

Abstract
In the field of immunology, there is an increasing interest in cellular energy metabolism and its outcome on immune cell effector function. Activation of immune cells leads to rapid metabolic changes that are central to cellular biology in order to support the effector responses. Therefore, the need for user-friendly and dependable assay technologies to address metabolic regulation and nutrient utilization in immune cells is an important need in this field. Redox-dye reduction-based Phenotype MicroArray (PM) assays, which measure NADH reduction as a readout, developed by Biolog Inc., provide a wide screening of metabolites both in bacteria and mammalian cells. In this study, we delineate a detailed protocol of a customized Biolog assay for investigation of a specific metabolic pathway of interest. The option to be able to easily customize this technology offers researchers with a convenient assay platform to methodically examine specific nutrient substrates or metabolic pathways of interest in a rapid and cost effective manner.
Keywords: Metabolism, Dendritic Cells, Biolog, Phenotypic MicroArray, Glucose, Glycogen
INTRODUCTION
There is a growing appreciation in the field of immunology that immune cell activation is accompanied by dramatic shifts in cellular metabolism, often highlighted by increased cellular uptake of glucose (1–9). Cellular utilization of specific metabolic pathways has traditionally been assessed by radioisotope labeling of metabolites and/or indirect measurements of enzymatic activities. While the techniques like carbon isotopic labeling, such as LC-MS approaches, provide reliable snapshots of cellular metabolic fluxes, they demand specialized expertise from the execution of experiments to data analysis. In this work, we outline an experimental method using Biolog Inc. Phenotype MicroArray (PM) technology to perform simple and rapid screenings of cellular energy-producing metabolic pathways that provides fast, cost effective, and clear results.
The Biolog PM assay technology was originally developed for microbial metabolic screening, based on assaying the ability of microorganisms to metabolize distinct nutrient substrates. This assay platform was subsequently developed for a variety of eukaryotic cell applications including assays for evaluating cancer cell drug targets, metabolic disorders, cancer cell metabolism, drug toxicity testing, and general metabolic pathway analyses. For these assays, cells are plated in a 96-well format where each well is pre-loaded with a single distinct metabolic substrate. After allowing cells to equilibrate to their nutrient environment and metabolism to occur, cells are then co-incubated with a colorimetric dye that is reduced in the presence of NADH. If cells are capable of metabolizing a substrate, NADH will be produced resulting in proportional reduction of the dye. This assay is based on the premise that the amount of NADH produced directly correlates the cell’s ability to utilize a particular substrate as an energy source (10). Although it is similar to the principle of MTT assays, the Biolog tetrazolium redox dye mix specifically optimized to measure cellular NADH production. It is also water soluble and readily adapted for metabolic rate measurements. This eliminates the solubility issues and the precipitation of formazan, thereby reducing the high background noise that is a typical obstacle for data interpretation in standard MTT methodologies (10). The intensity of color formation that results from dye reduction directly correlates with the amount of cellular NADH production driven by metabolism of a given substrate. In addition, cells can be assayed over time to provide kinetic analysis of substrate usage. Figure 1 provides a simple illustration of NADH production during glycolytic and mitochondrial metabolism which is read as dye reduction of the Biolog Inc. Dye Mix in these assays.
Figure 1.
