stress has been shown to have numerous adverse effects on health [1; 2]. Many of these effects are mediated through stress actions on the immune system [3; 4; 5]. It is important to elucidate the psychological and biological mechanisms by which chronic stressors weaken health, exacerbate disease, or inhibit mental and physical performance because that could enable the development of biobehavioral and pharmacological treatments designed to ameliorate or eliminate the harmful effects of chronic stress. However, it is also important to appreciate that the process of evolution did not select for the biological stress response to sicken, handicap, or kill us, but rather to help us survive [6]. A psycho-physiological stress response is one of nature’s fundamental survival mechanisms. Without a fight-or-flight stress response, a lion has no chance of catching a gazelle, just as the gazelle has no chance of escape. Thus, during short-term stress, multiple physiological systems are activated to enable survival. Dhabhar et al. first proposed that just as the short-term stress response prepares the cardiovascular, musculoskeletal and neuroendocrine systems for fight or flight, under certain conditions, stress may similarly prepare the immune system and the brain for challenges (e.g., wounding, infection, figuring out an escape route, tackling a job interview, running a race, etc.) that may be imposed by a stressor (e.g., predator, or, in modern times, a medical/surgical procedure, professional opportunity, athletic competition, etc.) [7; 8; 9; 10; 11]. Since then, numerous studies have shown in humans and animals, that short-term stress experienced at the time of immune activation induces a significant enhancement of the ensuing immune response. Studies have also shown short-term stress induced enhancement of mental performance. We propose that it is important to investigate the adaptive mechanisms and effects of the short-term stress response and to harness the psychological and biological mechanisms of the adaptive stress response, to enhance protection or performance under conditions of threat, challenge, or opportunity.
2. Stress: Definition, mediators, and individual differences
Even though the word “stress” generally has negative connotations, stress is a familiar and ubiquitous aspect of life, being a stimulant for some, but a burden for many others. Numerous definitions have been proposed for the concept of stress, each focusing on aspects of an internal or external challenge, disturbance, or stimulus; on stimulus perception by an organism; or on a physiological response to the stimulus [12; 13; 14]. An integrated definition states that stress is a constellation of events, consisting of a stimulus (stressor), that precipitates a reaction in the brain (stress perception), that activates physiological fight or flight systems in the body (stress response) [9]. Psychological, physiological, physical, or exercise-related stressors all activate biological stress responses involving the release of factors in the systemic circulation and locally within central and peripheral tissues. In the periphery, the stress response consists of the “big three” stress hormones: norepinephrine and epinephrine that are released by the sympathetic nervous system, and cortisol, that is induced following activation of the hypothalamic-pituitary-adrenal axis. Virtually every cell in the body expresses receptors for one or more of these “big three” hormones, that induce changes in almost all cells and tissues and inform them about the presence of a stressor. The peripheral stress response also includes other neuroendocrine factors such as adrenocorticotropin (ACTH), vasopressin [15], and oxytocin [15; 16], and cytokines [17] such as interleukin-6 [18] and inteleukin-1beta [19]. In the periphery, similar biological stress responses are observed under conditions that require protection (e.g. attack by a predator) [20; 21], performance (e.g. making a speech, or taking an exam, running a race) [22; 23; 24], or pleasure (e.g. sexual intercourse) [25; 26; 27; 28]. However, different types of stressors can differentially affect the relative proportions and the magnitude and duration of elevation of factors induced in the systemic circulation during short-term stress. It is important to recognize, that the biological stress response is the only pathway through which a stressor can affect the body.
Protective versus harmful effects of stress
Dhabhar et al., first proposed that short-term or acute stress induced enhancement of immune function may be an adaptive psycho-physiological mechanism that enhances immune protection following wounding, infection, vaccination, and perhaps even in the context of some types of cancer [7; 11; 29; 30]. Although this idea may sound similar to Hans Selye’s concept of “eustress,” it must be noted that Selye defined “eustress” largely in terms of the nature of the stressor, (i.e., whether it was pleasant as opposed to noxious) but stated that eustress and distress both cause “damage,” the former causing less damage than the latter [31]. In contrast, Dhabhar et al., have defined “good” versus “bad” stress in terms of the duration of the biological stress response and its adaptive versus deleterious effects, and have stated that stress does not always have deleterious effects, and in some cases can even have beneficial effects on brain, body, and health [3; 4; 7; 11; 29; 30; 32; 33].
It is known that stress can be harmful when it is chronic or long lasting [1; 34; 35; 36], however, it is often overlooked that a stress response has salubrious adaptive effects in the short run [32; 37]. Therefore, a major distinguishing characteristic of stress is the duration of the biological stress response. Short-term stress has been defined as stress that lasts for a period of minutes to hours, and chronic stress as stress that persists for several hours per day for weeks or months [9]. Dysregulation of the circadian cortisol rhythm is one marker that appears to coincide with the deleterious effects of chronic stress [9; 38; 39]. The intensity of stress can be gauged by the peak levels of stress hormones, neurotransmitters, and other physiological changes such as increases in heart rate and blood pressure, and could affect the amount of time for which these changes persist during stress and following the cessation of stress.
It is important to note that there are significant individual differences in stress perception, processing, appraisal, and coping [32; 40]. Such differences could be the result of genetic as well as experiential factors. Individual differences become especially salient while studying human subjects because stress perception, processing, appraisal, and coping mechanisms can have significant effects on the kinetics and peak levels of circulating stress hormones and on the duration for which these hormone levels are elevated. Studies showing differences in stress hormone receptors, reactivity and peak levels [41; 42], adaptation to stress [43], and in distribution and activation of adrenal steroid receptors and corticosteroid binding globulin levels [41; 44], suggest that genetic, experiential, as well as environmental factors play a role in establishing individual differences [41; 43; 44; 45]. The ability of humans to generate and experience psychological stressors even in the absence of external stressors can result in long-term activation of the physiological stress response that often has deleterious effects. The magnitude and duration of elevations in stress hormones can have significant effects on immune cell distribution and function [4; 8; 46; 47].
3. Revision of the dogma that immune function is suppressed during stress in order to conserve energy for survival responses
When viewed from an evolutionary perspective, immunosuppression under all stress conditions would not be adaptive because stress is an intrinsic part of life for most organisms, and dealing successfully with stressors enables survival. Moreover, most selection pressures, the chisels of evolution, are stressors. The brain perceives stressors, warns the body of danger, and promotes survival (e.g., when a gazelle sees a charging lion, the gazelle’s brain detects a threat and orchestrates a physiological response that enables the gazelle to flee). Stressful experiences often result in wounding or infection. Therefore, immunoenhancement, rather than immunosuppression, would be adaptive during short-term stress because it is unlikely that millions of years of evolution would select for a system exquisitely sculpted to escape the jaws and claws of a lion only to succumb to wounds and microbes [7; 29; 30]. In other words, just as the short-term stress response prepares the cardiovascular, musculoskeletal, and neuroendocrine systems for fight-or-flight, it should also prepare the immune system for challenges (wounding or infection) that are likely to result from stressful encounters (attack by a predator).
In contrast to the above discussion, it was (and still is) erroneously believed by many that stress-induced suppression of immune function is adaptive because immunosuppression during short-term stress conserves energy that is required to deal with the immediate demands imposed by the stressor. However, most mechanisms of immunosuppression expend, rather than conserve, energy. Moreover, the immune system is often critically needed for responding immediately to the actions of the stress-inducing agent (e.g., wounding by a predator). Thus, while ovulation, copulation, or digestion can wait for the cessation of stress, the immune response is not similarly dispensable during times of stress. Immune activation is critical for responding to the immediate demands of a stressful situation, especially when the situation results in wounding or infection. Furthermore, the time course for many proposed mechanisms for stress-induced immunosuppression, such as inhibition of prostaglandin synthesis, cytokine production, or leukocyte proliferation [48] is significantly longer than that seen during acute stress. While conservation of energy may play a role in stress-induced immunosuppression under some conditions, it is not likely to do so under all conditions of stress.
The energy-conservation hypothesis has also been invoked to suggest that adaptive immunity is suppressed and that only innate immunity is enhanced during acute stress [49; 50]. The underlying assumption for this hypothesis is that only innate immune responses are required for, and capable of, effective immunoprotection on a short time scale, and that inhibiting adaptive immunity would make more energy available to the innate immune system. There are several reasons for considering a revision of these assumptions and hypotheses: First, while classifications such as “innate” and “adaptive” are useful for conceptualization of different types of immune responses, it is important to keep in mind that in vivo immune responses consist of intricate and synchronous interactions among numerous proteins, cytokines, and cell types that include components of what were traditionally thought to be separate “innate” versus “adaptive” systems [51]. In general, most, if not all, components of an immune response are galvanized into action following immune activation although different components may predominate during different phases of the immune response. Second, it must be appreciated that suppressing an immune response does not necessarily conserve energy and, in fact, may even require additional expenditure of energy (e.g. energy is consumed during synthesis and/or release of immunosuppressive factors or during apoptosis). Third, the “adaptive” immune system is not designed solely to fight challenges that the “innate” system fails to overcome, unlike what is often erroneously claimed. An important function of adaptive immunity is to “memorize” previously encountered antigens/pathogens and to increase the overall efficiency with which a total, in vivo immune response is mounted against the antigen/pathogen upon subsequent exposure. In many instances, antigens and pathogens that activate an immune response may be those that the organism has previously encountered. In such cases, surveillance-memory T cells play a critical role in conferring protection by initiating the immune-response cascade and the sooner they are activated the more robust the protection. Thus, it would be counterproductive to specifically waste energy and resources during stress to suppress specific and powerful adaptive immune responses that are driven by already-present memory lymphocytes that the organism has invested considerable amounts of energy to acquire in the first place, and then to maintain for most if not all of its lifespan.
A variant of the energy-conservation hypothesis has been proposed to explain a transient short-term stress induced decline in immune function observed under some conditions in some invertebrate organisms such as crickets [52]. It has been suggested that short-term stressors (e.g., a cricket being tied to a stick, which is an extremely unnatural condition that could induce very high intensity stress) lead to immunosuppression in crickets because of octopamine (the insect analog of norepinephrine) driven competition for specific factors that are required for both lipid-derived mobilization of energy as well as for immune activation. However, octopamine suppresses immune function in crickets, but enhances immunity in other organisms such as the tobacco hornworm and cockroaches [53], suggesting that the relationship between octopamine and immune function in invertebrates is diverse and complex. It is also important to recognize that stress-induced immuno-suppression in some organisms, such as crickets, may simply reflect the fact that these organisms have not experienced selection pressures for long-term survival following wounding or infection (which could be due to their very short life-spans), and therefore have not evolved independent mechanisms to simultaneously support both the mobilization of energy and immune function.
4. The immune triad: Immuno-protection, Immuno-pathology, and Immuno-regulation
While investigating or discussing immune responses, it is useful to categorize them in terms of their principal cellular and molecular components. For example, innate, adaptive, Th1, Th2, Th17, etc. immune responses are all defined in terms of their cellular and cytokine components. In addition to these categories, it is also useful to define immune responses in terms of their integrated, functional, end-effects. Therefore, we have proposed that immune responses can be categorized as being immuno-protective, immuno-pathological, and immuno-regulatory/inhibitory [3; 11; 33]. It is important to bear in mind that while all these categories provide useful constructs with which to organize ideas, concepts, and models, an overall in vivo immune response is likely to consist of several types of responses with varying amounts of dominance from each category. The composition and nature of an immune response is also affected by, and changes with, time. Three major types of immune responses are defined below in terms of their functional end effects:
Immuno-protective responses are defined as responses that promote efficient wound healing, eliminate infections and cancer, and mediate vaccine-induced immunological memory [3; 11; 33]. Key characteristics of immuno-protection involve active immune surveillance, a rapid and robust response upon immune activation, efficient clearance of the activating agent or pathogen, followed by rapid resolution. Immuno-protective responses are critical for completion of the proliferative and remodeling phases of wound healing. Wound healing is important not only for frank wounds where the initiating event is tissue damage itself, but also for tissue-intrinsic “wounds” where the initiating event is an immune response precipitated by intracellular infection during which there can be collateral tissue damage. Innate and/or adaptive Type-1 or Type-2 immune responses can all confer immuno-protection depending on the type of the pathogen (viral, bacterial, protozoan, fungal, helminthic), on whether it is intra- or extra-cellular, and on the accompanying wounding conditions (sterile, infected, external or internal wounds).
Immuno-pathological responses are defined as those that are directed against self- (autoimmune disease like multiple sclerosis, arthritis, lupus) or innocuous antigens (asthma, allergies) and responses that involving chronic, non-resolving inflammation [3; 11; 33]. Immuno-pathology is also involved during low-level, long-term elevations in local and/or systemic inflammatory mediators (e.g., CRP or IL-6) that are thought to contribute to disorders like cardiovascular disease, obesity, and depression [54; 55; 56].
Immuno-regulatory responses are defined as those that involve immune cells and factors that regulate (mostly down-regulate) the function of other immune cells [3; 11; 33]. Although the previous concept of suppressor T cells became mired in controversy, recent studies suggest that there is an arm of the immune system that functions to inhibit immune responses [57; 58; 59]. For example, regulatory CD4+CD25+FoxP3+ T cells, IL-10, and TGF-beta have been shown to have immuno-regulatory/inhibitory functions. The physiological function of these factors is to keep pro-inflammatory, allergic, and autoimmune responses in check [59; 60]. However, it has also been suggested that immuno-regulatory/inhibitory factors may suppress anti-tumor immunity and be indicative of negative prognosis for cancer [39; 61; 62; 63].
5. Factors that determine whether stress enhances or suppresses immune function, and the potential health consequences of these effects of stress
Key factors that determine whether stress enhances or suppresses immune function include [3; 9; 11; 32]: 1) Effects of stress on immune cell distribution in the body. 2) The duration of stress. 3) Differential effects of physiologic versus pharmacologic concentrations of glucocorticoids, and the differential effects of endogenous (e.g. cortisol, corticosterone) versus synthetic (e.g., dexamethasone) glucocorticoids. 4) The timing of stressor or stress hormone exposure relative to the time of activation and ensuing time course of the immune response. It is important to recognize that factors such as gender, genetics, age, the route of administration and nature of the immunizing antigen, and time during the circadian cycle, additionally affect immune function, and could also affect the nature of the relationship between stress and immune function. It is also important to bear in mind that whether a stressor enhances or suppresses immune function, it is the end-effect of the immune response that determines whether the stress-immune interactions have beneficial or harmful effects on health (Figure 1).
Figure 1. Enhancing versus suppressive effects of stress on immune function and potential consequences for health outcomes.
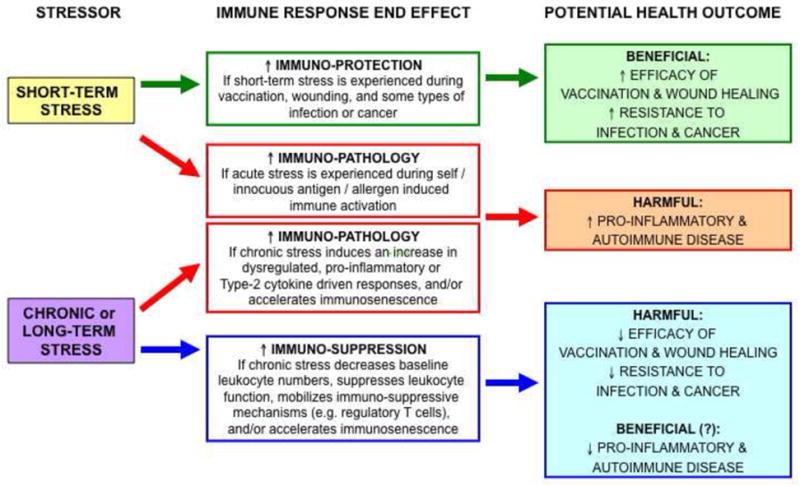
Short-term stress experienced during vaccination, wounding, or infection may enhance immuno-protective responses. Short-term stress experienced during immune activation in response to self/innocuous antigens or allergens, may excerbate pro-inflammatory and autoimmune disorders. Chronic stress-induced increases in pro-inflammatory or Type-2 cytokine mediated immune responses may also exacerbate inflammatory and autoimmune disease. Chronic stress induced suppression of immune responses may decrease the effectiveness of vaccination and wound healing and decrease resistance to infection and cancer. (Reprinted with permission from S. Karger AG, Basel: Enhancing versus suppressive effects of stress on immune function: implications for immunoprotection and immunopathology. Dhabhar, F.S., NeuroImmunoModulation, 16(5): p300–317, Copyright 2009.)
6. Stress-induced changes in immune cell distribution – From barracks, to boulevards, to potential battlefields
Effective immuno-protection requires rapid recruitment of leukocytes into sites of wounding, infection, surgery, or vaccination. Immune cells circulate continuously on surveillance pathways that take them from the blood, through various organs, lymphatic vessels and nodes, and back into the blood. This circulation is essential for the maintenance of an effective immune defense network [64]. The numbers and proportions of leukocytes in the blood provide an important representation of the state of distribution of leukocytes in the body and of the state of activation of the immune system. The ability of short-term stress to induce changes in leukocyte distribution within different body compartments is perhaps one of the most under-appreciated effects of stress and stress hormones on the immune system [3; 4; 7; 9; 47].
Numerous studies have shown that short-term stress induces significant changes in absolute numbers and relative proportions of leukocytes in the blood. Stress-induced changes in blood leukocyte numbers have been reported in fish [65], hamsters [66], mice [67; 68], rats [7; 29; 69; 70], rabbits [71], horses [72], non-human primates [73], and humans [74; 75; 76; 77; 78; 79]. This suggests that the phenomenon of stress-induced leukocyte redistribution has a long evolutionary lineage, and has important functional significance. Interestingly, changes in blood leukocyte numbers were used as a measure of stress before methods were available to directly assay stress hormones [80]. Studies have also shown that glucocorticoid [69; 81; 82], and catecholamine [47; 76; 83; 84; 85; 86] hormones induce rapid and significant changes in leukocyte distribution and that these hormones are the major mediators of the effects of stress.
