

Abstract
Cross education (CE) occurs after unilateral training whereby performance of the untrained contralateral limb is enhanced. A few studies have shown that CE can preserve or “spare” strength and size of an opposite immobilized limb, but the specificity (i.e., trained homologous muscle and contraction type) of these effects is unknown. The purpose was to investigate specificity of CE “sparing” effects with immobilization. The nondominant forearm of 16 participants was immobilized with a cast, and participants were randomly assigned to a resistance training (eccentric wrist flexion, 3 times/week) or control group for 4 weeks. Pre- and posttesting involved wrist flexors and extensors eccentric, concentric and isometric maximal voluntary contractions (via dynamometer), muscle thickness (via ultrasound), and forearm muscle cross-sectional area (MCSA; via peripheral quantitative computed tomography). Only the training group showed strength preservation across all contractions in the wrist flexors of the immobilized limb (training: −2.4% vs. control: −21.6%; P = 0.04), and increased wrist flexors strength of the nonimmobilized limb (training: 30.8% vs. control: −7.4%; P = 0.04). Immobilized arm MCSA was preserved for the training group only (training: 1.3% vs. control: −2.3%; P = 0.01). Muscle thickness differed between groups for the immobilized (training: 2.8% vs. control: −3.2%; P = 0.01) and nonimmobilized wrist flexors (training: 7.1% vs. control: −3.7%; P = 0.02). Strength preservation was nonspecific to contraction type (P = 0.69, = 0.03) yet specific to the trained flexors muscle. These findings suggest that eccentric training of the nonimmobilized limb can preserve size of the immobilized contralateral homologous muscle and strength across multiple contraction types.
NEW & NOTEWORTHY Unilateral strength training preserves strength, muscle thickness, and muscle cross-sectional area in an opposite immobilized limb. The preservation of size and strength was confined to the trained homologous muscle group. However, strength was preserved across multiple contraction types.
Keywords: bilateral transfer, cross education, cross transfer, eccentric strength, muscle lengthening
INTRODUCTION
Cross education (CE) of strength occurs when unilateral strength training produces performance improvement of the untrained contralateral limb (7, 36). Within the last decade, there is renewed scientific interest in CE, on account of evidence of preserved strength and muscle size in an opposite immobilized limb (26). The allure of an immobilization model in CE research is that disuse causes accelerated loss in neural drive, strength, and muscle size (10, 12, 13, 52) and the prospect of CE to attenuate these decrements can be explored.
To date, there are just four studies that have investigated the effects of CE in healthy participants with an immobilization model (20, 21, 40, 45). All four studies found that a CE intervention attenuated the strength loss in the immobilized limb. However, three of these studies also found a sparing effect for muscle size (20, 40, 45). The mechanisms of muscle size preservation are currently unclear. Current theory suggests CE is probably driven by neural mechanisms (7, 36) rather than muscle or morphological factors.
The discrepancy in findings for muscle size preservation between Farthing et al. (21) and the other three studies (20, 40, 45) warrants further investigation. All four studies used ultrasound to image muscle thickness. Although ultrasound is a valid and reliable as a muscle size measure (8), it is not considered a gold standard measure. Revisiting these size sparing effects with a measure of muscle cross-sectional area (MCSA) from peripheral quantitative computed tomography (pQCT) is an important step to confirm or refute the previous findings (26).
Regardless of ambiguity around sparing of muscle size, the strength sparing effects of CE are consistent in all prior studies. The mechanistic link, if any, between the strength and size sparing effects of CE is unknown. One approach to explore the link between size and strength sparing effects with CE is to investigate specificity. While CE of strength has been shown to be specific to the homologous muscle in the untrained contralateral limb (30, 37, 56), and to the contraction type (29) and velocity (18) used in training, the specificity of CE has never been tested for an immobilized limb. Specificity effects are convincing indices of the involvement of neural mechanisms (24); therefore, the sparing effects of CE are hypothesized to demonstrate specificity. However, specificity effects may present differently for an immobilized limb due to inhibited movement, altered corticospinal excitability (4, 15, 38, 43, 54), muscle atrophy and altered protein synthesis (1, 42), and potential systemic effects (26) that could contribute globally to muscle size sparing.
The observed sparing effect of CE has implications for improving rehabilitation of an injured or neurologically impaired limb (11, 39, 44), with the goal of restoring symmetry after unilateral injury (22). The concept of using CE for its sparing effects during rehabilitation from injury has the potential to reduce the total time of recovery, particularly if therapy is started before atrophy and strength deprivation begin to occur (22). Understanding the specificity of sparing effects of CE as it pertains to the type of contraction and antagonistic muscle pairs can inform the design of effective training programs to optimize rehabilitation and recovery from injuries. The purpose of this study was to examine muscle contraction type and task specificity of CE in an immobilized left forearm after 4 wk of eccentric (ECC) wrist flexion training of the right arm. The primary hypothesis was that the CE sparing effects on muscle strength and size would be specific to muscle (i.e., homologous wrist flexors, not extensors), task (i.e., flexion, not extension), and type [i.e., ECC strength not concentric (CON) and isometric (ISO) strength]. The secondary hypothesis was that unilateral ECC training would result in the sparing of MCSA in the immobilized limb, as measure by pQCT.
METHODS
Participants
Sixteen participants from the University of Saskatchewan student population volunteered to participate in this study (Table 1). Participants were randomly assigned to an immobilized control (n = 8) or training group (n = 8). Participants were right handed, as determined by a handedness questionnaire, healthy (i.e., no physical injuries or neurological conditions), and were classified as currently untrained (<6 mo resistance training experience in the previous year, where 1 mo of experience is equal to resistance training on average 3 times per week for 4 wk). Before the beginning of the study, written informed consent was obtained. This study conformed to the standards set by the Declaration of Helsinki and was approved by the University of Saskatchewan Behavioural and Biomedical Research Ethics Board.
Table 1.
Demographics
| Group/Sex | Age, yr | Height, cm | Weight, kg | WHQ | Training Experience, mo |
|---|---|---|---|---|---|
| Training (n = 8: 1 man and 7 women) | 20 ± 2 | 170.3 ± 10.1 | 77.2 ± 19.2 | 18.3 ± 2.4 | 2.3 ± 4.1 |
| Control (n = 8: 2 men and 6 women) | 23 ± 5 | 169.3 ± 8.5 | 85.7 ± 22.7 | 17.1 ± 2.5 | 2.9 ± 4.3 |
Data are means ± SD. WHQ, Waterloo handedness questionnaire.
Participant’s handedness was determined with the Waterloo Handedness Questionnaire (3). The questionnaire scores participants as either right (indicated by a positive score) or left handed (indicated by a negative score) on a scale of +20 representing strong right-handedness to −20 representing strong left-handedness. Participants were required to be right-handed for this study because previous literature has demonstrated greater CE of strength in right-handed individuals when training their dominant arm (16, 19). An a priori sample size calculation for the between-within factorial ANOVA design with power (1-β) = 0.95 and α = 0.05 was determined using G*Power 3.1 (23). An effect size estimate of 0.51 was calculated from a previous CE immobilization study (16). The sample size estimate was 16 participants (n = 8 per group) for the primary outcome measure of torque.
Intervention and Design
All participants received a cast on their left, nondominant, forearm for 4 wk according to our previous method (20, 21). Casts were placed by a physician, with the intent to immobilize the wrist, hand, thumb, and part of the fingers up to the middle phalanges. Notches were cut out of the cast at the proximal end, large enough for placement of electrodes for electromyography (EMG) monitoring of the wrist flexors during training sessions. The training group underwent strength training of the right wrist flexors three times per week while the immobilized control group did not train and were instructed to refrain from other forms of exercise during their participation in the study. Training involved maximal effort ECC isokinetic contractions performed through 80° of motion (40° flexion to 40° extension) with a fixed rotational velocity of 1.05 rad/s on an isokinetic dynamometer (Humac NORM; CSMi, Stoughton, MA). Strength training was progressive, commencing with two sets of eight repetitions and progressed in volume up to six sets of eight, with a taper to two sets for the last session. A 1-min rest was given between each set. Participants were prompted to position their immobilized limb in the same pronated orientation as the training limb and to relax it during all testing and training sessions to minimize mirror activity (30). The mirrored positioning of the immobilized limb was important in controlling for possible confounding effects of the orientation of the wrist and homologous or no-homologous mirror activity during unilateral movements (47).
Measures
Familiarization and testing sessions.
