

Dynamic management approaches protect endangered bycatch species but with much greater efficiency than existing static closures.
Abstract
Seafood is an essential source of protein for more than 3 billion people worldwide, yet bycatch of threatened species in capture fisheries remains a major impediment to fisheries sustainability. Management measures designed to reduce bycatch often result in significant economic losses and even fisheries closures. Static spatial management approaches can also be rendered ineffective by environmental variability and climate change, as productive habitats shift and introduce new interactions between human activities and protected species. We introduce a new multispecies and dynamic approach that uses daily satellite data to track ocean features and aligns scales of management, species movement, and fisheries. To accomplish this, we create species distribution models for one target species and three bycatch-sensitive species using both satellite telemetry and fisheries observer data. We then integrate species-specific probabilities of occurrence into a single predictive surface, weighing the contribution of each species by management concern. We find that dynamic closures could be 2 to 10 times smaller than existing static closures while still providing adequate protection of endangered nontarget species. Our results highlight the opportunity to implement near real-time management strategies that would both support economically viable fisheries and meet mandated conservation objectives in the face of changing ocean conditions. With recent advances in eco-informatics, dynamic management provides a new climate-ready approach to support sustainable fisheries.
INTRODUCTION
Unsustainable harvest of long-lived ocean predators has resulted in contemporary populations that are a fraction of their baseline biomass (1–4). The loss of marine predators can result in ecosystems crossing tipping points from healthy to degraded states and can lead to a significant loss of ecosystem services (5, 6). Even in fisheries where target fish stocks are currently managed at sustainable levels (2, 6, 7), incidental capture of nontarget species (“bycatch”) remains a significant global problem, threatening many populations of marine mammals, turtles, seabirds, and sharks (8–11). In addition, climate variability and change can create additional risks as productive pelagic habitats shift (12, 13), introducing new ecological interactions (14) and anthropogenic threats (15).
Bycatch mitigation solutions have included changes in fishing gear and methodology, acoustic deterrents, and temporal and spatial management measures such as marine protected areas closed to fishing (8, 16). However, spatial management approaches remain tied largely to static boundaries and coarse temporal scales (17), although the fluidity of interactions among marine predators and their environments has long been recognized (18, 19). Consequently, fixed time-area closures may not always encompass the core habitat of species of concern and may unnecessarily restrict fishing activity when bycatch risk is low. While many of these approaches have been successful for single species, managers are often faced with trading off protection of multiple protected species with sustaining economically viable fisheries (8, 9). Dynamic ocean management is an example of an ecological informatics (“eco-informatics”) approach that uses near real-time data streams to support sustainable use of marine resources (17, 19–22). Dynamic ocean management approaches are robust to climate variability and change, as they account for species movement or distributions at scales matching those of human activities in the oceans (12, 13, 15, 23). Here, we present a multispecies dynamic ocean management approach designed to address the long-standing problem of how to balance sustainable target catch with protected species bycatch.
We focus on the California drift gillnet (DGN) fishery as a case study in a highly dynamic pelagic system. The fishery targets broadbill swordfish (Xiphias gladius) in the California Current, a highly productive upwelling ecosystem and globally significant hotspot of marine biodiversity (24). An underexploited and economically valuable swordfish stock is targeted through overnight deployment of large mesh gillnets of several kilometers in length. This indiscriminant fishing technique can result in bycatch of protected species such as loggerhead (Caretta caretta) and leatherback (Dermochelys coriacea) turtles, small delphinids, beaked whales, and California sea lions (Zalophus californianus; Fig. 1). Because bycatch rates have exceeded management targets, the fishery has been legally mandated to implement mitigation measures (16, 25), including gear modifications and a vast static seasonal area closure [Pacific Leatherback Conservation Area (PLCA); 552,000 km2] implemented in 2001, designed to avoid critically endangered leatherback turtles and beaked whales in California waters (16, 25, 26). Nonetheless, catch rates of some nontarget species have remained high (25). While these management actions have successfully minimized bycatch of leatherback turtles, as intended, they have also resulted in a downsizing of the fishery by 90%, leading to harvest levels well below the sustainable swordfish quota (26), challenging the economic viability of this fishery.
Fig. 1. Map of tracking and fisheries observer data relative to the PLCA.
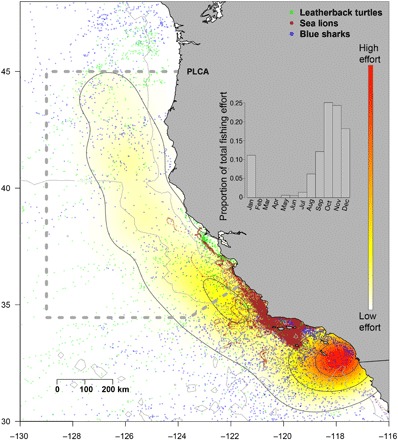
Fisheries observer data are shown as kernel densities, from low (yellow) to high (red) effort. The greatest fishing effort was spatially concentrated in the Southern California Bight. The three tagged species are shown as points, with leatherback turtles migrating from western Pacific breeding grounds to forage in the California Current, blue sharks migrating north-south in the California Current seasonally, and California sea lions remaining within the study area for the duration of our analyses. Inset: The majority of fishing effort (80%) has taken place between August and December throughout the period of the study, with a peak in October.
To address this problem, we take a novel, data-driven, multispecies approach that allows for reduction of bycatch rates while maintaining quota catch. This approach integrates multiple data streams to create an automatable tool that can predict daily relative catch and bycatch probabilities at previously unachievable spatial and temporal scales and provide these predictions to end users in near real time. We also use hindcasts to explore the dynamic nature of habitat intra- and interannually and to assess the efficacy of this tool by comparing its potential for mitigating bycatch relative to the existing static time-area closure (26).