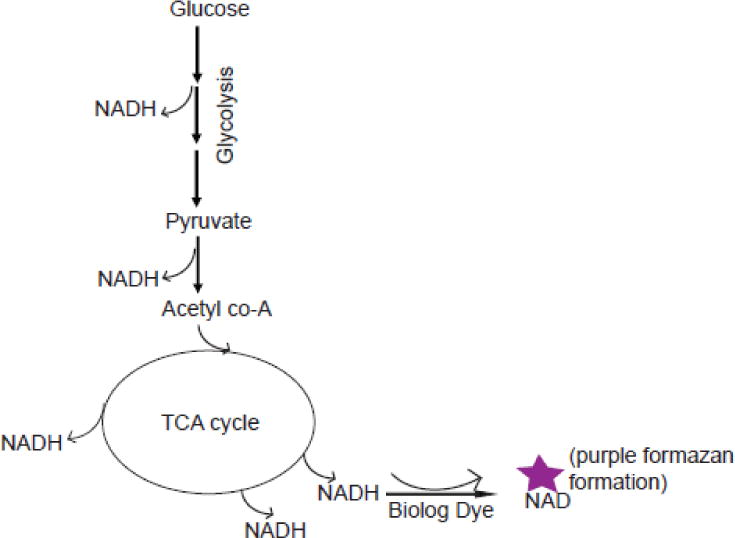
Illustration of NADH production (with dye reduction) from glucose metabolism
One of the major advantages of this assay, from the perspective of larger-scale screening assays, is that it can be accomplished with relatively few cells in a total volume of 100 μL (see procedure for details) in each well of a 96-microwell plate. Cells are originally plated in a minimal nutrient media (MC-0) for overnight incubation. During this incubation, the cells will utilize residual glucose provided in MC-0 before they adapt and switch to the saturating single nutrient source in each well of a PM plate. Cellular utilization of metabolic substrates and their kinetics can be simply measured by colorimetric readings of purple formazan formation from Biolog redox dye reduction. The advantage of this technology is that it not only allows an individual to perform a broad screening of multiple metabolites by using pre-designed PM plates, but also provides the flexibility to customize the assay for a thorough investigation of a single metabolite/metabolic pathway of interest, specifically tailored to individual’s research needs. Below, we delineate an illustrative example of how researchers can adapt this technology to screen for the efficacy of metabolic-inhibiting drugs for a specific research question.
In a recently published research article, we originally identified that dendritic cells (DCs) of the immune system can use glycogen as an energy substrate from screening a wide number of carbon substrates using Biolog inc., PM assay plates (11). In this screen, we identified that DCs can utilize not only single units of glucose but also various lengths of glucose polymers including the long-chain glucose polymer known as glycogen (11). To systematically examine glycogen usage by DCs, we customized this technology to allow us to specifically screen for the efficacy of drugs that targeted glycogen metabolism in our assay system (Figure 2). In this case, we built our custom assays using our substrates of interest, glycogen and glucose (Table 1). Because of the extreme flexibility of the assay, we were also able to use this approach to simultaneously examine cellular nutrient utilization in both resting and activated DCs. With this customized approach, we have successfully identified a metabolite of our interest, glycogen, and validated the specificity of multiple inhibitors of glycogen metabolism in DCs as well as other immune cell types at different activation stages. Because of a growing interest in the metabolic pathway utilization of immune cells in response to stimuli, we believe that this assay platform will be attractive to researchers in the field based on the flexibility to customize this technology to query a wide range of metabolic parameters in a quick and cost-effective way.
Figure 2.
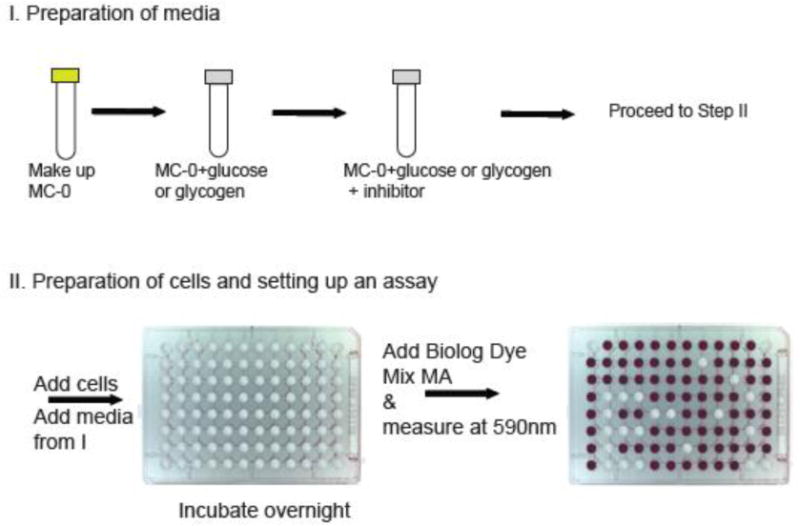
Workflow for a customized Biolog assay set up (96-well plate picture courtesy of Biolog Inc.)
