

Abstract
Burkholderia pseudomallei is a Gram-negative bacterium that causes melioidosis, which can be fatal in humans. Melioidosis is prevalent in the tropical regions of Southeast Asia and Northern Australia. Ecological data have shown that this bacterium can survive as a free-living organism in environmental niches, such as soil and water, as well as a parasite living in host organisms, such as ameba, plants, fungi, and animals. This review provides an overview of the survival and adaptation of B. pseudomallei to stressful conditions induced by hostile environmental factors, such as salinity, oxidation, and iron levels. The adaptation of B. pseudomallei in host cells is also reviewed. The adaptive survival mechanisms of this pathogen mainly involve modulation of gene and protein expression, which could cause alterations in the bacteria's cell membrane, metabolism, and virulence. Understanding the adaptations of this organism to environmental factors provides important insights into the survival and pathogenesis of B. pseudomallei, which may lead to the development of novel strategies for the control, prevention, and treatment of melioidosis in the future.
1. Introduction
Burkholderia pseudomallei is a Gram-negative bacterium that is the causative agent of melioidosis, an infectious disease of public-health significance in Southeast Asia and Northern Australia [1, 2]. In an endemic area, a mortality rate of 40% among septicemic melioidosis patients has been reported in Thailand [3]. In Australia, the mortality rate was about 21% [3]. In nature, B. pseudomallei can survive in diverse environmental niches, indicating an ability to sense and respond to changes in the environment via specific survival mechanisms. This review provides information on the adaptations of B. pseudomallei in response to various hostile environmental stress factors, such as salinity, oxidation, and iron content, as well as its adaptation in target cells. Essential changes in gene and protein expression to enable B. pseudomallei to adapt are explored. Future directions for B. pseudomallei research are also discussed.
2. Background Information for B. pseudomallei
B. pseudomallei was discovered in 1911 by Whitmore and his team as a bacterial agent associated with “glanders-like” disease [2]. Many years later, this bacterium was proven to cause melioidosis [4]. This life-threatening disease presents with a wide range of nonspecific signs and symptoms, including fever, pneumonia, acute septicemia, and chronic localized infection [4, 5]. Chronic infection can cause abscesses in various internal organs, such as the lungs, liver, spleen, kidneys, prostate gland, and skeletal muscles [5]. The routes of transmission of B. pseudomallei include inoculation via skin abrasion, inhalation, and ingestion [6]. Patients with diabetes, thalassemia, or renal disease or people who work in paddy fields have been reported to have a higher risk of melioidosis [7]. However, healthy individuals with no obvious risk factors can also become infected, albeit with significantly lower risk. Without appropriate treatment, the septicemic form of melioidosis can develop and is associated with >90% mortality rate [2]. Currently, no effective vaccine exists to prevent melioidosis.
B. pseudomallei is found in a wide range of ecological niches, including soil and surface water, and has also been found to adhere to the roots of legumes [8]. The wide variety of B. pseudomallei habitats may help explain the persistence of this bacterium in endemic areas. Several studies have indicated that climatic, physical, chemical, and biological factors could control the proliferation and survival of B. pseudomallei in the environment. For example, many studies have demonstrated an association between the frequency of recorded melioidosis cases and rainfall-related events [9–11]. Merritt and Inglis suggested that the levels of cloud cover during rainfall correspond to the levels of soil moisture and might provide appropriate conditions for the survival of B. pseudomallei [12]. Dense cloud cover may provide B. pseudomallei with protection from bactericidal UV wavelengths in sunlight [12]. Soil is considered a major reservoir of B. pseudomallei, a saprophytic organism. Thus, the physiochemistry of the soil is likely a key factor supporting the survival of B. pseudomallei. Manivanh et al. [13] found the prevalence of B. pseudomallei to be high at soil depths > 30 cm with high water content and low total nitrogen, carbon, and organic matter. Tong et al. [14] showed that B. pseudomallei can survive in soils with 20% moisture for 439 days, which is longer than without water, where bacterial survival was only 30 days. This may be because soil moisture supports the availability of bacterial nutrients and membrane integrity [8]. It has been demonstrated that B. pseudomallei can persist in harsh-environment, nutrient-limited settings, such as low-iron environments [15]. Conversely, Musa et al. [16] found that soil containing high levels of iron was strongly associated with B. pseudomallei persistence. It is possible that iron can regulate the expression of respiratory enzymes in the biological processes involved in B. pseudomallei survival [17]. Biological factors are important for B. pseudomallei to persist in the environment. It is evident that free-living amebae are an environmental reservoir for B. pseudomallei. The important amebae for B. pseudomallei are Acanthamoeba spp., Hartmannella spp., and Naegleria spp. [18]. This seems to be advantageous for B. pseudomallei survival in harmful conditions. B. pseudomallei has reportedly survived inside amebae in an environment contaminated with disinfectants and antibiotics [19]. However, not all ameba species can facilitate the persistence and dispersal of a particular bacterial pathogen in the environment, since some amebae isolated from endemic areas can antagonize B. pseudomallei [20], including Paravahlkampfia ustiana, Acanthamoeba spp., and isolate A-ST39-E1.