All participants underwent a familiarization session as an introduction to all strength and stimulation testing measures. Following familiarization, participants returned to the laboratory within 7 days for two separate pretesting sessions. Two pretesting sessions were used to determine variance of measures and to establish a stable baseline. An average of the two pretesting sessions for each strength, muscle activation, and muscle thickness measure was used in data analysis. Data collection occurred in two separate laboratories. Muscle thickness and strength measures were assessed by the primary researcher in one laboratory, while MCSA was collected in a separate laboratory by a single researcher blinded to group assignment. For pre- and posttesting, muscle thickness was always measured before strength. MCSA was scheduled before all other tests whenever possible or with a minimum of 48 h allotted for recovery. The scheduling of MCSA measures was strategic to avoid the potential influence of forearm muscle contractions (possible edema) on the accuracy of the measure. After the second pretesting session, participants received the nondominant forearm cast, initiating the intervention period. After the 4-wk intervention, participants returned to the laboratory for cast removal followed by posttesting. The order of posttesting was consistent with MCSA measured immediately after cast removal (5.7 ± 3.4 min) followed by muscle thickness and strength testing.
Peak torque.
All testing and training sessions used an identical setup and were supervised and completed on an isokinetic dynamometer. Testing sessions involved maximal effort isokinetic ECC, CON, and ISO muscle actions of the wrist flexors and extensors. The primary outcome measure was peak torque, which was recorded for each contraction type over three sets of one repetition separated by 30 s and used as a measure of contraction specific strength. The highest torque value achieved over the three maximal attempts was used as the strength value on each occasion. For each contraction type, wrist flexors were tested first followed by wrist extensors. The order of limb testing (left or right arm) was randomized and held constant for each participant for every testing session. A 1-min rest was given between each test. ECC and CON muscle actions were performed through 80° of motion (40° flexion to 40° extension) with a fixed rotational velocity of 1.05 rad/s, with ISO muscle actions performed with a neutral wrist (3-s MVC at 0° of flexion). ISO contractions were assessed first, followed by ECC and CON in a randomized order. Participants were seated in an upright position, with the elbow at 90°, and the forearm resting on a pad with the wrist in a pronated position grasping the dynamometer handle. Participants were instructed to rest their immobilized limb on their lap with the forearm in a similar pronated position to be consistent with the wrist orientation of the training limb. Verbal encouragement was provided by the same experimenter for each test. The precision error [coefficient of variation (CV%)] of our laboratory for the left and right forearm torque measures is 5.7 and 4.9%, respectively (19).
Muscle cross-sectional area.
Single operator (K. Björkman), blinded to the group randomization, obtained MCSA (cm2) from both forearms (at 65% site of radius length) using pQCT (XCT2000; Stratec, Medizintechnik, Germany) and our standard protocols (25). MCSA was analyzed using the BoneJ plugin (Version 1.3.11) for the open-source software ImageJ (48). We separated muscle tissue from subcutaneous fat using a threshold of 30 mg/cm3 and from bone using a threshold of 280 mg/cm3. Operator’s precision error (CV%RMS) of forearm MCSA was 1.8%, between repeated measures (>24 h apart) of another university student sample (Table 2) and comparable with earlier reported precision (25). MCSA from pQCT is highly correlated (r = 0.78–0.92) to MCSA derived from MRI (51), but it is unable to differentiate specific muscles (e.g., forearm flexors from extensors). Pre- and posttesting MCSA was assessed in both arms of each group (intervention and control) to assess the changes in the immobilized and nonimmobilized limb over the duration of the study. During pretesting, pQCT was measured within 7 days after the familiarization session; posttesting pQCT was measured immediately post cast removal.
Table 2.
Precision data for pQCT analysis of muscle cross-sectional area
| Muscle Cross-Sectional Area, cm2 |
|||||
|---|---|---|---|---|---|
| Participant Number | Measure One | Measure Two | Means | SDpooled/Means | CV%2 |
| 1 | 58.2 | 59.9 | 59.1 | 1.96 | 3.8 |
| 2 | 34.7 | 34.6 | 34.6 | 0.25 | 0.1 |
| 3 | 24.3 | 23.7 | 24.0 | 1.83 | 3.4 |
| 4 | 31.2 | 30.0 | 30.6 | 2.72 | 7.4 |
| 5 | 43.8 | 44.4 | 44.1 | 1.05 | 1.1 |
| 6 | 54.6 | 53.9 | 54.2 | 0.81 | 0.7 |
| 7 | 25.8 | 27.4 | 26.6 | 4.12 | 17.0 |
| 8 | 42.1 | 42.8 | 42.5 | 1.24 | 1.5 |
| 9 | 44.2 | 45.0 | 44.6 | 1.23 | 1.5 |
| 10 | 52.1 | 51.1 | 51.6 | 1.30 | 1.7 |
| 11 | 30.2 | 29.8 | 30.0 | 0.96 | 0.9 |
| 12 | 53.3 | 53.0 | 53.2 | 0.37 | 0.1 |
| 13 | 22.3 | 22.5 | 22.4 | 0.61 | 0.4 |
| 14 | 44.9 | 43.9 | 44.4 | 1.65 | 2.7 |
| 15 | 50.9 | 52.6 | 51.7 | 2.35 | 5.5 |
| 16 | 46.4 | 45.2 | 45.8 | 1.77 | 3.1 |
| Average | 3.2 | ||||
| CV%RMS | 1.8 | ||||
pQCT, quantitative computed tomography; CV%RMS, precision error coefficient of variation (%root mean square).
Muscle thickness.
Muscle thickness of the wrist flexor and extensor muscles during the pre- and posttesting sessions was assessed to investigate the specificity of muscle size sparing effects. Muscle thickness was assessed using ultrasound (LOGIQ e BTO8; GE Healthcare, Milwaukee, WI). Ultrasound has previously been used in our laboratory with precision errors for the left and right forearm muscles of 1.5 and 1.4%, respectively (19) and is a valid and reliable assessment of muscle thickness (6, 8, 33). The procedure involved placing a probe with transmission gel on the surface of the skin over the bulk of the muscle while the limb was at rest in neutral position. Anthropometric measures and landmarks on the arms (using nontoxic markers) and overhead transparency film were used to ensure accuracy on repeat occasions. Muscle thickness was determined by measuring a linear distance on the image of the muscle from the edge of the subcutaneous tissue to the edge of the bone. This method has been previously used in our laboratory on the wrist flexors (21); however, in the current study this method was also applied for the wrist extensors. Thickness measures were taken at a standardized location of one-third the distance between the medial epicondyle and radial styloid for the wrist flexors and one-third the distance between the lateral epicondyle and the ulna styloid for the wrist extensors. At each testing session, four measurements were recorded for each muscle, with the average of the two closest measures used for comparison.
Muscle activation.
EMG data were recorded during pre- and posttesting and during the 1st, 7th, and 12th training sessions. During pre- and posttesting, surface EMG (Grass EMG P511 AC amplifier; Grass Technologies, Middleton, WI; amplification of 1,000, bandwidth of 10 to 1,000 Hz; and VERMED NeuroPlus; 2.5 cm2, Ag/Ag chloride sensor) was used to measure muscle activity in the agonist and antagonist (wrist flexors and extensors) muscles and in the biceps brachii and triceps brachii of the trained and untrained limbs. Electrodes for the flexor carpi radialis (FCR) muscle were placed one-third of the distance from the medial epicondyle to the radial styloid following the recommendations from Buschbacher and Prahlow (5) and Zehr (55). The extensor carpi radialis (ECR) electrodes were placed on the medial side of the brachioradialis, at one-fifth (approximately three finger widths) of the distance from the lateral epicondyle on a line with the second metacarpal (55). The electrodes on the biceps brachii were placed one-third of the distance from the fossa cubit on a line between the fossa cubit and the medial acromion. The triceps brachii long head electrodes were placed at the 50% mark between the posterior crista of the acromion and the olecranon at two finger widths medial to the line as per surface electromyography for the noninvasive assessment of muscles guidelines (27). The EMG data collected in the upper arm (biceps brachii and triceps brachii) were used during testing to visually monitor muscle activation in real time only; no offline analysis of these data was conducted. Participants were instructed to relax the upper arm during contractions. During the 1st, 7th, and 12th training sessions, EMG was measured in wrist flexors of each arm and was used to determine the level of mirror activity occurring in the immobilized nontraining limb during strength training of the opposite wrist flexors.
Maximum electrically evoked contraction.
A constant current high-voltage stimulator (model DS7AH; Digitimer, Hertfordshire, UK) was used to supramaximally activate the wrist flexors and extensors during a 10% isometric MVC background contraction (34). Electrodes were manually pressed into the median nerve above the elbow, under the belly of the short head of the biceps brachii to ensure adequate stimulation of the nerve. Electrodes were also manually pressed into the radial nerve above the lateral epicondyle of the elbow for stimulation of the wrist extensors. A series of control twitches (0.5-ms pulses) were used to determine the milliamps (mA) current required to reach maximum M wave (Mmax). Stimulations started with a low current, barely detectable by the participant. The intensity was raised progressively until a plateau in the M wave occurred. The milliamps required to evoke a plateau in the peak-to-peak magnitude of the M wave plus 20% was used to ensure Mmax was reached and was recorded. The LabVIEW software was programmed to randomly administer five stimulations, and the average of the evoked tracings was used as the Mmax. M-wave data were filtered offline (MATLAB 2006b; MathWorks, Natick, MA) with a custom filter (band pass between 100 and 250 Hz) to help remove stimulation artifact. The Mmax was recorded at each testing occasion and used as a reference to normalize the within session EMG data recorded during each MVC test.
