

Abstract
The concept of ant slavery rests on the untested assumption that slave-making ants impose fitness costs on colonies of the species they raid. We tested that assumption by comparing the summertime seasonal productivity of Leptothorax spp. colonies in field exclosures without slavemakers, with a colony of the obligatory slave-making ant Protomognathus americanus, or with a colony of the obligatory slavemaker Leptothorax duloticus. Leptothorax longispinosus colonies placed in exclosures with P. americanus colonies did not differ significantly in any demographic attribute from colonies in exclosures without slavemakers. By contrast, Leptothorax curvispinosus colonies exposed to L. duloticus experienced significant reductions in dealate queens, workers, and larvae relative to control colonies exclosed without slavemakers. The pronounced difference in the impact of these slavemakers on their host-species populations correlates with differences in the behavior of the slavemakers observed in the laboratory and likely explains why P. americanus is more abundant than L. duloticus in nature. It seems that more advanced social parasites, like anatomical parasites, evolve to minimize their impact on their hosts, and thus can be regarded as “prudent social parasites.”
Keywords: social parasitism, life-history costs, prudent parasite
Ant slavery is a form of social parasitism in which members of the parasite species exploit the labor of captured host-species workers. Although the behavior of parasites has long been assumed to impose fitness costs on their host-species colonies, those costs have yet to be documented (1).
Slave-making ants raid the nests of other ant species and abscond with the brood. Although some captured brood may be eaten, parasite colonies rear many captured worker pupae; and host workers that eclose from captured broods subsequently augment the slavemaker colony's worker force (2–4). Slave raids disrupt the order of target colonies (5–9), and brood is lost to predation. Furthermore, slave workers not only care for slavemaker and captured host-species broods, they accompany their captors on raids (6, 10). Thus, the labor of slave workers apparently propagates slavemaker genes at the expense of genes shared with relatives.
To explain the apparent paradox of workers laboring on behalf of unrelated gynes, researchers have examined the mechanisms through which slave-species workers form a social attachment to their maternal colony. In virtually all ant species examined, newly eclosed workers are highly accepting of, and acceptable to, any adults to which they are exposed during the first days of their adult lives (11–14). The acceptance of adult nestmates by enslaved workers can thus be regarded as an artifact of a familiarity based mechanism that would normally ensure the acceptance of closely related nestmates and the rejection of non-nestmates. Early learning mechanisms are also thought to account for slave workers' acceptance of slavemaker broods (15–17), although Leptothorax longispinosus workers (commonly enslaved by the slave-making ant Protomognathus americanus) preferentially accept and tend slavemaker broods regardless of their prior experience with ant broods (18).
Such recognition errors are presumably costly, and thus selection should favor recognition mechanisms allowing discrimination against parasites (19). However, this notion rests on the assumption that slavemakers impose a fitness cost on the reproductives in raided host-species colonies and on enslaved workers without providing any compensating benefits. Enslaved workers may benefit, or at least salvage fitness, by producing their own sons, or by selectively rearing reproductives from their own maternal colony. In the laboratory, slave raids result in the decimation of the target colony because queens and workers that have been evicted from their nests have none of the options they might normally enjoy in the field. Under natural conditions, evicted queens and workers may join a surviving nest of their own colony (if the species in question is polydomous), attempt to secure adoption in another colony, or repossess a raided nest after the slavemakers have departed. Being raided does not necessarily represent a “dead end” for a target colony in the field. Further, local parasite-species colonies may benefit host-species colonies by eliminating predators or competitors, if such benefits exceed local parasite pressure. Attempts to measure the costs of slave raids must allow for such contingencies, and therefore must be conducted in a field setting.