B. pseudomallei is likely to be constantly exposed to a variety of stressful conditions, forcing it to adapt and to survive in environmental niches. B. pseudomallei contains a number of genes that are important for survival and adaptation. The genome sequence of B. pseudomallei has revealed that the bacterium has two large chromosomes (4.07 and 3.17 Mb), containing at least 16 genomic islands [21]. The major chromosome carries many genes associated with core functions, such as cell growth and metabolism, whereas the smaller chromosome carries genes encoding accessory functions, such as those required for bacterial adaptation and virulence. In addition to survival in the environment, upon infection, B. pseudomallei has the ability to survive inside a variety of host cells, and this is mediated by several virulence factors, such as type 3 secretion system (T3SS), type 5 secretion system (T5SS), type 6 secretion system (T6SS), lipopolysaccharide (LPS), and flagella, as well as numerous bacterial products and enzymes [22, 23]. These factors contribute to B. pseudomallei pathogenesis [22] and also occasionally facilitate its adaptation under hostile environmental conditions. This issue will be discussed further below.
3. Molecular Mechanisms for B. pseudomallei Adaptation in the Presence of Hostile and Environmental Factors
The unusual ability of B. pseudomallei to survive for months or years in the environment is evident by the persistence of this bacterium in melioidosis-endemic areas [8]. Indeed, B. pseudomallei showed the ability to survive in adverse environments, including lack of nutrients [24], limited oxygen [25, 26], and exposure to high salt concentrations [27, 28] and oxidative agents [29, 30]. Importantly, B. pseudomallei is able to sequester within human macrophages and lymphoreticular organs in a dormant or quiescent state for many years [8]. Various environmental and hostile stresses are critical factors contributing to the adaptive survival mechanisms of B. pseudomallei. Like other bacteria, B. pseudomallei possesses various mechanisms to modulate its gene expression for survival under stress (Table 1). The adaptation of B. pseudomallei to stress includes modulation of the expression of genes encoding important proteins, such as short-chain dehydrogenase/oxidoreductase (SDO) [31], acyl-CoA dehydrogenase [27], Burkholderia secretion apparatus (Bsa) T3SS [27], beta-lactamase-like protein [28], sigma factor E (RpoE) [27, 32], and heat-shock proteins [27, 32] for salt stress; KatG and KatE catalase enzymes [33], sigma factor S (RpoS) [34], succinyl-CoA: 3-ketoacid CoA transferase (SCOT) [34], and DpsA [35] for oxidative stress; Fur [36], pyochelin [37], pyoverdine [37], ornibactin [37], cepabactin [37], and biofilm formation-associated regulator [38] for iron stress; and ATP synthases [25], polyhydroxybutyrate synthase [25], pyruvate dehydrogenase [25], acetate kinase [25], alcohol dehydrogenase [25], motility-mediated proteins [25], stress-related proteins [25], and virulence factors [25] for oxygen stress. Most of these stress-response proteins have been observed to react with sera from melioidosis patients [39], potentially indicating their important roles in the adaptation of bacteria to survive under ecologically stressful conditions.