Data acquisition.
Custom software in LabVIEW (version 8.6) was used to obtain M waves from evoked contractions, and EMG and torque data during MVCs. All channels were acquired at a sampling rate of 1,000 Hz. To determine activation amplitude of the EMG data, the middle 1 s of the rectified burst activity from each voluntary contraction was used to determine the mean absolute value and the greatest amplitude recorded from the three repitions for each contraction type was used in analysis. An analog-to-digital converter (model PCI-6034E; National Instruments, Austin, TX) was used to convert the analog signals and display in LabVIEW.
Data Analysis
The study was a 2 × 2 × 2 × 3 × 2 factorial design [group (training, control) × arm (immobilized, nonimmobilized) × task (flexion, extension) × type (ECC, CON, ISO) × time (pre, posttraining)]. Strength data were analyzed with a 2 × 2 × 2 × 3 × 2 factorial ANOVA (group × arm × task × type × time) followed by further assessment of the significant two- and three-way interactions appropriate for the research questions related to the contraction type and task specificity. Strength data were also split by arm (immobilized, nonimmobilized) and task (wrist flexion, extension) for several breakdown analyses to better understand the trained limb vs. the CE sparing effect.
MCSA (via pQCT) was analyzed with a 2 × 2 × 2 (group × arm × time) factorial ANOVA, and muscle thickness (via ultrasound) was analyzed with a 2 × 2 × 2 × 2 [group × arm × muscle (flexors and extensors) × time] factorial ANOVA followed by further assessment of the significant two- and three-way interactions appropriate for the research questions related to the muscle specificity. Muscle size data were also split by arm (immobilized, nonimmobilized) and muscle (for muscle thickness; flexors, extensors) for several breakdown analyses to better understand the trained limb vs. the CE sparing effect.
EMG data were analyzed with a 2 × 2 × 2 × 2 × 3 × 2 factorial ANOVA [group × arm × muscle (agonist, antagonist) × task × type × time]. In addition, an EMG analysis of the mirror activation in the left, immobilized arm of the training group was conducted for the 1st, 7th, and 12th training sessions. The mirror activation analysis involved separate one-factor repeated-measures ANOVA to investigate differences among sessions (1, 7, and 12), repetitions (1–8), and sets (1–6).
Greenhouse-Geisser adjustments were used for violations of sphericity. Breakdown analyses followed where significant interactions were detected. Data analysis was completed using SPSS version 24. Significance was accepted at P < 0.05, and effect sizes are reported as partial eta-squared ().
RESULTS
Demographics
There were no participant dropouts during the study and maintained 100% adherence to all 12 training sessions. One participant reported having preexisting eczema in the palm of the immobilized hand, so the cast was trimmed (cut off ~1 cm at the distal end of the cast) to allow for treatment of the eczema without compromising the quality of immobilization. Groups were similar in height, weight, training experience, and handedness, P > 0.05. (Table 1).
Muscle Strength
The five-factor interaction for strength data did not reach significance, F(2,28) = 3.151, P = 0.058, = 0.184; however, significant three-way interactions were found for arm × type × task, F(2,28) = 3.447, P = 0.046, = 0.198; group × task × time, F(1,14) = 5.263, P = 0.038, = 0.273; and arm × task × time, F(1,14) = 7.027, P = 0.019, = 0.334.
The significant group × task × time interaction indicates muscle-specific CE effects to the trained homologous muscle group (i.e., flexors) but the nonspecific CE effects for contraction type. To simplify the interpretation, data were separated by task and arm and collapsed across contraction type. Significant group × time interactions were observed for the left, P = 0.049, = 0.249, and right wrist flexors, P = 0.038, = 0.272. For the right arm, wrist flexion strength changes were significantly different between the training (30.8%) and control (−7.4%) groups. For the immobilized left arm the changes for wrist flexion strength were significantly different between training (−2.4%) and control (−21.6%) groups. There were no group differences for the changes in wrist extension, but immobilized left arm extension strength significantly decreased pooled across group and contraction type (−13.2%, P < 0.001, = 0.709). See Fig. 1 and Table 3 for detailed torque changes.
Fig. 1.
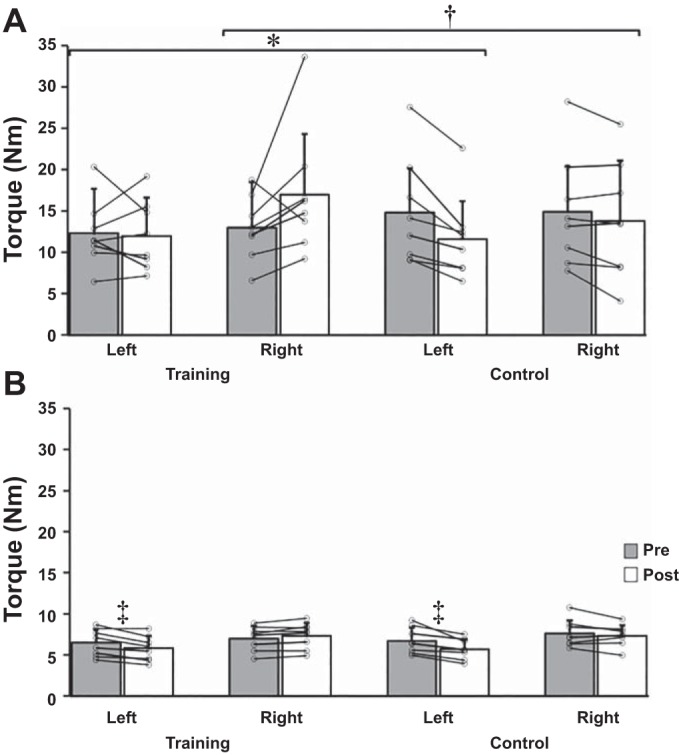
Changes in wrist flexion (A) and wrist extension torque (Nm; B) averaged across contraction types [concentric (CON), isometric (ISO), and eccentric (ECC)] from pre- to posttesting for each group. The left arm was immobilized during training. Data represent means ± SD in Nm. *P = 0.049, significant group × time interaction for left wrist flexion torque. †P = 0.038, significant group × time interaction for right wrist flexion torque. ‡P < 0.001, significant time main effect for left wrist extension torque.
Table 3.
Strength changes for contraction type and task
| Training |
Control |
|||
|---|---|---|---|---|
| Contractions (Type/Task/Arm) | Pre | Post | Pre | Post |
| ISO | ||||
| Flexion | ||||
| Left | 12.5 ± 5.3 | 12.0 ± 4.0 | 15.0 ± 6.7 | 11.2 ± 5.3 |
| Right | 12.3 ± 4.0 | 16.3 ± 7.1 | 14.2 ± 6.9 | 13.0 ± 6.1 |
| Extension | ||||
| Left | 6.2 ± 1.6 | 5.3 ± 1.6 | 6.3 ± 1.6 | 5.5 ± 1.1 |
| Right | 6.9 ± 1.6 | 7.1 ± 1.8 | 7.2 ± 1.6 | 7.1 ± 1.2 |
| CON | ||||
| Flexion | ||||
| Left | 9.4 ± 2.7 | 9.5 ± 4.1 | 11.7 ± 5.4 | 8.7 ± 4.7 |
| Right | 10.9 ± 4.2 | 13.3 ± 6.2 | 11.8 ± 5.8 | 11.1 ± 6.3 |
| Extension | ||||
| Left | 5.1 ± 1.3 | 4.5 ± 1.4 | 5.6 ± 1.1 | 4.6 ± 1.1 |
| Right | 5.5 ± 1.2 | 5.9 ± 1.1 | 6.3 ± 1.5 | 6.1 ± 1.2 |
| ECC | ||||
| Flexion | ||||
| Left | 14.9 ± 4.7 | 14.7 ± 4.7 | 17.8 ± 7.6 | 15.0 ± 5.5 |
| Right | 15.7 ± 4.4 | 21.3 ± 9.6 | 18.7 ± 7.9 | 17.4 ± 9.1 |
| Extension | ||||
| Left | 8.1 ± 2.4 | 7.5 ± 2.1 | 8.3 ± 2.2 | 7.1 ± 1.8 |
| Right | 8.7 ± 1.8 | 9.1 ± 2.0 | 9.3 ± 1.9 | 8.9 ± 1.9 |
| Pooled | ||||
| Flexion | ||||
| Left | 12.3 ± 5.4 | 12.0 ± 4.6 | 14.8 ± 5.4 | 11.6 ± 4.6 |
| Right | 13.0 ± 5.5 | 17.0 ± 7.3 | 14.9 ± 5.5 | 13.8 ± 7.3 |
| Extension | ||||
| Left | 6.5 ± 1.6 | 5.8 ± 1.5 | 6.7 ± 1.6 | 5.7 ± 1.2 |
| Right | 7.0 ± 1.5 | 7.3 ± 1.6 | 7.6 ± 1.6 | 7.3 ± 1.3 |
| Pooled | ||||
| Left | 9.4 ± 3.3 | 8.9 ± 2.9 | 10.8 ± 3.3 | 8.7 ± 2.9 |
| Right | 10.0 ± 3.5 | 12.2 ± 4.3 | 11.3 ± 3.5 | 10.6 ± 4.3 |
Data are means ± SD in Nm. There were no significant differences between contraction type [concentric (CON), isometric (ISO), and eccentric (ECC)]. Pooled data revealed significant differences as shown in Fig. 1.