Short-term stress induces an initial increase followed by a decrease in blood lymphocyte and monocyte numbers, and an increase in blood neutrophil numbers [4; 79]. Soon after the beginning of stress (order of minutes) or during mild short-term stress or exercise, stress hormones induce the body’s “soldiers” (leukocytes), to exit their “barracks” (spleen, lung, marginated pool and other organs) and enter the “boulevards” (blood vessels and lymphatics). This results in an increase in blood leukocyte numbers, the effect being most prominent for NK cells and granulocytes. As the stress response continues, stress hormones, acting largely through normal immune cell surveillance and trafficking mechanisms, induce leukocytes to exit the blood and take position at potential “battle stations” (skin, mucosal lining of gastro-intestinal and urinary-genital tracts, lung, liver, and lymph nodes) in preparation for immune challenges which may be imposed by the actions of the stressor [4; 7; 8; 30]. Such a redistribution of leukocytes results in a decrease in blood leukocyte numbers. Thus, short-term stress induces a redistribution of leukocytes from the barracks, through the boulevards, and to potential battlefields [3; 4; 9; 68]. It was hypothesized that such a leukocyte redistribution may enhance immune function in compartments to which immune cells traffic during stress, and subsequently demonstrated that a stress-induced redistribution of leukocytes from the blood to the skin and subcutaneous tissues is accompanied by a significant enhancement of skin immunity [30; 87; 88].
Since the blood is the most accessible and commonly used compartment for human studies, it is important to carefully evaluate how changes in blood immune parameters might affect in vivo immune function in the context of the specific experiment or study at hand even when stress is not the focus of a study. Moreover, because most blood collection procedures involve a certain amount of stress, because all patients or subjects will have experienced short-term and chronic stress, and because many studies of psychophysiological effects on immune function focus on stress, the effect of stress on blood leukocyte distribution becomes a factor of considerable importance.
7. Short-term stress-induced enhancement of innate/primary immune responses
Effective immunoprotection requires rapid recruitment of leukocytes into sites of wounding, surgery, infection, or vaccination. Using an implanted surgical sponge as an in vivo wound healing arena, Viswanathan et al. investigated the effects of short-term stress on immune cell infiltration into a wound site [68] and elucidated the kinetics, magnitude, subpopulation-, and chemoattractant-specificity of an acute stress-induced increase in leukocyte trafficking to a site of immune activation. Compared to non-stressed controls, mice that were acutely stressed before sponge implantation showed 200–300% higher neutrophil, macrophage, NK and T cell infiltration. These authors further quantified the effects of acute stress on lymphotactin- (LTN, a predominantly lymphocyte-specific chemokine), and TNF-α-(a pro-inflammatory cytokine) induced leukocyte infiltration. An additional increase in infiltration induced by acute stress was observed for neutrophils, only in response to TNF-α; macrophages, in response to TNF-α and LTN; and NK and T cells only in response to LTN. These results showed that acute stress initially increases trafficking of all major leukocyte subpopulations to a site of immune activation. Tissue damage-, antigen-, or pathogen-driven chemoattractants subsequently determine which subpopulations are recruited more vigorously. Such stress-induced increases in leukocyte trafficking may enhance immunoprotection during surgery, vaccination, or infection, but may also exacerbate immunopathology during inflammatory (cardiovascular disease, gingivitis) or autoimmune (psoriasis, arthritis, multiple sclerosis) diseases.
Studies were also conducted to investigate whether the primary immune response in the skin is enhanced when antigen exposure takes place following a stressful experience. Short-term stress experienced at the time of novel antigen exposure resulted in a significant enhancement of the ensuing immune response [37]. Compared to controls, mice restrained for 2.5 hours before primary immunization with keyhole limpet hemocyanin (KLH) showed a significantly enhanced immune response when re-exposed to KLH nine months later. This immunoenhancement was mediated by an increase in numbers of memory and effector helper T cells in sentinel lymph nodes at the time of primary immunization. Further analyses showed that the early stress-induced increase in T cell memory may have stimulated the robust increase in infiltrating lymphocyte and macrophage numbers observed months later at a novel site of antigen re-exposure. Enhanced leukocyte infiltration was driven by increased levels of the Type-1 cytokines, IL-2 and IFN-γ, and TNF-α, observed at the site of antigen re-exposure in animals that had been stressed at the time of primary immunization. Given the importance of inducing long-lasting increases in immunological memory during vaccination, Dhabhar et al. have suggested that the neuroendocrine stress response is nature’s adjuvant that could be psychologically and/or pharmacologically manipulated to safely increase vaccine effectiveness [3; 4; 6; 11; 32; 37].
A similar enhancement of the sensitization/immunization/induction phase of cell-mediated immunity by different types of stressors administered at the time of antigen exposure, has been observed in mice, rats, and non-human primates [89; 90; 91]. A series of elegant experiments also showed that short-term stress experienced at the time of sensitization resulted in a significant increase in the contact hypersensitivity (CHS) response [92]. Other studies further elucidated the molecular and cellular mediators of the immunoenhancing effects of short-term stress [93]. They showed that compared to non-stressed mice, acutely stressed animals showed significantly greater pinna swelling, leukocyte infiltration, and upregulated macrophage chemoattractant protein-1 (MCP-1), macrophage inflammatory protein-3α (MIP-3α), IL-1α, IL-1β, IL-6, TNF, and IFN-γ gene expression at the site of primary antigen exposure. Stressed animals also showed enhanced maturation and trafficking of dendritic cells from skin to lymph nodes, higher numbers of activated macrophages in skin and lymph nodes, increased T cell activation in lymph nodes, and enhanced recruitment of surveillance T cells to skin [37; 93]. These findings showed that important interactive components of innate (dendritic cells and macrophages) and adaptive (surveillance T cells) immunity are mediators of the stress-induced enhancement of a primary immune response. Such immunoenhancement during primary immunization may induce a long-term increase immunologic memory resulting in subsequent augmentation of the immune response during secondary antigen exposure.
8. Short-term stress-induced enhancement of adaptive/secondary immune responses
In addition to enhancing primary cutaneous immune responses, short-term stress experienced at the time of antigen re-exposure can also enhance secondary or recall responses in skin [30]. Compared to controls, mice that were acutely stressed at the time of antigen re-exposure showed a significantly larger number of infiltrating leukocytes at the site of the immune reaction. These results demonstrated that a relatively mild behavioral manipulation can enhance an important class of immune responses that mediate harmful (allergic dermatitis) as well as beneficial (resistance to certain viruses, bacteria, and tumors) aspects of immune function. Other studies have similarly shown enhancement of the elicitation/recall phase of cell-mediated immunity by different stressors administered at the time of antigen re-exposure, in mice, rats, hamsters, and non-human primates [66; 89; 90; 91]. It has also been shown that short-term stress enhanced CMI responses in both male and female mice [94], however, these authors did not observe the stress-induced enhancement of the sensitization phase of CMI [95] that has been reported by several independent groups as described above [37; 89; 90; 91; 92; 93; 96].
Taken together, the findings described in Sections 6, 7, and 8 have led to the suggestion that the short-term stress response is mother nature’s endogenous adjuvant that kicks into gear during times of stress that are often accompanied by wounding and pathogen entry and require protective immune responses to be mounted in order to heal wounds and fight pathogens
9. Short-term stress induced enhancement of immune function in the context of cancer
Given the importance of cutaneous cell-mediated immunity in elimination of immuno-responsive tumors such as squamous cell carcinoma (SCC) [97; 98], and given the immuno-enhancing effects of short-term stress, studies have examined the effects of short-term stress administered at the time of ultraviolet light (UV) exposure (minimum erythemal dose, 3-times/week) on gene expression of chemokines and cytokines, infiltration of helper and cytolytic T cells that are critical for controlling and/or eliminating SCC and on tumor incidence, number and size [99]. Compared to controls, the short-term stress group showed greater cutaneous T-cell attracting chemokine (CTACK)/CCL27, RANTES, IL-12, and IFN-γ gene expression, higher infiltrating T cell numbers, lower tumor incidence, and fewer tumors early, but not later during tumor development. These results suggest that activation of short-term stress physiology increased chemokine expression and T cell trafficking and/or function during/following UV exposure, and enhanced Type 1 cytokine-driven cell-mediated immunity that is crucial for resistance to SCC [99]. A stress-induced reduction in tumor burden has similarly been reported for murine sarcoma virus induced tumors [100].
Although much work remains to be done, these findings show that short-term stress enhances anti-tumor immunity just as it enhances other aspects of innate and adaptive immunity. These findings raise the tantalizing possibility that the physiological fight-or-flight stress response, and its adjuvant-like immuno-enhancing effects may provide a novel and important mechanism for enhancing immune system mediated tumor-detection/elimination that merits further investigation. These findings also suggest that the beneficial effects of exercise/physical activity in the context of cancer [101; 102; 103; 104; 105; 106], may be at least partially mediated by activation of short-term stress physiology and it’s adjuvant-like effects.
10. Endocrine mediators of stress-induced enhancement of immune function
Although much work remains to be done to identify molecular, cellular, and physiological mechanisms mediating the adjuvant-like, immuno-enhancing effects of short-term stress, studies have shown that corticosterone and epinephrine are important mediators of a short-term stress induced immuno-enhancement [87]. Adrenalectomy, which eliminates the glucocorticoid and epinephrine stress response, eliminated the stress-induced enhancement of cell-mediated immunity. An inverted-U/bell-shaped relationship was observed between the concentration of stress hormone and it’s effects on immune function: Low dose corticosterone or epinephrine administration significantly enhanced the immune response [87]. In contrast, high dose corticosterone, chronic corticosterone, or low dose dexamethasone were potently anti-inflammatory effects [87] as would be expected from their well-known use in the clinic [48]. These results suggested a novel role for physiological concentrations of adrenal stress hormones as endogenous immuno-enhancing agents. They also showed that hormones released during a short-term stress response may help prepare the immune system for potential challenges (e.g., wounding or infection) for which stress perception by the brain may serve as an early warning signal. Other studies have also suggested that physiological concentrations of glucocorticoid hormones mediate stress-induced enhancement of interferon production [107], skin contact hypersensitivity [94], and that the adjuvant like effects of stress on dendritic cell and CD8+ T cell migration and function, that mediate immuno-enhancement are driven by norepinephrine [92]. In a series of elegant studies, Sanders and colleagues have elucidated the role of the beta-adrenergic receptor in regulating lymphocyte function, and have shown that the level of activation is influenced by the time of receptor engagement relative to the state of activation and/or differentiation of the lymphocyte and by the cytokine milieu [108; 109]. Taken together, these studies suggest that endogenous stress hormones in physiological concentrations can have immuno-enhancing effects, while endogenous hormones at pharmacologic concentrations, and synthetic hormones, are immuno-suppressive.
11. Cytokine mediators of stress-induced enhancement of immune function
Since gamma interferon (IFNγ) is a critical cytokine mediator of cell mediated-immunity as well as delayed, and contact hypersensitivity, studies were conducted to elucidate the role of IFNγ as a local mediator of the stress-induced enhancement of skin immunity [88]. The effect of short-term stress on skin immunity was examined in wild-type and IFNγ receptor gene knockout mice (IFNγR−/−). Acutely stressed wild-type mice showed a significantly larger cell mediated immune response than non-stressed mice. In contrast, IFNγR−/− mice failed to show a stress-induced enhancement of skin immunity. Immuno-neutralization of IFNγ in wild-type mice significantly reduced the stress-induced enhancement of skin immunity [88]. In addition to IFNγ, stress-induced increases in gene expression of TNF, MCP-1, MIP-3α, IL-1α, IL-1β, and IL-6 (but not IL-4) have also been associated with enhancement of the immunization phase of cell-mediated immunity [37; 93].
Another important immunological effect of short-term stress is to induce a significant increase in concentrations of circulating cytokines such as IL-6 and IL-1β [17; 18; 19; 110; 111; 112]. Importantly, this increase is observed in response to psychological stressors such as the Trier Social Stress Test (TSST) and in the absence of immune activating events such as a wound, or antigen/pathogen inoculation. We suggest that such short-term stress-induced increases in circulating cytokines may be an additional systemic mechanism mediating stress-induced enhancement of immune function. Interestingly, short-term stress-induced increases in circulating cytokines are related to changes in emotional states experienced during stress. For example, IL-1β reactivity during stress is a significant mediator of the relationship between a decline in positive affect and cognitions during stress, and an increase in depressive symptoms one year later [19]. Such mediation is particularly salient given the known role of proinflammatory cytokines in inducing sickness behavior, depressive states, and depression [55; 113; 114; 115; 116] and in important reciprocal immune-to-neural signaling [55; 117; 118; 119].
In another interesting example, anger experienced during a stressor is related to a stress-induced increase in circulating IL-6, however, perceived social support mitigates the effects of anger on IL-6 stress reactivity such that the greater the amount of social support, the lower the stress reactivity of IL-6 [18]. In light of these findings, it has been suggested that short-term stress-induced increases in IL-6 and other pro-inflammatory cytokines may confer a survival advantage by facilitating immuno-enhancement during/following short-term stress [18]. We have speculated that individuals with low social support may be more likely to be “out on their own,” and have to fend for themselves, and as a result be more susceptible to attack and/or injury [18]. Therefore, such individuals may mount a more robust immunological stress response. Furthermore, an angry individual may be more likely to engage in an aggressive encounter, i.e., choose to fight rather than flee, and as a result may be more likely to need enhanced immune defenses to heal wounds (incurred during the fight) and to defend against accompanying pathogen entry. Such evolutionary underpinnings may partially explain the association among emotional states and stress-reactivity of proinflammatory cytokines. As with most psychological and biological processes, activating this response too frequently or for too long (especially in the absence of a wound or infection), may result in greater long-term exposure to proinflammatory factors resulting in their deleterious health consequences. Such chronic effects may underlie the proinflammatory milieu that is often observed during various disorders [55; 120] like major depression [111; 121; 122; 123; 124], alcohol addiction [125] and posttraumatic stress disorder [126; 127; 128], and in some cases may be facilitated by the genetic makeup of an individual [129].
12. Immunomodulatory effects of timing of stress or stress hormone administration relative to the timing of immune activation and the time course of the ensuing immune response
Under certain conditions, endogenous stress hormones have immunoenhancing effects while under other conditions these hormones suppress autoimmune and inflammatory reactions. It is possible that these differential effects are achieved by differences in overall glucocorticoid sensitivity of the affected immune response. At the beginning of an immune response, certain components such as leukocyte trafficking, antigen presentation, helper T cell function, leukocyte proliferation, cytokine and chemokine function, and effector cell function may be receptive to glucocorticoid-mediated immunoenhancement. In contrast, at a later, more advanced stage of an immune response these components may be more receptive to glucocorticoid-mediated immunosuppression. While this hypothesis needs to be tested through further experiments, studies examining the effects of corticosterone on T lymphocyte proliferation in vitro [130], support the hypothesis that there may be temporal differences in the receptivity of an immune response to the enhancing versus suppressive effects of endogenous glucocorticoid hormones. Thus, studies have shown that during the early stages of T cell activation, low levels of corticosterone potently enhance anti-TCR-induced lymphocyte proliferation. However, during later stages of culture, the same levels of corticosterone suppress T lymphocyte proliferation [130]. Furthermore, it has been shown that corticosterone had to be present during the process of TCR activation in order to enhance the proliferative response. If corticosterone was added to the culture system more than two hours after the initiation of TCR activation, the enhancement of lymphocyte proliferation was not observed. Sanders and colleagues have elegantly elucidated the role of the beta-adrenergic receptor in regulating lymphocyte function, showing that the level of activation is influenced by the time of receptor activation relative to the state of activation and/or differentiation of the lymphocyte and by the cytokine milieu [108; 109]. Similar bimodal effects of catecholamines dependent on the state (early versus late) of progression of rheumatic disease have also been shown [131]. It has been proposed that energy and volume regulation may be one important aspect of interactions between stress (and other) hormones and the immune system and that these factors may take on additional significance during chronic inflammatory conditions [132; 133].
13. The transition from adaptive to maladaptive effects of stress: When stress-related changes become chronic
We have defined chronic stress as that which results in stress-related biological changes that last for weeks, months, or years [9]. Figure 2 illustrates the steps and factors that mediate the transition from adaptive short-term stress to deleterious chronic stress. Time is represented on the “x” axis and changes in stress-related biological factors are represented on the “y” axis. The upright isosceles triangle (top panel) represents an adaptive short-term response. In order for a stress response to be adaptive it is critical that stress related biological changes occur rapidly (within minutes) and resolve back to baseline rapidly (within minutes to hours). Life involves a series of stress “hits” and responses for most organisms. Most humans and other organisms are built to handle this series of stress responses and can continue to mount adaptive/protective responses under such conditions especially if their psychological and physiological systems return to baseline/resting conditions between stress hits. The isosceles trapezoid (middle panel) represents a prolonged short-term stress response with delayed shutdown which results in greater overall area under the curve exposure to stress related factors compared to the prototypical adaptive stress response. Such prolonged stress responses can begin to have deleterious effects especially if they are part of a series of stress hits with little or no time of return to baseline/resting conditions between hits. The long rectangle (bottom panel) is a graphical representation of chronic stress that is known to have numerous deleterious effects on brain, body, and health, and mental and physical performance.
Figure 2. Steps and factors that mediate the transition from adaptive short-term stress to deleterious chronic stress.
