

ABSTRACT
Cyclic di-AMP (c-di-AMP) is an important second messenger in bacteria. In most Firmicutes, the molecule is required for growth in complex media but also toxic upon accumulation. In an article on their current study, Zarrella and coworkers present a suppressor analysis of a Streptococcus pneumoniae strain that is unable to degrade c-di-AMP (T. M. Zarrella, D. W. Metzger, and G. Bai, J Bacteriol 200:e00045-18, 2018, https://doi.org/10.1128/JB.00045-18). Their study identifies new links between c-di-AMP and potassium homeostasis and supports the hypothesis that c-di-AMP serves as a second messenger to report about the intracellular potassium concentrations.
KEYWORDS: second messenger, Streptococcus pneumoniae, potassium uptake, phosphodiesterase, diadenylate cyclase
TEXT
Bacteria use second messengers in order to process environmental information. These molecules—often specific nucleotides—are synthesized in response to primary signals and are then perceived by intracellular targets, typically protein or RNA molecules (1). Among all second messengers, cyclic di-AMP (c-di-AMP) is unique due to three exceptional features. First, c-di-AMP is the only second messenger that is essential for many bacteria that produce it, including the model organism Bacillus subtilis and the pathogens Staphylococcus aureus, Listeria monocytogenes, and Streptococcus pneumoniae, as well as the genome-reduced bacterium Mycoplasma pneumoniae (2–8). Second, the Firmicutes are unable to grow under standard conditions on complex media in the absence of c-di-AMP, but its accumulation poses problems for the bacteria as well; c-di-AMP has therefore been coined an “essential poison” (9). For B. subtilis and M. pneumoniae, loss of the phosphodiesterases that degrade c-di-AMP leads to cell death (8–10). Third, c-di-AMP is the only second messenger that modulates a biological process by binding to a protein and also to an RNA molecule that controls its expression. This was discovered in B. subtilis, where c-di-AMP limits potassium uptake by binding to the KtrA signaling subunit of the KtrAB potassium transporter as well as to the riboswitch in the untranslated region of the ktrAB mRNA. Binding of c-di-AMP to the riboswitch prevents transcription beyond the riboswitch and thus reduces the expression of the potassium transporter (11, 12).
The essential role of c-di-AMP for the bacterial physiology and its requirement for the virulence of several important pathogens have attracted considerable interest in the elucidation of the reason for its essentiality and for its functions in the cell (reviewed in references 13 to 15). The data obtained so far suggest key functions of c-di-AMP in the control of central metabolism, potassium homeostasis, and osmotic adaptation (6, 15–18). However, the identification of target proteins of so far unknown function (11, 18, 19) suggests that c-di-AMP may be involved in additional processes.
The making and breaking of cyclic di-AMP involve two classes of enzymes: the diadenylate cyclases and phosphodiesterases, respectively. The diadenylate cyclases are characterized by a conserved catalytically active domain, the so-called DAC (diadenylate cyclase) domain (13, 14, 20). Four types of cyclases have so far been investigated. They differ by the regulatory domains that are attached to the DAC domain (8, 14). B. subtilis carries genes that encode three diadenylate cyclases: DisA, CdaA, and CdaS. DisA and CdaS have been implicated in the control of DNA integrity and spore germination, respectively (21, 22). CdaA is the most widespread diadenylate cyclase, and it is the only cyclase in pathogenic Firmicutes (14). This membrane protein consists of an N-terminal transmembrane domain and the enzymatically active DAC domain, which is flanked by coiled-coil domains on each side (9, 20). As observed for the cyclases, many bacteria possess multiple c-di-AMP-degrading phosphodiesterases. One class of phosphodiesterases is characterized by conserved DHH/DHH1 domains, whereas the enzymes of the second class possess an HD domain. L. monocytogenes and B. subtilis have a phosphodiesterase of each class (9, 23). S. aureus only encodes one c-di-AMP-degrading enzyme of the DHH/DHH1 family, whereas a second cytoplasmic DHH/DHH1 class protein degrades the linear dinucleotide pApA rather than c-di-AMP (24). In S. pneumoniae, two DHH/DHH1-type phosphodiesterases are present. One of these enzymes cleaves c-di-AMP to two molecules of AMP, whereas the second enzyme produces pApA (25). Loss of these enzymes results in the accumulation of c-di-AMP and loss of virulence of the bacteria (25, 26).