To examine the costs that the obligatory slavemakers P. americanus and Leptothorax duloticus impose on colonies of their Leptothorax host species, we contrasted changes in demographics of host-species colonies confined in exclosures with a slavemaker colony to those of host colonies exclosed without a slavemaker colony. P. americanus has been regarded as a highly derived slave-making species (20), given the low level of aggression manifested in its slave raids (6, 9). Unlike L. duloticus (21), P. americanus raiding parties injure few adult members of raided nests (6, 22, 23), and P. americanus colonies rear slave-species queens and males from captured broods (6, 24). Further, unlike L. duloticus, with the exception of raids that occur in late summer, P. americanus raiders seldom expand their colony by moving into raided nests (6, 22, 25, 26). Although P. americanus functions as an obligatory slavemaker, its impact on local host-species populations is predicted to be less than that of L. duloticus given the tendencies of P. americanus to “husband” its hosts.
Materials and Methods
Colonies of the slavemaker P. americanus and its host L. longispinosus were collected in their acorn- or hickory-nut nests from sites in the regional municipalities of Peel and Halton, Ontario, Canada, in late May of 1994 and 1996. Similarly, colonies of the slavemaker L. duloticus and its host Leptothorax curvispinosus were collected from acorns and hickory nuts in the area of Hell, MI, in late May of 1998. In the laboratory, colonies were censused within 14 days of collection for the number of eggs, larvae, prepupae, worker pupae, male pupae, queen pupae, adult workers, adult males, alate queens, and dealate queens in each. After the census, colonies selected for the experiment were induced to move into an acorn that had been kept frozen at −70°C for at least 24 h. Freezing the intact acorn killed any inhabitants and allowed the experimental colonies to establish themselves in a natural nest before the onset of the field experiment.
Pairs of circular exclosures (1.5 m in diameter) were established in mixed deciduous forest by clearing the intended area of cavity-nesting ants. The borders of the exclosures consisted of 30-cm-high garden edging, buried to a depth of 10 cm, and rimmed along the top inner 5 cm with a bead of petroleum jelly. To exclude vertebrate predators, each exclosure was covered with a 1-m-high cone of “chicken wire” supported by wooden doweling. Five host-species nests (L. longispinosus in Ontario or L. curvispinosus in Michigan) were arranged equidistantly around the periphery of the exclosure about 10 cm inside the outer boundary. In addition to these five nests, we placed a slavemaker nest (P. americanus in Ontario or L. duloticus in Michigan) in the center of one exclosure in each pair (slavemaker treatment) and a sixth host-species nest in the center of the other exclosure (host-only treatment). Host-species nests were matched across treatments as well as possible for number of queens, workers, larvae, and pupae. To simulate natural conditions more closely, a vacant acorn (produced by freezing) was placed about 5 cm from each introduced nest. For the trials in Ontario involving P. americanus, 6 exclosure pairs were established by June 15, 1994, and 5 exclosure pairs were established by June 20, 1996. For trials in Michigan involving L. duloticus, seven exclosure pairs were established by May 28, 1998. Exclosures were inspected visually once a week to ensure that the petroleum jelly barrier and the chicken wire covers remained uncompromised. All nests from the exclosures were collected and preserved in 70% ethanol during the first week of September of the same year they were established. We repeated the census procedure on all nests and, after censusing, recorded the exclosure from which the nest had been recovered.
Although the colonies that we introduced were uniquely identifiable by a colony number recorded on a small wooden stake beside each nest, we had no way of knowing whether colonies remained in their original nests. Thus, in our analyses, we considered demography at the level of exclosure populations; that is, as pooled values for the nests in each exclosure. No statistically significant differences were detected in the initial demography of populations established in the two exclosure types in any year the experiment was staged. We used ANOVAs to evaluate the significance of treatment type (slavemaker vs. host-only) for each demographic cohort for both the P. americanus and L. duloticus data, adjusting the resultant probability values from those tests by using the sequential Bonferroni technique (27) to preserve an experimentwise type I error of 0.05. Further, we contrasted the frequency of occupied and unoccupied acorn nests in slavemaker vs. host-only exclosures by using G tests on 2 × 2 contingency tables. Finally, we evaluated whether the impact on host-species colonies was proportional to the size of slavemaker colonies by regressing host-colony demographics (the number of dealate queens, workers, worker pupae, larvae, and eggs) on the total number of slavemaker workers and the number of enslaved Leptothorax workers within slavemaker exclosures. Significance values for those regression analyses were adjusted by using the sequential Bonferroni technique to maintain an experimentwise error rate of 0.05.