MATERIALS AND METHODS
We used a data-driven, multispecies predictive habitat modeling framework termed EcoCast to create daily predictive surfaces that quantify relative target catch-bycatch probabilities over the domain of the fishery. We first determined habitat preferences of the target species, broadbill swordfish, and three non-target species that interact with the fishery [leatherback turtle, blue shark (Prionace glauca), and California sea lions]. We used two independent species distribution data sets—National Oceanic and Atmospheric Administration (NOAA) fisheries observer data from the DGN fishery (1990–2014) and satellite-linked tracking data collected during the Tagging of Pacific Predators (24) program (2001–2009)—to determine the species’ presence and absence and to sample contemporaneous environmental conditions (Fig. 1 and fig. S1; see Supplemental Methods). The fishery had observers on board for 4 to 23% of the vessels annually from 1990 to 2014, and these data were used to derive swordfish catch (n = 5430 catch events), blue shark bycatch (n = 4591 catch events), and absence sets with no target or bycatch of species (n = 4074 and 4931 sets for swordfish and blue sharks, respectively) for use in species-specific models [see the study of Scales et al. (20)]. Sample sizes of bycatch events for sea lions (n = 157 catch events) and leatherback turtles (n = 25 catch events) were insufficient to build robust models. Argos tracking data were obtained for blue sharks (n = 70 individuals), female California sea lions (n = 75 individuals), and leatherback turtles (n = 20 individuals). Details on tagging methodology are available for sea lions (27, 28), leatherback turtles (26, 29), and blue sharks (24). All tracks were standardized using a state space model, generating a daily regular position including uncertainty estimates to match availability of remotely sensed environmental data (30).
Pseudo-absences (hereafter called absences) for tracking data were generated by creating a set of 20 correlated random walks for each individual, which started at the tagging location and matched the total duration of the tag (fig. S2). Each step was determined by sampling a paired step length and turning angle from the distribution of observed step lengths and turning angles from the corresponding track [see the study of Hazen et al. (31)]. The correlated random walks thus recreate movement characteristics that are similar to the original track but move independently of the underlying environment to sample space available to, but not used by, focal individuals. We collated tracking data and randomly selected absences into a master data set, combining locations received from all individuals sampled from each population. To account for sampling bias and reduce spatial autocorrelation issues, presences and absences from tracking data were randomly and iteratively sub-sampled 1000 times.
Presence and absence data sets were used to sample remotely sensed environmental variables using date, location, and mean position error. All environmental data fields were resampled to a common extent and resolution (0.25° × 0.25° pixel size) that matched the coarsest resolution product. Mean values were sampled at each location over a radius encompassing the mean position error in tracking data (0.5°), and SD was calculated over a 1° × 1° window. Environmental data sets were extracted via Southwest Fisheries Science Center (SWFSC)/Environmental Research Division’s ERDDAP, when available (32), including sea surface temperature (SST) from Pathfinder and GHRSST (mean and SD), zonal wind (y-wind) speed as a proxy for upwelling from Quikscat and Ascat (mean), and chlorophyll a (chl-a) from SeaWiFS and MODIS Aqua (mean). Additional variables were extracted from Aviso+ and CMEMS (Copernicus Marine Environment Monitoring Service) servers, including sea surface height anomaly (SSHa; mean and SD), and eddy kinetic energy (EKE) calculated from zonal and meridional current fields (mean). Sea surface temperature SD, SSHa SD, and EKE provide metrics of mesoscale activity at multiple scales. Lunar illumination, a proxy for the amount of moonlight reaching the sea surface, was calculated using the lunar package v.0.1-14 in R (v. 3.4.0), and bathymetry (mean and SD; that is, bottom depth and roughness) was acquired from ETOPO1. Although our current product may not be considered high resolution for all ocean uses, product resolution can be improved as new satellite products come online or as new ocean-modeled products become operationalized (20, 33).
We built and validated a suite of species distribution models using boosted regression trees with a binomial (presence-absence) response to quantify the probability of the presence of each of our focal species as a function of combined environmental covariates (20, 34). Resultant models then were used to predict relative habitat suitability for each of the focal species at daily time steps (figs. S3 to S5). We examined the spatiotemporal variability in daily predictions at 0.25° resolution over the main period of operation of the fishery (August to December) in two contrasting years—2012, a year of typical oceanographic conditions in the California Current, and 2015, an unusually warm year resulting from a marine heatwave and the strong El Niño event of 2015–2016 (13).
To create an integrated multispecies predictive surface, we weighted each layer by the relative management risk of the focal species before averaging across layers. Species risk weightings were determined on the basis of management concern, discussion with fishers and managers, and fishery bycatch rates such that critically endangered leatherback turtles were given values twice the weighting of blue sharks and over 10 times that of sea lions. Prediction layers for each species were combined into a single surface by multiplying the layer by the species weighting, summing the layers, and then normalizing the range of values in the final predictive surface from −1 (low catch and high bycatch) to 1 (high catch and low bycatch). We assessed a suite of potential species weightings based on management concern (fig. S6) to illustrate how they influence the EcoCast predictions. Given that the weightings are arithmetically determined, increasing bycatch risk for a species by a factor of 2 would also increase risk in the integrated surface proportionally, albeit with different spatial patterns.