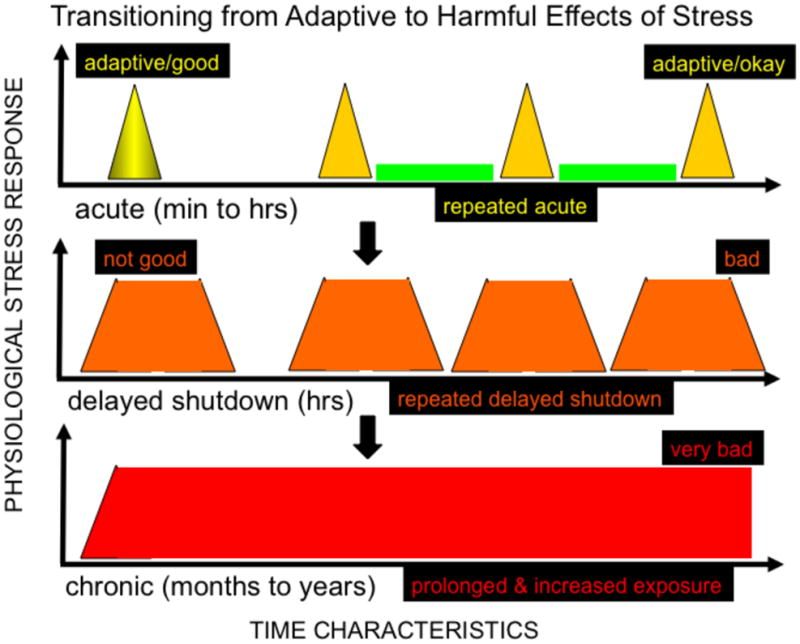
Time is represented on the “x” axis and changes in stress-related biological factors are represented on the “y” axis. The upright yellow isosceles triangle (top panel) represents an adaptive short-term stress response. In order for a stress response to be adaptive, it is critical that stress related biological changes occur rapidly (within minutes) and resolve back to baseline rapidly (within minutes to hours). Life involves a series of stress “hits” and responses. Most humans and other organisms are built to handle this series of stress responses and can continue to mount adaptive/protective responses under such conditions especially if their psychological and physiological systems return to baseline/resting conditions (green bars between yellow triangles) between stress hits. The isosceles trapezoid (middle panel) represents a prolonged short-term stress response with delayed shutdown which results in greater overall area under the curve exposure to stress related factors compared to the prototypical adaptive stress response. Such prolonged stress responses can begin to have deleterious effects especially if they are part of a series of stress hits with little or no time of return to baseline/resting conditions between hits. The long red rectangle (bottom panel) is a graphical representation of chronic stress that is known to have numerous deleterious effects on brain, body, and health, and mental and physical performance. An important characteristic of chronic stress is that the stressor(s), stress perception, or aspects of the stress response, persist for long periods (months to years) of time and result in an overall increase in exposure to, and/or sustained changes in, stress-related biological factors. Rather than stress in general, it is chronic stress that suppresses endogenous defenses (such as wound healing and immuno-protective responses), and repair/restoration mechanisms (such as DNA repair, telomere lengthening by telomerase, and anti-oxidant systems), and mental and physical performance.
An important characteristic of chronic stress is that the stressor(s), stress perception, or aspects of the stress response, persist for long periods (months to years) of time and result in an overall increase in exposure to, and/or sustained changes in, stress-related biological factors. The concept of “allostatic load” has been proposed to describe the “psycho-physiological wear and tear” that takes place while different biological systems work to stay within a range of equilibrium (allostasis) in response to demands placed by internal or external chronic stressors (for review see: [1; 13; 134; 135]). A disruption of the circadian cortisol rhythm is a critical indicator and/or mediator of the beginning of the deleterious effects of chronic stress [9]. Other harbingers of the deleterious effects of chronic stress include the disruption or dysregulation of stress-related and other physiological processes such as sleep, metabolism, and behavior. We suggest that rather than stress in general, it is chronic stress that suppresses endogenous defenses (such as wound healing and immuno-protective responses), and repair/restoration mechanisms (such as DNA repair, telomere lengthening by telomerase, and anti-oxidant systems), and mental and physical performance.
14. Chronic stress-induced suppression/dysregulation of immune function
In contrast to short-term stressors, chronic stress has been shown to suppress or dysregulate immune function. This topic has been the subject of many excellent reviews (such as: [5; 35; 131; 136; 137; 138; 139; 140; 141]). In addition to significant personal and health-related costs of chronic stress, the economic cost to industry arising from work-related stress in the United States alone is thought to be more than $ 300 billion [142].
14.1 Effects of increasing the intensity, duration, and chronicity of stress
Short-term stress administered for 2h prior to antigenic challenge, significantly enhanced skin cell-mediated immunity [9]. Increasing the duration of stress from 2h to 5h produced the same magnitude immuno-enhancement. Interestingly, increasing the intensity of short-term stress produced a significantly larger enhancement of the immune response that was accompanied by increasing magnitudes of leukocyte redeployment. In contrast, immuno-suppression was observed when chronic stress exposure was begun 3 weeks before primary immunization and either discontinued following immunization, or continued an additional week until re-exposure to the antigen, or extended for one week after re-exposure [9]. Interestingly, short-term stress induced redistribution of peripheral blood lymphocytes was attenuated with increasing duration of stressor exposure and correlated with attenuated glucocorticoid responsivity. These results suggested that stress-induced alterations in lymphocyte redeployment may play an important role in mediating the bi-directional effects of stress on cutaneous cell-mediated immunity [9]. An association between chronic stress and reduced skin cell mediated immunity has also been reported in human subjects [143; 144].
14.2 Effects of chronic stress on leukocyte numbers, mobilization, and trafficking
A chronic stress-induced decrease in baseline leukocyte numbers and leukocyte mobilization and trafficking from the blood to other body compartments is an important mediator of stress induced suppression of immune function [9]. In human and animal studies, chronic stress has also been shown to suppress different immune parameters examples of which include: CMI [145], antibody production [146; 147], NK activity [34; 148; 149; 150], leukocyte proliferation [148; 150; 151], skin homograft rejection [152], virus-specific T cell and NK cell activity [153], and anti-mycobacterial activity of macrophages from susceptible mouse strains [154].
14.3 Chronic stress induced acceleration of immunosenescence
Accelerated biological aging is another important mechanism through which chronic stress suppresses/dysregulates immune function. In a seminal study, Epel et al. showed that blood lymphocytes and monocytes from women reporting high chronic stress levels have significantly shorter telomeres compared to leukocytes from women reporting low stress [155]. Immune cell telomerase activity was also lower in the high stress women indicating a chronic stress induced decrease in their ability to rebuild shortened telomeres [155]. The study concluded that “women with the highest levels of perceived stress had telomeres that were shorter on average by the equivalent of at least one decade of additional aging compared to low stress women [155].” Epel et al have also shown that the rate of telomere shortening predicts death from cardiovascular disease [156], and has significant deleterious effects [157]. Thus, chronic stress induced telomere attrition can have significant deleterious effects on immune function because it could lead to DNA replication errors and is also likely to result in suppression of immuno-protection and exacerbation of immune dysregulation and immuno-pathology.
15. Chronic stress and suppression of protective immune responses during wound healing, vaccination, and infection
Chronic stress-induced suppression of protective immunity has been the subject of many excellent reviews [5; 35; 131; 136; 137; 138; 139; 140; 141; 158]. Briefly, chronic stress has been shown to suppress wound healing [159; 160; 161; 162; 163] and vaccine [35; 158; 164; 165; 166] related immune responses. Chronic stress has also been shown to increase susceptibility to viral [153; 167; 168; 169; 170; 171] and bacterial infection [172; 173].
16. Chronic stress and cancer
Numerous studies have investigated the effects of chronic stress in the context of cancer [174; 175; 176]. In light of the immuno-suppressive effects of long-term stress, and given the importance of cell-mediated immunity in elimination of immuno-responsive tumors like squamous cell carcinoma [97], studies have also investigated the effects of chronic stress on cancer emergence [39] and progression [39; 177; 178; 179; 180]. Chronic stress significantly accelerated the emergence and progression of squamous cell carcinoma (SCC). Compared to non-stressed controls, chronically stressed mice had lower IFN-γ, CCL27/CTACK, and CD3ε gene expression and lower CD4+ and CD8+ T cells infiltrating within and around tumors. Chronically stressed mice also showed a shorter median time to first tumor and reached 50% incidence six weeks earlier than controls. Interestingly, stressed mice had higher numbers of tumor infiltrating and circulating regulatory/suppressor T cells than non-stressed mice. These studies showed that chronic stress increased susceptibility to UV-induced squamous cell carcinoma by suppressing skin immunity, Type 1 cytokines, and protective T cells, and increasing active immuno-suppressive mechanisms mediated by regulatory/suppressor T cells [39]. Similarly, studies have shown that a high-anxious behavioral phenotype, that is likely to be associated with increased susceptibility to chronic stress, is associated with suppressed anti-tumor immunity, and increased susceptibility to the emergence and progression of squamous cell carcinoma [181].
17. Chronic stress and autoimmune disease
Given the immuno-suppressive effects of chronic stress, it may be hypothesized that under certain conditions, chronic stress could ameliorate autoimmune diseases. A few preclinical studies suggest that this may be the case. Levine et al. demonstrated that the administration of prolonged restraint stress to rats before the induction of experimental allergic encephalomyelitis (EAE) resulted in a suppression of the incidence and severity of disease [182]. Rogers et al showed that exposure of rats to a variety of stressors results in a marked suppression of the clinical and histological manifestations of type II collagen-induced arthritis [183]. Similarly, Griffin et al. demonstrated suppression of EAE by chronic stress [184]. In an elegant series of experiments, Stefanski et al. recently showed that severe (but not moderate) social stress significantly reduced susceptibility to collagen-induced arthritis in Wistar rats, and that this effect was mediated by decreases in CD4, CD8 T cell numbers and macrophage infiltration at the site of collagen injection [185].
One would not recommend chronically stressing anyone, leave alone patients with autoimmune disease. However, there may be lessons to be learned from the above-mentioned studies. Important questions for future studies include: 1) What are the physiological conditions and mechanisms under which chronic stress can exert immuno-suppressive effects in the absence of inducing proinflammatory effects? 2) Does a chronic stress-induced increase in regulatory/suppressor T (Tregs) [39], regulatory B cells [181], NK cells, dendritic cells or monocytes/macrophages mediate suppression of autoimmune responses? 3) Is chronic stress induced amelioration of autoimmune disease observed in human subjects? 4) If so, could some of the biological mechanisms mediating chronic stress-induced amelioration of autoimmune reactions be safely and selectively harnessed to treat autoimmune diseases without administering chronic stress? Clearly, more research is warranted into investigating whether chronic stress ameliorates autoimmune reactions in humans, delineating the conditions under which such amelioration is observed, and elucidating mechanisms with the goal of identifying targets for pharmacological or biobehavioral interventions.
Importantly, it has also been suggested that chronic stress-induced exacerbation of inflammatory diseases such as rheumatoid arthritis may be mediated by a loss of immuno-suppression that is normally driven by sympathetic nerves that innervate the inflamed tissue, and by systemic secretion of cortisol through cytokine-induced activation [186; 187] of the hypothalamic-pituitary-adrenal axis [188; 189]. Sternberg et. al., initially showed that a defect in inflammation induced activation of the HPA-axis, resulting in a reduction/loss of the anti-inflammatory effects of endogenous glucocorticoids, is an important factor in the progression of autoimmune diseases [42; 190].
18. Exercise, stress, and immune function: Beneficial versus deleterious effects
The process of exercising induces a physiological stress response and increases circulating concentrations of adrenaline (epinephrine), noradrenaline (norepinephrine), cortisol, and other stress-related factors including cytokines [191; 192; 193]. Understanding the psychological, physiological, and health effects of exercise in the context of stress and stress physiology is critical for several important reasons: 1) A hitherto unappreciated but critical mechanism mediating the salubrious effects of exercise could be through its optimization of the beneficial, survival-promoting effects of the short-term stress response [3; 11; 194]. Regular exercise may help keep the short-term stress response oiled, fine-tuned, and ready for fight or flight. This idea also makes sense from an evolutionary perspective because regular and robust physical activity is an intrinsic part of life in nature. The “conveniences” of modern societies might cause unintended harm by decreasing our levels of physical activity and making exercise optional in our day-to-day lives. Studies have shown that physical activity can modulate cancer-related pathways and improve some biomarkers associated with better prognosis [195]. In keeping with this idea, recently conducted mouse studies have shown that exposure to short-term stress (three times per week) in a manner that mimics exercise-induced activation of short-term stress physiology, significantly enhanced anti-tumor immunity and decreased tumor burden [99]. These findings suggest that regular activation of the short-term stress response, in a frequency that does not induce chronic stress, may be one mechanism mediating findings from human studies showing that moderate and regular physical activity reduces the risk of cancer occurrence [196; 197], progression, and mortality [198]. 2) Intense prolonged exercise [199] or exercising under extreme environmental conditions [200], may lead to chronic exposure to stress hormones that make the individual susceptible to the deleterious health effects of chronic stress. Exercise-induced pain, exhaustion, or injury could also induce psychological stress. 3) When performed regularly and in moderation, exercise could be a factor in ameliorating the deleterious health effects of chronic stress and increased allostatic load (viz. the physiological cost that results from ongoing adaptive efforts to maintain homeostasis in response to stressors) [1; 201; 202; 203]. The type, intensity, duration and frequency of exercise and the conditions under which it should be performed in order to effectively reduce the stress burden of different individuals need to be better understood and defined. It is likely that one would need different strokes for different folks, i.e., running could serve as a “stress optimizer” for some while others would benefit from aerobics, swimming, dancing or yoga. The most beneficial results are likely to arise when the physical as well as psychosocial aspects of the exercise are matched with factors such as the fitness, capability, temperament, personality, etc., of the exercising individual. 4) The psychosocial stress status of an individual may positively or negatively affect the relationship between exercise and health. For example, compared to a low-stress individual, a chronically stressed individual may react differently to the effects of exercise [204]. Furthermore, the stress status of an individual could affect their motivation or ability to exercise.
These are areas of research that are ripe for investigation and are relevant for the well-being of recreational and elite athletes as well as armed forces and other professions for whom exercise is a critical aspect of training and job-performance. Physical activity and exercise are potent stimulators of the physiological stress response. Therefore, many health effects of exercise are likely to be mediated through stress and immune factors in addition to cardiovascular, neuromuscular, and other factors.
19. Short-term stress induced immunoenhancement: From bench to bedside
Dhabhar et al. have proposed that a psycho-physiological stress response is one of nature’s fundamental survival mechanisms that could be therapeutically harnessed to augment immune function during vaccination, wound healing or infection [3; 4; 33; 37; 205]. These adjuvant-like immuno-enhancing effects of short-term stress likely evolved because many stressful situations (aggression, accident) result in immune activation (wounding, infection) and vice versa. Interestingly, in modern times, many medical procedures involving immune activation (vaccination, surgery) also induce a stress response. Preclinical findings initially lent support to this hypothesis and have since been replicated in studies involving human subjects.
Based on a series of preclinical findings, we hypothesized that patients who show an a priori defined adaptive immune cell redistribution (“barracks to boulevards to battlefields,” as described in Section 6) during the short-term stress of surgery, will also show significantly enhanced postsurgical recovery [79]. To test this hypothesis, we enumerated blood immune cell redistribution profiles induced by short-term surgery stress in patients undergoing knee surgery. An adaptive immune cell redistribution was defined a priori as a blood count profile that showed an increase (relative to baseline) in absolute numbers of circulating lymphocytes or monocytes at the beginning of stress/surgery, and a decrease (relative to baseline or peak numbers) in lymphocyte or monocyte numbers towards the end of stress/surgery. Results showed that patients who showed the predefined “adaptive” lymphocyte redistribution profiles during surgery showed significantly enhanced recovery as measured by the Lysholm Scale at 1, 3, 8, 16, 24, and 48 weeks after surgery [79]. In contrast, knee function of patients who showed a maladaptive immune cell redistribution response, plateaued at eight weeks following surgery, and failed to reach the maximum recovery levels shown by patients who showed adaptive immune cell redistribution during the short-term stress of surgery [79]. Thus, the stress induced immune cell redistribution profile measured on the day of surgery, predicted the trajectory of recovery for almost one year after surgery [79]. These results may be applicable to a wide range of surgical procedures because the physiological stress response, that is known to drive stress-induced changes in immune cell distribution [4; 7; 30; 69; 87; 206], is likely to be similar during different types of surgery.
While these findings need to be replicated, they lay the foundation for important clinical applications (summarized from Rosenberger et al.) [79]: 1) Quantifying blood immune cell numbers is quick, straightforward and inexpensive. Therefore, changes in blood leukocyte redistribution could be monitored during surgery to inform the surgeon about whether additional para- or post-surgical intervention is indicated for enhancing recovery, especially for patients who show maladaptive immune cell redistribution. 2) The specific effects of the principal stress hormones, epinephrine, norepinephrine, and cortisol on immune cell redistribution patterns and subpopulation specificity are largely known [4]. This suggests that these hormones could be administered at specific times during surgery to induce “adaptive” leukocyte redistribution in patients showing a maldaptive response. 3) Further elucidation of mechanisms mediating adaptive versus maladaptive immune cell redistribution during stress is important because it would support the formulation of pharmacological, bio-behavioral, and psychological interventions to maximize adaptive surgery stress induced leukocyte redistribution and the related enhancement of recovery. 4) It has been hypothesized that the ability to mount adaptive immune cell redistribution responses during stress may be a trait-like characteristic that is expressed in a similar manner across different stressors [79]. If this hypothesis is confirmed, it raises the possibility of prospectively identifying, patients who are likely to show adaptive versus maladaptive stress-induced changes in immune cell distribution by administering a “stress test” prior to surgery. Prospective identification coupled with stress-optimization interventions could maximize the probability of a patient showing adaptive leukocyte redistribution during surgery, leading to enhanced recovery following surgery. The significant personal and economic benefits of enhanced recovery include reduced morbidity, fewer days lost from work or sports activities, more complete and long-lasting return to life activities, reduced risk of re-injury, and reduced individual and societal healthcare costs.
Similarly, preclinical studies initially demonstrated that short-term stress experienced during primary [37; 93] or secondary [30; 87; 88; 92] antigen exposure significantly enhances the ensuing immune response. Based on these laboratory studies, an elegant series of clinical studies has shown that adjuvant effects of short-term psychological stress, or exercise stress, administered before vaccination, can enhance vaccine-induced immunity in human subjects [207; 208; 209]. In terms of further mechanistic parallels between basic and human subjects studies, it has been shown that a short-term stress-induced enhancement of skin immunity in mice is mediated by enhanced maturation and trafficking of dendritic cells from skin to draining lymph nodes, larger numbers of activated macrophages in skin and lymph nodes, and increased T cell activation in lymph nodes [93]. These findings are in agreement with studies that showed that short-term psychological stress in human participants induces a significant decrease in epidermal Langerhans’ cells that the authors suggest represents a trafficking of these cells from the skin to draining lymph nodes [210], a phenomenon that has been described as having striking similarities in “mice and men” [211].