Results
Fewer individuals of each caste/age cohort were recovered from L. longispinosus colonies in slavemaker exclosures than from exclosures that did not contain a P. americanus colony, although none of those differences were statistically significant (Table 1). Further, the ratio of occupied to unoccupied nests did not differ between exclosure types (Table 1).
Table 1.
L. longispinosus colony demographics in population exclosures containing a P. americanus colony (Experimental; n = 11) vs. exclosures without a slavemaker colony (Control; n = 11)
| Cohort | Control | Experimental | F1,20 | P | Padj |
|---|---|---|---|---|---|
| Dealate queens | 8.55 ± 1.50 | 5.91 ± 1.41 | 1.64 | 0.21 | 0.82 |
| Alate queens | 2.18 ± 0.91 | 0.36 ± 0.20 | 3.78 | 0.07 | 0.46 |
| Males | 1.73 ± 0.59 | 0.36 ± 0.24 | 4.57 | 0.05 | 0.41 |
| Workers | 290.09 ± 31.44 | 210.73 ± 45.10 | 2.08 | 0.16 | 0.82 |
| Queen pupae | 0.36 ± 0.20 | 0.00 ± 0.00 | 3.20 | 0.09 | 0.53 |
| Male pupae | 0.09 ± 0.09 | 0.00 ± 0.00 | 1.00 | 0.33 | 0.82 |
| Worker pupae | 6.18 ± 1.96 | 3.36 ± 1.56 | 1.27 | 0.27 | 0.82 |
| Larvae | 380.45 ± 39.76 | 282.45 ± 57.78 | 1.95 | 0.18 | 0.82 |
| Eggs | 5.27 ± 1.43 | 2.00 ± 0.59 | 4.49 | 0.05 | 0.41 |
| Nests occupied | 69 | 70 | |||
| Nests unoccupied | 46 | 51 | Gadj = 0.04, P = 0.84 | ||
Values shown are mean ± SE except for nest occupancy where numerical frequencies are shown. Probability values from ANOVA are adjusted (Padj) by using the sequential Bonferroni technique.
By contrast, L. curvispinosus nests in exclosures with the slavemaker L. duloticus had significantly fewer dealate queens, workers, and larvae after recovery than conspecific nests exclosed without a slavemaker colony (Table 2). The ratio of occupied to unoccupied nests also differed between exclosure types: a preponderance of unoccupied nests was evident in exclosures with L. duloticus, whereas the reverse was true in exclosures containing only L. curvispinosus (Table 2).
Table 2.
L. curvispinosus colony demographics in population exclosures containing an L. duloticus colony (Experimental; n = 7) vs. exclosures without a slavemaker colony (Control; n = 7)
| Cohort | Control | Experimental | F1,12 | P | Padj |
|---|---|---|---|---|---|
| Dealate queens | 8.29 ± 0.71 | 4.00 ± 0.69 | 18.62 | 0.001 | 0.005 |
| Alate queens | 0.14 ± 0.14 | 0.00 ± 0.00 | 1.00 | 0.34 | 1.0 |
| Males | 0.00 ± 0.00 | 0.00 ± 0.00 | — | — | — |
| Workers | 302.00 ± 29.07 | 113.00 ± 30.08 | 20.42 | 0.0007 | 0.004 |
| Queen pupae | 0.00 ± 0.00 | 0.00 ± 0.00 | — | — | — |
| Male pupae | 0.14 ± 0.14 | 0.00 ± 0.00 | 1.00 | 0.34 | 1.0 |
| Worker pupae | 11.14 ± 5.42 | 2.29 ± 0.84 | 2.61 | 0.13 | 0.52 |
| Larvae | 305.57 ± 24.93 | 95.43 ± 24.61 | 36.00 | 0.0001 | 0.0007 |
| Eggs | 8.57 ± 2.46 | 5.86 ± 4.18 | 0.31 | 0.59 | 1.0 |
| Nests occupied | 48 | 31 | |||
| Nests unoccupied | 34 | 48 | Gadj = 5.25, P = 0.02 | ||
Values shown are mean ± SE except for nest occupancy where numerical frequencies are shown. Probability values from ANOVA are adjusted (Padj) by using the sequential Bonferroni technique.