Target catch maintenance was equally as important as bycatch reduction in this management scenario. As such, pixels with low integrated risk and low swordfish suitable habitat (<25%) were given a 0 value to ensure that fishing effort was not directed to areas of extremely low bycatch yet also low target catch. The top quartiles of species-specific predictions were plotted in time to illustrate how they changed throughout the season for a normal (2012) and anomalously warm year (2015; fig. S5). In addition, the persistence of bycatch risk for leatherback turtles within the study area was explored over the fishing season (August through November) to examine the efficiency of dynamic closures compared to the 552,000 km2 static Pelagic Leatherback Conservation Area (PLCA). Daily pixels with leatherback turtle habitat suitability greater than 0.25 (top three quartiles) were considered to be a conservative threshold for leatherback bycatch risk. This value was chosen based on previous critical habitat threshold estimates of 0.75 for top predators (12, 35). Suitable-habitat pixels were summed across the 2012 and 2015 fishing seasons independently and divided by the total number of days, resulting in values of 0 to 100% persistence of turtle habitat per pixel. Hypothetical dynamic closures were then compared to the spatial area covered by the Pelagic Leatherback Conservation Area. The calculations for leatherbacks alone included more conservative habitat cutoffs (25%) than that of the integrated multispecies time series (50%). This resulted in smaller closed areas when considering multiple species compared to the conservative approach used when protecting leatherbacks alone.
RESULTS
Species distribution models
We developed habitat suitability models for four focal species (broadbill swordfish, blue shark, California sea lion, and leatherback turtle) over the California Current domain (Fig. 2). Bathymetry was the most informative variable in predicting the presence of all species, followed by sea surface temperature, SSHa, and chl-a concentration (Table 1). When examining the partial effect of a single variable alone (sea surface temperature and bathymetry), all species except sea lions showed overlapping habitat preferences, indicating that a multiple covariate approach is required to tease apart spatial patterns in habitat use (figs. S3 to S5). Our models had AUC (Area Under Receiver Operating Curve) values of 0.72 to 0.93 using a 75% training and 25% test data set but were lower when predicting upon specific years (0.66 to 0.85; table S1). Suitable habitat for the target species, swordfish, was more broadly distributed across the area of operation of the fishery than for nontarget species, highlighting an ability to occupy a broad ecological niche, resulting in multiple fishable regions with low bycatch risk. For example, a suitable habitat for sea lions was limited to coastal areas, while suitable habitat for the leatherback turtle was more patchily distributed offshore. Predictions of habitat suitability for blue sharks generated from models using fisheries-independent (satellite tracking) and fisheries observer data were broadly similar, although the observer-based model predictions showed a more dominant influence of mesoscale variability, possibly as a result of increased fishing effort and catchability at mesoscale features such as fronts and eddies (Fig. 2 and figs. S3 and S4).
Fig. 2. Habitat suitability predictions for individual species for 1 August 2012 that inform the EcoCast tool, from low (white) to high (blue).
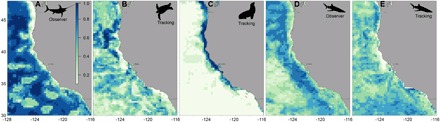
(A) Swordfish (observer) were predicted to occupy the broadest portion of the California Current. (B) Leatherback turtles (tracking) were predicted to occupy waters offshore of the upwelling front, yet avoiding newly upwelled waters. (C) California sea lions (tracking) occupied cool, nearshore waters with proximity to their haul-out sites. (D) Blue sharks (tracking) were also broadly distributed and avoided the coolest upwelled waters nearshore. (E) Blue sharks (observer) were more closely tied to mesoscale features, incorporating both species distribution and fisheries catchability.
Table 1. Variable weightings indicating the importance in final boosted regression tree models for tracking and observer data.
Bathymetry and temperature were reliably the two most important predictors in modeling habitat. Bold numbers highlight the three most important factors for each model. NA, not applicable.
| Observer | Tracking | ||||
| Swordfish | Blue shark | Blue shark | Leatherback | Sea lion | |
| Bottom depth | 32.9 | 47.2 | 15.6 | 14.6 | 49.1 |
| SST mean | 18.0 | 8.0 | 49.3 | 34.7 | 14.3 |
| SSHa | 10.3 | 8.2 | 4.7 | 11.2 | 5.4 |
| Chl-a | 7.9 | 2.7 | 11.0 | 8.9 | 12.2 |
| y-wind | 5.7 | 3.7 | 4.4 | 6.1 | 1.9 |
| Lunar phase | 5.5 | 3.5 | NA | NA | NA |
| Bottom roughness | 5.4 | 5.5 | 4.2 | 11.4 | 3.0 |
| SST SD | 5.3 | 6.7 | 3.2 | 5.1 | 2.8 |
| SSHa SD | 5.1 | 10.2 | 4.3 | NA | 6.0 |
| EKE | 3.9 | 4.4 | 3.4 | 6.1 | 5.3 |
Integrated EcoCast models
Species-specific habitat suitability predictions were integrated to generate a weighted daily predictive surface for all four species using contemporaneous remotely-sensed data fields. In our case study, both bycatch-only and multispecies-integrated predictions showed differing spatial patterns between 2012 and 2015, a normal year and an anomalously warm year, respectively (Fig. 3 and see Supplemental Methods). In 2012, pixels with high bycatch risk were concentrated largely around mesoscale structures outside of the upwelling domain (Fig. 3, A and C, and movie S1). In 2015, pixels with high bycatch risk were close to shore, with less mesoscale influence offshore (Fig. 3, B and D). Both 2012 and 2015 had pixels with persistently high bycatch risk directly south of the PLCA and a narrow band of pixels around 40°N of low risk. Many of the features visible in daily predictions were stable across days if not weeks (movies S1 to S4). Our results suggest that interannual variability can be as important as seasonal variability when designing dynamic management approaches (Fig. 4).
Fig. 3. Predicted risk surfaces for bycatch species and integrated EcoCast product for 1 August 2012 and 2015.