Table 1.
An example of customized plate layout for validating DC utilization of glucose and glycogen metabolic pathways with Biolog plates and the Biolog assay platform with Bone-marrow-derived Dendritic Cells (BMDCs) and Glycogen Phosphorylase (PYG) inhibitor.
| MC-0 (no cells) | MC-0 only (with cells) | glucose | Glycogen | Glucose+ PYG inhibitor | Glycogen + PYG inhibitor |
|---|---|---|---|---|---|
| BMDCs | BMDCs | BMDCs | BMDCs | BMDCs | |
| BMDCs | BMDCs | BMDCs | BMDCs | BMDCs | |
| BMDCs | BMDCs | BMDCs | BMDCs | BMDCs |
MATERIALS
*a96-well micro plates (Biolog Inc.); Other 96-well flat bottom plates can be substituted. Please see discussion for more information.
*bBiolog dye mix MA (6x) (Biolog Inc.) (for some cell types dye mix MB may work better)
*cIFM-1 (Biolog Inc.); This can be substituted with phenol red free, glucose and glutamine free RPMI.
Glutamine 2mM
5% Fetal Bovine Serum (FBS) (or dialyzed FBS if limiting exogenous glucose is desired)
Bovine pancreatic glycogen, 5mg/mL concentration (Sigma aldrich)
CP91149 (CP) glycogen phosphorylase inhibitor I (Selleckham)
DAB, 1,4-Dideoxy-1,4-imino-D-arabinitol hydrochloride, glycogen phosphorylase inhibitor II (Santa Cruz)
Glucose 10mM
Multi-channel pipette and reservoir (optional)
Plate reader at 590nm wavelength
*These reagents/supplies are obtained from Biolog Inc.,; however, options and limitations to substitute *a and *c with alternative sources are mentioned in discussion.
PROCEDURE
Murine Bone-marrow -derived Dendritic Cells (BMDCs) are used as an example in this protocol
I. Reagent preparation
-
1. Assay Media (MC-0)
MC-0 assay media supplemented with 5% FBS is prepared with the following reagents: for 25mL of MC-0 medium, add 1.25mL of FBS, 500μL of Pen Strep, and 50μL of 200mM stock of glutamine into a 23.5mL of IF-M1 medium. (NOTE 1)
Glycogen containing MC-0 medium is prepared from the MC-0 medium from 1a to get a 10mg/mL (2x concentration). Glucose containing MC-0 medium with a final concentration 10mM (2x concentration) is prepared similarly by using the MC-0 medium from 1a.
For dose titrations of pharmacological inhibitors, for example glycogen phosphorylase inhibitor CP and DAB, inhibitor dilutions are prepared in glycogen containing MC-0 medium from Ia.
-
Assay Media (IFM-1)
For testing a certain pharmacological inhibitor such as glucose transporter-1 inhibitor (STF31), it is best to use IF-M1, without the addition of serum to limit the contribution of exogenous glucose by serum. In this case, glucose containing IF-M1 medium can be prepared by a certain concentration of glucose in IF-M1 medium with a range of concentration of the inhibitor.
II. Preparation of cells
BMDCs are generated as previously described (1). Briefly, bone marrow cells are flushed from femurs of 6–10 week old C57/Bl6J mice, and cultured in CDCM, which is composed of RPMI, 10% fetal bovine serum, 5mM glutamine, 1mM beta-mercaptoethanol, and 10IU Pen/Strep, supplemented in 20ng/μL of murine GM-CSF. The cells are cultured for 7 days, with a medium change every other day.
Day 7 BMDCs, differentiated in GM-CSF, are washed 2x in PBS to rid of any trace of culture medium (See NOTE 2). After the final wash, the cells are re-suspended in MC-0 basal medium from 1a at 1×10ˆ6/mL concentration. Proceed to III-1 to seed the cells.
If using other cell types such as T or B lymphocytes, the cell number must be adjusted (See Note 2) as mentioned in Table 1. If using human monocyte-derived DCs, cell numbers must also be adjusted due to differences in cell sizes. Furthermore, the redox dye mixes MA and MB should be compared for each cell type to see which works best.