Neither the number of P. americanus workers (Table 3) nor the number of L. longispinosus workers enslaved in their colonies (Table 4) had any significant relationship to abundance within any demographic cohort of unenslaved nests within experimental exclosures. However, the number of L. duloticus workers did have a significant inverse relationship with the number of unenslaved L. curvispinosus workers in experimental exclosures, explaining 70% of the variation in unenslaved worker abundance (Table 5). Further, the number of enslaved L. curvispinosus workers in those exclosures had significant inverse relationships with both unenslaved dealate queens and workers, and tended to affect both the number of larvae and eggs recovered from unenslaved nests (Table 6).
Table 3.
L. longispinosus colony demographics relative to the number of P. americanus workers in experimental populations (n = 11) as evaluated by simple linear regression
| Cohort | Least squares equation | F1,9 | P | Padj |
r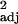
|
|---|---|---|---|---|---|
| Dealate queens | y = −0.32x + 7.69 | 0.55 | 0.48 | 0.64 | 0.00 |
| Workers | y = −19.31x + 317.79 | 2.31 | 0.16 | 0.49 | 0.12 |
| Worker pupae | y = 0.49x + 0.66 | 1.11 | 0.33 | 0.64 | 0.01 |
| Larvae | y = −27.69x + 436.03 | 3.10 | 0.11 | 0.45 | 0.17 |
| Eggs | y = −0.35x + 3.94 | 5.91 | 0.04 | 0.19 | 0.33 |
Table 4.
L. longispinosus colony demographics relative to the number of enslaved L. longispinosus workers in experimental populations (n = 11) as evaluated by simple linear regression
| Cohort | Least squares equation | F1,9 | P | Padj |
r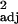
|
|---|---|---|---|---|---|
| Dealate queens | y = −0.01x + 6.61 | 0.11 | 0.75 | 0.75 | 0.00 |
| Workers | y = −0.96x + 298.36 | 2.02 | 0.19 | 0.75 | 0.09 |
| Worker pupae | y = 0.02x + 1.09 | 1.04 | 0.33 | 0.75 | 0.00 |
| Larvae | y = −1.21x + 392.62 | 1.93 | 0.20 | 0.75 | 0.09 |
| Eggs | y = −0.02x + 3.51 | 4.24 | 0.07 | 0.35 | 0.24 |
Table 5.
L. curvispinosus colony demographics relative to the number of L. duloticus workers in experimental populations (n = 7) as evaluated by simple linear regression
| Cohort | Least squares equation | F1,5 | P | Padj |
r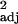
|
|---|---|---|---|---|---|
| Dealate queens | y = −0.17x + 5.91 | 7.09 | 0.04 | 0.16 | 0.50 |
| Workers | y = −8.16x + 207.38 | 15.19 | 0.01 | 0.05 | 0.70 |
| Worker pupae | y = −0.04x + 2.69 | 0.09 | 0.77 | 0.86 | 0.00 |
| Larvae | y = −3.97x + 141.41 | 1.82 | 0.24 | 0.72 | 0.12 |
| Eggs | y = 0.47x + 0.46 | 0.73 | 0.43 | 0.86 | 0.00 |
Table 6.