MUTATIONS AFFECTING THE MAJOR DIADENYLATE CYCLASE CdaA PREVENT c-di-AMP ACCUMULATION
As mentioned above, not only is c-di-AMP essential in Gram-positive bacteria, its accumulation is also detrimental to the cells or even toxic (3, 9). In particular, several studies have revealed an increased sensitivity to salt and other stress factors, which is often accompanied by resistance to β-lactam antibiotics (2, 25, 27–30 [see reference 15 for review]). Based on the impaired growth of strains lacking the phosphodiesterases, suppressor mutations have been identified that provide novel insights into c-di-AMP signaling. In B. subtilis, such strains that lack both phosphodiesterases rapidly lyse and acquire suppressor mutations that allow survival (9). An analysis of those mutants revealed that in all cases the major diadenylate cyclase CdaA was affected: frameshift mutations resulted in the expression of truncated CdaA proteins that lack the catalytically relevant DGA and RHR motifs in the DAC domain or in a deletion of the C-terminal eight amino acids of CdaA. The inactivation of CdaA in B. subtilis upon loss of the phosphodiesterases made DisA the only expressed diadenylate cyclase in these mutants. Simultaneous deletion of the genes encoding both phosphodiesterases and DisA provoked a mutation in the ribosomal binding site of the cdaA mRNA, resulting in reduced CdaA synthesis (9). Taken together, these results suggested that CdaA is the major diadenylate cyclase in B. subtilis and that c-di-AMP produced by this enzyme is the cause of c-di-AMP toxicity. Zhu et al. have isolated suppressor mutants that overcome the salt sensitivity of Lactococcus lactis strains lacking the single c-di-AMP-degrading phosphodiesterase (28). With an impressive set of more than 60 distinct mutations affecting CdaA, and thus reduced levels of c-di-AMP, this study convincingly reinforced the conclusion that c-di-AMP accumulation causes osmosensitivity in L. lactis (28). The current study by Zarrella et al. (31) adds another flavor to the adaptation of c-di-AMP-accumulating bacteria to stress conditions (Fig. 1). S. pneumoniae lacking both phosphodiesterases exhibits reduced growth on complex media and increased sensitivity to stress conditions, including osmotic and heat stresses (31). The suppressor mutants that are more resistant to heat stress carry mutations, and two distinct mutations affecting the single diadenylate cyclase CdaA are involved in this resistance: a V76G substitution at the outer side of the third transmembrane helix, and a frameshift at the C terminus-encoding end of the cdaA gene that results in the addition of a 32-amino-acid tail. Interestingly, a mutation corresponding to the V76G mutation was also found in the L. lactis suppressor screen, and mutations affecting the C terminus of the protein were detected in both B. subtilis and L. lactis (9, 28). Both mutations resulted in a severe reduction of the c-di-AMP concentrations and restored the wild-type levels of the nucleotide. Interestingly, one of the mutants had an additional substitution affecting the potassium transporter TrkH, but this mutation made only a minor contribution to the reduction of the c-di-AMP levels (31).
FIG 1.
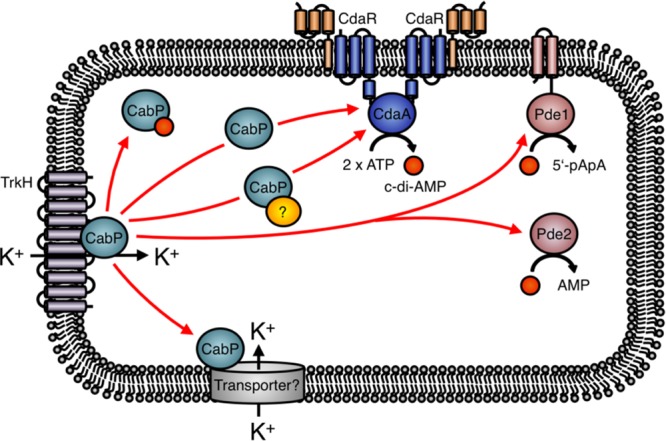
c-di-AMP signaling in Streptococcus pneumoniae. CdaR directly interacts with the diadenylate cyclase CdaA and regulates its activity (7). The red arrows indicate the possibilities of how the Trk family protein CabP modulates the cellular c-di-AMP level. The CabP-dependent control of c-di-AMP synthesis or degradation might also depend on additional factors, as indicated by the yellow circle. Moreover, CabP could control the activity of an unknown additional potassium transporter, and the changes in cellular potassium concentration could then affect the c-di-AMP levels.