Table 1.
Stress-regulated genes for Burkholderia pseudomallei adaptation.
| Type of stress | Stress regulated genes | ||
|---|---|---|---|
| Membrane structure | Metabolism | Virulence | |
| Salinity | |||
| High level | gene encoding Acyl-CoA dehydrogenase [27] and cyaB [27] |
gene encoding SDO [31] | bsa-encoded genes (bipD and bopE) [27], rpoE [27], mucB [27], groEL [32], and htpG [32] |
| Low level | - | - | - |
| Oxidative condition | |||
| High level | rpoE [29] and speG [29] | rpoS [34], scoA [34], cysM [34], panB [34], pdxJ [34], gene encoding SCOT [34], and genes encoding KatG and KatE [33] | dpsA [35] |
| Low level | - | - | - |
| Iron | |||
| High level | - | fur [36] | mba [37], pch [37], and bhu/hmu [17] |
| Low level | - | BPSS0495 [17] | fur [36], bfmR [38], and genes encoding T6SS [76] |
| Oxygen | |||
| High level | - | - | - |
| Low level | - | Genes encoding ATP synthases (atpA and atpD), arginine and pyruvate fermentation (aceE, arcD and tatA), electron transport proteins (aarC,cydA, cydB, mocA, and BPSL1260), and polyhydroxybutyrate synthase (bdhA-2) [25] | Genes encoding flagella-mediated motility (flgA, flgC, flgK,flgM, fliF, fliJ, fliK, and pilT), stress-related proteins (clpB, rpoH, and rpoS), and virulence factors (bopE, bipC, bipD, orgA,and pilA) [25] |
3.1. Adaptation to Salt Stress
In Thailand, the highest incidence of melioidosis and the highest prevalence of B. pseudomallei are found in the northeast, where saline soil and water are abundant [15]. This raises the possibility that B. pseudomallei can adapt to saline conditions and gain a selective ecological advantage over other soil microorganisms. Consistent with this, B. pseudomallei infection has been reported in cystic fibrosis patients, who have higher salt concentrations in their lungs than healthy individuals [27]. Several studies have revealed that exposure to high salinity influences B. pseudomallei survival and virulence, by adjusting the expression of genes and proteins involved in bacterial physiology, virulence, and metabolism [27, 31, 40]. A possible mechanism by which B. pseudomallei adapts to counter salinity stress is shown in Figure 1.
Figure 1.

Mechanism of Burkholderia pseudomallei adaptation in response to high salt stress.
In general, when bacteria encounter salt stress, they recognize environmental stress with an osmosensor [41]. It has been reported that an adenylate cyclase (CyaB) acts as an osmosensor in the Gram-negative saprophytic bacterium Myxococcus xanthus [42]. Under salt stress, the expression of B. pseudomallei adenylate cyclase is increased [27]. Adenylate cyclase might function as an osmosensor in B. pseudomallei or might be involved in the transmission of the signal. However, the exact role of adenylate cyclase in adaptation to salt stress is still unknown.
Under salt-stress conditions, there is evidence of severely impaired growth and morphology in B. pseudomallei [27, 43]. In our previous study, B. pseudomallei K96243 demonstrated growth impairment during culturing in LB containing 470 mM NaCl [27]. Moreover, morphological alteration from rod to coccoid was found in B. pseudomallei adaptation to high salt stress [43]. Changes from the rod to the coccoid form increase the cell membrane surface, which might benefit nutrient uptake by the bacterium [43]. B. pseudomallei also changes its membrane in response to salt stress. B. pseudomallei showed upregulated expression of Acyl-CoA dehydrogenase during high salt stress [27]. Acyl-CoA dehydrogenases are involved in changes in bacterial membrane fluidity during salt tolerance [44]. Acyl-CoA dehydrogenases may therefore play a role in adjusting the bacterial membrane lipid composition, modifying the types of fatty acids present, and altering the structures of phospholipids when B. pseudomallei encounters high salt levels.