L. curvispinosus colony demographics relative to the number of enslaved L. curvispinosus workers in experimental populations (n = 7) as evaluated by simple linear regression
| Cohort | Least squares equation | F1,5 | P | Padj |
r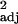
|
|---|---|---|---|---|---|
| Dealate queens | y = −0.02x + 5.79 | 24.61 | 0.004 | 0.02 | 0.80 |
| Workers | y = −0.91x + 187.70 | 16.03 | 0.01 | 0.04 | 0.72 |
| Worker pupae | y = −0.01x + 2.98 | 0.46 | 0.53 | 0.53 | 0.00 |
| Larvae | y = −0.64x + 147.93 | 6.43 | 0.05 | 0.15 | 0.48 |
| Eggs | y = 0.11x − 3.18 | 6.86 | 0.05 | 0.15 | 0.49 |
Discussion
Our results reveal that slavemakers impose costs on their hosts, but that short-term costs differ substantially between the two slave-making species examined. None of our tests revealed any statistically significant influence of P. americanus on L. longispinosus colonies. However, the general trend toward reductions in each demographic cohort among colonies exclosed with P. americanus relative to controls suggests that fitness costs would be evident when amortized over the life of a L. longispinosus queen. Studies of longer duration are required to document such costs.
In stark contrast, L. duloticus colonies significantly reduced the number of free-living L. curvispinosus queens, workers, and larvae in their exclosures. Further, the magnitude of those losses increased with increases in the size of the slavemaker colony. The increase in the proportion of vacant nests in just over 3 months where L. duloticus was present suggests that L. duloticus may routinely decimate local Leptothorax populations. Robust Leptothorax colonies expand by occupying multiple nests (28–30). Thus, Leptothorax colonies confronted with local L. duloticus seem to be at a distinct disadvantage, whereas free-living colonies persist when faced with local P. americanus.
We do not know whether invertebrate predators contributed to losses of ants in our exclosures. Competition for nest sites, which can be significant in limiting Leptothorax populations (30), can be assayed by considering the number of colonies of potential competitors that became established in the exclosures. Colonies of potential Leptothorax competitors (Tapinoma and Lasius sp.) became established twice in host-only exclosures, but only once in a slavemaker exclosure for trials involving P. americanus, and twice in both the host-only and L. duloticus exclosures in Michigan. Thus, there is no compelling evidence that local slavemakers reduce interspecific competition on their Leptothorax hosts.
The differential impact of the two slave-making species is not a product of variation in the power of our statistical tests. Eleven pairs of exclosures were ultimately established for tests involving P. americanus, and only seven pairs were used in assaying the impact of L. duloticus. Thus, if anything, our tests would be biased toward detecting costs imposed by P. americanus and not L. duloticus. Further, the parasite pressure experienced by Leptothorax nests in our study far exceeds that typical in nature. In our experimental exclosures, the ratio of slavemaker to host-species nests was 1:5, which greatly exceeds the relative abundance of parasite nests in the field (T.M.A., personal observation). Thus, our tests are not overly conservative. Further, although susceptibility to parasites may differ between L. longispinosus and L. curvispinosus, we chose to test each slavemaker with its most common host, thereby obtaining data representative of natural interactions. Finally, although assaying impacts in different years and in different geographical locations is not ideal, the design itself is a product of our findings and ethical constraints. We performed tests on P. americanus between 1994 and 1996, and after attempting to publish those data, we found reviewers attributed the failure to detect costs to aspects of our experimental technique instead of the low impact of P. americanus on host colonies. Thus, to validate and extend our findings, we chose to replicate the experiment with L. duloticus in Michigan. Because L. duloticus and its host L. curvispinosus do not occur in the region where our P. americanus work was conducted, and because establishing exclosures containing L. duloticus and L. curvispinosus would have imposed the unethical risk of introducing these species into an area where they do not naturally occur, we were forced to conduct the experiment with L. duloticus in a locality to which it is native. Despite the resultant methodological untidiness, our findings at the very least indicate that the impact of slave-making parasites on host populations varies. Indeed, Foitzik et al. (31) have demonstrated geographical variation in the impact of P. americanus on its hosts and concomitant variation in the defensive behavior of hosts that they interpret as a product of the coevolutionary relationship between that social parasite and its host species.