A LINK BETWEEN POTASSIUM UPTAKE AND c-di-AMP SYNTHESIS
In all organisms studied so far, at least one of the c-di-AMP target molecules is involved in potassium homeostasis, and in several more intensively studied bacteria, such as S. aureus, L. monocytogenes, and B. subtilis, even multiple potassium-related targets have been detected. In these three bacteria, c-di-AMP not only binds to the cytoplasmic gating subunits of potassium transporters to control their activity but also binds to proteins and RNA molecules that control the expression of potassium transporters (14). Interestingly, c-di-AMP binds KdpD, a sensor kinase of a two-component system that activates expression of the high-affinity potassium uptake system KdpABC in S. aureus and L. monocytogenes. B. subtilis does not possess the Kdp system and relies on the KtrAB and the newly discovered KimA transporters for high-affinity uptake of potassium (17, 32, 33). The ktrAB and kimA genes for these transporters are each under the control of a c-di-AMP-responsive riboswitch that causes premature transcription termination upon binding of the second messenger (12, 17). Thus, potassium uptake is reduced as long as the intracellular concentration of c-di-AMP is high. This connection between potassium homeostasis and c-di-AMP signaling is also reinforced by the current study of Zarrella et al. (31) (Fig. 1). First of all, a mutation in TrkH, the integral membrane subunit of the potassium transporter TrkH-CabP, contributes to the decrease in the c-di-AMP levels in one of the suppressor mutants that were isolated as a result of c-di-AMP accumulation (described above). CabP, the gating component of this transporter, has previously been identified as a target of c-di-AMP in S. pneumoniae (34). This protein is similar to the c-di-AMP-binding KtrA proteins from B. subtilis and other Firmicutes. Interestingly, the TrkH variant isolated by Zarrella et al. allows improved growth of the strain lacking any c-di-AMP-degrading phosphodiesterases on complex media. As c-di-AMP binding to CabP inhibits potassium uptake, the mutant TrkH protein may exhibit improved potassium transport. The subunits of the TrkH-CabP complex have different effects on the intracellular c-di-AMP levels: while a deletion of the trkH gene has no effect, the deletion of cabP results in a reduction of the c-di-AMP levels by about 50%. This finding indicates that CabP is involved in the control of c-di-AMP homeostasis, even though the current work could not yet identify the precise mechanism. In S. aureus, KtrA controls the activity of two distinct membrane proteins, KtrB and KtrD. Thus, it is tempting to speculate that CabP serves as gating subunit not only for TrkH but also for a second membrane-spanning transporter. Indeed, the genome of S. pneumoniae also encodes the protein SPD_0429, which is most similar to KtrB. Thus, deletion of trkH may not be sufficient to reduce potassium uptake due to the activity of SPD_0429. Loss of CabP as the single gating subunit for both transporters would in any case result in reduced potassium uptake. In B. subtilis, reduced potassium levels also result in reduced c-di-AMP levels (17). Thus, the presence of a second potassium transporter subunit nicely explains the seemingly contradictory results obtained with the trkH and cabP mutants.
c-di-AMP, A KEY PLAYER IN CONTROLLING OSMOLYTE TRANSPORT IN GRAM-POSITIVE BACTERIA?