Stress induced enhancement of pro-inflammatory and autoimmune disorders
While short-term stress-induced enhancement of immuno-protective responses have been appreciated relatively recently, stress-induced exacerbations of pro-inflammatory (e.g. dermatitis [212; 213], cardiovascular disease [214; 215], periodontal disease [216], and asthma [212; 217; 218]) and autoimmune (e.g., psoriasis [219; 220], arthritis [221], multiple sclerosis [222]) diseases are well-known and frequently observed in the clinic. We have suggested that stress-induced exacerbation of proinflammatory and autoimmune diseases may be partially mediated by mechanisms that are similar to those that enhance protective immune responses during stress [3; 4; 205]. Therefore, it would be beneficial for future studies to: 1) Determine the extent to which stress-induced exacerbation of such disorders is mediated by immuno-enhancing mechanisms activated during short-term stress versus immuno-dysregulatory mechanisms activated during chronic stress. 2)Determine the extent to which stress induces the onset of disease, and the extent to which stress exacerbates ongoing disease. 3) Use more standardized psychological and physiological measures of stress and where possible also of the immune parameters affected by stress.
20. Short-term stress induced immunoenhancement: Sex Differences
The overall immune-enhancing effects of short-term stress that are described here are similar in women and men [79; 223]. However, a few findings suggest that further investigation of sex differences in stress-induced enhancement of immune function would be beneficial and could enable the development of interventions designed to maximally harness the protective effects of short-term stress in women and men. For example, sex differences were observed in the knee surgery study by Rosenberger et al. that is described in Section 19: Women and men who showed the adaptive profile of lymphocyte redistribution during surgery, showed improved recovery compared to individuals who showed a maladaptive profile of lymphocyte redistribution [79]. Interestingly, women who were high lymphocyte redistributors showed enhanced early recovery, while men who were high lymphocyte redistributors showed higher maximum knee function [79]. In the case of stress induced enhancement of vaccination induced immune responses, it has been shown that eccentric exercise before the administration of influenza vaccine induced differential effects in women and men, with the antibody response being enhanced in women, and cell-mediated immunity being enhanced in men [223]. Findings such as these indicate that further investigation of sex differences in stress-induced enhancement of immune function is warranted.
21. Short-term stress induced enhancement of mental/cognitive and physical performance
As is the case with the effects of stress on immune function, chronic stress can suppress mental and physical performance. However, it is likely that a short-term stress response can enhance performance under some conditions. Indeed, studies have shown that aroused, excited, or anxious emotional states, all of which can induce a short-term stress response, can also have beneficial effects for mental performance. For example, it has been shown that amongst students preparing to take a practice Graduate Record Examination (GRE) those who were told that arousal improves performance, showed a significantly larger increase in salivary alpha amylase (indicating a more robust activation of the sympathetic nervous system) and performed significantly better on the GRE-math section, than students who were not given this information. Interestingly, these same students also performed better on the match section of the actual GRE [224]. Studies have also shown that higher anxiety increases the input of the motor pathway when an individual is processing threatening or negative social stimuli. The authors interpret their findings as showing that: “threat tunes neural processing in [a] fast, selective, yet attention-independent fashion in sensory and motor systems for different adaptive purposes” [225]. Another study showed that when subjects reappraised anxiety as excitement by stating, “I am excited,” before making a speech, they felt more excited, spoke longer, and were rated as being more competent and persuasive than subjects who reappraised their anxiety as calmness by stating, “I am calm,” before making their speech [226]. Similar effects were observed in a math performance task [226].
Taken together, these studies show that reappraisal of a stressor as a challenge/opportunity instead of a threat can lead to better performance. Here we hypothesize that in the context of the effects of short-term stress on mental performance, the reappraisal process may serve to optimize the short-term stress response, and/or prepare the individual’s brain circuitry and overall psychology to better receive stress signals to perform better during stress, and/or amerliorate/eliminate specific effects of stress physiology that inhibit certain aspects of cognitive function. All elements of these hypotheses need to be tested rigorously. Importantly, one study has indeed shown that compared to controls, subjects who were induced to reappraise their arousal by being told that arousal is functional and adaptive, subsequently showed a more optimized/adaptive stress response (“increased cardiac efficiency, lower vascular resistance, and decreased attentional bias”) during the Trier Social Stress Test [227].
Numerous studies have also shown memory enhancement following exposure to stress or stress hormones. McIntyre and Roozendall have shown that epinephrine and cortisol released during stress or administered during exposure to emotionally arousing experiences, enhance memory for such experiences [228]. Marin et al. showed that short-term stress enhances emotional memories that are reactivated just before exposure to stress [229]. Other studies have shown that the magnitude of the systemic norepinephrine response to emotionally arousing stimuli predicts and perhaps mediates, the strength of long-term memory for those stimuli [230]. Segal et al. showed that norepinephrine released during exercise undertaken following a learning task enhances memory in patients with mild cognitive impairment and normal controls [231]. Interestingly, it has also been shown that glucocorticoid hormones administered at the time of memory acquisition induced memory enhancement but only in the presence of increased noradrenergic activation [232; 233].
It has also been shown that a high-intensity short-term stress response can inhibit rather than enhances mental performance [234]. We suggest that the memory disrupting effects of high-intensity stressors may be mediated by an inverted-U shaped relationship between stress hormones and mental performance, similar to that observed with stress hormone induced enhancement of immune function [87]. Indeed, studies conducted by Conrad et al. and Lupien et al. do show an inverted U-shaped relationship between stress hormone concentrations and memory [235; 236]. Short-term stress could also appear to inhibit performance if the test being used to measure mental performance is not ethologically relevant for a fight-or-flight situation (e.g. a mouse may not care to look for food when it is being chased by a fox). We suggest that in human subjects, training and practice may be important factors that teach an individual to harness their fight-or-flight response to increase performance even for tasks that are not ethologically relevant in a fight-or-flight context (e.g. harnessing your anxiety during a math exam, job interview, or while making a speech even though there is no imminent threat to survival during any of these situations).
Similar to its effects on mental performance, we propose that short-term stress acting through stress hormones also enhances physical performance. Although evidence is not as abundant as it is for enhancement of immune function, studies have begun to elucidate mechanisms. It has been shown that endogenous cortisol concentrations are positively related with muscle strength especially in the context of lower testosterone concentrations [237]. Studies using pharmacological blockade of catecholamine receptors have indicated a role for catecholamine hormones in increasing exercise performance (mean peak VO2 and cumulative work/time to exhaustion) [238]. Kim et al. have shown that acute administration of capsaicin increases endurance swimming capacity in mice, and that this increase is mediated by adrenal stress hormone induced enhancement of fatty acid utilization [239].
It is likely that stress-induced enhancement of mental and physical performance is harnessed by athletes, dancers, performers, entertainers, first responders, emergency room and intensive care personnel, surgeons, armed forces personnel, special operations forces, and many individuals in other walks of life who are highly successful. Such individuals may deal successfully with stress with some even appearing to thrive on it. Interestingly, it has also been reported that short-term enhances prosocial behavior [240]. We suggest that this could in turn contribute to enhancement of mental and physical performance. Importantly, a large portion of the beneficial effects of practice and training could be through optimizing the short-term stress response and it’s adaptive effects, and through preparing and enabling the individual to harness these effects during times of stress. This hypothesis needs to be further tested. Taken together, the findings discussed in section 21 suggest that as is the case for short-term stress induced enhancement of immune function [4; 7; 69; 87], multiple elements of the physiological fight-or-flight response mediate stress-induced enhancement of mental and physical performance and that the relationship between stress hormones and mental and physical performance may also be represented by an inverted U or bell-shaped curve.
22. Effects of stress on protection and performance: The good, the bad, and the beautiful
The GOOD: The short-term stress response experienced at the time of immune activation may naturally increase immunoprotection during surgery, vaccination, infection, or cancer. Similarly, short-term stress could enhance mental and physical performance. The BAD: Under some conditions, immune-enhancement driven by short-term stress or immune-dysregulation driven by long-term stress, can exacerbate pro-inflammatory (dermatitis, cardiovascular disease, gingivitis) and autoimmune diseases (psoriasis, arthritis, multiple sclerosis) diseases that are known to be exacerbated by stress [219; 241; 242; 243]. Moreover, chronic stress can delay wound healing [35], suppress vaccine responses [35], and increase susceptibility to infections [244] and cancer [39; 177; 178; 181]. Chronic stress can also impair mental performance [245]. The BEAUTIFUL: Preclinical and clinical studies showing short-term stress induced enhancement of immune function during surgery [68; 79], vaccination [30; 37; 87; 88; 92; 93; 209] and cancer [99], raise the tantalizing possibility that the physiological fight-or-flight stress response and its adjuvant-like immuno-enhancing effects may provide a novel and important mechanism for enhancing immune protection. Similarly, short-term stress physiology could be harnessed to enhance mental and physical performance. Further studies could lead to the development of treatments that induce a short-term stress response (e.g., consistent moderate exercise, virtual-reality stressors, or pharmacological agents) to boost protective immunity and performance.
23. Staying On The Good Side Of The Stress Spectrum
Dhabhar et al. have proposed the concept of the STRESS SPECTRUM to understand, optimize, and harness the beneficial effects of short-term stress, and minimize the harmful effects of chronic stress [3; 6; 8; 9; 11] (Figure 3). One end of the Stress Spectrum is characterized by GOOD STRESS, i.e., short-duration stress that results in physiological conditions that enhance immuno-protection and mental and physical performance. An important characteristic of good stress is a rapid physiological stress response mounted in the presence of the stressor, followed by a rapid shut-down of the response upon cessation of the stressor. While the sympathetic nervous system and hypothalamic-pituitary-adrenal axis are the major drivers of the fight-or-flight response, the parasympathetic nervous system, together with negative feedback loops within the fight-or-flight response, are the major drivers of recovery from stress. The parasympathetic nervous system drives the rest-and-digest response and mediates recovery and a return to resting state following stress. The opposite end of the spectrum is characterized by BAD STRESS or DISTRESS, i.e., chronic or long-term stress that can result in dysregulation or suppression of immune function, and can inhibit mental and physical performance. An important characteristic of chronic stress is that the physiological response either persists long after the stressor has ceased, or is activated repeatedly to result in an overall increase in exposure to stress hormones and/or dysregulation of stress-related and other physiological processes such as sleep, metabolism, and behavior. A disruption of the circadian cortisol rhythm is a critical indicator and/or mediator of the beginning phase of the deleterious effects of chronic stress [9; 11]. The Stress Spectrum also describes the RESTING ZONE of low/no stress that represents a state of health maintenance/restoration (Figure 3). The extent, speed, and efficiency with which an organism returns to its resting state after stress depends partly on RESILIENCE, which we define as the capacity of psycho-physiological systems to recover from challenging conditions. Psychological and physiological resilience factors determine the overall effects of stress on an individual [3; 32].
Figure 3. The Stress Spectrum[3; 6; 11; 32; 205].
We have proposed the concept of the STRESS SPECTRUM to understand, optimize, and harness the beneficial effects of short-term stress, and minimize the harmful effects of chronic stress. One end of the spectrum is represented by GOOD stress that involves a rapid biological stress response mounted in the presence of the stressor, followed by a rapid shut-down of the response upon cessation of the stressor. Such responses induce physiological conditions that are likely to enhance protective immunity, mental and physical performance, and overall health. The opposite end of the spectrum is represented by BAD stress that involves chronic or long-term biological changes that are likely to result in dysregulation or suppression of immune function, a decrease in mental and physical performance, and an increased likelihood of disease. Short- and/or long- term stress is generally superimposed on a psycho-physiological RESTING STATE, or “green zone” of low/no stress that also represents a state of health maintenance/restoration. While the sympathetic nervous system and hypothalamic-pituitary-adrenal axis are the major drivers of the fight-or-flight response, the parasympathetic nervous system is the major driver of recovery from stress. The parasympathetic nervous system, together with negative feedback loops within physiological fight-or-flight systems, drives the rest-and-digest response that mediates recovery from stress and a return to resting state. In order to maintain health one needs to optimize GOOD stress, maximize the RESTING ZONE, and minimize BAD stress. This is likely to involve a multi-pronged approach [3; 6; 11; 32]: Sleep of a quality and duration that helps one feel rested in the morning, a moderate and healthy diet, and consistent and moderate exercise or physical activity, are three LIFESTYLE FACTORS that are likely to enable one to stay on the “good” side of the stress spectrum. Effective perspective-setting, appraisal/reappraisal, and coping mechanisms, social support, authenticity, genuine gratitude, and compassion towards others and oneself, are likely to provide PSYCHO-SOCIAL BUFFERS against bad stress, and to enable one to stay on the “good” side of the stress spectrum. Additionally, depending on individual preferences, ACTIVITIES such as, meditation, yoga, religious beliefs and activities, nature walks or hikes, exercise, dancing, music, art, craft, fishing, painting, etc. may also reduce BAD stress, extend the RESTING ZONE, and optimize GOOD stress. Such personal activities are likely to involve different strokes for different folks and need not always be meditative or reflective in nature. (Reprinted with permission from Springer Nature: Effects of stress on immune function: the good, the bad, and the beautiful. Dhabhar, F.S., Immunologic Research, 58, p 193–210, Copyright 2014.)
The longer one experiences chronic stress, the greater the chances of there being detrimental health effects. However, because most organisms are stress resilient, it often takes prolonged exposure to chronic stress to break down physiological systems. In order to stay healthy, one needs to minimize chronic stress, maximize the resting zone of low/no stress, and optimize the short-term/fight-or-flight stress response so that it is mounted rapidly and robustly when needed and shut down immediately after the cessation of stress (Figure 3). Attaining and maintaining physical, mental and emotional well-being involves a multi-pronged approach [3; 6; 246] (further studies are needed to test some of the following recommendations): Sleep, a moderated healthy diet, and exercise or physical activity [247], are three LIFESTYLE FACTORS that are likely to enable one to stay on the good side of the stress spectrum. In order to harness the beneficial effets of sleep, different individuals may require different amounts of sleep, and the same individual may require different amounts of sleep at different times in their life. Therefore, it is important to recognize that sleep of a quality and duration that helps one feel rested in the morning, is likely to be critical for optimizing ones Stress Spectrum. With regards to exercise, consistent moderate exercise is more sustainable and likely to confer greater stress-optimization and health benefits than inconsistent high intensity exercise. Similarly, with regards to diet, a moderate, enjoyable diet may be more sustainable and beneficial in the long run than extreme and hard-to-maintain dietary interventions. PSYCHO-SOCIAL FACTORS that can buffer against chronic stress, and enable one to stay on the good side of the stress spectrum include: The individual’s perception of the situation as being under their control[248; 249; 250; 251; 252], positive emotion regulation and reappraisal [253], coping ability/capacity/strategy[254][255], and perception of social support[18][256]. In contrast, when an individual appraises a stressor where the demands of dealing with the stressor exceed their resources and coping mechanisms, then such a situation is likely to contribute to chronic stress. While scientific studies are needed to test these hypotheses, it is likely that factors such as authenticity, gratitude, and compassion towards others and oneself may also buffer against chronic stress. In addition to lifestyle and psycho-social factors, depending on the personality and preferences of the individual, ACTIVITIES such as, meditation [257], mindfulness [258], yoga [259], religious beliefs and activities [260], exercise [247], dancing, music, art, craft, painting, [261; 262], contact with nature [263], as well as fishing, boating, birding, etc. (scientific studies are needed to investigate the effects of these activities) may also reduce BAD STRESS, maximize the RESTING ZONE, and optimize GOOD STRESS. Such personal activities are likely to involve different strokes for different folks and need not all be meditative or contemplative in nature [3; 11].
The Stress Spectrum, taken together with other information presented in this review, shows that the duration, intensity/concentration, and timing of exposure to stressor-induced physiological activation (neurotransmitters, hormones, and their molecular, cellular, organ-level and systemic effects) are critical for determining whether stress will enhance or inhibit immune function and mental and physical performance. While there is significant evidence in support of our Stress Spectrum Model (Figure 3) [3; 6; 11; 32], different aspects of the model need to be further investigated in preclinical, translational, and human subjects studies.
24. Conclusion
It is important to continue to elucidate mechanisms and translate findings described here from bench to bedside. However, this work is important because stress is a ubiquitous part of life, and can positively or negatively affect health and mental and physical performance. Chronic stress has long-been known to play a role in the etiology of numerous diseases, and exacts a tremendous cost from society. In contrast, short-term stress is one nature’s fundamental survival mechanisms that could be harnessed clinically to safely and effectively enhance immuno-protection, and functionally to enhance mental and physical performance. It is hoped that this review will provide a conceptual framework and targets for further investigation of mechanisms and conditions under which the adaptive and protective aspects of short-term stress or exercise can be optimized and harnessed, and for developing pharmacological and/or biobehavioral interventions to enhance health, healing, and mental and physical performance.
Highlights.
Stress has a “bad” reputation that deserves revision given recent preclinical and clinical findings
Chronic stress (weeks/months/years) has deleterious effects on brain, body & health
Fight-or-flight stress (minutes to hours) enhances immune-protection, and mental/physical performance
It would be beneficial to harness fight-or-flight physiology to enhance protection/performance during threat/challenge/opportunity
“The Stress Spectrum” provides a model for optimizing and harnessing “good” stress and minimizing “bad” stress
Acknowledgments
I am deeply grateful to Bruce McEwen for the invaluable support, guidance, and encouragement that he has given me starting even before I joined his laboratory at The Rockefeller University. He has shown all of us (who have been fortunate enough to be mentored by him) how to conduct innovative and rigorous science while caring for all those you work with. I thank all current and previous members of my laboratory, and particularly, Dr. Kavitha Viswanathan, Dr. Alison Saul, Jean Tillie, Christine Daugherty, Nicole Taylor, and Alesha Cox-Harris, whose work and publications are among those discussed in this manuscript. I also thank my mentors, colleagues and collaborators for their significant contributions. I am grateful for all the studies that we have done together and the fun that we have had doing them. I am also grateful for support from the following agencies and foundations that has been critical for enabling us to conduct our research: The NIH (AI48995, AR46299, CA107498), The Office of Naval Research, The Dana Foundation, and The John D. and Catherine T. MacArthur Foundation.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.McEwen BS. Protective and Damaging Effects of Stress Mediators. N Engl J Med. 1998;338:171–179. doi: 10.1056/NEJM199801153380307. [DOI] [PubMed] [Google Scholar]
- 2.Ader R. Psychoneuroimmunology IV. Academic Press; San Diego: 2007. [Google Scholar]
- 3.Dhabhar FS. Enhancing versus suppressive effects of stress on immune function: implications for immunoprotection and immunopathology. NeuroImmunoModulation. 2009;16:300–17. doi: 10.1159/000216188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Dhabhar FS, Malarkey WB, Neri E, McEwen BS. Stress-induced redistribution of immune cells - From barracks to boulevards to battlefields: A tale of three hormones. Psychoneuroendocrinology. 2012;37:1345–1368. doi: 10.1016/j.psyneuen.2012.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Padro CJ, Sanders VM. Neuroendocrine regulation of inflammation. Semin Immunol. 2014 doi: 10.1016/j.smim.2014.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dhabhar FS. Positive Effects of Stress. TED@Vancouver. 2012 http://talentsearch.ted.com/video/Firdaus-Dhabhar-The-positive-ef OR https://www.youtube.com/watch?v=nsc83N-Q1q4.