III. Setting up the experiment
BMDCs are seeded at 50,000 cells per well in technical duplicates or triplicates per condition. To do this, add 50μL of 1×10ˆ6/mL BMDCs prepared in II-1 to each well of a 96-microwell Biolog plate (NOTE 3). Then add 50 μL of the prepared medium (at 2x concentration) from I-1 (containing carbon substrates of interest with and without the inhibitors). The total volume in each well will be 100 μL. Note that the basal media alone, either MC-0 or IF-M1, without the cells should be included for background correction.
Incubate the plate at 37°C in 5%CO2 for overnight.
The next morning, add 20 μL of Biolog dye Mix MA (6x) to each well containing 100 μL total medium volume (NOTE 4). This gives a final concentration of tetrazolium at 500 μM. Then, immediately measure the tetrazolium reduction at 590nm using a spectrophotometric plate reader (NOTE 2). This will be a time zero “0”, baseline, reading. Return the plate to the incubator and continue reading the assay at every 15 minute for the first hour. After the first hour, continue the assay with hourly readings for 6 hours or till the color change/the value of the kinetic reading is plateaued. This will provide the kinetics of formazan production that is linear with time (NOTE 5)
The same day analysis (incubation) can also be done using IF-M1 as a base medium without addition of serum (See Note 1 and the Protocol for IF-M1 based assay media). Then allow the cells for an hour or two of incubation time before adding the Biolog dye MA for kinetics readings. Upon dye addition, continue reading the assay using a colorimetric plate reader as mentioned above.
DATA ANALYSIS
- Data are analyzed by normalizing to hour “0” (baseline) readings of each corresponding media:
- Check raw readings to see if the readouts from replicates are approximately the same.
- Average the replicates; the readings of technical replicates should be in a close range to each other.
- For background correction, subtract the readings of MC-0 (no cells) from each group.
- Next, normalize all the kinetic values to the baseline hour “0” by dividing the readings of each time point to the hour “0” readings of corresponding groups.
- Graph the normalized values, with the baseline hour “0” as a starting point to represent relative increase (fold change) over baseline.
An example graph of glycogen utilization in mouse BMDCs with an inhibitor dose titration are shown in Figure 3A. A dose titration for glycogen phosphorylase pharmacological inhibitor, CP, in activated T cells is shown in Figure 3B.
Figure 3.
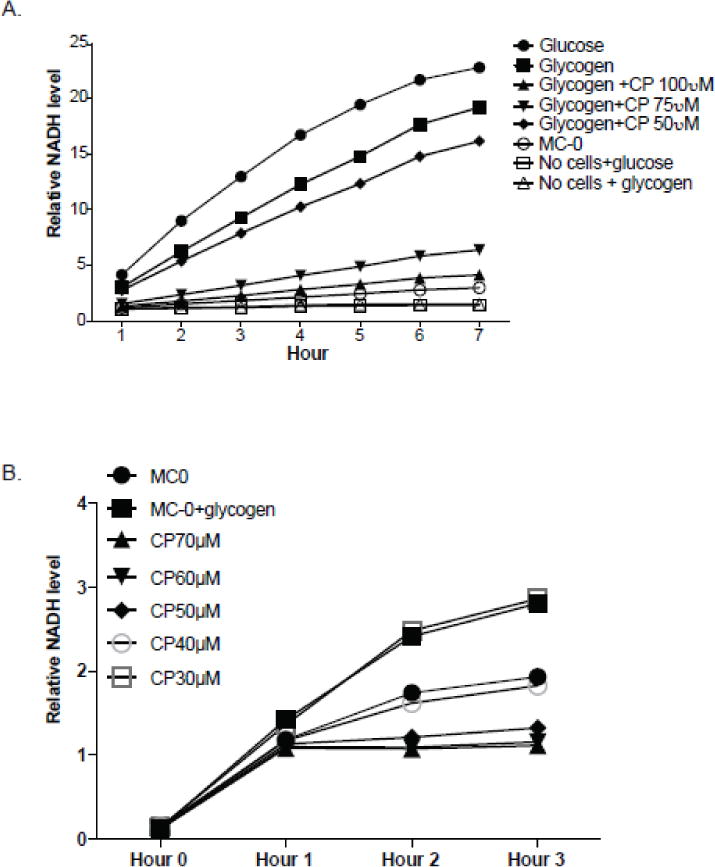
(A) Titration of CP in BMDCs. BMDCs differentiated in GM-CSF were seeded at 50,000 cells/well in a 96-well Biolog microplate plate in a total volume of 100μL corresponding media and incubated overnight. 20μL of Biolog Redox Dye Mix MA was added the next morning. The assay was read hourly from basal time 0 (right after dye addition) up to 7 hours. Data were analyzed by dividing the average readings of each time point by basal time “0” of the corresponding medium. A representative of at least 3independent experiments. (B) Titration of CP in activated T cells. Activated T cells were seeded at 100,000 cells/well in a 96-well Biolog microplate in a total volume of 100μL of corresponding media and incubated overnight. CP were titrated in MC-0+glycogen medium. 20μL of Biolog Redox Dye Mix MA was added to each well the next morning. The assay was read hourly from basal time “0” through hour 3. Data were analyzed by dividing the average readings of each time point by basal time “0” of the corresponding medium. A representative of 3 independent experiments.