In the past few years, several studies have demonstrated that perturbations of c-di-AMP metabolism affect the integrity of the protective cell wall in Gram-positive bacteria, including streptococci (for a recent review, see reference 15). For instance, elevated cellular concentrations of c-di-AMP due to the loss of the phosphodiesterase GdpP in L. monocytogenes, L. lactis, and B. subtilis reduced the susceptibility of the bacteria to β-lactams (2, 28, 35). A Streptococcus suis mutant tolerating large amounts of lysozyme had acquired a mutation in the cdaA diadenylate cyclase gene, which probably alters the cellular c-di-AMP concentration (36). In contrast, a decrease of the cellular c-di-AMP levels in B. subtilis and L. monocytogenes caused cell lysis (2, 7, 17). Moreover, strains that synthesize smaller amounts of c-di-AMP are more susceptible to cell wall-targeting antibiotics (15). Therefore, it has been concluded that c-di-AMP is essential for proper cell envelope synthesis (2, 7, 35). However, today we know that the nucleotide rather indirectly affects the integrity of the cell envelope by modulating the cellular turgor, a physical variable that has to be tightly adjusted depending on the osmotic pressure of the environment (15). For B. subtilis, L. monocytogenes, and S. aureus, it has been shown that c-di-AMP becomes dispensable for the cell if the bacteria are cultivated in a chemically defined medium that does not lead to the excessive accumulation of osmolytes and amino acids (6, 16, 17). Thus, many c-di-AMP-related cell wall phenotypes in Firmicutes can be explained by the perturbation of osmolyte homeostasis.
As described above, c-di-AMP plays a crucial role in controlling the activities of potassium uptake systems in a variety of Gram-positive bacteria, including S. pneumoniae. Recently, it has been uncovered that c-di-AMP also controls the uptake of other osmolytes, such as the compatible solutes carnitine and glycine-betaine. c-di-AMP inhibits uptake of carnitine via the high-affinity OpuC ATP-binding cassette (ABC) transport systems in L. monocytogenes and S. aureus (29, 37). The OpuC transport system consists of the subunits OpuCA, OpuCB, OpuCC, and OpuCD (38). OpuCA is a cytoplasmic protein that contains tandem cystathionine-β-synthase (CBS) and ABC domains, of which the latter fuels the transport activity of the Opu system. c-di-AMP was shown to inhibit carnitine uptake by binding in the cleft between the two CBS domains of the OpuCA subunit (29). Thus, c-di-AMP prevents uptake of carnitine to toxic levels, as is the case for the osmolyte potassium. It is interesting to note that many bacterial ABC transporters involved in the transport of compatible solutes contain tandem CBS domains (39). In B. subtilis and L. lactis, the CBS domains in the osmoregulatory ABC transporter OpuA, which transports the osmolyte glycine-betaine, sense the ionic strength and control the transport activity (39, 40). However, it has to be tested whether c-di-AMP also controls the activity of the OpuA transport system in B. subtilis and L. lactis. Interestingly, c-di-AMP also binds to the CBS domain-containing proteins CbpA and CbpB from L. monocytogenes (18). The OpuCA homolog of S. pneumoniae (designated PotA) does not possess CBS domains, and it is therefore not involved in the c-di-AMP-dependent uptake of osmolytes. However, in future studies it will be interesting to elucidate whether c-di-AMP is also a key player of osmolyte homeostasis in S. pneumoniae as is the case in other Firmicutes. For this purpose, a saturating suppressor screen with an S. pneumoniae cdaA mutant isolated in the current study could be a straightforward approach to identify novel c-di-AMP-binding proteins that are probably involved in the transport of osmoactive substances. Moreover, it is essential to elucidate how the diadenylate cyclases and the phosphodiesterases that synthesize and degrade c-di-AMP, respectively, sense the potassium concentrations and/or the cellular turgor to adjust uptake and export of osmolytes. The study by Zarrella et al. (31) reveals that the S. pneumoniae c-di-AMP effector protein CabP modulates the synthesis of the nucleotide, probably by controlling the activity of the diadenylate cyclase CdaA (31). As discussed above, CabP could affect the c-di-AMP pool by modulating potassium uptake via TrkH and SPD_0429. As a consequence, the alteration of the c-di-AMP concentration could be due to a change in diadenylate cyclase activity that responds to potassium. Therefore, the precise role of CabP in controlling the cellular c-di-AMP levels should be elaborated in future studies (Fig. 1).
ACKNOWLEDGMENT
This work was supported by grants from the Deutsche Forschungsgemeinschaft via Priority Program SPP 1897 (to J.S. and F.M.C.).
The views expressed in this Commentary do not necessarily reflect the views of the journal or of ASM.
Footnotes
For the article discussed, see https://doi.org/10.1128/JB.00045-18.