The influences of salt stress on the pathogenicity of B. pseudomallei have been studied intensively [27, 28]. NaCl-exposed B. pseudomallei secreted many effector proteins, including the beta-lactamase-like protein, which led to greater survival after treatment with beta-lactam antibiotics [28]. Indeed, high salt stress resulted in the increased invasion of B. pseudomallei into A549 human lung respiratory epithelial cells, by increasing the expression and secretion of Bsa T3SS proteins [27]. Bsa T3SS is an important virulence factor for B. pseudomallei invasion and intracellular replication. High salt stress can increase the transcription of bipD and bopE genes, which encode the Bsa translocon component and the virulence-associated effector involved in actin dynamics, respectively. Besides, the increased Bsa T3SS may participate in the enhanced plaque formation of B. pseudomallei observed after exposure to NaCl [31].
An alternative model of T3SS triggering under salt stress has been linked with MucA-mediated coordination of alginate production in P. aeruginosa [45]. Alginate production is known to be activated by high salt conditions [46]. A comparison of global gene expression of mucA mutant- and wild-type strains under T3SS-inducing conditions showed the downregulation of T3SS genes and upregulation of genes involved in alginate biosynthesis. Under high salt conditions, the upregulation of sigma factor rpoE was observed in B. pseudomallei, suggesting a role for rpoE in tolerance to environmental stress [30]. Similarly, the upregulation of rpoE was observed; it was postulated to be involved in the regulation of T3SS in P. aeruginosa. Therefore, rpoE might play a role in controlling B. pseudomallei T3SS expression under high-salinity conditions, as described for P. aeruginosa.
B. pseudomallei can also alter bacterial metabolism under salt stress by upregulating the expression of short-chain dehydrogenase/oxidoreductase (SDO) [31]. SDO, an important enzyme in the metabolic pathways [47], catalyzes the NADPH-dependent reduction of many compounds, such as sugars, aldehydes, and ketones [48]. Recently, the induction of SDO activity during salt stress has been shown to be linked to the adaptation and pathogenesis of B. pseudomallei, by facilitating the invasion of host cells [31]. However, further experiments are required to investigate the underlying mechanism.
More recently, salt stress was found to increase thermal resistance, oxidative resistance, and plaque formation, while decreasing the motility of B. pseudomallei [32]. The resistance of B. pseudomallei to heat and oxidative stress may result from the increased gene expression of stress-response cellular components, such as sigma factor rpoE, and heat-shock proteins groEL and htpG in B. pseudomallei under high-salinity conditions [32]. Inactivation of the rpoE operon increased the susceptibility of B. pseudomallei to killing by menadione and hydrogen peroxide (H2O2) and high osmolarity [30]. Furthermore, it has been demonstrated that rpoE regulated a heat-inducible promoter of the rpoH gene in B. pseudomallei [49]. These data imply that RpoE plays an important role in the increased resistance of B. pseudomallei in response to heat and oxidative stress. Taken together, the evidence suggests that adaptive changes induced by salt stress may aid B. pseudomallei survival and/or persistence in various environments.
3.2. Adaptation to Oxidative Stress
Reactive oxygen species (ROS) can be generated by living organisms and chemical processes that occur in the environment. For example, H2O2 is produced by the oxidation of metals and sulfur species, or by UV radiation [50]. Some organic peroxides are produced by plant [51] and animal hosts [52] as defense mechanisms against microbial pathogens [53]. ROS play a role in controlling early B. pseudomallei infection by threatening and inhibiting the intracellular growth of B. pseudomallei. Thus, to survive, B. pseudomallei must possess a mechanism to adapt to this hostile factor, as shown in Figure 2.
Figure 2.