- 7.Dhabhar FS, Miller AH, McEwen BS, Spencer RL. Effects of stress on immune cell distribution -- dynamics and hormonal mechanisms. J Immunology. 1995;154:5511–5527. [PubMed] [Google Scholar]
- 8.Dhabhar FS, McEwen BS. Bidirectional Effects Of Stress & Glucocorticoid Hormones On Immune Function: Possible Explanations For Paradoxical Observations. In: Ader R, Felten DL, Cohen N, editors. Psychoneuroimmunology - Third Edition. Academic Press; San Diego: 2001. pp. 301–338. [Google Scholar]
- 9.Dhabhar FS, McEwen BS. Acute stress enhances while chronic stress suppresses immune function in vivo: A potential role for leukocyte trafficking. Brain Behavior & Immunity. 1997;11:286–306. doi: 10.1006/brbi.1997.0508. [DOI] [PubMed] [Google Scholar]
- 10.Dhabhar FS. Good Stress, Bad Stress, and Health, Invited Lecture at The Center for Compassion and Altruism Research and Education. Stanford University; 2014. [Google Scholar]
- 11.Dhabhar FS. Effects of stress on immune function: the good, the bad, and the beautiful. Immunologic research. 2014;58:193–210. doi: 10.1007/s12026-014-8517-0. [DOI] [PubMed] [Google Scholar]
- 12.Goldstein DS, McEwen B. Allostasis, homeostats, and the nature of stress. Stress. 2002;5:55–8. doi: 10.1080/102538902900012345. [DOI] [PubMed] [Google Scholar]
- 13.McEwen BS. The end of stress as we know it. Dana Press; Washington, DC: 2002. [Google Scholar]
- 14.Sapolsky RM. The influence of social hierarchy on primate health. Science. 2005;308:648–652. doi: 10.1126/science.1106477. [DOI] [PubMed] [Google Scholar]
- 15.Jezova D, Jurankova E, Mosnarova A, Kriska M, Skultetyova I. Neuroendocrine response during stress with relation to gender differences. Acta Neurobiol Exp (Wars) 1996;56:779–85. doi: 10.55782/ane-1996-1183. [DOI] [PubMed] [Google Scholar]
- 16.Jong TR, Menon R, Bludau A, Grund T, Biermeier V, Klampfl SM, Jurek B, Bosch OJ, Hellhammer J, Neumann ID. Salivary oxytocin concentrations in response to running, sexual self-stimulation, breastfeeding and the TSST: The Regensburg Oxytocin Challenge (ROC) study. Psychoneuroendocrinology. 2015;62:381–8. doi: 10.1016/j.psyneuen.2015.08.027. [DOI] [PubMed] [Google Scholar]
- 17.Steptoe A, Hamer M, Chida Y. The effects of acute psychological stress on circulating inflammatory factors in humans: a review and meta-analysis. Brain Behav Immun. 2007;21:901–12. doi: 10.1016/j.bbi.2007.03.011. [DOI] [PubMed] [Google Scholar]
- 18.Puterman E, Epel ES, O’Donovan A, Prather AA, Aschbacher K, Dhabhar FS. Anger Is Associated with Increased IL-6 Stress Reactivity in Women, But Only Among Those Low in Social Support. Int J Behav Med. 2013 doi: 10.1007/s12529-013-9368-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Aschbacher K, Epel E, Wolkowitz OM, Prather AA, Puterman E, Dhabhar FS. Maintenance of a positive outlook during acute stress protects against pro-inflammatory reactivity and future depressive symptoms. Brain Behav Immun. 2012;26:346–52. doi: 10.1016/j.bbi.2011.10.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Roseboom PH, Nanda SA, Bakshi VP, Trentani A, Newman SM, Kalin NH. Predator threat induces behavioral inhibition, pituitary-adrenal activation and changes in amygdala CRF-binding protein gene expression. Psychoneuroendocrinology. 2007;32:44–55. doi: 10.1016/j.psyneuen.2006.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Vendruscolo LF, Vendruscolo JC, Terenina-Rigaldie E, Raba F, Ramos A, Takahashi RN, Mormede P. Genetic influences on behavioral and neuroendocrine responses to predator-odor stress in rats. Neurosci Lett. 2006;409:89–94. doi: 10.1016/j.neulet.2006.07.026. [DOI] [PubMed] [Google Scholar]
- 22.Kirschbaum C, Pirke KM, Hellhammer DH. The ‘Trier Social Stress Test’--a tool for investigating psychobiological stress responses in a laboratory setting. Neuropsychobiology. 1993;28:76–81. doi: 10.1159/000119004. [DOI] [PubMed] [Google Scholar]
- 23.Foley P, Kirschbaum C. Human hypothalamus-pituitary-adrenal axis responses to acute psychosocial stress in laboratory settings. Neurosci Biobehav Rev. 2010;35:91–6. doi: 10.1016/j.neubiorev.2010.01.010. [DOI] [PubMed] [Google Scholar]
- 24.Henze GI, Zankert S, Urschler DF, Hiltl TJ, Kudielka BM, Pruessner JC, Wust S. Testing the ecological validity of the Trier Social Stress Test: Association with real-life exam stress. Psychoneuroendocrinology. 2017;75:52–55. doi: 10.1016/j.psyneuen.2016.10.002. [DOI] [PubMed] [Google Scholar]
- 25.Kotwica G, Franczak A, Okrasa S, Koziorowski M, Kurowicka B. Effects of mating stimuli and oxytocin on plasma cortisol concentration in gilts. Reprod Biol. 2002;2:25–37. [PubMed] [Google Scholar]
- 26.Veronesi MC, Tosi U, Villani M, Govoni N, Faustini M, Kindahl H, Madej A, Carluccio A. Oxytocin, vasopressin, prostaglandin F(2alpha), luteinizing hormone, testosterone, estrone sulfate, and cortisol plasma concentrations after sexual stimulation in stallions. Theriogenology. 2010;73:460–7. doi: 10.1016/j.theriogenology.2009.09.028. [DOI] [PubMed] [Google Scholar]
- 27.Fox CA, Fox B. A comparative study of coital physiology, with special reference to the sexual climax. J Reprod Fertil. 1971;24:319–36. doi: 10.1530/jrf.0.0240319. [DOI] [PubMed] [Google Scholar]
- 28.Leuner B, Glasper ER, Gould E. Sexual experience promotes adult neurogenesis in the hippocampus despite an initial elevation in stress hormones. PLoS One. 2010;5:e11597. doi: 10.1371/journal.pone.0011597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Dhabhar FS, Miller AH, Stein M, McEwen BS, Spencer RL. Diurnal and stress-induced changes in distribution of peripheral blood leukocyte subpopulations. Brain Behav Immun. 1994;8:66–79. doi: 10.1006/brbi.1994.1006. [DOI] [PubMed] [Google Scholar]
- 30.Dhabhar FS, McEwen BS. Stress-induced enhancement of antigen-specific cell-mediated immunity. J Immunology. 1996;156:2608–2615. [PubMed] [Google Scholar]
- 31.Selye H. Stress without distress. Nal Penguin Inc; New York: 1974. [Google Scholar]
- 32.Dhabhar FS, McEwen BS. Bidirectional effects of stress on immune function: Possible explanations for salubrious as well as harmful effects. In: Ader R, editor. Psychoneuroimmunology IV. Elsevier Academic Press; San Diego: 2007. pp. 723–760. [Google Scholar]
- 33.Dhabhar FS. A Hassle a Day May Keep the Pathogens Away: The Fight-Or-Flight Stress Response and the Augmentation of Immune Function. Integrative and comparative biology. 2009;49:215–236. doi: 10.1093/icb/icp045. [DOI] [PubMed] [Google Scholar]
- 34.Irwin M, Patterson T, Smith TL, Caldwell C, Brown SA, Gillin CJ, Grant I. Reduction of immune function in life stress and depression. Biol Psychiatry. 1990;27:22–30. doi: 10.1016/0006-3223(90)90016-u. [DOI] [PubMed] [Google Scholar]
- 35.Glaser R, Kiecolt-Glaser JK. Stress-induced immune dysfunction: implications for health. Nat Rev Immunol. 2005;5:243–51. doi: 10.1038/nri1571. [DOI] [PubMed] [Google Scholar]
- 36.Chrousos GP, Kino T. Glucocorticoid action networks and complex psychiatric and/or somatic disorders. Stress. 2007;10:213–9. doi: 10.1080/10253890701292119. [DOI] [PubMed] [Google Scholar]
- 37.Dhabhar FS, Viswanathan K. Short-Term Stress Experienced At The Time Of Immunization Induces A Long-lasting Increase In Immunological Memory. Am J Physiol Regul Integr Comp Physiol. 2005;289:R738–744. doi: 10.1152/ajpregu.00145.2005. [DOI] [PubMed] [Google Scholar]
- 38.Sephton S, Spiegel D. Circadian disruption in cancer: a neuroendocrine-immune pathway from stress to disease? Brain Behav Immun. 2003;17:321–8. doi: 10.1016/s0889-1591(03)00078-3. [DOI] [PubMed] [Google Scholar]
- 39.Saul AN, Oberyszyn TM, Daugherty C, Kusewitt D, Jones S, Jewell S, Malarkey WB, Lehman A, Lemeshow S, Dhabhar FS. Chronic stress and susceptibility to skin cancer. J Nat Cancer Institute. 2005;97:1760–1767. doi: 10.1093/jnci/dji401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Gunnar M, Quevedo K. The neurobiology of stress and development. Annu Rev Psychol. 2007;58:145–73. doi: 10.1146/annurev.psych.58.110405.085605. [DOI] [PubMed] [Google Scholar]
- 41.Dhabhar FS, McEwen BS, Spencer RL. Stress response, adrenal steroid receptor levels, and corticosteroid-binding globulin levels -- a comparison between Sprague Dawley, Fischer 344, and Lewis rats. Brain Research. 1993;616:89–98. doi: 10.1016/0006-8993(93)90196-t. [DOI] [PubMed] [Google Scholar]
- 42.Sternberg EM, Hill JM, Chrousos GP, Kamilaris T, Listwak SJ, Gold PW, Wilder RL. Inflammatory mediator-induced hypothalamic-pituitary-adrenal axis activation is defective in streptococcal cell wall arthritis-susceptible Lewis rats. Proc Natl Acad Sci USA. 1989;86:2374–2378. doi: 10.1073/pnas.86.7.2374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Dhabhar FS, McEwen BS, Spencer RL. Adaptation to prolonged or repeated stress -- Comparison between rat strains showing intrinsic differences in reactivity to acute stress. Neuroendocrinology. 1997;65:360–368. doi: 10.1159/000127196. [DOI] [PubMed] [Google Scholar]
- 44.Dhabhar FS, Miller AH, McEwen BS, Spencer RL. Differential activation of adrenal steroid receptors in neural and immune tissues of Sprague Dawley, Fischer 344, and Lewis rats. J Neuroimmunol. 1995;56:77–90. doi: 10.1016/0165-5728(94)00135-b. [DOI] [PubMed] [Google Scholar]
- 45.Gomez-Serrano M, Tonelli L, Listwak S, Sternberg E, Riley AL. Effects of cross fostering on open-field behavior, acoustic startle, lipopolysaccharide-induced corticosterone release, and body weight in Lewis and Fischer rats. Behav Genet. 2001;31:427–36. doi: 10.1023/a:1012742405141. [DOI] [PubMed] [Google Scholar]
- 46.Schwab CL, Fan R, Zheng Q, Myers LP, Hebert P, Pruett SB. Modeling and predicting stress-induced immunosuppression in mice using blood parameters. Toxicol Sci. 2005;83:101–13. doi: 10.1093/toxsci/kfi014. [DOI] [PubMed] [Google Scholar]
- 47.Benschop RJ, Rodriguez-Feuerhahn M, Schedlowski M. Catecholamine-induced leukocytosis: early observations, current research, and future directions. Brain, Behavior, & Immunity. 1996;10:77–91. doi: 10.1006/brbi.1996.0009. [DOI] [PubMed] [Google Scholar]
- 48.Schleimer RP, Claman HN, Oronsky A, editors. Anti-inflammatory steroid action: Basic and clinical aspects. Academic Press Inc; San Diego: 1989. [Google Scholar]
- 49.Dopp JM, Miller GE, Myers HF, Fahey JL. Increased natural killer-cell mobilization and cytotoxicity during marital conflict. Brain Behav Immun. 2000;14:10–26. doi: 10.1006/brbi.1999.0567. [DOI] [PubMed] [Google Scholar]
- 50.Segerstrom SC, Miller GE. Psychological stress and the human immune system: a meta-analytic study of 30 years of inquiry. Psychol Bull. 2004;130:601–30. doi: 10.1037/0033-2909.130.4.601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Vivier E, Malissen B. Innate and adaptive immunity: specificities and signaling hierarchies revisited. Nat Immunol. 2005;6:17–21. doi: 10.1038/ni1153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Adamo SA. Bidirectional connections between the immune and the nervous system in insects. In: Beckage NE, editor. Insect Immunology. Academic Press; Amsterdam, Boston: 2008. p. 348. [Google Scholar]
- 53.Baines D, Downer RG. Octopamine enhances phagocytosis in cockroach hemocytes: involvement of inositol trisphosphate. Archives of Insect Biochemistry & Physiology. 1994;26:249–61. doi: 10.1002/arch.940260402. [DOI] [PubMed] [Google Scholar]
- 54.Van Gaal LF, Mertens IL, De Block CE. Mechanisms linking obesity with cardiovascular disease. Nature. 2006;444:875–880. doi: 10.1038/nature05487. [DOI] [PubMed] [Google Scholar]
- 55.Dantzer R, O’Connor JC, Freund GG, Johnson RW, Kelley KW. From inflammation to sickness and depression: when the immune system subjugates the brain. Nature Reviews Neuroscience. 2008;9:46–56. doi: 10.1038/nrn2297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Maes MA. A review on the acute phase response in major depression. Rev Neurosci. 1993;4:407–416. doi: 10.1515/revneuro.1993.4.4.407. [DOI] [PubMed] [Google Scholar]
- 57.Simpson E. Special regulatory T-cell review: Regulation of immune responses--examining the role of T cells. Immunology. 2008;123:13–6. doi: 10.1111/j.1365-2567.2007.02775.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Piccirillo CA. Regulatory T cells in health and disease. Cytokine. 2008;43:395–401. doi: 10.1016/j.cyto.2008.07.469. [DOI] [PubMed] [Google Scholar]
- 59.Wing K, Sakaguchi S. Regulatory T cells exert checks and balances on self tolerance and autoimmunity. Nat Immunol. 2010;11:7–13. doi: 10.1038/ni.1818. [DOI] [PubMed] [Google Scholar]
- 60.Bluestone JA, Tang Q. How do CD4+CD25+ regulatory T cells control autoimmunity? Curr Opin Immunol. 2005;17:638–42. doi: 10.1016/j.coi.2005.09.002. [DOI] [PubMed] [Google Scholar]
- 61.Finn OJ. Cancer Immunology. N Engl J Med. 2008;358:2704–2715. doi: 10.1056/NEJMra072739. [DOI] [PubMed] [Google Scholar]
- 62.Olson BM, McNeel DG. Monitoring regulatory immune responses in tumor immunotherapy clinical trials. Front Oncol. 2013;3:109. doi: 10.3389/fonc.2013.00109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Whiteside TL. Regulatory T cell subsets in human cancer: are they regulating for or against tumor progression? Cancer Immunol Immunother. 2014;63:67–72. doi: 10.1007/s00262-013-1490-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Sprent J, Tough DF. Lymphocyte life-span and memory. Science. 1994;265:1395–1400. doi: 10.1126/science.8073282. [DOI] [PubMed] [Google Scholar]
- 65.Pickford GE, Srivastava AK, Slicher AM, Pang PKT. The stress response in the abundance of circulating leukocytes in the Killifish, Fundulus heteroclitus. I The cold-shock sequence and the effects of hypophysectomy. J Exp Zool. 1971;177:89–96. doi: 10.1002/jez.1401770110. [DOI] [PubMed] [Google Scholar]
- 66.Bilbo SD, Dhabhar FS, Viswanathan K, Saul A, Yellon SM, Nelson RJ. Short day lengths augment stress-induced leukocyte trafficking and stress-induced enhancement of skin immune function. Proc Natl Acad Sci U S A. 2002;99:4067–72. doi: 10.1073/pnas.062001899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Jensen MM. Changes in leukocyte counts associated with various stressors. J Reticuloendothelial Soc. 1969;8:457–465. [PubMed] [Google Scholar]
- 68.Viswanathan K, Dhabhar FS. Stress-induced enhancement of leukocyte trafficking into sites of surgery or immune activation. PNAS, USA. 2005;102:5808–5813. doi: 10.1073/pnas.0501650102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Dhabhar FS, Miller AH, McEwen BS, Spencer RL. Stress-induced changes in blood leukocyte distribution -- role of adrenal steroid hormones. J Immunology. 1996;157:1638–1644. [PubMed] [Google Scholar]
- 70.Rinder CS, Mathew JP, Rinder HM, Tracey JB, Davis E, Smith BR. Lymphocyte and monocyte subset changes during cardiopulmonary bypass: effects of aging and gender [see comments] Journal of Laboratory & Clinical Medicine. 1997;129:592–602. doi: 10.1016/s0022-2143(97)90193-1. [DOI] [PubMed] [Google Scholar]
- 71.Toft P, Svendsen P, Tonnesen E, Rasmussen JW, Christensen NJ. Redistribution of lymphocytes after major surgical stress. Acta Anesthesiol Scand. 1993;37:245–249. doi: 10.1111/j.1399-6576.1993.tb03708.x. [DOI] [PubMed] [Google Scholar]
- 72.Snow DH, Ricketts SW, Mason DK. Hematological responses to racing and training exercise in Thoroughbred horses, with particular reference to the leukocyte response. Equine Vet J. 1983;15:149–154. doi: 10.1111/j.2042-3306.1983.tb01742.x. [DOI] [PubMed] [Google Scholar]