Muscle Cross-Sectional Area
A three-factor ANOVA revealed a significant interaction for group × arm × time, F(1,14) = 7.328, P = 0.017, = 0.344. To breakdown the three-way interaction, data were split by arm and separate group × time ANOVA tests were run. A significant group × time interaction was detected for the left, immobilized arm only, P = 0.012, = 0.375, indicating that the change in MCSA for the left arm for the training group (1.3%) was different from the change in the control group (−2.3%).
Further analysis involved splitting data by group and running separate arm × time ANOVA tests to compare between arm differences. These analyses revealed a significant arm × time interaction for the control group only, P = 0.019, = 0.566, indicating that the change in the left arm (−2.3%) was different from the change in the right arm (1.2%). For the training group, the change in the left, immobilized arm (1.3%) was not significantly different from the change in the right, trained arm (1.2%) (Fig. 2 and Table 4).
Fig. 2.
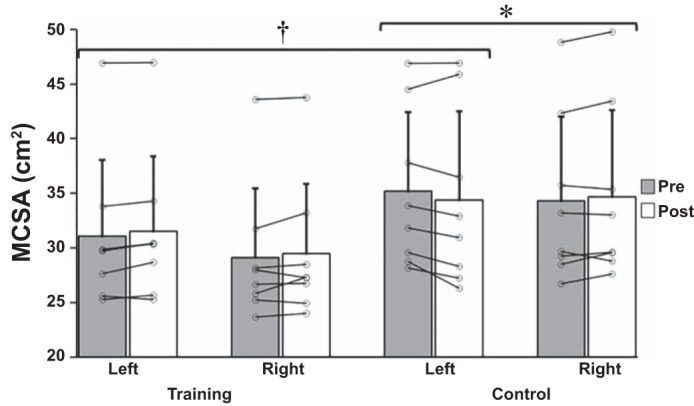
Muscle cross-sectional area (MCSA; cm2) changes for the left, immobilized and right arms of the training and control groups from pre- to posttesting. Data represent means ± SD in cm2. *P = 0.019, significant arm × time interaction for control group only. †P = 0.012, significant group × time interaction for the left arm only.
Table 4.
Muscle size changes
| Group/Arm/Measure | Pre | Post |
|---|---|---|
| Training | ||
| Left | ||
| Flexor MT | 3.27 ± 0.46 | 3.36 ± 0.56 |
| Extensor MT | 1.60 ± 0.27 | 1.68 ± 0.28 |
| MCSA | 31.10 ± 6.95 | 31.52 ± 6.87 |
| Right | ||
| Flexor MT | 3.53 ± 0.55 | 3.78 ± 0.51 |
| Extensor MT | 1.65 ± 0.33 | 1.69 ± 0.29 |
| MCSA | 29.13 ± 6.32 | 29.48 ± 6.40 |
| Control | ||
| Left | ||
| Flexor MT | 3.75 ± 0.73 | 3.63 ± 0.68 |
| Extensor MT | 1.69 ± 0.20 | 1.64 ± 0.25 |
| MCSA | 35.18 ± 7.23 | 34.38 ± 8.12 |
| Right | ||
| Flexor MT | 3.74 ± 0.62 | 3.60 ± 0.53 |
| Extensor MT | 1.68 ± 0.14 | 1.72 ± 0.14 |
| MCSA | 34.29 ± 7.71 | 34.66 ± 7.96 |
Data are means ± SD. MCSA, muscle cross-sectional area in cm2; MT, muscle thickness in cm.
Muscle Thickness
Normality assessments of the control group determined that the left flexor muscle thickness raw change was skewed (Zskew = 2.75) and kurtotic (Zkurt = 3.13). One participant was removed from the analysis because it was determined that a positive change in muscle thickness of 0.68 cm (20%) in the immobilized limb of a control participant was a result of measurement error. After the removal of the outlier, there were no violations of normality as determined by skewness (Zskew = 1.14) and kurtosis (Zkurt = 0.59).
A four-factor omnibus factorial ANOVA revealed a significant interaction for group × arm × muscle × time, F(1,13) = 6.037, P = 0.029, = 0.317. To breakdown the four-way interaction, data were split by arm and by muscle, and separate group × time ANOVA tests were run. Significant group × time interactions were found for the right, P = 0.026, = 0.327, and left wrist flexors, P = 0.011, = 0.405, but not for wrist extensor muscles of either limb.
Data were then split by group, and separate arm × muscle × time ANOVA tests were run. A significant arm × muscle × time interaction was detected for the training group only, P = 0.006, = 0.684. Further breakdown of the three-way interaction for the training group involved splitting these data by muscle (flexors, extensors), but this did not reveal any arm × time interactions.
The ANOVA tests for muscle thickness revealed that the change in the untrained, immobilized left flexors of the training group (2.8%) was significantly different from control (−3.2%). The tests also revealed the change in the right flexors of the training group (7.1%) was different from the control group (−3.7%) (Fig. 3 and Table 4).
Fig. 3.
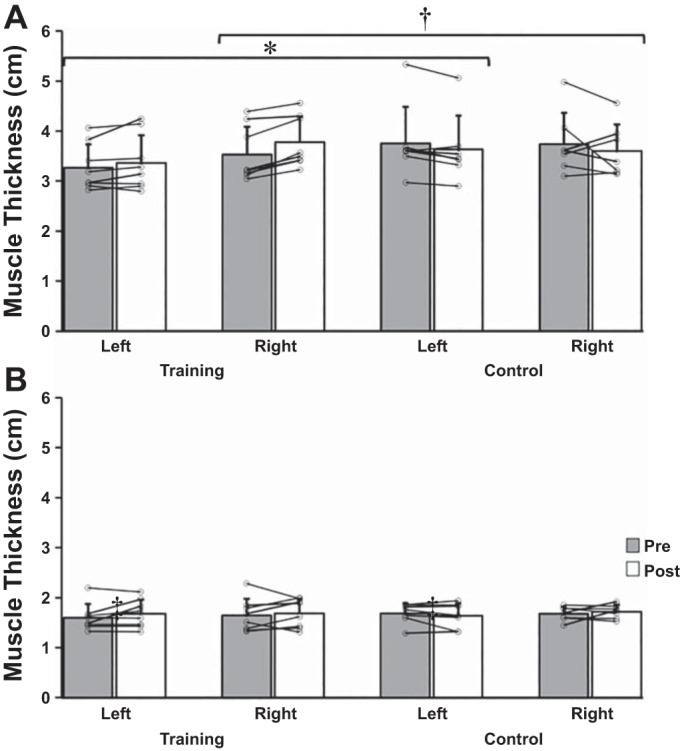
Changes in muscle thickness for wrist flexors (A) and wrist extensors (B) between groups. Note the observed significant effects only for the wrist flexors. Data represent means ± SD in cm. *P = 0.011, significant group × time interaction for the left wrist flexors. †P = 0.026, significant group × time interaction for the right wrist flexors.
Muscle Activation
The six-factor omnibus factorial ANOVA failed to reach significance, F(2,28) = 0.306, P = 0.739, = 0.021. A significant interaction was observed for group × muscle × type × task × time, P = 0.004, = 0.331.
To breakdown the significant interaction, data were split by group and separate muscle × task × type × time ANOVA tests were run. No significant interactions were observed that included a time factor. Further breakdown involved splitting these data by arm and examining between group differences. No significant interactions including group or time factors were observed for the immobilized and nonimmobilized limb (Table 5, Flexion, and Table 6, Extension).
Table 5.