Nevertheless, there are independent reasons to believe that P. americanus and L. duloticus impact host populations differentially. Alloway (6) observed in the laboratory that P. americanus raiding parties seldom killed target-colony adults and never occupied raided nests. In contrast, L. duloticus raiding parties killed most adults in raided nests and, in 2 of 10 raids, emigrated into the raided nest (6). In addition, field and laboratory studies indicate that P. americanus colonies regularly rear captured host-species queen and male pupae to adulthood, whereas L. duloticus colonies rarely if ever do so (T.M.A., unpublished results). The differential impact of the two slavemaker species documented in the present study is consistent with those differences in raiding and brood-rearing behaviors.
Under some circumstances, selection might favor social parasites that limit the damage imposed on their hosts, providing a social-parasite analogy to what Holmes (32) has termed “prudent parasitism.” This “prudence” or stewardship of the host resource should allow its practitioners to achieve higher population densities than those that wantonly exploit their hosts. It is thus no coincidence that P. americanus has been reported to achieve local population densities two to three times higher than those found for L. duloticus (6). Indeed, whereas the tactics of P. americanus would allow prolonged residence in the same nest, it is not surprising that L. duloticus would move to occupy raided nests; only in this manner would L. duloticus enjoy access to additional Leptothorax slaves. Studies examining costs imposed by other slave-making ants in relation to the purported length of their evolutionary association with their hosts and the relationship of costs to the abundance of those slavemakers should prove particularly enlightening in evaluating the “prudent social parasite” hypothesis.
It has been suggested for hosts of avian brood parasites that the prolonged evolutionary association between a social parasite and its host may result in adjustments of the host's life history such that costs are minimized (33). Although such modifications cannot be addressed by the present data, evolutionary responses to selection pressures imposed by slave-making ants may be constrained in Leptothorax by benefits associated with polydomy and polygyny. The occupation of multiple nests (polydomy) and incorporation of multiple egg-laying queens in a single colony (polygyny) facilitate colony growth and provide insurance against accident (28). These processes, however, also enforce relaxed discrimination through enhanced variation in recognition signatures. Although the minimal loss of brood to P. americanus may be compensated for by benefits of polydomy and polygyny, that same flexibility in social discrimination may render Leptothorax colonies vulnerable to less prudent parasites like L. duloticus. Tradeoffs of this nature will become evident only through longer-term studies (over the life of Leptothorax queens) that simultaneously consider costs and benefits across multiple levels of the community.
Acknowledgments
We thank R. L. Baker for discussions provoking our interest in documenting costs and F. J. Aznar, A. O. Bush, K. S. Ellison, W. J. Gallaway, J. O. Murie, S. C. Smith, T. J. Underwood, B. Wallace, and several anonymous reviewers for comments on previous versions of the manuscript. We also thank L. Harewood, T. Howard, M. Lobosalmo, J. Melbye, and F. Trampus for assistance in executing the study. A. Fleming and R. Nussbaum are thanked for allowing access to freezers used in producing vacant nests, and both R. Nussbaum and J. Bolgos are thanked for facilitating access to the Edwin S. George Reserve in Michigan. Funding for this research was provided by the Natural Sciences and Engineering Research Council of Canada and the Brandon University Research Committee.