RESULTS
To validate the ability of DCs to metabolize glycogen from the PM screening assay in our recently published article (11), we modified the PM assay technology, in which untreated BMDCs were seeded in minimal nutrient medium (MC-0), MC-0 that has glycogen or glucose (control), and a range of dilutions of glycogen phosphorylase inhibitor, CP, in MC-0+glycogen. MC-0 medium alone with no cells was also included for background subtraction. Workflow schematics is shown in Figure 2. As shown in figure 3A, NADH levels from the MC-0+glycogen group are comparable to those of MC-0+glucose. Based on our previously published findings (2, 11), it is not surprising that the cells release such a high level of NADH from MC-0+glucose as glucose is considered a primary fuel source in these cells. To confirm that DCs can generate NADH from glycogen, we titrated CP in a range of doses. Optimal dosage of the inhibitor is determined by comparing the NADH of inhibitor-containing groups to that of MC-0 only group. We identified that a 100μM dosage of CP provides optimal inhibition of glycogen phosphorylase.
Since BMDCs are partially adherent cells, we are interested in whether we could observe similarly in non-adherent immune cells. Thus, we set up a similar assay using activated T cells as in Fig. 3B and measured the kinetics of NADH production. Although T cells can generate NADH, the optimal inhibitory dose of CP is much lower than that of BMDCs, which, we believe, is most likely due to a difference in cell size.
DISCUSSION
We have previously utilized pre-designed Biolog inc., PM assay plates to investigate the effect of inhibition of mechanistic target of rapamycin (mTOR) in various nutrient sources (2). In our recent publication, we also utilized PM-M1 plates to screen for potential nutrient sources that can serve as an intracellular carbon storage (11). While the pre-designed PM assays are extremely beneficial in screening a mass number of nutrient sources, the idea of utilizing this technology to conveniently investigate a specific carbon source or metabolic pathway prompted us to modify this assay by using a combination of in-house reagents and Biolog inc., products.
Although the assay adaptation and procedure relatively simple, there are several steps that require attention in order to achieve optimal results. In this paper, we examined and validated glycogen utilization in BMDCs and activated T cells. The seeding density for both types of cells are different due to differences in cell size. It is best to optimize the cell number for each cell type of interest in order to obtain optimal assay signal. Too few or too many cells will result in inconsistent assay signal and high variability among technical replicates. It is recommended that the cells be in a monolayer (if adherent). If the wells are too crowded, nutrients in the basal medium can be exhausted quickly even before dye addition, and this can result in inaccurate readings. Table 2 describes the approximate cell numbers for each immune cell type we tested, for a 96-well microplate from Biolog inc. Other types of 96-well flat bottom plates can be used, in which case, the cell number and total medium volume per well should be adjusted. Extreme deviation from a monolayer of cells can cause adverse effects on nutrient utilization and impact the rate of NADH production.
Table 2.