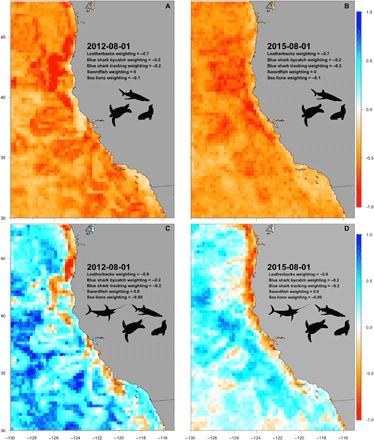
(see movies S1 to S4 for the entire season). Bycatch-only model results for an (A) average year, 2012, in the California Current and (B) an anomalously warm year, 2015, for leatherback turtles, blue sharks, and California sea lions. Integrated EcoCast model predictions for (C) 2012 and (D) 2015 incorporate swordfish in addition to the three aforementioned bycatch species. Values range from −1 (low catch and high bycatch) to 1 (high catch and low bycatch). Risk weightings reflect management concern with leatherbacks and swordfish having the highest, followed by blue sharks, and California sea lions having the lowest (see the Supplementary Materials for details and sensitivity analysis). Risk weightings can be adjusted dependent on management priorities, such as when the fishing season progresses or priorities change.
Fig. 4. Comparison of a dynamic ocean management (DOM) approach relative to the existing seasonal PLCA.
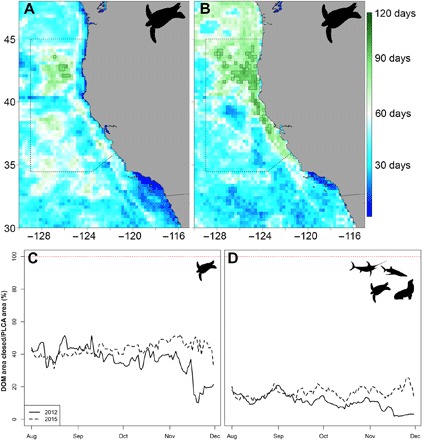
Here, we test two management objectives: a dynamic closure based on a conservative 25% habitat suitability threshold for leatherbacks and a dynamic closure based on the 50% threshold for EcoCast. (A) Nine pixels (dark green, outlined in black) had high turtle bycatch risk for 90 or more days of the 2012 fishing season compared to (B) 79 pixels in the 2015 fishing season. In addition, 86% of the pixels with high turtle bycatch risk for 60 or more days of the season were within the PLCA in 2012 compared to 63% of pixels in 2015. Time series of the ratio of DOM to PLCA area for (C) leatherback protection and (D) integrated EcoCast risk prediction highlight the efficacy of a dynamic approach compared to static protection across contrasting years in the California Current.
EcoCast evaluation
Particular attention was given to one focal species, the leatherback turtle, owing to its protected species status and influence on the design of the existing time-area closure. We quantified persistence of leatherback bycatch risk in each pixel by counting the number of days in which predicted habitat suitability was above a 25% threshold during the August to December fishing season. The temporal patterns in bycatch risk for leatherbacks were explored daily over the fishing season to examine the efficiency of dynamic closures compared to the static PLCA. While the PLCA includes a significant portion of leatherback habitat in the California Current (Fig. 4, A and B), leatherback turtle habitat was more variable in space and time in an average year (2012) compared to a warm year (2015). We found that a dynamic closure would have required about half the area to achieve comparable bycatch risk reduction in August to September of 2012 and would have required only 11% of the PLCA area near the end of November (Fig. 4, C and D). In comparison, suitable habitat for leatherback turtles in 2015 was greater in area and more persistent, with a dynamic closure of 33 to 43% of the PLCA area needed to achieve similar levels of bycatch risk reduction. The same calculation was performed with a 50% threshold for the integrated risk product from August to December, resulting in dynamic closures ranging from 3 to 22% of the PLCA area in 2012 and from 13 to 28% in 2015. Overall, the PLCA is well placed for reducing leatherback turtle bycatch; however, it also greatly reduces fishing opportunity compared to dynamic closures.
DISCUSSION
Bycatch remains one of the most significant global barriers to fisheries sustainability, despite a range of mitigation efforts implemented in individual fisheries (11, 16, 25). Innovation in effective fishery management tools is urgently required to reduce the ecological impacts of non-selective fisheries. Our data-driven, multispecies dynamic ocean management approach provides a novel solution to reducing bycatch while supporting sustained catch rates through coupling high-resolution Earth Observation data with fisheries observer and fisheries-independent data sets. Single-species dynamic ocean management approaches have been applied elsewhere, such as the voluntary framework to reduce loggerhead turtle bycatch in the central North Pacific (36) and the mandatory framework that limits tuna fishing opportunities in the waters of Eastern Australia when bycatch quota is not available (37). However, this is the first example of a multispecies tool that can predict both catch probability and bycatch risk in near real time.
Our analyses suggest that dynamic ocean management can support effective bycatch reduction while maintaining fishing opportunity, thereby allowing fisheries managers to meet dual objectives of ecological and economic sustainability. Our findings suggest that, by tracking daily oceanographic conditions, the California swordfish DGN fishery could access currently closed fishing areas while still protecting leatherback turtles. Previous findings have shown that the timing of the PLCA is well placed to reduce bycatch (26). Our hindcast model predictions show that the current static closure is well placed spatially, although at times overly conservative. When protecting the top 75% of predicted leatherback habitat, dynamic closures were at most half of the size of the current PLCA. When protecting the top 50% of high bycatch and low target catch areas from integrated EcoCast risk products, dynamic closures were a quarter of the size, highlighting the opportunity cost of the current seasonal closure. Variability in closure size was high during the fishing season, as the proportion of predicted suitable habitat for leatherbacks within the existing protected area varied by 16 and 28% within 2012 and 2015, respectively. The interannual variability between these 2 years was up to 41%, suggesting that annual adjustments to closed areas could be an improvement when daily adjustments are not feasible. Intra- and interannual variability in dynamic closure size was muted when considering multiple bycatch species. Holistically, the results provide evidence that an eco-informatics approach that responds to ocean conditions at coarse temporal scales can be more effective than static boundaries in simultaneously supporting fishing opportunities and reducing bycatch.