- 73.Morrow-Tesch JL, McGlone JJ, Norman RL. Consequences of restraint stress on natural killer cell activity, behavior, and hormone levels in Rhesus Macaques (Macaca mulatta) Psychoneuroendocrinol. 1993;18:383–395. doi: 10.1016/0306-4530(93)90013-b. [DOI] [PubMed] [Google Scholar]
- 74.Herbert TB, Cohen S. Stress and immunity in humans: a meta-analytic review. Psychosom Med. 1993;55:364–379. doi: 10.1097/00006842-199307000-00004. [DOI] [PubMed] [Google Scholar]
- 75.Schedlowski M, Jacobs R, Stratman G, Richter S, Hädike A, Tewes U, Wagner TOF, Schmidt RE. Changes of natural killer cells during acute psychological stress. J Clin Immunol. 1993;13:119–126. doi: 10.1007/BF00919268. [DOI] [PubMed] [Google Scholar]
- 76.Mills PJ, Ziegler MG, Rehman J, Maisel AS. Catecholamines, catecholamine receptors, cell adhesion molecules, and acute stressor-related changes in cellular immunity. Adv Pharmacol. 1998;42:587–90. doi: 10.1016/s1054-3589(08)60819-4. [DOI] [PubMed] [Google Scholar]
- 77.Redwine L, Mills PJ, Sada M, Dimsdale J, Patterson T, Grant I. Differential immune cell chemotaxis responses to acute psychological stress in Alzheimer caregivers compared to non-caregiver controls. Psychosom Med. 2004;66:770–5. doi: 10.1097/01.psy.0000138118.62018.87. [DOI] [PubMed] [Google Scholar]
- 78.Bosch JA, Berntson GG, Cacioppo JT, Dhabhar FS, Marucha PT. Acute stress evokes selective mobilization of T cells that differ in chemokine receptor expression: A potential pathway linking immunologic reactivity to cardiovascular disease. Brain, Behavior and Immunity. 2003;17:251–259. doi: 10.1016/s0889-1591(03)00054-0. [DOI] [PubMed] [Google Scholar]
- 79.Rosenberger PH, Ickovics JR, Epel E, Nadler E, Jokl P, Fulkerson JP, Tillie JM, Dhabhar FS. Surgery stress induced immune cell redistribution profiles predict short- and long-term postsurgical recovery: A prospective study. Journal of Bone and Joint Surgery. 2009;91:2783–2794. doi: 10.2106/JBJS.H.00989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Hoagland H, Elmadjian F, Pincus G. Stressful psychomotor performance and adrenal cortical function as indicated by the lymphocyte reponse. J Clin Endocrinol. 1946;6:301–311. doi: 10.1210/jcem-6-4-301. [DOI] [PubMed] [Google Scholar]
- 81.Fauci AS, Dale DC. The effect of in vivo hydrocortisone on subpopulations of human lymphocytes. J Clin Invest. 1974;53:240–246. doi: 10.1172/JCI107544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Fauci AS, Dale DC. The effect of hydrocortisone on the kinetics of normal human lymphocytes. Blood. 1975;46:235–243. [PubMed] [Google Scholar]
- 83.Carlson SL, Fox S, Abell KM. Catecholamine modulation of lymphocyte homing to lymphoid tissues. Brain, Behavior, & Immunity. 1997;11:307–20. doi: 10.1006/brbi.1997.0501. [DOI] [PubMed] [Google Scholar]
- 84.Benschop RJ, Oostveen FG, Heijnen CJ, Ballieux RE. Beta 2-adrenergic stimulation causes detachment of natural killer cells from cultured endothelium. Eur J Immunol. 1993;23:3242–3247. doi: 10.1002/eji.1830231230. [DOI] [PubMed] [Google Scholar]
- 85.Redwine L, Snow S, Mills P, Irwin M. Acute psychological stress: effects on chemotaxis and cellular adhesion molecule expression. Psychosom Med. 2003;65:598–603. doi: 10.1097/01.psy.0000079377.86193.a8. [DOI] [PubMed] [Google Scholar]
- 86.Mills PJ, Meck JV, Waters WW, D’Aunno D, Ziegler MG. Peripheral leukocyte subpopulations and catecholamine levels in astronauts as a function of mission duration. Psychosom Med. 2001;63:886–90. doi: 10.1097/00006842-200111000-00006. [DOI] [PubMed] [Google Scholar]
- 87.Dhabhar FS, McEwen BS. Enhancing versus suppressive effects of stress hormones on skin immune function. PNAS, USA. 1999;96:1059–1064. doi: 10.1073/pnas.96.3.1059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Dhabhar FS, Satoskar AR, Bluethmann H, David JR, McEwen BS. Stress-Induced Enhancement of Skin Immune Function: A Role For IFNg. PNAS, USA. 2000;97:2846–2851. doi: 10.1073/pnas.050569397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Blecha F, Barry RA, Kelley KW. Stress-induced alterations in delayed-type hypersensitivity to SRBC and contact sensitivity to DNFB in mice. Proc Soc Exp Biol Med. 1982;169:239–246. doi: 10.3181/00379727-169-41338. [DOI] [PubMed] [Google Scholar]
- 90.Wood PG, Karol MH, Kusnecov AW, Rabin BS. Enhancement of antigen-specific humoral and cell-mediated immunity by electric footshock stress in rats. Brain, Behav, Immun. 1993;7:121–134. doi: 10.1006/brbi.1993.1014. [DOI] [PubMed] [Google Scholar]
- 91.Coe CL, Lubach G, Ershler WB. Immunological consequences of maternal separation in infant primates. In: Lewis M, Worobey J, editors. Infant stress and coping. Jossey-Bass Inc; New York, NY: 1989. pp. 64–91. [DOI] [PubMed] [Google Scholar]
- 92.Saint-Mezard P, Chavagnac C, Bosset S, Ionescu M, Peyron E, Kaiserlian D, Nicolas JF, Berard F. Psychological stress exerts an adjuvant effect on skin dendritic cell functions in vivo. J Immunol. 2003;171(8):4073–80. doi: 10.4049/jimmunol.171.8.4073. [DOI] [PubMed] [Google Scholar]
- 93.Viswanathan K, Daugherty C, Dhabhar FS. Stress as an endogenous adjuvant: augmentation of the immunization phase of cell-mediated immunity. International Immunology. 2005;17:1059–1069. doi: 10.1093/intimm/dxh286. [DOI] [PubMed] [Google Scholar]
- 94.Flint MS, Miller DB, Tinkle SS. Restraint-induced modulation of allergic and irritant contact dermatitis in male and female B6.129 mice. Brain Behav Immun. 2000;14:256–69. doi: 10.1006/brbi.2000.0604. [DOI] [PubMed] [Google Scholar]
- 95.Flint MS, Valosen JM, Johnson EA, Miller DB, Tinkle SS. Restraint stress applied prior to chemical sensitization modulates the development of allergic contact dermatitis differently than restraint prior to challenge. J Neuroimmunol. 2001;113:72–80. doi: 10.1016/s0165-5728(00)00413-6. [DOI] [PubMed] [Google Scholar]
- 96.Bilbo SD, Hotchkiss AK, Chiavegatto S, Nelson RJ. Blunted stress responses in delayed type hypersensitivity in mice lacking the neuronal isoform of nitric oxide synthase. J Neuroimmunol. 2003;140:41–8. doi: 10.1016/s0165-5728(03)00175-9. [DOI] [PubMed] [Google Scholar]
- 97.Kripke ML. Ultraviolet radiation and immunology: something new under the sun-- presidential address. Cancer Res. 1994;54:6102–5. [PubMed] [Google Scholar]
- 98.Granstein RD, Matsui MS. UV radiation-induced immunosuppression and skin cancer. Cutis. 2004;74:4–9. [PubMed] [Google Scholar]
- 99.Dhabhar FS, Saul AN, Daugherty C, Holmes TH, Bouley DM, Oberyszyn TM. Short-Term stress enhances cellular immunity and increases early resistance to squamous cell carcinoma. Brain, Behavior, and Immunity. 2010;24:127–37. doi: 10.1016/j.bbi.2009.09.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Amkraut A, Solomon GF. Stress and murine sarcoma virus (Moloney)-induced tumors. Cancer Res. 1972;32:1428–33. [PubMed] [Google Scholar]
- 101.Schmitz KH. Exercise for secondary prevention of breast cancer: moving from evidence to changing clinical practice. Cancer prevention research (Philadelphia, Pa) 2011;4:476–80. doi: 10.1158/1940-6207.CAPR-11-0097. [DOI] [PubMed] [Google Scholar]
- 102.Pierce JP, Stefanick ML, Flatt SW, Natarajan L, Sternfeld B, Madlensky L, Al-Delaimy WK, Thomson CA, Kealey S, Hajek R, Parker BA, Newman VA, Caan B, Rock CL. Greater survival after breast cancer in physically active women with high vegetable-fruit intake regardless of obesity. J Clin Oncol. 2007;25:2345–51. doi: 10.1200/JCO.2006.08.6819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Speck RM, Courneya KS, Masse LC, Duval S, Schmitz KH. An update of controlled physical activity trials in cancer survivors: a systematic review and meta-analysis. J Cancer Surviv. 2010;4:87–100. doi: 10.1007/s11764-009-0110-5. [DOI] [PubMed] [Google Scholar]
- 104.Holick CN, Newcomb PA, Trentham-Dietz A, Titus-Ernstoff L, Bersch AJ, Stampfer MJ, Baron JA, Egan KM, Willett WC. Physical activity and survival after diagnosis of invasive breast cancer. Cancer Epidemiol Biomarkers Prev. 2008;17:379–86. doi: 10.1158/1055-9965.EPI-07-0771. [DOI] [PubMed] [Google Scholar]
- 105.McNeely ML, Campbell KL, Rowe BH, Klassen TP, Mackey JR, Courneya KS. Effects of exercise on breast cancer patients and survivors: a systematic review and meta-analysis. CMAJ. 2006;175:34–41. doi: 10.1503/cmaj.051073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Betof AS, Dewhirst MW, Jones LW. Effects and potential mechanisms of exercise training on cancer progression: a translational perspective. Brain Behav Immun. 2013;30(Suppl):S75–87. doi: 10.1016/j.bbi.2012.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Solomon GF, Merigan TC, Levine S. Variation in adrenal cortical hormones within physiologic ranges, stress and interferon production in mice. Riv Patol Nerv Ment. 1966;87:74–9. doi: 10.3181/00379727-126-32370. [DOI] [PubMed] [Google Scholar]
- 108.Nance DM, Sanders VM. Autonomic innervation and regulation of the immune system (1987–2007) Brain Behav Immun. 2007;21:736–45. doi: 10.1016/j.bbi.2007.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Sanders VM. The beta2-adrenergic receptor on T and B lymphocytes: do we understand it yet? Brain Behav Immun. 2012;26:195–200. doi: 10.1016/j.bbi.2011.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Altemus M, Rao B, Dhabhar FS, Ding W, Granstein R. Stress-induced changes in skin barrier function in healthy women. J Investigative Dermatology. 2001;117:309–317. doi: 10.1046/j.1523-1747.2001.01373.x. [DOI] [PubMed] [Google Scholar]
- 111.Pace TW, Mletzko TC, Alagbe O, Musselman DL, Nemeroff CB, Miller AH, Heim CM. Increased stress-induced inflammatory responses in male patients with major depression and increased early life stress. Am J Psychiatry. 2006;163:1630–3. doi: 10.1176/ajp.2006.163.9.1630. [DOI] [PubMed] [Google Scholar]
- 112.Prather AA, Puterman E, Epel ES, Dhabhar FS. Poor sleep quality potentiates stress-induced cytokine reactivity in postmenopausal women with high visceral abdominal adiposity. Brain, behavior, and immunity. 2013 doi: 10.1016/j.bbi.2013.09.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Capuron L, Miller AH. Immune system to brain signaling: neuropsychopharmacological implications. Pharmacol Ther. 2011;130:226–38. doi: 10.1016/j.pharmthera.2011.01.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Maes M. Depression is an inflammatory disease,but cell-mediated immune activation is the key component of depression. Prog Neuropsychopharmacol Biol Psychiatry. 2011;35:664–75. doi: 10.1016/j.pnpbp.2010.06.014. [DOI] [PubMed] [Google Scholar]
- 115.Miller AH, Haroon E, Raison CL, Felger JC. Cytokine targets in the brain: impact on neurotransmitters and neurocircuits. Depress Anxiety. 2013;30:297–306. doi: 10.1002/da.22084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Andreasson A, Arborelius L, Erlanson-Albertsson C, Lekander M. A putative role for cytokines in the impaired appetite in depression. Brain Behav Immun. 2007;21:147–52. doi: 10.1016/j.bbi.2006.08.002. [DOI] [PubMed] [Google Scholar]
- 117.Quan N. Brain’s firewall: blood-brain barrier actively regulates neuroimmune information flow. Brain Behav Immun. 2006;20:447–8. doi: 10.1016/j.bbi.2006.03.004. [DOI] [PubMed] [Google Scholar]
- 118.Quan N. In-depth conversation: Spectrum and kinetics of neuroimmune afferent pathways. Brain Behav Immun. 2014 doi: 10.1016/j.bbi.2014.02.006. No of pages, 8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Besedovsky HO, Rey AD. Physiology of psychoneuroimmunology: a personal view. Brain Behav Immun. 2007;21:34–44. doi: 10.1016/j.bbi.2006.09.008. [DOI] [PubMed] [Google Scholar]
- 120.Schedlowski M, Engler H, Grigoleit JS. Endotoxin-induced experimental systemic inflammation in humans: a model to disentangle immune-to-brain communication. Brain Behav Immun. 2014;35:1–8. doi: 10.1016/j.bbi.2013.09.015. [DOI] [PubMed] [Google Scholar]
- 121.Dhabhar FS, Burke HM, Epel ES, Mellon SH, Rosser R, Reus VI, Wolkowitz OM. Low serum IL-10 concentrations and loss of regulatory association between IL-6 and IL-10 in adults with major depression. J Psychiatr Res. 2009;43:962–9. doi: 10.1016/j.jpsychires.2009.05.010. [DOI] [PubMed] [Google Scholar]
- 122.Pace TW, Hu F, Miller AH. Cytokine-effects on glucocorticoid receptor function: relevance to glucocorticoid resistance and the pathophysiology and treatment of major depression. Brain Behav Immun. 2007;21:9–19. doi: 10.1016/j.bbi.2006.08.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Pace TW, Miller AH. Cytokines and glucocorticoid receptor signaling. Relevance to major depression. Ann N Y Acad Sci. 2009;1179:86–105. doi: 10.1111/j.1749-6632.2009.04984.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Dantzer R, O’Connor JC, Lawson MA, Kelley KW. Inflammation-associated depression: from serotonin to kynurenine. Psychoneuroendocrinology. 2011;36:426–36. doi: 10.1016/j.psyneuen.2010.09.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Kelley KW, Dantzer R. Alcoholism and inflammation: neuroimmunology of behavioral and mood disorders. Brain Behav Immun. 2011;25(Suppl 1):S13–20. doi: 10.1016/j.bbi.2010.12.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Altemus M, Cloitre M, Dhabhar FS. Cellular Immune Response in Adult Women with PTSD Related to Childhood Abuse. American Journal of Psychiatry. 2003;160:1705–1707. doi: 10.1176/appi.ajp.160.9.1705. [DOI] [PubMed] [Google Scholar]
- 127.Pace TW, Heim CM. A short review on the psychoneuroimmunology of posttraumatic stress disorder: from risk factors to medical comorbidities. Brain Behav Immun. 2011;25:6–13. doi: 10.1016/j.bbi.2010.10.003. [DOI] [PubMed] [Google Scholar]
- 128.Wieck A, Grassi-Oliveira R, Hartmann do Prado C, Teixeira AL, Bauer ME. Neuroimmunoendocrine interactions in post-traumatic stress disorder: focus on long-term implications of childhood maltreatment. Neuroimmunomodulation. 2014;21:145–51. doi: 10.1159/000356552. [DOI] [PubMed] [Google Scholar]
- 129.Fredericks CA, Drabant EM, Edge MD, Tillie JM, Hallmayer J, Ramel W, Kuo JR, Mackey S, Gross JJ, Dhabhar FS. Healthy young women with serotonin transporter SS polymorphism show a pro-inflammatory bias under resting and stress conditions. Brain Behav Immun. 2009;24:350–357. doi: 10.1016/j.bbi.2009.10.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Wiegers GJ, Labeur MS, Stec IE, Klinkert WE, Holsboer F, Reul JM. Glucocorticoids accelerate anti-T cell receptor-induced T cell growth. J Immunol. 1995;155:1893–1902. [PubMed] [Google Scholar]
- 131.Straub RH, Bijlsma JW, Masi A, Cutolo M. Role of neuroendocrine and neuroimmune mechanisms in chronic inflammatory rheumatic diseases--the 10-year update. Semin Arthritis Rheum. 2013;43:392–404. doi: 10.1016/j.semarthrit.2013.04.008. [DOI] [PubMed] [Google Scholar]
- 132.Straub RH. Evolutionary medicine and chronic inflammatory state--known and new concepts in pathophysiology. J Mol Med (Berl) 2012;90:523–34. doi: 10.1007/s00109-012-0861-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Straub RH. Interaction of the endocrine system with inflammation: a function of energy and volume regulation. Arthritis Res Ther. 2014;16:203. doi: 10.1186/ar4484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Juster RP, McEwen BS, Lupien SJ. Allostatic load biomarkers of chronic stress and impact on health and cognition. Neurosci Biobehav Rev. 2010;35:2–16. doi: 10.1016/j.neubiorev.2009.10.002. [DOI] [PubMed] [Google Scholar]