Guidelines for different immune cell numbers for a 96-well microplate set-up.
| Cell Type | Cell # per well of a 96-well Biolog microplate |
|---|---|
| BMDCs (Murine Bone marrow-derived DCs) | 50,000 |
| Human monocyte-derived DCs | 50,000 – 80,000 |
| Lymphocytes – naïve or activated B and T cells | 100,000 |
In our assays, we incubated the cells overnight in MC-0 medium to allow the cells to utilize minimal nutrients present in MC-0 before they switch to the saturating single nutrient source. This incubation time can be adjusted to the same day or a longer period by varying the serum concentration (usually from a range of 0 to 20% FBS) in IF-M1 media. In some cases, an extended incubation period might be necessary after the addition of reduction dye to be able to observe distinct purple color formation. This is likely due to slow metabolic responses in some cells. These adjustments depend on the metabolic needs or activities of an individual cell type of interest. Serum can also be omitted in IF-M1 medium if the same day analysis is performed or if testing for a certain substrate or pharmacological inhibitor. Regardless of serum concentration, the end goal is to obtain a linear reading of NADH over a certain period of time.
The choice of Biolog dye mix MA or MB depends on the cell type, and in this work we characterize the use of Dye Mix MA for myeloid (2) and lymphoid immune cells (Fig 3). For other cell types, extensive information about which dye to use can be found at:http://biolog.com/pdf/pmmlit/00P%20133rC%20Redox%20Dye%20Mix%20Brochure%20JUL07.pdf. While other commercial formazan dyes are commercially available, we found that, based on our experience, Biolog Dye Mix provides the most reliable results, likely due to the fact that 1) a wide range of serum concentration does not impact the spectral absorbance of the dye and 2) colored chemical inhibitors cause very little interference in absorption. Of note, these Dye Mixes should be protected from light as discoloration can occur and create a higher background, thus potentially skewing the data collected.
While we found this assay platform extremely useful and intituitive, one major draw back could be found in rapidly proliferating cells, such as activated T cells. The rate of proliferation could impact the results due to a varying degree of final cell numbers over a period of kinetic readings or during overnight incubation. Such problem can be potentially solved by performing the same day assay and multiple replicates of experiments. Another point to consider is that one has to choose the right type of base medium, IF-M1 or IF-M2 depending on the type of metabolic pathways or nutrients of interest. For instance, if there is interest in examining the metabolism of a specific amino acid, it’s best to substitute IF-M2, which lacks all 20 amino acids, for IF-M1, and subsequently add back the amino acid/s of choice.
We measured dye reduction kinetically, according to the guidelines by Biolog inc., until the kinetics readings reach linear, which varies from one cell type to another depending on their metabolic activities. For our purpose, we typically normalize the data to baseline readings at time point “0” after dye addition so that our baseline is a value of “1” and kinetic production of NADH reflects a fold-change of signal above baseline. One of the real strengths of the Biolog assay platform, is the ability to couple specific nutrient conditions to energetic production over a dynamic kinetic range. This is in distinct contrast to many other platorms in the field where researchers are forced to choose between either nutrient or energetic constraints. In addition, other kinetic assay (such as the Extracellular Flux Analysis from Agilent/Seahorse Biociences) are not directly coupled to the production of reducing equivalents. Overall, the benefits of this assay outweigh its limitations, and this customizable assay platform presents a powerful tool to better understand complex metabolic networks at the cellular level.
Acknowledgments
The authors would like to acknowledge Dr. Ralph Budd, Dr. Paula Deming, and the VCIID COBRE for extensive support; Dr. Barry Bochner for technical advice and manuscript proofreading; Michael Secinaro for providing activated T cells for some of the described assays. Funding sources: 2016 AAI Careers in Immunology Fellowship (P.T. and E.A.), UVM College of Nursing and Health Sciences Incentive Grant (E.A.), UVM start-up Funds (E.A.), P30GM118228 (E.A.), 1R21AI135385-01A1 (E.A.).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
CONFLICT OF INTEREST STATEMENT
The authors declare no conflict of interest related to this work.