Flexion EMG changes
| Training |
Control |
|||
|---|---|---|---|---|
| Flexion Contractions (Type/Muscle/Arm) | Pre | Post | Pre | Post |
| ISO | ||||
| Agonist (FCR) | ||||
| Left | 0.214 ± 0.111 | 0.159 ± 0.080 | 0.210 ± 0.098 | 0.154 ± 0.066 |
| Right | 0.197 ± 0.088 | 0.273 ± 0.182 | 0.200 ± 0.099 | 0.215 ± 0.110 |
| Antagonist (ECR) | ||||
| Left | 0.042 ± 0.021 | 0.049 ± 0.025 | 0.076 ± 0.040 | 0.037 ± 0.027 |
| Right | 0.037 ± 0.018 | 0.049 ± 0.016 | 0.062 ± 0.028 | 0.047 ± 0.028 |
| CON | ||||
| Agonist (FCR) | ||||
| Left | 0.188 ± 0.075 | 0.143 ± 0.100 | 0.181 ± 0.082 | 0.128 ± 0.057 |
| Right | 0.192 ± 0.085 | 0.214 ± 0.108 | 0.189 ± 0.096 | 0.184 ± 0.109 |
| Antagonist (ECR) | ||||
| Left | 0.034 ± 0.022 | 0.037 ± 0.029 | 0.057 ± 0.032 | 0.024 ± 0.016 |
| Right | 0.029 ± 0.015 | 0.039 ± 0.013 | 0.049 ± 0.019 | 0.038 ± 0.029 |
| ECC | ||||
| Agonist (FCR) | ||||
| Left | 0.245 ± 0.114 | 0.188 ± 0.105 | 0.210 ± 0.090 | 0.177 ± 0.054 |
| Right | 0.249 ± 0.110 | 0.266 ± 0.193 | 0.200 ± 0.088 | 0.216 ± 0.099 |
| Antagonist (ECR) | ||||
| Left | 0.032 ± 0.012 | 0.057 ± 0.046 | 0.061 ± 0.036 | 0.029 ± 0.016 |
| Right | 0.030 ± 0.011 | 0.047 ± 0.016 | 0.049 ± 0.017 | 0.037 ± 0.024 |
| Pooled | ||||
| Agonist (FCR) | ||||
| Left | 0.216 ± 0.140 | 0.164 ± 0.090 | 0.201 ± 0.124 | 0.153 ± 0.054 |
| Right | 0.213 ± 0.128 | 0.251 ± 0.159 | 0.196 ± 0.132 | 0.205 ± 0.104 |
| Antagonist (ECR) | ||||
| Left | 0.036 ± 0.024 | 0.047 ± 0.034 | 0.064 ± 0.052 | 0.030 ± 0.020 |
| Right | 0.032 ± 0.020 | 0.045 ± 0.014 | 0.053 ± 0.028 | 0.041 ± 0.025 |
Data represent means ± SD in normalized units. FCR, flexor carpi radialis; ECR, extensor carpi radialis. There were no significant differences between groups regardless of muscle (flexors, extensors), task (flexion, extension) or contraction type (ECC, CON, ISO).
Table 6.
Extension EMG changes
| Training |
Control |
|||
|---|---|---|---|---|
| Extension Contractions (Type/Muscle/Arm) | Pre | Post | Pre | Post |
| ISO | ||||
| Agonist (ECR) | ||||
| Left | 0.124 ± 0.025 | 0.118 ± 0.060 | 0.175 ± 0.065 | 0.128 ± 0.052 |
| Right | 0.118 ± 0.038 | 0.108 ± 0.041 | 0.160 ± 0.059 | 0.135 ± 0.033 |
| Antagonist (FCR) | ||||
| Left | 0.031 ± 0.021 | 0.036 ± 0.038 | 0.064 ± 0.065 | 0.017 ± 0.011 |
| Right | 0.032 ± 0.020 | 0.051 ± 0.034 | 0.043 ± 0.029 | 0.033 ± 0.030 |
| CON | ||||
| Agonist (ECR) | ||||
| Left | 0.118 ± 0.028 | 0.097 ± 0.033 | 0.182 ± 0.074 | 0.118 ± 0.035 |
| Right | 0.121 ± 0.033 | 0.106 ± 0.041 | 0.150 ± 0.044 | 0.128 ± 0.029 |
| Antagonist (FCR) | ||||
| Left | 0.034 ± 0.022 | 0.044 ± 0.043 | 0.053 ± 0.046 | 0.019 ± 0.009 |
| Right | 0.030 ± 0.020 | 0.036 ± 0.026 | 0.032 ± 0.018 | 0.032 ± 0.027 |
| ECC | ||||
| Agonist (ECR) | ||||
| Left | 0.132 ± 0.023 | 0.118 ± 0.070 | 0.165 ± 0.069 | 0.123 ± 0.042 |
| Right | 0.124 ± 0.037 | 0.092 ± 0.031 | 0.151 ± 0.053 | 0.130 ± 0.042 |
| Antagonist (FCR) | ||||
| Left | 0.037 ± 0.024 | 0.043 ± 0.032 | 0.061 ± 0.047 | 0.024 ± 0.021 |
| Right | 0.037 ± 0.020 | 0.054 ± 0.045 | 0.040 ± 0.018 | 0.041 ± 0.029 |
| Pooled | ||||
| Agonist (ECR) | ||||
| Left | 0.124 ± 0.032 | 0.111 ± 0.048 | 0.174 ± 0.096 | 0.123 ± 0.042 |
| Right | 0.121 ± 0.048 | 0.102 ± 0.034 | 0.153 ± 0.072 | 0.131 ± 0.034 |
| Antagonist (FCR) | ||||
| Left | 0.034 ± 0.032 | 0.041 ± 0.034 | 0.059 ± 0.076 | 0.020 ± 0.014 |
| Right | 0.033 ± 0.028 | 0.047 ± 0.034 | 0.038 ± 0.028 | 0.035 ± 0.028 |
Data represent means ± SD in normalized units. FCR, flexor carpi radialis; ECR, extensor carpi radialis. There were no significant differences between groups regardless of muscle (flexors, extensors), task (flexion, extension) or contraction type (ECC, CON, ISO).
M-wave data were stable between pre- and posttesting sessions for the left wrist flexors (pre: 1.49 ± 1.03 mV; post: 1.57 ± 0.99 mV), left wrist extensors (pre: 1.65 ± 1.07 mV; post: 1.60 ± 0.73 mV), right wrist flexors (pre: 1.60 ± 0.96 mV; post: 1.49 ± 1.16 mV), and right wrist extensors (pre: 1.90 ± 0.95 mV; post: 2.04 ± 0.89 mV).
Mirror Activity (Coactivation of the Nontraining Limb)
The mean EMG activity of the left, immobilized, wrist flexors of the training group measured during the 1st, 7th and 12th training sessions was, on average, 5.6% of pretesting isometric MVC. The mirror EMG activity (normalized to baseline isometric MVC) was not significantly different among sessions 1, 7, and 12 (range: 0.047 ± 0.017 to 0.085 ± 0.046), P = 0.077, = 0.360, among repititions 1 to 8 (range: 0.051 ± 0.021 to 0.061 ± 0.024), P = 0.553, = 0.108, or among sets 1 to 6 (range: 0.047 ± 0.026 to 0.061 ± 0.022), P = 0.142, = 0.245, during training sessions.
DISCUSSION
The major finding of this study is that the CE sparing effects of muscle strength were specific to the trained homologous muscle in the contralateral limb but were not specific to contraction type. ECC wrist flexion training of the nonimmobilized limb preserved ECC, CON, and ISO strength in the contralateral, immobilized wrist flexors. Importantly, this was the first study to identify muscle size sparing effects of CE with muscle area measures via pQCT. The current study has therefore replicated previously observed sparing effects of CE with a robust measure of muscle size (26). The confirmation of muscle size and strength sparing effects across multiple measures builds confidence in the reproducibility of CE sparing effects and their clinical relevance for improving recovery from unilateral injury or impairment.
Muscle Strength
In prior CE sparing studies, the average trained limb strength improvement was ~24%, accompanied by preservation of strength in the untrained, immobilized limb. Note that the average decrease in immobilized limb strength for nontraining control groups in prior CE sparing studies averaged ~12% across studies; therefore, the CE sparing effect can be estimated as about half of the trained limb effect (20, 21, 40, 45). As previously mentioned, the magnitude of CE of strength is typically ~50% of the trained limb effect in studies not involving immobilization models (7). The current data merge well with these prior studies, which, on average, report that the immobilized limb of the training group shows no change in strength (0.3%, NS) and a significant decline of 13.2% for the immobilized limb of the nontraining control group (20, 21, 40, 45). The combined evidence to date suggests the strength sparing effect of CE amounts to ~12% for arm immobilization protocols of a 3- to 4-wk duration (20, 21, 40, 45).
Previous research has found CE to be highly specific, where transfer effects reside primarily in the contralateral homologous muscle group, with the same velocity or joint angle used in training of the opposite limb (18, 50, 53). A novel aspect of the current study was the intent to investigate muscle size and strength in the wrist flexor and extensor muscles of the trained and contralateral, immobilized arm. CE attenuated strength loss in the homologous immobilized wrist flexors, which is congruent with muscle specificity of CE effects in studies without immobilization and indicates that the mechanisms of CE specificity are likely unaltered in the presence of immobilization.