Moreover, using historical species distribution data to designate static or seasonal closures puts these areas at the risk of losing ecological relevance as species’ distributions shift with a changing climate (38). Anomalous ocean conditions place stress on static management approaches and are likely to become more frequent in the future (39), highlighting the need for climate-ready fisheries management (40). Dynamic ocean management tools can be used to test the efficacy of existing closures under increasing climate variability or long-term change. As the distributions and conservation status of populations of nontarget species change, eco-informatics tools can adapt to varying conservation and exploitation-based economic objectives. The implementation of marine spatial planning is both a sociopolitical and an ecological enterprise, often requiring significant stakeholder engagement when balancing across multiple conflicting needs (41). While static approaches require a new implementation phase to adjust, dynamic ocean management approaches are designed to adjust management boundaries as new information becomes available (42).
With the burgeoning growth of technological products and capacity, there are greater opportunities for eco-informatics approaches to meet challenging, and often conflicting, management mandates in both marine and terrestrial systems (43). Dynamic modeling approaches confer opportunity to proactively address multiple management objectives using hindcasts, real-time data, seasonal forecasts, and climate projections, when available (44). Management approaches that are based on dynamic species-environment relationships offer a potential win-win outcome for protected species and fisheries and provide climate-ready solutions to respond to both ocean variability and directional change. While no model is perfect, the iterative and adaptable design of dynamic ocean management approaches can improve as new data become available and new analytical techniques are developed. The continued development of eco-informatics and dynamic approaches will provide an important pathway to support long-term economic sustainability in concert with conservation of marine biodiversity.
Supplementary Material
Acknowledgments
We thank the scientific teams and all those who supported animal tagging efforts including the Tagging of Pacific Predators program and equally the SWFSC fisheries observer program that collected bycatch data aboard drift gillnet vessels. We are grateful to the numerous captains and crews who provided ship time and logistical support and NOAA regional managers including H. Taylor and T. Fahy who provided feedback and support along the way. We also thank L. Hazen of Stanford’s Center for Ocean Solutions for logistical and meeting support toward achieving the NASA project goals. This project was a brainchild of the late D. Foley whose career was dedicated to incorporating oceanographic data into fisheries management. Funding: This manuscript is the culmination of a NASA Ecoforecasting funded project (NNH12ZDA001N-ECOF) with additional support from California Sea Grant, NOAA’s Bycatch Reduction Engineering Program, NOAA’s West Coast Regional Office, and NOAA’s Integrated Ecosystem Assessment program. Author contributions: This NASA modeling study was initiated by E.L.H., S.J.B., R.L.L., L.B.C., and S.M.M. Tracking data were collected by D.P.C., H.D., S.K., S.R.B., and T.E. Observer and tracking data were compiled by S.M.M., and oceanographic data were compiled by E.L.H., S.J.B., S.M.M., D.K.B., and K.L.S. Analyses were led by E.L.H. and K.L.S. with significant contribution from S.M.M., D.K.B., and H.W. Figures were created by E.L.H. and K.L.S. The manuscript was drafted by E.L.H. and edited by all co-authors. Competing interests: The authors declare that they have no competing interests. Data and materials availability: All data needed to evaluate the conclusions in the paper are present in the paper and/or the Supplementary Materials. Additional data related to this paper may be requested from the authors. Fisheries observer data are available from NOAA’s Observer Program upon request in adherence with U.S. confidentiality laws (www.st.nmfs.noaa.gov/observer-home/). Tagging data are available via IOOS’s Animal Telemetry Network (https://ioos.noaa.gov/project/atn/). Code and updated EcoCast URLs are available via github (https://github.com/elhazen/EcoCast-SciAdv). Correspondence and requests for analyzed data should be addressed to E.L.H. (elliott.hazen@noaa.gov).
SUPPLEMENTARY MATERIALS
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/4/5/eaar3001/DC1
Supplementary Methods
table S1. Species-specific model deviance explained and cross-validation using area under the curve statistics.
fig. S1. Kernel density plot of fisheries effort and tracking data for leatherback turtles, California sea lions, and blue sharks.
fig. S2. Sample track with three randomly selected pseudotracks for all three satellite-tracked species.
fig. S3. Partial response curves from boosted regression trees for sea surface temperature, bathymetry, chl-a, and SSHa across all species models.
fig. S4. Species-specific predictions with error bounds from boosted regression tree model fitting process.
fig. S5. Time series of species habitat in a normal (2012) and anomalously warm (2015) year.
fig. S6. Sensitivity analysis of EcoCast bycatch and integrated risk under varying species weightings to highlight their influence on the final product.
fig. S7. Operational tool for exploring EcoCast weightings available to managers to assess how varying scenarios change the integrated risk surface.
movie S1. Animation of daily bycatch predictions for the August to December 2012 fishing season, with red pixels representing high risk and white representing low risk.
movie S2. Animation of daily bycatch predictions for the August to December 2015 fishing season, with red pixels representing high bycatch risk and white representing low risk.
movie S3. Animation of daily integrated predictions for the August to December 2012 fishing season, with red pixels representing high bycatch risk and low target catch and with blue pixels representing high target catch and low bycatch risk.
movie S4. Animation of daily integrated predictions for the August to December 2015 fishing season, with red pixels representing high bycatch risk and low target catch and with blue pixels representing high target catch and low bycatch risk.