- 135.Danese A, McEwen BS. Adverse childhood experiences, allostasis, allostatic load, and age-related disease. Physiol Behav. 2012;106:29–39. doi: 10.1016/j.physbeh.2011.08.019. [DOI] [PubMed] [Google Scholar]
- 136.Solomon GF. Emotions, stress, the central nervous system, and immunity. Ann N Y Acad Sci. 1969;164:335–43. doi: 10.1111/j.1749-6632.1969.tb14048.x. [DOI] [PubMed] [Google Scholar]
- 137.Vitlic A, Lord JM, Phillips AC. Stress, ageing and their influence on functional, cellular and molecular aspects of the immune system. Age (Dordr) 2014:1–7. doi: 10.1007/s11357-014-9631-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Webster Marketon JI, Glaser R. Stress hormones and immune function. Cell Immunol. 2008;252:16–26. doi: 10.1016/j.cellimm.2007.09.006. [DOI] [PubMed] [Google Scholar]
- 139.Fagundes CP, Glaser R, Kiecolt-Glaser JK. Stressful early life experiences and immune dysregulation across the lifespan. Brain Behav Immun. 2013;27:8–12. doi: 10.1016/j.bbi.2012.06.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Sanders VM, Straub RH. Norepinephrine, The Beta-Adrenergic Receptor, and Immunity. Brain, Behavior, and Immunity. 2002;16:290–332. doi: 10.1006/brbi.2001.0639. [DOI] [PubMed] [Google Scholar]
- 141.Irwin MR. Human psychoneuroimmunology: 20 Years of discovery. Brain, Behavior, and Immunity. 2008;22:129–139. doi: 10.1016/j.bbi.2007.07.013. [DOI] [PubMed] [Google Scholar]
- 142.APA. Stress In America, Stress In America. American Psychological Association; Washington, DC: 2007. pp. 1–19. [Google Scholar]
- 143.Smith A, Vollmer-Conna U, Bennett B, Wakefield D, Hickie I, Lloyd A. The relationship between distress and the development of a primary immune response to a novel antigen. Brain Behav Immun. 2004;18:65–75. doi: 10.1016/s0889-1591(03)00107-7. [DOI] [PubMed] [Google Scholar]
- 144.Sephton SE, Dhabhar FS, Keuroghlian AS, Giese-Davis J, McEwen BS, Ionan AC, Spiegel D. Depression, cortisol, and suppressed cell-mediated immunity in metastatic breast cancer. Brain Behav Immun. 2009;23:1148–55. doi: 10.1016/j.bbi.2009.07.007. [DOI] [PubMed] [Google Scholar]
- 145.Kelley KW, Greenfield RE, Evermann JF, Parish SM, Perryman LE. Delayed-type hypersensitivity, contact sensitivity, and PHA skin-test responses of heat- and cold-stressed calves. Am J Vet Res. 1982;43:775–779. [PubMed] [Google Scholar]
- 146.Edwards EA, Dean LM. Effects of crowding of mice on humoral antibody formation and protection to lethal antigenic challenge. Psychosom Med. 1977;39:19–24. doi: 10.1097/00006842-197701000-00003. [DOI] [PubMed] [Google Scholar]
- 147.Fleshner M, Laudenslager ML, Simons L, Maier SF. Reduced serum antibodies associated with social defeat in rats. Physiol & Behav. 1989;45:1183–1187. doi: 10.1016/0031-9384(89)90107-8. [DOI] [PubMed] [Google Scholar]
- 148.Bartrop R, Lazarus L, Luckhurst E, Kiloh LG, Penny R. Depressed lymphocyte function after bereavement. The Lancet. 1977;1:834–836. doi: 10.1016/s0140-6736(77)92780-5. [DOI] [PubMed] [Google Scholar]
- 149.Kiecolt-Glaser JK, Garner W, Speicher C, Penn GM, Holliday J, Glaser R. Psychosocial modifiers of immunocompetence in medical students. Psychosom Med. 1984;46:7–14. doi: 10.1097/00006842-198401000-00003. [DOI] [PubMed] [Google Scholar]
- 150.Cheng GJ, Morrow-Tesch JL, Beller DI, Levy EM, Black PH. Immunosuppression in mice induced by cold water stress. Brain, Behavior, & Immunity. 1990;4:278–291. doi: 10.1016/0889-1591(90)90032-l. [DOI] [PubMed] [Google Scholar]
- 151.Regnier JA, Kelley KW. Heat- and cold-stress suppresses in vivo and in vitro cellular immune response of chickens. Am J Vet Res. 1981;42:294–299. [PubMed] [Google Scholar]
- 152.Wistar R, Hildemann WH. Effect of stress on skin transplantation immunity in mice. Science. 1960;131:159–160. doi: 10.1126/science.131.3394.159. [DOI] [PubMed] [Google Scholar]
- 153.Bonneau RH, Sheridan JF, Feng N, Glaser R. Stress-induced effects on cell-mediated innate and adaptive memory components of the murine immune response to herpes simplex virus infection. Brain, Behavior, Immunity. 1991;5:274–295. doi: 10.1016/0889-1591(91)90023-4. [DOI] [PubMed] [Google Scholar]
- 154.Brown DH, Zwilling BS. Activation of the hypothalamic-pituitary-adrenal axis differentially affects the anti-mycobacterial activity of macrophages from BCG-resistant and susceptible mice. J Neuroimmunol. 1994;53:181–187. doi: 10.1016/0165-5728(94)90028-0. [DOI] [PubMed] [Google Scholar]
- 155.Epel E, Blackburn EH, Lin J, Dhabhar FS, Adler NE, Morrow JD, Cawthon RM. Accelerated telomere shortening in response to life stress. PNAS. 2004;101:17312–17315. doi: 10.1073/pnas.0407162101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Epel ES, Merkin SS, Cawthon R, Blackburn EH, Adler NE, Pletcher MJ, Seeman TE. The rate of leukocyte telomere shortening predicts mortality from cardiovascular disease in elderly men. Aging (Albany NY) 2009;1:81–8. doi: 10.18632/aging.100007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Blackburn EH, Epel ES. Telomeres and adversity: Too toxic to ignore. Nature. 2012;490:169–71. doi: 10.1038/490169a. [DOI] [PubMed] [Google Scholar]
- 158.Cohen S, Miller GE, Rabin BS. Psychological stress and antibody response to immunization: a critical review of the human literature. Psychosom Med. 2001;63:7–18. doi: 10.1097/00006842-200101000-00002. [DOI] [PubMed] [Google Scholar]
- 159.Glaser R, Rabin B, Chesney M, Cohen S, Natelson B. Stress-induced immunomodulation: implications for infectious diseases? [see comments] JAMA. 1999;281:2268–70. doi: 10.1001/jama.281.24.2268. [DOI] [PubMed] [Google Scholar]
- 160.Marucha PT, Kiecolt-Glaser JK, Favagehi M. Mucosal wound healing is impaired by examination stress. Psychosomatic Medicine. 1998;60:362–365. doi: 10.1097/00006842-199805000-00025. [DOI] [PubMed] [Google Scholar]
- 161.Mercado AM, Padgett DA, Sheridan JF, Marucha PT. Altered kinetics of IL-1 alpha, IL-1 beta, and KGF-1 gene expression in early wounds of restrained mice. Brain Behav Immun. 2002;16:150–62. doi: 10.1006/brbi.2001.0623. [DOI] [PubMed] [Google Scholar]
- 162.Sheridan JF, Padgett DA, Avitsur R, Marucha PT. Experimental models of stress and wound healing. World J Surg. 2004;28:327–30. doi: 10.1007/s00268-003-7404-y. [DOI] [PubMed] [Google Scholar]
- 163.Christian LM, Graham JE, Padgett DA, Glaser R, Kiecolt-Glaser JK. Stress and wound healing. Neuroimmunomodulation. 2006;13:337–46. doi: 10.1159/000104862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Glaser R, Kiecolt-Glaser JK, Bonneau RH, Malarkey W, Kennedy S, Hughes J. Stress-induced modulation of the immune response to recombinant hepatitis B vaccine. Psychosom Med. 1991;54:22–29. doi: 10.1097/00006842-199201000-00005. [DOI] [PubMed] [Google Scholar]
- 165.Glaser R, Rice J, Speicher CE, Stout JC, Kiecolt-Glaser JK. Stress depresses interferon production by leukocytes concomitant with a decrease in natural killer cell activity. Behav Neurosci. 1986;100:675–678. doi: 10.1037//0735-7044.100.5.675. [DOI] [PubMed] [Google Scholar]
- 166.Kiecolt-Glaser JK, Glaser R. Chronic stress and mortality among older adults [editorial; comment] Jama. 1999;282:2259–60. doi: 10.1001/jama.282.23.2259. [DOI] [PubMed] [Google Scholar]
- 167.Fagundes CP, Glaser R, Johnson SL, Andridge RR, Yang EV, Di Gregorio MP, Chen M, Lambert DR, Jewell SD, Bechtel MA, Hearne DW, Herron JB, Kiecolt-Glaser JK. Basal Cell Carcinoma: Stressful Life Events and the Tumor Environment. Arch Gen Psychiatry. 2012;69:618–26. doi: 10.1001/archgenpsychiatry.2011.1535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Cohen S, Doyle WJ, Skoner DP, Rabin BS, Gwaltney JM., Jr Social ties and susceptibility to the common cold [see comments] Jama. 1997;277:1940–4. [PubMed] [Google Scholar]
- 169.Cohen S, Frank E, Doyle WJ, Skoner DP, Rabin BS, Gwaltney JM., Jr Types of stressors that increase susceptibility to the common cold in healthy adults [see comments] Health Psychology. 1998;17:214–23. doi: 10.1037//0278-6133.17.3.214. [DOI] [PubMed] [Google Scholar]
- 170.Cohen S, Hamrick N, Rodriguez MS, Feldman PJ, Rabin BS, Manuck SB. Reactivity and vulnerability to stress-associated risk for upper respiratory illness. Psychosom Med. 2002;64:302–10. doi: 10.1097/00006842-200203000-00014. [DOI] [PubMed] [Google Scholar]
- 171.Cohen S, Line S, Manuck SB, Rabin BS, Heise ER, Kaplan JR. Chronic social stress, social status, and susceptibility to upper respiratory infections in nonhuman primates [see comments] Psychosomatic Medicine. 1997;59:213–21. doi: 10.1097/00006842-199705000-00001. [DOI] [PubMed] [Google Scholar]
- 172.Rojas IG, Padgett DA, Sheridan JF, Marucha PT. Stress-induced susceptibility to bacterial infection during cutaneous wound healing. Brain Behav Immun. 2002;16:74–84. doi: 10.1006/brbi.2000.0619. [DOI] [PubMed] [Google Scholar]
- 173.Rozlog LA, Kiecolt-Glaser JK, Marucha PT, Sheridan JF, Glaser R. Stress and immunity: implications for viral disease and wound healing. J Periodontol. 1999;70:786–92. doi: 10.1902/jop.1999.70.7.786. [DOI] [PubMed] [Google Scholar]
- 174.Antoni MH, Lutgendorf SK, Cole SW, Dhabhar FS, Sephton SE, McDonald PG, Stefanek M, Sood AK. The influence of bio-behavioral factors on tumour biology: pathways and mechanisms. Nature Reviews Cancer. 2006;6:240–248. doi: 10.1038/nrc1820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Chida Y, Hamer M, Wardle J, Steptoe A. Do stress-related psychosocial factors contribute to cancer incidence and survival? Nat Clin Pract Oncol. 2008;5:466–75. doi: 10.1038/ncponc1134. [DOI] [PubMed] [Google Scholar]
- 176.Sood AK, Lutgendorf SK. Stress influences on anoikis. Cancer prevention research (Philadelphia, Pa) 2011;4:481–5. doi: 10.1158/1940-6207.CAPR-10-0358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Ben-Eliyahu S, Yirmiya R, Liebeskind JC, Taylor AN, Gale RP. Stress increases metastatic spread of a mammary tumor in rats: evidence for mediation by the immune system. Brain, Behav, Immun. 1991;5:193–205. doi: 10.1016/0889-1591(91)90016-4. [DOI] [PubMed] [Google Scholar]
- 178.Thaker PH, Han LY, Kamat AA, Arevalo JM, Takahashi R, Lu C, Jennings NB, Armaiz-Pena G, Bankson JA, Ravoori M, Merritt WM, Lin YG, Mangala LS, Kim TJ, Coleman RL, Landen CN, Li Y, Felix E, Sanguino AM, Newman RA, Lloyd M, Gershenson DM, Kundra V, Lopez-Berestein G, Lutgendorf SK, Cole SW, Sood AK. Chronic stress promotes tumor growth and angiogenesis in a mouse model of ovarian carcinoma. Nat Med. 2006;12:939–44. doi: 10.1038/nm1447. [DOI] [PubMed] [Google Scholar]
- 179.Levi B, Benish M, Goldfarb Y, Sorski L, Melamed R, Rosenne E, Ben-Eliyahu S. Continuous stress disrupts immunostimulatory effects of IL-12. Brain Behav Immun. 2011;25:727–35. doi: 10.1016/j.bbi.2011.01.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Goldfarb Y, Sorski L, Benish M, Levi B, Melamed R, Ben-Eliyahu S. Improving postoperative immune status and resistance to cancer metastasis: a combined perioperative approach of immunostimulation and prevention of excessive surgical stress responses. Ann Surg. 2011;253:798–810. doi: 10.1097/SLA.0b013e318211d7b5. [DOI] [PubMed] [Google Scholar]
- 181.Dhabhar FS, Saul AN, Holmes TH, Daugherty C, Neri E, Tillie JM, Kusewitt DF, Oberyszyn TM. High anxiety is associated with higher chronic stress burden, lower protective immunity, and increased cancer progression. PLoS ONE. 2012;7 doi: 10.1371/journal.pone.0033069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Levine S, Saltzman A. Nonspecific stress prevents relapses of Experimental Allergic Encephalomyelitis in rats. Brain, Behavior, and Immunity. 1987;1:336–341. doi: 10.1016/0889-1591(87)90036-5. [DOI] [PubMed] [Google Scholar]
- 183.Rogers MP, Trentham DE, McCune WJ, Ginsberg BI, Rennke HG, Reich P, David JR. Effect of psychological stress on the induction of arthritis in rats. Arthritis & Rheumatism. 1980;23:1337–1342. doi: 10.1002/art.1780231202. [DOI] [PubMed] [Google Scholar]
- 184.Griffin AC, Warren DL, Wolny AC, Whitacre CC. Suppression of experimental autoimmune encephalomyelitis by restraint stress. J Neuroimmunology. 1993;44:103–116. doi: 10.1016/0165-5728(93)90273-2. [DOI] [PubMed] [Google Scholar]
- 185.Stefanski V, Hemschemeier SK, Schunke K, Hahnel A, Wolff C, Straub RH. Differential effect of severe and moderate social stress on blood immune and endocrine measures and susceptibility to collagen type II arthritis in male rats. Brain Behav Immun. 2013;29:156–65. doi: 10.1016/j.bbi.2012.12.019. [DOI] [PubMed] [Google Scholar]
- 186.Besedovsky HO, del Rey A. The cytokine-HPA axis feed-back circuit. Z Rheumatol. 2000;59(Suppl 2):II/26–30. doi: 10.1007/s003930070014. [DOI] [PubMed] [Google Scholar]
- 187.del Rey A, Besedovsky HO. The cytokine-HPA axis circuit contributes to prevent or moderate autoimmune processes. Z Rheumatol. 2000;59:II/31–5. doi: 10.1007/s003930070015. [DOI] [PubMed] [Google Scholar]
- 188.Straub RH, Kalden JR. Stress of different types increases the proinflammatory load in rheumatoid arthritis. Arthritis Res Ther. 2009;11:114. doi: 10.1186/ar2712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Del Rey A, Wolff C, Wildmann J, Randolf A, Straub RH, Besedovsky HO. When immune-neuro-endocrine interactions are disrupted: experimentally induced arthritis as an example. Neuroimmunomodulation. 2010;17:165–8. doi: 10.1159/000258714. [DOI] [PubMed] [Google Scholar]
- 190.Sternberg EM. Neuroendocrine regulation of autoimmune/inflammatory disease. J Endocrinol. 2001;169:429–35. doi: 10.1677/joe.0.1690429. [DOI] [PubMed] [Google Scholar]
- 191.Hackney AC. Exercise as a stressor to the human neuroendocrine system. Medicina (Kaunas) 2006;42:788–97. [PubMed] [Google Scholar]
- 192.Pedersen BK, Hoffman-Goetz L. Exercise and the immune system: regulation, integration, and adaptation. Physiol Rev. 2000;80:1055–81. doi: 10.1152/physrev.2000.80.3.1055. [DOI] [PubMed] [Google Scholar]
- 193.Pedersen BK. Special feature for the Olympics: effects of exercise on the immune system: exercise and cytokines. Immunol Cell Biol. 2000;78:532–5. doi: 10.1111/j.1440-1711.2000.t01-11-.x. [DOI] [PubMed] [Google Scholar]
- 194.Walsh NP, Gleeson M, Pyne DB, Nieman DC, Dhabhar FS, Shephard RJ, Oliver SJ, Bermon S, Kajeniene A. Position statement. Part two: Maintaining immune health. Exerc Immunol Rev. 2011;17:64–103. [PubMed] [Google Scholar]
- 195.Winzer BM, Whiteman DC, Reeves MM, Paratz JD. Physical activity and cancer prevention: a systematic review of clinical trials. Cancer Causes Control. 2011;22:811–26. doi: 10.1007/s10552-011-9761-4. [DOI] [PubMed] [Google Scholar]
- 196.Friedenreich CM. The role of physical activity in breast cancer etiology. Semin Oncol. 2010;37:297–302. doi: 10.1053/j.seminoncol.2010.05.008. [DOI] [PubMed] [Google Scholar]
- 197.Friedenreich CM, Neilson HK, Lynch BM. State of the epidemiological evidence on physical activity and cancer prevention. European Journal of Cancer. 2010;46:2593–2604. doi: 10.1016/j.ejca.2010.07.028. [DOI] [PubMed] [Google Scholar]
- 198.Clague J, Bernstein L. Physical activity and cancer. Curr Oncol Rep. 2012;14:550–8. doi: 10.1007/s11912-012-0265-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Gleeson M. Immune function in sport and exercise. J Appl Physiol. 2007;103:693–9. doi: 10.1152/japplphysiol.00008.2007. [DOI] [PubMed] [Google Scholar]
- 200.Walsh NP, Whitham M. Exercising in environmental extremes: a greater threat to immune function? Sports Med. 2006;36:941–76. doi: 10.2165/00007256-200636110-00003. [DOI] [PubMed] [Google Scholar]