REFERENCES
- 1.Gomelsky M. 2011. cAMP, c-di-GMP, c-di-AMP, and now cGMP: bacteria use them all! Mol Microbiol 79:562–565. doi: 10.1111/j.1365-2958.2010.07514.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Luo Y, Helmann JD. 2012. Analysis of the role of Bacillus subtilis σM in β-lactam resistance reveals an essential role for c-di-AMP in peptidoglycan homeostasis. Mol Microbiol 83:623–639. doi: 10.1111/j.1365-2958.2011.07953.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Mehne FMP, Gunka K, Eilers H, Herzberg C, Kaever V, Stülke J. 2013. Cyclic-di-AMP homeostasis in Bacillus subtilis: both lack and high-level accumulation of the nucleotide are detrimental for cell growth. J Biol Chem 288:2004–2017. doi: 10.1074/jbc.M112.395491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Song JH, Ko KS, Lee JY, Baek JY, Oh WS, Yoon HS, Jeong JY, Chun J. 2005. Identification of essential genes in Streptococcus pneumoniae by allelic replacement mutagenesis. Mol Cells 19:365–374. [PubMed] [Google Scholar]
- 5.Chaudhuri RR, Allen AG, Owen PJ, Shalom G, Stone K, Harrison M, Burgis TA, Lockyer M, Garcia-Lara J, Foster SJ, Pleasance SJ, Peters SE, Maskell DJ, Charles IG. 2009. Comprehensive identification of essential Staphylococcus aureus genes using transposon-mediated differential hybridization (TMDH). BMC Genomics 10:291. doi: 10.1186/1471-2164-10-291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Whiteley AT, Pollock AJ, Portnoy DA. 2015. The PAMP c-di-AMP is essential for Listeria growth in macrophages and rich but not minimal medium due to a toxic increase in (p)ppGpp. Cell Host Microbe 17:788–798. doi: 10.1016/j.chom.2015.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Rismondo J, Gibhardt J, Rosenberg J, Kaever V, Halbedel S, Commichau FM. 2016. Phenotypes associated with the essential diadenylate cyclase CdaA and its potential regulator CdaR in the human pathogen Listeria monocytogenes. J Bacteriol 198:416–426. doi: 10.1128/JB.00845-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Blötz C, Treffon K, Kaever V, Schwede F, Hammer E, Stülke J. 2017. Identification of the components involved in cyclic di-AMP signaling in Mycoplasma pneumoniae. Front Microbiol 8:1328. doi: 10.3389/fmicb.2017.01328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Gundlach J, Mehne FMP, Herzberg C, Kampf J, Valerius O, Kaever V, Stülke J. 2015. An essential poison: synthesis and degradation of cyclic di-AMP in Bacillus subtilis. J Bacteriol 197:3265–3274. doi: 10.1128/JB.00564-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Huynh TN, Woodward JJ. 2016. Too much of a good thing: regulated depletion of c-di-AMP in the bacterial cytoplasm. Curr Opin Microbiol 30:22–29. doi: 10.1016/j.mib.2015.12.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Corrigan RM, Campeotto I, Jeganathan T, Roelofs KG, Lee VT, Gründling A. 2013. Systematic identification of conserved bacterial c-di-AMP receptor proteins. Proc Natl Acad Sci U S A 110:9084–9089. doi: 10.1073/pnas.1300595110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Nelson JW, Sudarsan N, Furukawa K, Weingerg Z, Wang JX, Breaker RR. 2013. Riboswitches in eubacteria sense the second messenger cyclic di-AMP. Nat Chem Biol 9:834–839. doi: 10.1038/nchembio.1363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Corrigan RM, Gründling A. 2013. Cyclic di-AMP: another second messenger enters the fray. Nat Rev Microbiol 11:513–524. doi: 10.1038/nrmicro3069. [DOI] [PubMed] [Google Scholar]
- 14.Commichau FM, Dickmanns A, Gundlach J, Ficner R, Stülke J. 2015. A jack of all trades: the multiple roles of the unique essential second messenger cyclic di-AMP. Mol Microbiol 97:189–204. doi: 10.1111/mmi.13026. [DOI] [PubMed] [Google Scholar]