REFERENCES AND NOTES
- 1.Myers R. A., Worm B., Rapid worldwide depletion of predatory fish communities. Nature 423, 280–283 (2003). [DOI] [PubMed] [Google Scholar]
- 2.Worm B., Sandow M., Oschlies A., Lotze H. K., Myers R. A., Global patterns of predator diversity in the open oceans. Science 309, 1365–1369 (2005). [DOI] [PubMed] [Google Scholar]
- 3.Collette B. B., Carpenter K. E., Polidoro B. A., Juan-Jordá M. J., Boustany A., Die D. J., Elfes C., Fox W., Graves J., Harrison L. R., McManus R., Minte-Vera C. V., Nelson R., Restrepo V., Schratwieser J., Sun C.-L., Amorim A., Brick Peres M., Canales C., Cardenas G., Chang S.-K., Chiang W.-C., de Oliveira Leite N. Jr, Harwell H., Lessa R., Fredou F. L., Oxenford H. A., Serra R., Shao K.-T., Sumaila R., Wang S.-P., Watson R., Yáñez E., High value and long life—Double jeopardy for tunas and billfishes. Science 333, 291–292 (2011). [DOI] [PubMed] [Google Scholar]
- 4.Pons M., Branch T. A., Melnychuk M. C., Jensen O. P., Brodziak J., Fromentin J. M., Harley S. J., Haynie A. C., Kell L. T., Maunder M. N., Parma A. M., Restrepo V. R., Sharma R., Ahrens R., Hilborn R., Effects of biological, economic and management factors on tuna and billfish stock status. Fish Fish. 18, 1–21 (2017). [Google Scholar]
- 5.Estes J. A., Terborgh J., Brashares J. S., Power M. E., Berger J., Bond W. J., Carpenter S. R., Essington T. E., Holt R. D., Jackson J. B. C., Marquis R. J., Oksanen L., Oksanen T., Paine R. T., Pikitch E. K., Ripple W. J., Sandin S. A., Scheffer M., Schoener T. W., Shurin J. B., Sinclair A. R. E., Soulé M. E., Virtanen R., Wardle D. A., Trophic downgrading of planet Earth. Science 333, 301–306 (2011). [DOI] [PubMed] [Google Scholar]
- 6.Worm B., Barbier E. B., Beaumont N., Emmett Duffy J., Folke C., Halpern B. S., Jackson J. B. C., Lotze H. K., Micheli F., Palumbi S. R., Sala E., Selkoe K. A., Stachowicz J. J., Watson R., Impacts of biodiversity loss on ocean ecosystem services. Science 314, 787–790 (2006). [DOI] [PubMed] [Google Scholar]
- 7.Worm B., Hilborn R., Baum J. K., Branch T. A., Collie J. S., Costello C., Fogarty M. J., Fulton E. A., Hutchings J. A., Jennings S., Jensen O. P., Lotze H. K., Mace P. M., McClanahan T. R., Minto C., Palumbi S. R., Parma A. M., Ricard D., Rosenberg A. A., Watson R., Zeller D., Rebuilding global fisheries. Science 325, 578–585 (2009). [DOI] [PubMed] [Google Scholar]
- 8.Lewison R. L., Crowder L. B., Read A. J., Freeman S. A., Understanding impacts of fisheries bycatch on marine megafauna. Trends Ecol. Evol. 19, 598–604 (2004). [Google Scholar]
- 9.Lewison R. L., Crowder L. B., Wallace B. P., Moore J. E., Cox T., Zydelis R., McDonald S., DiMatteo A., Dunn D. C., Kot C. Y., Bjorkland R., Kelez S., Soykan C., Stewart K. R., Sims M., Boustany A., Read A. J., Halpin P., Nichols W. J., Safina C., Global patterns of marine mammal, seabird, and sea turtle bycatch reveal taxa-specific and cumulative megafauna hotspots. Proc. Natl. Acad. Sci. U.S.A. 111, 5271–5276 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Sims D. W., Queiroz N., Fish stocks: Unlimited by-catch limits recovery. Nature 531, 448 (2016). [DOI] [PubMed] [Google Scholar]
- 11.Beddington J. R., Agnew D. J., Clark C. W., Current problems in the management of marine fisheries. Science 316, 1713–1716 (2007). [DOI] [PubMed] [Google Scholar]
- 12.Hazen E. L., Jorgensen S., Rykaczewski R. R., Bograd S. J., Foley D. G., Jonsen I. D., Shaffer S. A., Dunne J. P., Costa D. P., Crowder L. B., Block B. A., Predicted habitat shifts of Pacific top predators in a changing climate. Nat. Clim. Change 3, 234–238 (2013). [Google Scholar]
- 13.Di Lorenzo E., Mantua N., Multi-year persistence of the 2014/15 North Pacific marine heatwave. Nat. Clim. Change 6, 1042–1047 (2016). [Google Scholar]
- 14.Poloczanska E. S., Brown C. J., Sydeman W. J., Kiessling W., Schoeman D. S., Moore P. J., Brander K., Bruno J. F., Buckley L. B., Burrows M. T., Duarte C. M., Halpern B. S., Holding J., Kappel C. V., O’Connor M. I., Pandolfi J. M., Parmesan C., Schwing F., Thompson S. A., Richardson A. J., Global imprint of climate change on marine life. Nat. Clim. Change 3, 919–925 (2013). [Google Scholar]
- 15.Maxwell S. M., Hazen E. L., Bograd S. J., Halpern B. S., Breed G. A., Nickel B., Teutschel N. M., Crowder L. B., Benson S., Dutton P. H., Bailey H., Kappes M. A., Kuhn C. E., Weise M. J., Mate B., Shaffer S. A., Hassrick J. L., Henry R. W., Irvine L., McDonald B. I., Robinson P. W., Block B. A., Costa D. P., Cumulative human impacts on marine predators. Nat. Commun. 4, 2688 (2013). [DOI] [PubMed] [Google Scholar]