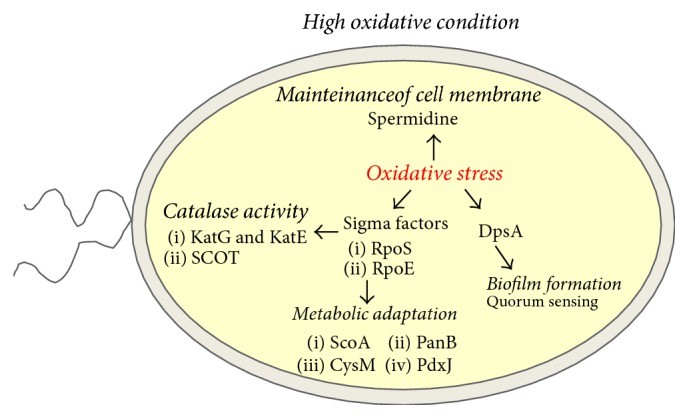
Mechanism of Burkholderia pseudomallei adaptation in response to oxidative stress.
The response of B. pseudomallei to oxidative stress is regulated by sigma (σ) factors [54], which are groups of proteins required for RNA synthesis. σ factors bind to the core of RNA polymerase to initiate RNA synthesis [55]. σ factors can be classified into 2 families: the σ 54 family and the σ 70 family [56]. Members of the σ 70 family are responsible for the expression of all essential genes, while members of the σ 54 family are mostly involved in nitrogen metabolism-associated genes. B. pseudomallei contains several σ factors, including RpoC (σC) [57], RpoN (σN) [58], RpoE (σE) [30], and RpoS (σS) [34]. RpoE and RpoS are members of the σ 70 family which play an important role in response to extracellular stress [55]. The rpoE gene of B. pseudomallei was activated during bacterial exposure to oxidative stress conditions [29]. When B. pseudomallei is exposed to H2O2-induced oxidative stress, the σE regulon turns on the expression of the speG gene involved in maintaining the levels of the polyamine, spermidine [29]. Spermidine helps B. pseudomallei to survive oxidative stress and plays vital roles in cell survival, by synchronizing biological processes such as Ca2+, Na+, and K+ -ATPase, to maintain membrane potential and control intracellular pH and volume during oxidative stress [59]. In addition to rpoE activation, the B. pseudomallei rpoS gene was activated during bacterial exposure to oxidative stress conditions [54]. RpoS controls the expression of genes encoding KatG and KatE catalase enzymes when B. pseudomallei is exposed to H2O2 [33]. RpoS also upregulates proteins involved in the response to oxidative stress, including succinyl-CoA: 3-ketoacid-coenzyme A transferase subunit A (ScoA), cysteine synthase B (CysM), 3-methyl-2-oxobutanoate hydroxymethyltransferase (PanB), and pyridoxal phosphate biosynthetic protein (PdxJ) and other proteins, which are universal-stress- and hypothetical oxidative-stress-responsive proteins [34]. When B. pseudomallei is exposed to oxidative stress, RpoS downregulates SCOT (a dimeric enzyme containing subunits A and B) expression to reduce endogenous ROS [34]. This mechanism enables the bacterium to reduce ROS intracellularly.
In addition to the genes and proteins regulated by σ factors mentioned above, the DNA-binding protein DpsA is involved in B. pseudomallei adaptation during exposure to oxidative stress [35]. DpsA plays a major role in protecting B. pseudomallei from oxidative stress through increased transcription of the katG (catalase peroxidase) promoter [60]. Moreover, dpsA gene expression is regulated in a cell population density-dependent manner via N-acylhomoserine lactone- (AHL-) dependent quorum sensing (QS). In several Gram-negative bacteria, QS is involved in biofilm formation, which is dependent on LuxI-type AHL synthases and LuxR-type transcriptional regulator proteins [35]. B. pseudomallei can produce biofilm, which may offer protection against hostile conditions, such as antibiotic treatment, salinity, and immune response [7, 8, 61]. Although it remains to be determined how B. pseudomallei triggers QS systems after exposure to oxidative stress, it is likely that biofilm formation and virulence-factor production are important survival mechanisms for B. pseudomallei in response to oxidative stress [35, 60].