- 201.Woods JA, Vieira VJ, Keylock KT. Exercise, inflammation, and innate immunity. Immunol Allergy Clin North Am. 2009;29:381–93. doi: 10.1016/j.iac.2009.02.011. [DOI] [PubMed] [Google Scholar]
- 202.Phillips AC, Burns VE, Lord JM. Stress and exercise: Getting the balance right for aging immunity. Exerc Sport Sci Rev. 2007;35:35–9. doi: 10.1097/jes.0b013e31802d7008. [DOI] [PubMed] [Google Scholar]
- 203.Woods JA, Lowder TW, Keylock KT. Can exercise training improve immune function in the aged? Ann N Y Acad Sci. 2002;959:117–27. doi: 10.1111/j.1749-6632.2002.tb02088.x. [DOI] [PubMed] [Google Scholar]
- 204.Fondell E, Lagerros YT, Sundberg CJ, Lekander M, Balter O, Rothman KJ, Balter K. Physical activity, stress, and self-reported upper respiratory tract infection. Med Sci Sports Exerc. 2011;43:272–9. doi: 10.1249/MSS.0b013e3181edf108. [DOI] [PubMed] [Google Scholar]
- 205.Dhabhar FS. Psychological stress and immunoprotection versus immunopathology in the skin. Clin Dermatol. 2013;31:18–30. doi: 10.1016/j.clindermatol.2011.11.003. [DOI] [PubMed] [Google Scholar]
- 206.Dhabhar FS. Enhancing versus suppressive effects of stress on immune function -- The role of leukocyte trafficking and the kinetic and concentration conditions of exposure to stress hormones (New Investigator Award) Brain, Behavior, Immunity. 2002;16:785–798. doi: 10.1016/s0889-1591(02)00036-3. [DOI] [PubMed] [Google Scholar]
- 207.Edwards KM, Burns VE, Reynolds T, Carroll D, Drayson M, Ring C. Acute stress exposure prior to influenza vaccination enhances antibody response in women. Brain Behav Immun. 2006;20:159–68. doi: 10.1016/j.bbi.2005.07.001. [DOI] [PubMed] [Google Scholar]
- 208.Edwards KM, Burns VE, Adkins AE, Carroll D, Drayson M, Ring C. Meningococcal A vaccination response is enhanced by acute stress in men. Psychosom Med. 2008;70:147–51. doi: 10.1097/PSY.0b013e318164232e. [DOI] [PubMed] [Google Scholar]
- 209.Edwards KM, Burns VE, Carroll D, Drayson M, Ring C. The acute stress-induced immunoenhancement hypothesis. Exerc Sport Sci Rev. 2007;35:150–5. doi: 10.1097/JES.0b013e3180a031bd. [DOI] [PubMed] [Google Scholar]
- 210.Kleyn CE, Schneider L, Saraceno R, Mantovani C, Richards HL, Fortune DG, Cumberbatch M, Dearman RJ, Terenghi G, Kimber I, Griffiths CE. The effects of acute social stress on epidermal Langerhans’ cell frequency and expression of cutaneous neuropeptides. J Invest Dermatol. 2008;128:1273–9. doi: 10.1038/sj.jid.5701144. [DOI] [PubMed] [Google Scholar]
- 211.Griffiths CE, Dearman RJ, Cumberbatch M, Kimber I. Cytokines and Langerhans cell mobilisation in mouse and man. Cytokine. 2005;32:67–70. doi: 10.1016/j.cyto.2005.07.011. [DOI] [PubMed] [Google Scholar]
- 212.Chida Y, Hamer M, Steptoe A. A bidirectional relationship between psychosocial factors and atopic disorders: a systematic review and meta-analysis. Psychosom Med. 2008;70:102–16. doi: 10.1097/PSY.0b013e31815c1b71. [DOI] [PubMed] [Google Scholar]
- 213.Arndt J, Smith N, Tausk F. Stress and atopic dermatitis. Curr Allergy Asthma Rep. 2008;8:312–7. doi: 10.1007/s11882-008-0050-6. [DOI] [PubMed] [Google Scholar]
- 214.Black PH, Garbutt LD. Stress, inflammation and cardiovascular disease. J Psychosom Res. 2002;52:1–23. doi: 10.1016/s0022-3999(01)00302-6. [DOI] [PubMed] [Google Scholar]
- 215.Belkic KL, Landsbergis PA, Schnall PL, Baker D. Is job strain a major source of cardiovascular disease risk? Scand J Work Environ Health. 2004;30:85–128. doi: 10.5271/sjweh.769. [DOI] [PubMed] [Google Scholar]
- 216.Hildebrand HC, Epstein J, Larjava H. The influence of psychological stress on periodontal disease. J West Soc Periodontol Periodontal Abstr. 2000;48:69–77. [PubMed] [Google Scholar]
- 217.Wright RJ, Rodriguez M, Cohen S. Review of psychosocial stress and asthma: an integrated biopsychosocial approach. Thorax. 1998;53:1066–74. doi: 10.1136/thx.53.12.1066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Chen E, Miller GE. Stress and inflammation in exacerbations of asthma. Brain Behav Immun. 2007;21:993–9. doi: 10.1016/j.bbi.2007.03.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Al’Abadie MS, Kent GG, Gawkrodger DJ. The relationship between stress and the onset and exacerbation of psoriasis and other skin conditions. Brit J Dermatol. 1994;130:199–203. doi: 10.1111/j.1365-2133.1994.tb02900.x. [DOI] [PubMed] [Google Scholar]
- 220.Fortune DG, Richards HL, Griffiths CE. Psychologic factors in psoriasis: consequences, mechanisms, and interventions. Dermatol Clin. 2005;23:681–94. doi: 10.1016/j.det.2005.05.022. [DOI] [PubMed] [Google Scholar]
- 221.Straub RH, Dhabhar FS, Bijlsma JW, Cutolo M. How psychological stress via hormones and nerve fibers may exacerbate rheumatoid arthritis. Arthritis Rheum. 2005;52:16–26. doi: 10.1002/art.20747. [DOI] [PubMed] [Google Scholar]
- 222.Artemiadis AK, Anagnostouli MC, Alexopoulos EC. Stress as a Risk Factor for Multiple Sclerosis Onset or Relapse: A Systematic Review. Neuroepidemiology. 2011;36:109–120. doi: 10.1159/000323953. [DOI] [PubMed] [Google Scholar]
- 223.Edwards KM, Burns VE, Allen LM, McPhee JS, Bosch JA, Carroll D, Drayson M, Ring C. Eccentric exercise as an adjuvant to influenza vaccination in humans. Brain Behav Immun. 2007;21:209–17. doi: 10.1016/j.bbi.2006.04.158. [DOI] [PubMed] [Google Scholar]
- 224.Jamieson JP, Mendes WB, Blackstock E, Schmader T. Turning the knots in your stomach into bows: Reappraising arousal improves performance on the GRE. J Exp Soc Psychol. 2010;46:208–212. doi: 10.1016/j.jesp.2009.08.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.El Zein M, Wyart V, Grezes J. Anxiety dissociates the adaptive functions of sensory and motor response enhancements to social threats. Elife. 2015;4 doi: 10.7554/eLife.10274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Brooks AW. Get excited: reappraising pre-performance anxiety as excitement. J Exp Psychol Gen. 2014;143:1144–58. doi: 10.1037/a0035325. [DOI] [PubMed] [Google Scholar]
- 227.Jamieson JP, Nock MK, Mendes WB. Mind over matter: reappraising arousal improves cardiovascular and cognitive responses to stress. J Exp Psychol Gen. 2012;141:417–22. doi: 10.1037/a0025719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.McIntyre CK, Roozendaal B. Adrenal Stress Hormones and Enhanced Memory for Emotionally Arousing Experiences. In: Bermudez-Rattoni F, editor. Neural Plasticity and Memory: From Genes to Brain Imaging. Boca Raton (FL): 2007. [PubMed] [Google Scholar]
- 229.Marin MF, Pilgrim K, Lupien SJ. Modulatory effects of stress on reactivated emotional memories. Psychoneuroendocrinology. 2010;35:1388–96. doi: 10.1016/j.psyneuen.2010.04.002. [DOI] [PubMed] [Google Scholar]
- 230.Segal SK, Cahill L. Endogenous noradrenergic activation and memory for emotional material in men and women. Psychoneuroendocrinology. 2009;34:1263–71. doi: 10.1016/j.psyneuen.2009.04.020. [DOI] [PubMed] [Google Scholar]
- 231.Segal SK, Cotman CW, Cahill LF. Exercise-induced noradrenergic activation enhances memory consolidation in both normal aging and patients with amnestic mild cognitive impairment. J Alzheimers Dis. 2012;32:1011–8. doi: 10.3233/JAD-2012-121078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Segal SK, Simon R, McFarlin S, Alkire M, Desai A, Cahill LF. Glucocorticoids interact with noradrenergic activation at encoding to enhance long-term memory for emotional material in women. Neuroscience. 2014;277:267–72. doi: 10.1016/j.neuroscience.2014.06.059. [DOI] [PubMed] [Google Scholar]
- 233.van Stegeren AH, Wolf OT, Everaerd W, Scheltens P, Barkhof F, Rombouts SA. Endogenous cortisol level interacts with noradrenergic activation in the human amygdala. Neurobiol Learn Mem. 2007;87:57–66. doi: 10.1016/j.nlm.2006.05.008. [DOI] [PubMed] [Google Scholar]
- 234.Taverniers J, Van Ruysseveldt J, Smeets T, von Grumbkow J. High-intensity stress elicits robust cortisol increases, and impairs working memory and visuo-spatial declarative memory in Special Forces candidates: A field experiment. Stress. 2010;13:323–33. doi: 10.3109/10253891003642394. [DOI] [PubMed] [Google Scholar]
- 235.Conrad CD, Lupien SJ, McEwen BS. Support for a bimodal role for type II adrenal steroid receptors in spatial memory. Neurobiol Learn Mem. 1999;72:39–46. doi: 10.1006/nlme.1998.3898. [DOI] [PubMed] [Google Scholar]
- 236.Lupien SJ, Wilkinson CW, Briere S, Menard C, Ng Ying Kin NM, Nair NP. The modulatory effects of corticosteroids on cognition: studies in young human populations. Psychoneuroendocrinology. 2002;27:401–16. doi: 10.1016/s0306-4530(01)00061-0. [DOI] [PubMed] [Google Scholar]
- 237.Crewther BT, Thomas AG, Stewart-Williams S, Kilduff LP, Cook CJ. Is salivary cortisol moderating the relationship between salivary testosterone and hand-grip strength in healthy men? Eur J Sport Sci. 2017;17:188–194. doi: 10.1080/17461391.2016.1220628. [DOI] [PubMed] [Google Scholar]
- 238.Gullestad L, Hallen J, Medbo JI, Gronnerod O, Holme I, Sejersted OM. The effect of acute vs chronic treatment with beta-adrenoceptor blockade on exercise performance, haemodynamic and metabolic parameters in healthy men and women. Br J Clin Pharmacol. 1996;41:57–67. doi: 10.1111/j.1365-2125.1996.tb00159.x. [DOI] [PubMed] [Google Scholar]
- 239.Kim KM, Kawada T, Ishihara K, Inoue K, Fushiki T. Increase in swimming endurance capacity of mice by capsaicin-induced adrenal catecholamine secretion. Biosci Biotechnol Biochem. 1997;61:1718–23. doi: 10.1271/bbb.61.1718. [DOI] [PubMed] [Google Scholar]
- 240.von Dawans B, Fischbacher U, Kirschbaum C, Fehr E, Heinrichs M. The social dimension of stress reactivity: acute stress increases prosocial behavior in humans. Psychol Sci. 2012;23:651–60. doi: 10.1177/0956797611431576. [DOI] [PubMed] [Google Scholar]
- 241.Amkraut AA, Solomon CF, Kraemer HC. Stress, early experience and adjuvant-induced arthritis in the rat. Psychosom Med. 1971;33:203–214. doi: 10.1097/00006842-197105000-00002. [DOI] [PubMed] [Google Scholar]
- 242.Ackerman KD, Heyman R, Rabin BS, Anderson BP, Houck PR, Frank E, Baum A. Stressful life events precede exacerbations of multiple sclerosis. Psychosom Med. 2002;64:916–920. doi: 10.1097/01.psy.0000038941.33335.40. [DOI] [PubMed] [Google Scholar]
- 243.Garg A, Chren MM, Sands LP, Matsui MS, Marenus KD, Feingold KR, Elias PM. Psychological stress perturbs epidermal permeability barrier homeostasis: implications for the pathogenesis of stress-associated skin disorders. Arch Dermatol. 2001;137:53–9. doi: 10.1001/archderm.137.1.53. [DOI] [PubMed] [Google Scholar]
- 244.Cohen S, Tyrrell DAJ, Smith AP. Psychological stress and susceptibility to the common cold. New Engl J Med. 1991;325:606–612. doi: 10.1056/NEJM199108293250903. [DOI] [PubMed] [Google Scholar]
- 245.Sandi C. Memory Impairments Associated with Stress and Aging. In: Bermudez-Rattoni F, editor. Neural Plasticity and Memory: From Genes to Brain Imaging. Boca Raton (FL): 2007. [PubMed] [Google Scholar]
- 246.Dhabhar FS. Harnessing stress on the path to peace. TEDx Hayward. 2013 https://www.youtube.com/watch?v=u2QFJbE1EBs.
- 247.Esch T, Stefano GB. Endogenous reward mechanisms and their importance in stress reduction, exercise and the brain. Arch Med Sci. 2010;6:447–55. doi: 10.5114/aoms.2010.14269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Corah NL, Bissell GD, Illig SJ. Effect of perceived control on stress reduction in adult dental patients. J Dent Res. 1978;57:74–6. doi: 10.1177/00220345780570012601. [DOI] [PubMed] [Google Scholar]
- 249.Astin JA. Stress reduction through mindfulness meditation. Effects on psychological symptomatology, sense of control, and spiritual experiences. Psychother Psychosom. 1997;66:97–106. doi: 10.1159/000289116. [DOI] [PubMed] [Google Scholar]
- 250.Griffin JM, Fuhrer R, Stansfeld SA, Marmot M. The importance of low control at work and home on depression and anxiety: do these effects vary by gender and social class? Soc Sci Med. 2002;54:783–98. doi: 10.1016/s0277-9536(01)00109-5. [DOI] [PubMed] [Google Scholar]
- 251.Steptoe A, Kunz-Ebrecht S, Owen N, Feldman PJ, Rumley A, Lowe GD, Marmot M. Influence of socioeconomic status and job control on plasma fibrinogen responses to acute mental stress. Psychosom Med. 2003;65:137–44. doi: 10.1097/01.psy.0000039755.23250.a7. [DOI] [PubMed] [Google Scholar]
- 252.Bosma H, Stansfeld SA, Marmot MG. Job control, personal characteristics, and heart disease. J Occup Health Psychol. 1998;3:402–9. doi: 10.1037//1076-8998.3.4.402. [DOI] [PubMed] [Google Scholar]
- 253.Garland EL, Hanley AW, Goldin PR, Gross JJ. Testing the mindfulness-to-meaning theory: Evidence for mindful positive emotion regulation from a reanalysis of longitudinal data. PLoS One. 2017;12:e0187727. doi: 10.1371/journal.pone.0187727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Smith SA. Mindfulness-based stress reduction: an intervention to enhance the effectiveness of nurses’ coping with work-related stress. Int J Nurs Knowl. 2014;25:119–30. doi: 10.1111/2047-3095.12025. [DOI] [PubMed] [Google Scholar]
- 255.Nejati S, Zahiroddin A, Afrookhteh G, Rahmani S, Hoveida S. Effect of Group Mindfulness-Based Stress-Reduction Program and Conscious Yoga on Lifestyle, Coping Strategies, and Systolic and Diastolic Blood Pressures in Patients with Hypertension. J Tehran Heart Cent. 2015;10:140–8. [PMC free article] [PubMed] [Google Scholar]
- 256.Cacioppo JT, Cacioppo S, Capitanio JP, Cole SW. The neuroendocrinology of social isolation. Annu Rev Psychol. 2015;66:733–67. doi: 10.1146/annurev-psych-010814-015240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257.Dharmawardene M, Givens J, Wachholtz A, Makowski S, Tjia J. A systematic review and meta-analysis of meditative interventions for informal caregivers and health professionals. BMJ Support Palliat Care. 2016;6:160–9. doi: 10.1136/bmjspcare-2014-000819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258.Sharma M, Rush SE. Mindfulness-based stress reduction as a stress management intervention for healthy individuals: a systematic review. J Evid Based Complementary Altern Med. 2014;19:271–86. doi: 10.1177/2156587214543143. [DOI] [PubMed] [Google Scholar]
- 259.Riley KE, Park CL. How does yoga reduce stress? A systematic review of mechanisms of change and guide to future inquiry. Health Psychol Rev. 2015;9:379–96. doi: 10.1080/17437199.2014.981778. [DOI] [PubMed] [Google Scholar]
- 260.Williams DR, Sternthal MJ. Spirituality, religion and health: evidence and research directions. Med J Aust. 2007;186:S47–50. doi: 10.5694/j.1326-5377.2007.tb01040.x. [DOI] [PubMed] [Google Scholar]
- 261.Sandmire DA, Rankin NE, Gorham SR, Eggleston DT, French CA, Lodge EE, Kuns GC, Grimm DR. Psychological and autonomic effects of art making in college-aged students. Anxiety Stress Coping. 2016;29:561–9. doi: 10.1080/10615806.2015.1076798. [DOI] [PubMed] [Google Scholar]
- 262.Martin L, Oepen R, Bauer K, Nottensteiner A, Mergheim K, Gruber H, Koch SC. Creative Arts Interventions for Stress Management and Prevention-A Systematic Review. Behav Sci (Basel) 2018;8 doi: 10.3390/bs8020028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263.Bjornstad S, Patil GG, Raanaas RK. Nature contact and organizational support during office working hours: Benefits relating to stress reduction, subjective health complaints, and sick leave. Work. 2015;53:9–20. doi: 10.3233/WOR-152211. [DOI] [PubMed] [Google Scholar]