- 16.Senko J., White E. R., Heppell S. S., Gerber L. R., Comparing bycatch mitigation strategies for vulnerable marine megafauna. Anim. Conserv. 17, 5–18 (2014). [Google Scholar]
- 17.Lewison R., Hobday A. J., Maxwell S., Hazen E., Hartog J. R., Dunn D. C., Briscoe D., Fossette S., O’Keefe C. E., Barnes M., Abecassis M., Bograd S., Bethoney N. D., Bailey H., Wiley D., Andrews S., Hazen L., Crowder L. B., Dynamic ocean management: Identifying the critical ingredients of dynamic approaches to ocean resource management. BioScience 65, 486–498 (2015). [Google Scholar]
- 18.Hyrenbach K. D., Forney K. A., Dayton P. K., Marine protected areas and ocean basin management. Aquat. Conserv. Mar. Freshw. Ecosyst. 10, 437–458 (2000). [Google Scholar]
- 19.Grantham H. S., Game E. T., Lombard A. T., Hobday A. J., Richardson A. J., Beckley L. E., Pressey R. L., Huggett J. A., Coetzee J. C., van der Lingen C. D., Petersen S. L., Merkle D., Possingham H. P., Accommodating dynamic oceanographic processes and pelagic biodiversity in marine conservation planning. PLOS ONE 6, e16552 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Scales K. L., Hazen E. L., Maxwell S. M., Dewar H., Kohin S., Jacox M. G., Edwards C. A., Briscoe D. K., Crowder L. B., Lewison R. L., Bograd S. J., Fit to predict? Eco-informatics for predicting the catchability of a pelagic fish in near real-time. Ecol. Appl. 27, 2313–2329 (2017). [DOI] [PubMed] [Google Scholar]
- 21.Dunn D. C., Maxwell S. M., Boustany A. M., Halpin P. N., Dynamic ocean management increases the efficiency and efficacy of fisheries management. Proc. Natl. Acad. Sci. U.S.A. 113, 668–673 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Maxwell S. M., Hazen E. L., Lewison R. L., Dunn D. C., Bailey H., Bograd S. J., Briscoe D. K., Fossette S., Hobday A. J., Bennett M., Benson S., Caldwell M. R., Costa D. P., Dewar H., Eguchi T., Hazen L., Kohin S., Sippel T., Crowder L. B., Dynamic ocean management: Defining and conceptualizing real-time management of the ocean. Mar. Policy 58, 42–50 (2015). [Google Scholar]
- 23.Hobday A. J., Alexander L. V., Perkins S. E., Smale D. A., Straub S. C., Oliver E. C. J., Benthuysen J. A., Burrows M. T., Donat M. G., Feng M., Holbrook N. J., Moore P. J., Scannell H. A., Gupta A. S., Wernberg T., A hierarchical approach to defining marine heatwaves. Prog. Oceanogr. 141, 227–238 (2016). [Google Scholar]
- 24.Block B. A., Jonsen I. D., Jorgensen S. J., Winship A. J., Shaffer S. A., Bograd S. J., Hazen E. L., Foley D. G., Breed G. A., Harrison A.-L., Ganong J. E., Swithenbank A., Castleton M., Dewar H., Mate B. R., Shillinger G. L., Schaefer K. M., Benson S. R., Weise M. J., Henry R. W., Costa D. P., Tracking apex marine predator movements in a dynamic ocean. Nature 475, 86–90 (2011). [DOI] [PubMed] [Google Scholar]
- 25.Northridge S., Coram A., Kingston A., Crawford R., Disentangling the causes of protected-species bycatch in gillnet fisheries. Conserv. Biol. 31, 686–695 (2016). [DOI] [PubMed] [Google Scholar]
- 26.Eguchi T., Benson S. R., Foley D. G., Forney K. A., Predicting overlap between drift gillnet fishing and leatherback turtle habitat in the California Current Ecosystem. Fish. Oceanogr. 26, 17–33 (2017). [Google Scholar]
- 27.Kuhn C. E., Costa D. P., Interannual variation in the at-sea behavior of California sea lions (Zalophus californianus). Mar. Mamm. Sci. 30, 1297–1319 (2014). [Google Scholar]
- 28.Briscoe D. K., Fossette S., Scales K. L., Hazen E. L., Bograd S. J., Maxwell S. M., McHuron E. A., Robinson P. W., Kuhn C., Costa D. P., Crowder L. B., Lewison R. L., Characterizing habitat suitability for a central-place forager in a dynamic marine environment. Ecol. Evol. 8, 2788–2801 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Benson S. R., Eguchi T., Foley D. G., Forney K. A., Bailey H., Hitipeuw C., Samber B. P., Tapilatu R. F., Rei V., Ramohia P., Pita J., Dutton P. H., Large-scale movements and high-use areas of western Pacific leatherback turtles, Dermochelys coriacea. Ecosphere 2, 1–27 (2011). [Google Scholar]
- 30.Jonsen I. D., Flemming J. M., Myers R. A., Robust state–space modeling of animal movement data. Ecology 86, 2874–2880 (2005). [Google Scholar]
- 31.Hazen E. L., Palacios D. M., Forney K. A., Howell E. A., Becker E., Hoover A. L., Irvine L., DeAngelis M., Bograd S. J., Mate B. R., Bailey H., WhaleWatch: A dynamic management tool for predicting blue whale density in the California Current. J. Appl. Ecol. 54, 1415–1428 (2016). [Google Scholar]
- 32.R. A. Simons, ERDDAP (NOAA/NMFS/SWFSC/ERD, 2017); https://coastwatch.pfeg.noaa.gov/erddap.