Unilateral training with ECC muscle actions results in greater CE of strength in the contralateral homologous muscle compared with CON or ISO muscle actions (18, 29). Hortobágyi et al. (29) observed a greater global training effect when training with ECC muscle actions compared with CON, meaning that both CON and ISO strength improved, albeit to a lesser extent, in the contralateral homologous muscle from the ECC training. In the present study, participants in the training group attended three sessions per week of isolated ECC wrist flexion actions, which resulted in a similar increase in strength of the right wrist flexors across all contraction types (Fig. 1). Additionally, a preservation of strength regardless of contraction type was apparent in the left, immobilized wrist flexors. ECC muscle actions compared with CON and ISO are known to cause greater increases in intracortical facilitation and larger decreases in intracortical inhibition (31, 32), which could explain larger transfer effects with ECC, and may contribute to the observed global strength sparing effect in the contralateral homologous muscle group in the current study.
Muscle Size
Before this study, the muscle size sparing effect was only observed using ultrasound measures of muscle thickness (20, 40, 45). The advantage of concurrent pQCT and ultrasound imaging of muscle is the ability to compare whole MCSA and site-specific muscle thickness (i.e., forearm flexors and extensors). Verifying muscle size sparing effects with a measure of MCSA is critical for progression in the field (26). The CE sparing effects for muscle thickness revealed support for muscle-specific effects to contralateral homologous wrist flexors (Fig. 3); however, immobilization did not result in significant decreases in muscle thickness for the extensors of either group. Therefore, the site-specific nature of muscle size sparing effects for flexors vs. extensors is less convincing than for strength. Further supporting the preservation of muscle thickness was the confirmation of muscle size preservation with pQCT-derived MCSA (Fig. 2). Since the pQCT MCSA method cannot differentiate flexor vs. extensor muscles, and there were no detectable changes in extensor muscle thickness, more work is needed to confirm site-specific muscle size sparing of CE.
This study provides novel insight into possible mechanisms of CE sparing. Currently, the dominant theories of CE effects have proposed possible mechanisms for strength and skill transfer effects to reside primarily in the brain (49). CE is thought to be a neural phenomenon where changes in cortical processes and motor engrams positively impact the neural drive to the contralateral limb (17, 49). This is understandable since CE effects typically do not present with evidence of alterations in muscle volume (16, 36, 49), especially without immobilization. There is no apparent candidate mechanism that accounts for muscle size changes with CE in the untrained limb, unless there is concurrent evidence of direct voluntary or involuntary muscle activation in the nontraining limb of at least 15% of maximum, which has been shown to induce small amounts of hypertrophy after 12 wk of training (28). Our findings suggest activation of the resting immobilized limb during training (~5.6% MVC) over a 4-wk period is not likely to preserve the muscle, although it cannot be ruled out. This presents the possibility that an alternate mechanism is involved in CE sparing effects, which may be independently activated or driven by the nervous system. It at least suggests the preservation of muscle strength and size via CE are related but could be driven by overlapping and independent mechanisms.
The regulation of muscle atrophy with immobilization or disuse occurs through two primary processes; elevated muscle protein breakdown and a decrease in muscle protein synthesis (MPS), with a decrease in MPS found to be the leading mechanism (46). While the mechanisms of muscle size preservation with CE are currently unknown, and a direct connection between neural contributions and the regulation of MPS and muscle protein breakdown is not clear, it remains possible that the unilateral ECC training of the wrist flexors in the present study led to muscle size preservation in the contralateral limb by influencing the balance of protein regulation. One possible mechanism may be in the neural regulation of the protein kinase B (AKT) and mammalian target of rapamycin (mTOR) pathway (26). AKT and mTOR are important protein complexes that play a role in the modulation of gene expression, cell development, growth, and survival and are upregulated in the nervous system during cellular stress (9, 41). The AKT and mTOR pathway is upregulated with skeletal muscle hypertrophy and downregulated with muscle atrophy caused by disuse (2). Investigating AKT and mTOR pathway modulation with CE to an immobilized limb is an intriguing prospect in understanding the muscle size sparing effects observed.
Muscle Activation and Mirror Activity
In the present study, the EMG data did not reveal significant between groups differences for any of the tested contraction types or tasks. The EMG data are inconclusive and further investigation with a larger sample size is needed. Muscle activation at levels as low as 10% one repetition maximum have been shown to increase strength (35). Farthing et al. (21) proposed that, although unlikely, it is possible for the CE sparing effects observed in past literature to be attributed to high levels (>10% MVC) of mirror activity during the unilateral training intervention in the opposite limb, because mirror activity was not monitored under the cast. Of the four studies to investigate the CE sparing effects in healthy participants, only Magnus et al. (40) monitored mirror activity during a sling model of disuse. Both Magnus et al. (40) and the current data show CE training produced low levels of mirror activity in the immobilized limb. Although unlikely, it remains possible that the reported 5.6% in the current study and the 3.1% (biceps brachii) and 6.1% (triceps brachii) reported in the Magnus et al. (40) study contributed at least some to the observed sparing effects of size and strength in the immobilized limb. The contribution of mirror activity to size and/or strength sparing effects of CE remains unclear, but the current hypothesis that cortical contributions are primarily responsible for the transfer effects remains viable (49).
Clinical Relevance
The findings of this study have broad clinical implications for rehabilitation from unilateral injury or impairment. The data suggest that training of the nonimmobilized, or noninjured limb, can benefit an opposite homologous immobilized or injured limb through CE effects. Although there is already published evidence that CE can benefit recovery from wrist fracture (39) and facilitate affected limb strength in chronic stroke patients (11), the current finding of muscle-specific sparing effects in healthy volunteers suggests that targeted training involving multiple muscles is needed to attenuate the loss in size and strength of an opposite limb in a clinical setting. Furthermore, the global sparing of strength across multiple contraction types after training only ECC suggests that more efficient strength rehabilitation programs can be administered. Rehabilitation should focus on complete joint symmetry by training both agonist and antagonistic pairs of both distal and proximal muscle groups. Taken together, the current literature demonstrates a clear potential benefit of CE for improving recovery from unilateral immobilization or injury (20, 21, 39, 40, 45).
Study Strengths and Limitations
The study began in September (late summer) and ran through the fall and winter semesters at the University of Saskatchewan. The strategic timing of participant recruitment is a likely cause for the 100% adherence to all 12 training sessions without any participant dropouts. Another strength to this study was that the MCSA data were collected by a researcher who was blinded to group assignment, which increases confidence in the outcome.
While mirror activation in the immobilized limb was monitored during three testing sessions, the muscle activity was only monitored in the wrist flexors. The lack of wrist extensor monitoring is a noted limitation to this study. Another limitation was the use of the wrist joint model for investigating specificity of CE sparing effects. Although the wrist is clinically relevant (39) and convenient for cast immobilization, there are several muscles that make up the wrist flexors and extensors, making EMG recording and muscle imaging of these muscles difficult to conduct. Although we detected significant effects for several of our main outcomes, the sample size was small for the complexity of the design. Recruiting participants for immobilization studies is difficult; therefore, replication of these findings with larger samples is important.
Although the 4-wk immobilization period was longer than most of the previous CE sparing literature (20, 21, 45), a longer immobilization period would have been beneficial to increase the severity of strength loss and atrophy in the immobilized limb. This would provide a better estimate of clinical studies of unilateral wrist fractures, which demonstrate much more profound losses in strength and function after 4–6 wk of immobilization (39). Immobilization due to unilateral injury may be prescribed for durations longer than 4 wk, and to improve the clinical relevance of these CE sparing effects, longer immobilization periods are recommended for future research. Future research would benefit from using MRI to assess specific muscle volume adaptations to determine the specificity of the sparing effects. Future research would also benefit from the use of functional imaging or brain stimulation to investigate the specificity of CE sparing effects. Using functional imaging or brain stimulation has the potential to offer clear evidence of the neural mechanisms contributing to the observed sparing effects of strength and size. As mentioned, future research investigating the specificity of CE may benefit by recording EMG of both the contralateral agonist (wrist flexors) and antagonist (wrist extensors) muscles during training sessions to more closely monitor the contributions of mirror activity in the resting, immobilized limb. Finally, the sparing effects of CE have not yet been shown in an older population, and the effects may not be the same as shown here for young, healthy volunteers.
Conclusion
This study provides novel insight into the specificity of CE sparing effects in an immobilized limb. The finding that immobilized limb strength was preserved across contraction types for the contralateral homologous muscle after only training with ECC muscle actions confirms that immobilization does not alter the specificity effects previously reported in nonimmobilization CE studies. There were no direct measures of cortical or corticospinal activation changes or altered excitability in the present study; therefore, no definitive conclusions can be made from these data to support neural mechanisms (49). However, the data suggest the possibility of an alternate site-specific mechanism of CE sparing that can somehow offset the effects of disuse in a homologous muscle. Although the cause of the muscle size sparing remains unclear, its confirmation brings new insight into possible contributing mechanisms of CE sparing effects. Regardless of the mechanisms, CE appears to be a relevant and practical exercise modality to attenuating the loss commonly associated with immobilization and is viable for consideration in clinical settings such as unilateral orthopedic or neurological injury (14, 39).