- 15.Commichau FM, Gibhardt J, Halbedel S, Gundlach J, Stülke J. 2018. A delicate connection: c-di-AMP affects cell integrity by controlling osmolyte transport. Trends Microbiol 26:175–185. doi: 10.1016/j.tim.2017.09.003. [DOI] [PubMed] [Google Scholar]
- 16.Zeden MS, Schuster CF, Bowman L, Zhong Q, Williams HD, Gründling A. 2018. Cyclic di-adenosine monophosphate (c-di-AMP) is required for osmotic regulation in Staphylococcus aureus but dispensable for viability in anaerobic conditions. J Biol Chem 293:3180–3200. doi: 10.1074/jbc.M117.818716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Gundlach J, Herzberg C, Kaever V, Gunka K, Hoffmann T, Weiß M, Gibhardt J, Thürmer A, Hertel D, Daniel R, Bremer E, Commichau FM, Stülke J. 2017. Control of potassium homeostasis is an essential function of the second messenger cyclic di-AMP in Bacillus subtilis. Sci Signal 10:eaal3011. doi: 10.1126/scisignal.aal3011. [DOI] [PubMed] [Google Scholar]
- 18.Sureka K, Choi PH, Precit M, Delince M, Pensinger DA, Huynh TN, Jurado AR, Goo YA, Sadilek M, Iavarone AT, Sauer JD, Tong L, Woodward JJ. 2014. The cyclic dinucleotide c-di-AMP is an allosteric regulator of metabolic enzyme function. Cell 158:1389–1401. doi: 10.1016/j.cell.2014.07.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gundlach J, Dickmanns A, Schröder-Tittmann K, Neumann P, Kaesler J, Kampf J, Herzberg C, Hammer E, Schwede F, Kaever V, Tittmann K, Stülke J, Ficner R. 2015. Identification, characterization and structure analysis of the c-di-AMP binding PII-like signal transduction protein DarA. J Biol Chem 290:3069–3080. doi: 10.1074/jbc.M114.619619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Rosenberg J, Dickmanns A, Neumann P, Gunka K, Arens J, Kaever V, Stülke J, Ficner R, Commichau FM. 2015. Structural and biochemical analysis of the essential diadenylate cyclase CdaA from Listeria monocytogenes. J Biol Chem 290:6596–6606. doi: 10.1074/jbc.M114.630418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gandara C, Alonso JC. 2015. DisA and c-di-AMP act at the intersection between DNA-damage response and stress homeostasis in exponentially growing Bacillus subtilis cells. DNA Repair 27:1–8. doi: 10.1016/j.dnarep.2014.12.007. [DOI] [PubMed] [Google Scholar]
- 22.Mehne FMP, Schröder-Tittmann K, Eijlander RT, Herzberg C, Hewitt L, Kaever V, Lewis RJ, Kuipers OP, Tittmann K, Stülke J. 2014. Control of the diadenylate cyclase CdaS in Bacillus subtilis: an autoinhibitory domain limits c-di-AMP production. J Biol Chem 289:21098–21107. doi: 10.1074/jbc.M114.562066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Huynh TN, Luo S, Pensinger D, Sauer JD, Tong L, Woodward JJ. 2015. An HD-domain phosphodiesterase mediates cooperative hydrolysis of c-di-AMP to affect bacterial growth and virulence. Proc Natl Acad Sci U S A 112:E747–E756. doi: 10.1073/pnas.1416485112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bowman L, Zeden MS, Schuster CF, Kaever V, Gründling A. 2016. New insights into the cyclic di-adenosine monophosphate (c-di-AMP) degradation pathway and the requirement of the cyclic dinucleotide for acid stress resistance in Staphylococcus aureus. J Biol Chem 291:26970–26986. doi: 10.1074/jbc.M116.747709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bai Y, Yang J, Eisele LE, Underwood AJ, Koestler BJ, Waters CM, Metzger DW, Bai G. 2013. Two DHH subfamily 1 proteins in Streptococcus pneumoniae possess cyclic di-AMP phosphodiesterase activity and affect bacterial growth and virulence. J Bacteriol 195:5123–5132. doi: 10.1128/JB.00769-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Cron LE, Stol K, Burghout P, van Selm S, Simonetti ER, Bootsma HJ, Hermans PW. 2011. Two DHH subfamily 1 proteins contribute to pneumococcal virulence and confer protection against pneumococcal disease. Infect Immun 79:3697–3710. doi: 10.1128/IAI.01383-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wang X, Davlieva M, Reyes J, Panesso