- 33.Edwards C. A., Moore A. M., Hoteit I., Cornuelle B. D., Regional ocean data assimilation. Ann. Rev. Mar. Sci. 7, 21–42 (2015). [DOI] [PubMed] [Google Scholar]
- 34.Elith J., Leathwick J. R., Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 40, 677–697 (2009). [Google Scholar]
- 35.Kappes M. A., Shaffer S. A., Tremblay Y., Foley D. G., Palacios D. M., Robinson P. W., Bograd S. J., Costa D. P., Hawaiian albatrosses track interannual variability of marine habitats in the North Pacific. Prog. Oceanogr. 86, 246–260 (2010). [Google Scholar]
- 36.Howell E. A., Kobayashi D. R., Parker D. M., Balazs G. H., Polovina J. J., TurtleWatch: A tool to aid in the bycatch reduction of loggerhead turtles Caretta caretta in the Hawaii-based pelagic longline fishery. Endanger. Species Res. 5, 267–278 (2008). [Google Scholar]
- 37.Hobday A. J., Hartmann K., Near real-time spatial management based on habitat predictions for a longline bycatch species. Fisheries Manag. Ecol. 13, 365–380 (2006). [Google Scholar]
- 38.McLeod E., Salm R., Green A., Almany J., Designing marine protected area networks to address the impacts of climate change. Front. Ecol. Environ. 7, 362–370 (2009). [Google Scholar]
- 39.Boyd P. W., Cornwall C. E., Davison A., Doney S. C., Fourquez M., Hurd C. L., Lima I. D., McMinn A., Biological responses to environmental heterogeneity under future ocean conditions. Glob. Chang. Biol. 22, 2633–2650 (2016). [DOI] [PubMed] [Google Scholar]
- 40.Wilson J. R., Lomonico S., Bradley D., Sievanen L., Dempsey T., Bell M., McAfee S., Costello C., Szuwalski C., McGonigal H., Fitzgerald S., Gleason M., Adaptive comanagement to achieve climate-ready fisheries. Conserv. Lett. 2018, e12452 (2018). [Google Scholar]
- 41.Chuenpagdee R., Pascual-Fernández J. J., Szeliánszky E., Alegret J. L., Fraga J., Jentoft S., Marine protected areas: Re-thinking their inception. Mar. Policy 39, 234–240 (2013). [Google Scholar]
- 42.Song A. M., Scholtens J., Stephen J., Bavinck M., Chuenpagdee R., Transboundary research in fisheries. Mar. Policy 76, 8–18 (2017). [Google Scholar]
- 43.Purves D., Scharlemann J. P. W., Harfoot M., Newbold T., Tittensor D. P., Hutton J., Emmott S., Ecosystems: Time to model all life on Earth. Nature 493, 295–297 (2013). [DOI] [PubMed] [Google Scholar]
- 44.Tommasi D., Stock C. A., Hobday A. J., Methot R., Kaplan I. C., Eveson J. P., Holsman K., Miller T. J., Gaichas S., Gehlen M., Pershing A., Vecchi G. A., Msadek R., Delworth T., Eakin C. M., Haltuch M. A., Séférian R., Spillman C. M., Hartog J. R., Siedlecki S., Samhouri J. F., Muhling B., Asch R. G., Pinsky M. L., Saba V. S., Kapnick S. B., Gaitan C. F., Rykaczewski R. R., Alexander M. A., Xue Y., Pegion K. V., Lynch P., Payne M. R., Kristiansen T., Lehodey P., Werner F. E., Managing living marine resources in a dynamic environment: The role of seasonal to decadal climate forecasts. Prog. Oceanogr. 152, 15–49 (2017). [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/4/5/eaar3001/DC1
Supplementary Methods
table S1. Species-specific model deviance explained and cross-validation using area under the curve statistics.
fig. S1. Kernel density plot of fisheries effort and tracking data for leatherback turtles, California sea lions, and blue sharks.
fig. S2. Sample track with three randomly selected pseudotracks for all three satellite-tracked species.
fig. S3. Partial response curves from boosted regression trees for sea surface temperature, bathymetry, chl-a, and SSHa across all species models.
fig. S4. Species-specific predictions with error bounds from boosted regression tree model fitting process.
fig. S5. Time series of species habitat in a normal (2012) and anomalously warm (2015) year.
fig. S6. Sensitivity analysis of EcoCast bycatch and integrated risk under varying species weightings to highlight their influence on the final product.
fig. S7. Operational tool for exploring EcoCast weightings available to managers to assess how varying scenarios change the integrated risk surface.
movie S1. Animation of daily bycatch predictions for the August to December 2012 fishing season, with red pixels representing high risk and white representing low risk.
movie S2. Animation of daily bycatch predictions for the August to December 2015 fishing season, with red pixels representing high bycatch risk and white representing low risk.
movie S3. Animation of daily integrated predictions for the August to December 2012 fishing season, with red pixels representing high bycatch risk and low target catch and with blue pixels representing high target catch and low bycatch risk.
movie S4. Animation of daily integrated predictions for the August to December 2015 fishing season, with red pixels representing high bycatch risk and low target catch and with blue pixels representing high target catch and low bycatch risk.