References
- 1.Davies N B, Bourke A F G, Brooke M d L. Trends Ecol Evol. 1989;4:274–278. doi: 10.1016/0169-5347(89)90202-4. [DOI] [PubMed] [Google Scholar]
- 2.Wilson E O. Evolution (Lawrence, Kans) 1975;29:108–119. doi: 10.1111/j.1558-5646.1975.tb00819.x. [DOI] [PubMed] [Google Scholar]
- 3.Alloway T M. Am Nat. 1980;115:247–261. [Google Scholar]
- 4.Buschinger A, Ehrhardt W, Winter U. Z Tierpsychol. 1980;53:245–264. [Google Scholar]
- 5.Creighton W S. Psyche. 1927;34:11–29. [Google Scholar]
- 6.Alloway T M. Anim Behav. 1979;27:202–210. [Google Scholar]
- 7.Buschinger A. Insectes Sociaux. 1983;30:235–240. [Google Scholar]
- 8.Stuart R J, Alloway T M. Behavior. 1983;85:58–90. [Google Scholar]
- 9.Topoff H. Am Sci. 1990;78:520–528. [Google Scholar]
- 10.Alloway T M. In: The Biology of Social Insects. Breed M D, Michener C D, Evans H E, editors. Boulder, CO: Westview; 1982. pp. 261–265. [Google Scholar]
- 11.Carlin N F, Hölldobler B. Science. 1983;222:1027–1029. doi: 10.1126/science.222.4627.1027. [DOI] [PubMed] [Google Scholar]
- 12.Morel L, Vander Meer R K. In: Chemistry and Biology of Social Insects. Eder J, Rembold H, editors. Munich: Verlag J. Peperny; 1987. pp. 471–472. [Google Scholar]
- 13.Alloway T M, Ryckman D. Behavior. 1991;118:235–243. [Google Scholar]
- 14.Stuart R J. Behav Ecol Sociobiol. 1992;30:403–408. [Google Scholar]
- 15.Jaisson P. Behavior. 1975;52:1–37. [Google Scholar]
- 16.Le Moli F, Passetti M. Atti Soc Ital Sci Nat Mus Civ Stor Nat Milano. 1977;118:49–64. [Google Scholar]
- 17.Le Moli F. Boll Zool. 1980;47:207–212. [Google Scholar]
- 18.Alloway T M, Hare J F. Behavior. 1989;110:93–105. [Google Scholar]
- 19.Lorenzana J C, Sealy S G. Stud Av Biol. 1999;18:241–253. [Google Scholar]
- 20.Stuart R J, Alloway T M. Anim Behav. 1985;33:1080–1088. [Google Scholar]
- 21.Buschinger A. In: Biosystematics of Social Insects. Howse P E, Clément J-L, editors. New York: Academic; 1981. pp. 211–222. [Google Scholar]
- 22.Wesson L G., Jr Trans Am Ent Soc. 1939;65:97–122. [Google Scholar]
- 23.Wesson L G., Jr Bull Brooklyn Ent Soc. 1940;35:73–83. [Google Scholar]
- 24.Sturtevant A H. Psyche. 1927;34:1–9. [Google Scholar]
- 25.Buschinger A, Alloway T M. Psyche. 1977;83:233–242. [Google Scholar]
- 26.Del Rio Pesado M G, Alloway T M. Psyche. 1983;90:151–162. [Google Scholar]
- 27.Rice W R. Evolution (Lawrence, Kans) 1989;43:223–225. doi: 10.1111/j.1558-5646.1989.tb04220.x. [DOI] [PubMed] [Google Scholar]
- 28.Alloway T M, Buschinger A, Talbot M, Stuart R, Thomas C. Psyche. 1982;89:249–274. [Google Scholar]
- 29.Stuart R J. Psyche. 1985;92:71–81. [Google Scholar]
- 30.Herbers J M. Behav Ecol Sociobiol. 1986;19:115–122. [Google Scholar]
- 31.Foitzik S, DeHeer C J, Hunjan D N, Herbers J M. Proc R Soc London Ser B. 2001;268:1139–1146. doi: 10.1098/rspb.2001.1627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Holmes J C. In: Coevolution. Futuyma D J, Slatkin M, editors. Sunderland, MA: Sinauer; 1988. pp. 161–185. [Google Scholar]
- 33.Brooker M, Brooker L. Behav Ecol. 1996;7:395–407. [Google Scholar]