D, Arias CA, Shamoo Y. 2017. Novel phosphodiesterase of the GdpP family modulates cyclic di-AMP levels in response to cell membrane stress in daptomycin-resistant enterococci. Antimicrob Agents Chemother 61:e01422-16. doi: 10.1128/AAC.01422-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zhu Y, Pham TH, Nhiep TH, Vu NM, Marcellin E, Chakrabortti A, Wang Y, Waanders J, Lo R, Huston WM, Bansal N, Nielsen LK, Liang ZX, Turner MS. 2016. Cyclic di-AMP synthesis by the diadenylate cyclase CdaA is modulated by the peptidoglycan biosynthesis enzyme GlmM in Lactococcus lactis. Mol Microbiol 99:1015–1027. doi: 10.1111/mmi.13281. [DOI] [PubMed] [Google Scholar]
- 29.Huynh TN, Choi PH, Sureka K, Ledvina HE, Campillo J, Tong L, Woodward JJ. 2016. Cyclic di-AMP targets the cystathione beta-synthase domain of the osmolyte transporter OpuC. Mol Microbiol 102:233–243. doi: 10.1111/mmi.13456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Griffiths JM, O'Neill AJ. 2012. Loss of function of the GdpP protein leads to joint β-lactam/glycopeptide tolerance in Staphylococcus aureus. Antimicrob Agents Chemother 56:579–581. doi: 10.1128/AAC.05148-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Zarrella TM, Metzger DW, Bai G. 2018. Stress suppressor screening leads to detection of regulation of cyclic di-AMP homeostasis by a Trk-family effector protein in Streptococcus pneumoniae. J Bacteriol 200:e00045-18. doi: 10.1128/JB.00045-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Holtmann G, Bakker EP, Uozumi N, Bremer E. 2003. KtrAB and KtrCD: two K+ uptake systems in Bacillus subtilis and their role in adaptation to hypertonicity. J Bacteriol 185:1289–1298. doi: 10.1128/JB.185.4.1289-1298.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Gundlach J, Herzberg C, Hertel D, Thürmer A, Daniel R, Link H, Stülke J. 2017. Adaptation of Bacillus subtilis to life at extreme potassium limitation. mBio 8:e00861-17. doi: 10.1128/mBio.00861-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Bai Y, Yang J, Zarrella TM, Zhang Y, Metzger DW, Bai G. 2014. Cyclic di-AMP impairs potassium uptake mediated by a cyclic di-AMP binding protein in Streptococcus pneumoniae. J Bacteriol 196:614–623. doi: 10.1128/JB.01041-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Witte CE, Whiteley AT, Burke TP, Sauer JD, Portnoy DA, Woodward JJ. 2013. Cyclic di-AMP is critical for Listeria monocytogenes growth, cell wall homeostasis, and establishment of infection. mBio 4:e00282-13. doi: 10.1128/mBio.00282-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wichgers Schreur PJ, van Weeghel C, Rebel JM, Smits MA, van Putten JP, Smith HE. 2012. Lysozyme resistance in Streptococcus suis is highly variable and multifactorial. PLoS One 7:e36281. doi: 10.1371/journal.pone.0036281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Schuster CF, Bellows LE, Tosi T, Campeotto I, Corrigan RM, Freemont P, Gründling A. 2016. The second messenger c-di-AMP inhibits the osmolyte uptake system OpuC in Staphylococcus aureus. Sci Signal 9:ra81. doi: 10.1126/scisignal.aaf7279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hoffmann T, Bremer E. 2017. Guardians in a stressful world: the Opu family of compatible solute transporters from Bacillus subtilis. Biol Chem 398:193–214. doi: 10.1515/hsz-2016-0265. [DOI] [PubMed] [Google Scholar]
- 39.Biemans-Oldehinkel E, Mahmood NA, Poolman B. 2006. A sensor for intracellular ionic strength. Proc Natl Acad Sci U S A 103:10624–10629. doi: 10.1073/pnas.0603871103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Horn C, Bremer E, Schmitt L. 2003. Nucleotide dependent monomer/dimer equilibrium of OpuAA, the nucleotide binding protein of the osmotically regulated ABC transporter OpuA from Bacillus subtilis. J Mol Biol 334:403–419. doi: 10.1016/j.jmb.2003.09.079. [DOI] [PubMed] [Google Scholar]