GRANTS
This research was funded by the National Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant RGPIN 2016 - 0529. Justin Andrushko received a M.Sc. scholarship from the Canadian Institutes of Health Research (CIHR) to carry out this research.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
J.W.A. and J.P.F. conceived and designed research; J.W.A. and K.M.B. performed experiments; J.W.A. analyzed data; J.W.A., J.L.L., S.A.K., and J.P.F. interpreted results of experiments; J.W.A. prepared figures; J.W.A. drafted manuscript; J.W.A., J.L.L., K.M.B., S.A.K., and J.P.F. edited and revised manuscript; J.W.A., J.L.L., K.M.B., S.A.K., and J.P.F. approved final version of manuscript.
ACKNOWLEDGMENTS
We acknowledge Chris Buttinger for the contribution of applying the casts throughout the study and all of the participants that were adherent and dedicated to making this study a success. We also thank Dr. Tibor Hortobágyi for providing helpful comments on an earlier version of the manuscript.
REFERENCES
- 1.Atherton PJ, Greenhaff PL, Phillips SM, Bodine SC, Adams CM, Lang CH. Control of skeletal muscle atrophy in response to disuse: clinical/preclinical contentions and fallacies of evidence. Am J Physiol Endocrinol Metab 311: E594–E604, 2016. doi: 10.1152/ajpendo.00257.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bodine SC, Stitt TN, Gonzalez M, Kline WO, Stover GL, Bauerlein R, Zlotchenko E, Scrimgeour A, Lawrence JC, Glass DJ, Yancopoulos GD. Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nat Cell Biol 3: 1014–1019, 2001. doi: 10.1038/ncb1101-1014. [DOI] [PubMed] [Google Scholar]
- 3.Bryden MP. Measuring handedness with questionnaires. Neuropsychologia 15: 617–624, 1977. doi: 10.1016/0028-3932(77)90067-7. [DOI] [PubMed] [Google Scholar]
- 4.Burianová H, Sowman PF, Marstaller L, Rich AN, Williams MA, Savage G, Al-Janabi S, de Lissa P, Johnson BW. Adaptive motor imagery: a multimodal study of immobilization-induced brain plasticity. Cereb Cortex 26: 1072–1080, 2016. doi: 10.1093/cercor/bhu287. [DOI] [PubMed] [Google Scholar]
- 5.Buschbacher RM, Prahlow ND. Manual of nerve conduction studies. J Clin Neuromuscul Dis 2: 120, 2000. doi: 10.1097/00131402-200012000-00015. [DOI] [Google Scholar]
- 6.Candow DG, Chilibeck PD, Facci M, Abeysekara S, Zello GA. Protein supplementation before and after resistance training in older men. Eur J Appl Physiol 97: 548–556, 2006. doi: 10.1007/s00421-006-0223-8. [DOI] [PubMed] [Google Scholar]
- 7.Carroll TJ, Herbert RD, Munn J, Lee M, Gandevia SC. Contralateral effects of unilateral strength training: evidence and possible mechanisms. J Appl Physiol (1985) 101: 1514–1522, 2006. doi: 10.1152/japplphysiol.00531.2006. [DOI] [PubMed] [Google Scholar]
- 8.Cartwright MS, Demar S, Griffin LP, Balakrishnan N, Harris JM, Walker FO. Validity and reliability of nerve and muscle ultrasound. Muscle Nerve 47: 515–521, 2013. doi: 10.1002/mus.23621. [DOI] [PubMed] [Google Scholar]
- 9.Chong ZZ, Li F, Maiese K. Oxidative stress in the brain: novel cellular targets that govern survival during neurodegenerative disease. Prog Neurobiol 75: 207–246, 2005. doi: 10.1016/j.pneurobio.2005.02.004. [DOI] [PubMed] [Google Scholar]
- 10.Clark BC, Issac LC, Lane JL, Damron LA, Hoffman RL. Neuromuscular plasticity during and following 3 wk of human forearm cast immobilization. J Appl Physiol (1985) 105: 868–878, 2008. doi: 10.1152/japplphysiol.90530.2008. [DOI] [PubMed] [Google Scholar]
- 11.Dragert K, Zehr EP. High-intensity unilateral dorsiflexor resistance training results in bilateral neuromuscular plasticity after stroke. Exp Brain Res 225: 93–104, 2013. doi: 10.1007/s00221-012-3351-x. [DOI] [PubMed] [Google Scholar]
- 12.Duchateau J, Hainaut K. Effects of immobilization on contractile properties, recruitment and firing rates of human motor units. J Physiol 422: 55–65, 1990. doi: 10.1113/jphysiol.1990.sp017972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Eastlack ME, Okereke E, Esterhai JL, Abdus S, Kelley M, Vanderborne K. Muscle changes during casting and therapy. Phys Ther 79: pS64, 1999. [Google Scholar]
- 14.Ehrensberger M, Simpson D, Broderick P, Monaghan K. Cross-education of strength has a positive impact on post-stroke rehabilitation: a systematic literature review. Top Stroke Rehabil 23: 126–135, 2016. doi: 10.1080/10749357.2015.1112062. [DOI] [PubMed] [Google Scholar]
- 15.Facchini S, Romani M, Tinazzi M, Aglioti SM. Time-related changes of excitability of the human motor system contingent upon immobilisation of the ring and little fingers. Clin Neurophysiol 113: 367–375, 2002. doi: 10.1016/S1388-2457(02)00009-3. [DOI] [PubMed] [Google Scholar]
- 16.Farthing JP. Cross-education of strength depends on limb dominance: implications for theory and application. Exerc Sport Sci Rev 37: 179–187, 2009. [DOI] [PubMed] [Google Scholar]
- 17.Farthing JP, Borowsky R, Chilibeck PD, Binsted G, Sarty GE. Neuro-physiological adaptations associated with cross-education of strength. Brain Topogr 20: 77–88, 2007. doi: 10.1007/s10548-007-0033-2. [DOI] [PubMed] [Google Scholar]
- 18.Farthing JP, Chilibeck PD. The effect of eccentric training at different velocities on cross-education. Eur J Appl Physiol 89: 570–577, 2003. doi: 10.1007/s00421-003-0841-3. [DOI] [PubMed] [Google Scholar]
- 19.Farthing JP, Chilibeck PD, Binsted G. Cross-education of arm muscular strength is unidirectional in right-handed individuals. Med Sci Sports Exerc 37: 1594–1600, 2005. doi: 10.1249/01.mss.0000177588.74448.75. [DOI] [PubMed] [Google Scholar]
- 20.Farthing JP, Krentz JR, Magnus CR. Strength training the free limb attenuates strength loss during unilateral immobilization. J Appl Physiol (1985) 106: 830–836, 2009. doi: 10.1152/japplphysiol.91331.2008. [DOI] [PubMed] [Google Scholar]
- 21.Farthing JP, Krentz JR, Magnus CR, Barss TS, Lanovaz JL, Cummine J, Esopenko C, Sarty GE, Borowsky R. Changes in functional magnetic resonance imaging cortical activation with cross education to an immobilized limb. Med Sci Sports Exerc 43: 1394–1405, 2011. doi: 10.1249/MSS.0b013e318210783c. [DOI] [PubMed] [Google Scholar]
- 22.Farthing JP, Zehr EP. Restoring symmetry: clinical applications of cross-education. Exerc Sport Sci Rev 42: 70–75, 2014. doi: 10.1249/JES.0000000000000009. [DOI] [PubMed] [Google Scholar]
- 23.Faul F, Erdfelder E, Lang A-G, Buchner A. G*Power 3: a flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav Res Methods 39: 175–191, 2007. doi: 10.3758/BF03193146. [DOI] [PubMed] [Google Scholar]
- 24.Folland JP, Williams AG. The adaptations to strength training : morphological and neurological contributions to increased strength. Sports Med 37: 145–168, 2007. doi: 10.2165/00007256-200737020-00004. [DOI] [PubMed] [Google Scholar]
- 25.Frank-Wilson AW, Johnston JD, Olszynski WP, Kontulainen SA. Measurement of muscle and fat in postmenopausal women: precision of previously reported pQCT imaging methods. Bone 75: 49–54, 2015. doi: 10.1016/j.bone.2015.01.016. [DOI] [PubMed] [Google Scholar]
- 26.Hendy AM, Lamon S. The cross-education phenomenon: brain and beyond. Front Physiol 8: 297, 2017. doi: 10.3389/fphys.2017.00297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hermens HJ, Freriks B, Disselhorst-Klug C, Rau G. Development of recommendations for SEMG sensors and sensor placement procedures. J Electromyogr Kinesiol 10: 361–374, 2000. doi: 10.1016/S1050-6411(00)00027-4. [DOI] [PubMed] [Google Scholar]