References
- 1.Amiel E, Everts B, Freitas TC, King IL, Curtis JD, Pearce EL, Pearce EJ. Inhibition of mechanistic target of rapamycin promotes dendritic cell activation and enhances therapeutic autologous vaccination in mice. Journal of immunology (Baltimore, Md : 1950) 2012;189(5):2151–8. doi: 10.4049/jimmunol.1103741. Epub 2012/07/25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Amiel E, Everts B, Fritz D, Beauchamp S, Ge B, Pearce EL, Pearce EJ. Mechanistic target of rapamycin inhibition extends cellular lifespan in dendritic cells by preserving mitochondrial function. Journal of immunology (Baltimore, Md : 1950) 2014;193(6):2821–30. doi: 10.4049/jimmunol.1302498. Epub 2014/08/12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Everts B, Amiel E, Huang SC, Smith AM, Chang CH, Lam WY, Redmann V, Freitas TC, Blagih J, van der Windt GJ, Artyomov MN, Jones RG, Pearce EL, Pearce EJ. TLR-driven early glycolytic reprogramming via the kinases TBK1-IKKvarepsilon supports the anabolic demands of dendritic cell activation. Nature immunology. 2014;15(4):323–32. doi: 10.1038/ni.2833. Epub 2014/02/25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Everts B, Amiel E, van der Windt GJ, Freitas TC, Chott R, Yarasheski KE, Pearce EL, Pearce EJ. Commitment to glycolysis sustains survival of NO-producing inflammatory dendritic cells. Blood. 2012;120(7):1422–31. doi: 10.1182/blood-2012-03-419747. Epub 2012/07/13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Krawczyk CM, Holowka T, Sun J, Blagih J, Amiel E, DeBerardinis RJ, Cross JR, Jung E, Thompson CB, Jones RG, Pearce EJ. Toll-like receptor-induced changes in glycolytic metabolism regulate dendritic cell activation. Blood. 2010;115(23):4742–9. doi: 10.1182/blood-2009-10-249540. Epub 2010/03/31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Frauwirth KA, Riley JL, Harris MH, Parry RV, Rathmell JC, Plas DR, Elstrom RL, June CH, Thompson CB. The CD28 signaling pathway regulates glucose metabolism. Immunity. 2002;16(6):769–77. doi: 10.1016/s1074-7613(02)00323-0. [DOI] [PubMed] [Google Scholar]
- 7.Freemerman AJ, Johnson AR, Sacks GN, Milner JJ, Kirk EL, Troester MA, Macintyre AN, Goraksha-Hicks P, Rathmell JC, Makowski L. Metabolic reprogramming of macrophages: glucose transporter 1 (GLUT1)-mediated glucose metabolism drives a proinflammatory phenotype. The Journal of biological chemistry. 2014;289(11):7884–96. doi: 10.1074/jbc.M113.522037.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Macintyre AN, Gerriets VA, Nichols AG, Michalek RD, Rudolph MC, Deoliveira D, Anderson SM, Abel ED, Chen BJ, Hale LP, Rathmell JC. The glucose transporter Glut1 is selectively essential for CD4 T cell activation and effector function. Cell metabolism. 2014;20(1):61–72. doi: 10.1016/j.cmet.2014.05.004.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Maciver NJ, Jacobs SR, Wieman HL, Wofford JA, Coloff JL, Rathmell JC. Glucose metabolism in lymphocytes is a regulated process with significant effects on immune cell function and survival. Journal of leukocyte biology. 2008;84(4):949–57. doi: 10.1189/jlb.0108024.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bochner BR, Siri M, Huang RH, Noble S, Lei XH, Clemons PA, Wagner BK. Assay of the multiple energy-producing pathways of mammalian cells. PLoS One. 2011;6(3):e18147. doi: 10.1371/journal.pone.0018147.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Thwe PM, Pelgrom L, Cooper R, Beauchamp S, Reisz JA, D’Alessandro A, Everts B, Amiel E. Cell-Intrinsic Glycogen Metabolism Supports Early Glycolytic Reprogramming Required for Dendritic Cell Immune Responses. Cell metabolism. 26(3):558–67.e5. doi: 10.1016/j.cmet.2017.08.012. [DOI] [PMC free article] [PubMed] [Google Scholar]