- 28.Holm L, Reitelseder S, Pedersen TG, Doessing S, Petersen SG, Flyvbjerg A, Andersen JL, Aagaard P, Kjaer M. Changes in muscle size and MHC composition in response to resistance exercise with heavy and light loading intensity. J Appl Physiol (1985) 105: 1454–1461, 2008. doi: 10.1152/japplphysiol.90538.2008. [DOI] [PubMed] [Google Scholar]
- 29.Hortobágyi T, Lambert NJ, Hill JP. Greater cross education following training with muscle lengthening than shortening. Med Sci Sports Exerc 29: 107–112, 1997. doi: 10.1097/00005768-199701000-00015. [DOI] [PubMed] [Google Scholar]
- 30.Hortobágyi T, Richardson SP, Lomarev M, Shamim E, Meunier S, Russman H, Dang N, Hallett M. Interhemispheric plasticity in humans. Med Sci Sports Exerc 43: 1188–1199, 2011. doi: 10.1249/MSS.0b013e31820a94b8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Howatson G, Taylor MB, Rider P, Motawar BR, McNally MP, Solnik S, DeVita P, Hortobágyi T. Ipsilateral motor cortical responses to TMS during lengthening and shortening of the contralateral wrist flexors. Eur J Neurosci 33: 978–990, 2011. doi: 10.1111/j.1460-9568.2010.07567.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kidgell DJ, Frazer AK, Daly RM, Rantalainen T, Ruotsalainen I, Ahtiainen J, Avela J, Howatson G. Increased cross-education of muscle strength and reduced corticospinal inhibition following eccentric strength training. Neuroscience 300: 566–575, 2015. doi: 10.1016/j.neuroscience.2015.05.057. [DOI] [PubMed] [Google Scholar]
- 33.Krentz JR, Quest B, Farthing JP, Quest DW, Chilibeck PD. The effects of ibuprofen on muscle hypertrophy, strength, and soreness during resistance training. Appl Physiol Nutr Metab 33: 470–475, 2008. doi: 10.1139/H08-019. [DOI] [PubMed] [Google Scholar]
- 34.Lagerquist O, Zehr EP, Docherty D. Increased spinal reflex excitability is not associated with neural plasticity underlying the cross-education effect. J Appl Physiol (1985) 100: 83–90, 2006. doi: 10.1152/japplphysiol.00533.2005. [DOI] [PubMed] [Google Scholar]
- 35.Laidlaw DH, Kornatz KW, Keen DA, Suzuki S, Enoka RM. Strength training improves the steadiness of slow lengthening contractions performed by old adults. J Appl Physiol (1985) 87: 1786–1795, 1999. doi: 10.1152/jappl.1999.87.5.1786. [DOI] [PubMed] [Google Scholar]
- 36.Lee M, Carroll TJ. Cross education: possible mechanisms for the contralateral effects of unilateral resistance training. Sports Med 37: 1–14, 2007. doi: 10.2165/00007256-200737010-00001. [DOI] [PubMed] [Google Scholar]
- 37.Lee M, Hinder MR, Gandevia SC, Carroll TJ. The ipsilateral motor cortex contributes to cross-limb transfer of performance gains after ballistic motor practice. J Physiol 588: 201–212, 2010. doi: 10.1113/jphysiol.2009.183855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Lissek S, Wilimzig C, Stude P, Pleger B, Kalisch T, Maier C, Peters SA, Nicolas V, Tegenthoff M, Dinse HR. Immobilization impairs tactile perception and shrinks somatosensory cortical maps. Curr Biol 19: 837–842, 2009. doi: 10.1016/j.cub.2009.03.065. [DOI] [PubMed] [Google Scholar]
- 39.Magnus CR, Arnold CM, Johnston G, Dal-Bello Haas V, Basran J, Krentz JR, Farthing JP. Cross-education for improving strength and mobility after distal radius fractures: a randomized controlled trial. Arch Phys Med Rehabil 94: 1247–1255, 2013. doi: 10.1016/j.apmr.2013.03.005. [DOI] [PubMed] [Google Scholar]
- 40.Magnus CR, Barss TS, Lanovaz JL, Farthing JP. Effects of cross-education on the muscle after a period of unilateral limb immobilization using a shoulder sling and swathe. J Appl Physiol (1985) 109: 1887–1894, 2010. doi: 10.1152/japplphysiol.00597.2010. [DOI] [PubMed] [Google Scholar]
- 41.Maiese K. Driving neural regeneration through the mammalian target of rapamycin. Neural Regen Res 9: 1413–1417, 2014. doi: 10.4103/1673-5374.139453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Marimuthu K, Murton AJ, Greenhaff PL. Mechanisms regulating muscle mass during disuse atrophy and rehabilitation in humans. J Appl Physiol (1985) 110: 555–560, 2011. doi: 10.1152/japplphysiol.00962.2010. [DOI] [PubMed] [Google Scholar]
- 43.Opie GM, Evans A, Ridding MC, Semmler JG. Short-term immobilization influences use-dependent cortical plasticity and fine motor performance. Neuroscience 330: 247–256, 2016. doi: 10.1016/j.neuroscience.2016.06.002. [DOI] [PubMed] [Google Scholar]
- 44.Papandreou M, Billis E, Papathanasiou G, Spyropoulos P, Papaioannou N. Cross-exercise on quadriceps deficit after ACL reconstruction. J Knee Surg 26: 51–58, 2013. [DOI] [PubMed] [Google Scholar]
- 45.Pearce AJ, Hendy A, Bowen WA, Kidgell DJ. Corticospinal adaptations and strength maintenance in the immobilized arm following 3 weeks unilateral strength training. Scand J Med Sci Sports 23: 740–748, 2013. doi: 10.1111/j.1600-0838.2012.01453.x. [DOI] [PubMed] [Google Scholar]
- 46.Phillips SM, McGlory C. CrossTalk proposal: The dominant mechanism causing disuse muscle atrophy is decreased protein synthesis. J Physiol 592: 5341–5343, 2014. doi: 10.1113/jphysiol.2014.273615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Post M, Bakels R, Zijdewind I. Inadvertent contralateral activity during a sustained unilateral contraction reflects the direction of target movement. J Neurosci 29: 6353–6357, 2009. doi: 10.1523/JNEUROSCI.0631-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Rantalainen T, Nikander R, Heinonen A, Daly RM, Sievänen H. An open source approach for regional cortical bone mineral density analysis. J Musculoskelet Neuronal Interact 11: 243–248, 2011. [PubMed] [Google Scholar]
- 49.Ruddy KL, Carson RG. Neural pathways mediating cross education of motor function. Front Hum Neurosci 7: 397, 2013. doi: 10.3389/fnhum.2013.00397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Seger JY, Arvidsson B, Thorstensson A, Seger JY. Specific effects of eccentric and concentric training on muscle strength and morphology in humans. Eur J Appl Physiol Occup Physiol 79: 49–57, 1998. doi: 10.1007/s004210050472. [DOI] [PubMed] [Google Scholar]
- 51.Sherk VD, Bemben MG, Palmer IJ, Bemben DA. Effects of filtering methods on muscle and fat cross-sectional area measurement by pQCT: a technical note. Physiol Meas 32: N65–N72, 2011. doi: 10.1088/0967-3334/32/12/N01. [DOI] [PubMed] [Google Scholar]
- 52.Suetta C, Aagaard P, Rosted A, Jakobsen AK, Duus B, Kjaer M, Magnusson SP. Training-induced changes in muscle CSA, muscle strength, EMG, and rate of force development in elderly subjects after long-term unilateral disuse. J Appl Physiol (1985) 97: 1954–1961, 2004. doi: 10.1152/japplphysiol.01307.2003. [DOI] [PubMed] [Google Scholar]
- 53.Weir JP, Housh TJ, Weir LL, Johnson GO. Effects of unilateral isometric strength training on joint angle specificity and cross-training. Eur J Appl Physiol Occup Physiol 70: 337–343, 1995. doi: 10.1007/BF00865031. [DOI] [PubMed] [Google Scholar]
- 54.Zanette G, Manganotti P, Fiaschi A, Tamburin S. Modulation of motor cortex excitability after upper limb immobilization. Clin Neurophysiol 115: 1264–1275, 2004. doi: 10.1016/j.clinph.2003.12.033. [DOI] [PubMed] [Google Scholar]
- 55.Zehr EP. Considerations for use of the Hoffmann reflex in exercise studies. Eur J Appl Physiol 86: 455–468, 2002. doi: 10.1007/s00421-002-0577-5. [DOI] [PubMed] [Google Scholar]
- 56.Zhou S. Chronic neural adaptations to unilateral exercise: mechanisms of cross education. Exerc Sport Sci Rev 28: 177–184, 2000. [PubMed] [Google Scholar]