3.3. Adaptation to Iron Concentrations
Iron is an essential microelement that contributes to the adaptation of B. pseudomallei to specific environmental niches, such as the soil and the host. The proposed adaptation of the B. pseudomallei response to iron content is shown in Figure 3. Iron plays a role as a cofactor of enzymes in cellular functions and metabolic processes. Therefore, an increase in iron concentration enhances the growth of B. pseudomallei [62], changes the bacterial morphology from rod form to coccoid form, and increases biofilm formation [61]. Furthermore, B. pseudomallei intracellular survival and MNGC formation cultured in A549 cell lines supplemented with iron are greater than in a non-iron-supplemented group [63]. The plaque-forming efficiency that indicates the severity of B. pseudomallei infected HeLa cells is increased in the presence of iron [63]. This raises the possibility that conditions with increased iron stores, such as thalassemia, are considered to increase the risk of acquiring melioidosis. In Thai adults, thalassemia was associated with an 11-fold increase in melioidosis compared with other patients with sepsis [64]. A recent study, reporting from the period from 2001 to 2010, showed that thalassemia was a major risk factor for melioidosis among Malaysian children [65]. Meanwhile, low-iron conditions were found to limit the growth of B. pseudomallei [63] and decrease the virulence of this bacterium [66]. A study of mice infected with B. pseudomallei showed that iron deprivation decreases bacterial load in visceral organs such as the lungs, liver, and spleen, which was associated with the improved survival of mice [66]. These studies indicate that iron is an important factor in B. pseudomallei infection.
Figure 3.
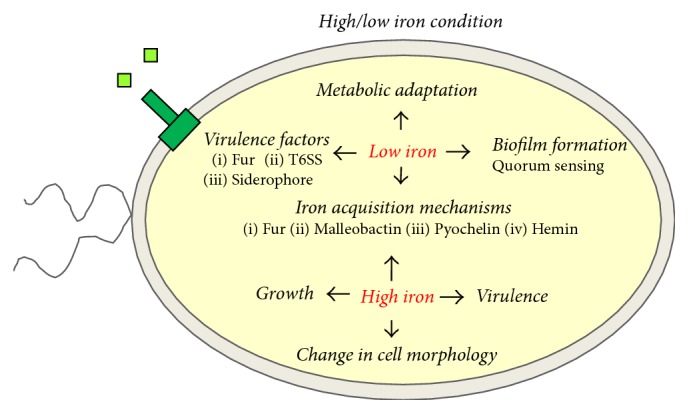
Mechanism of Burkholderia pseudomallei adaptation in response to iron content.
Generally, free iron is limited in physiological habitats and sequestered by the host by the iron‐binding proteins such as transferrin and lactoferrin. As a result, the bacterium must employ mechanisms of iron uptake regulation for survival under iron-restricted conditions. The iron regulators function under low-iron conditions by the expression of genes encoding an iron-acquisition system. Among these are the iron regulator gene “fur” (ferric uptake regulator), genes coding for iron-binding proteins, that is, siderophore (also called malleobactin), pyochelin, pyoverdine, ornibactin, cepabactin, and heme-hemin receptors, as well as a variety of genes involved in the metabolic pathway, that is, ferredoxin, NADH dehydrogenase, cytochrome oxidase, and ATP synthases [17, 37, 67].
Loprasert and coworkers reported that B. pseudomallei adapts itself in iron-limited conditions by upregulating the iron-acquisition system via the fur gene, which encodes a regulatory protein, Fur (ferric uptake regulatory) protein [36]. The Fur protein represses the transcription of iron-regulated promoters in response to increased intracellular iron concentrations. The Fur protein is also involved in the expression of toxins and bacterial virulence determinants in other bacteria [67, 68]. In B. pseudomallei, the Fur protein functions as a positive regulator of FeSOD (ferric-superoxide dismutase) and peroxidase to reduce free radicals and oxidative stress [36]. These enzymes influence the virulence of many bacteria [69]. However, the role of Fur in the virulence of B. pseudomallei has not, to date, been demonstrated.
The primary siderophore (malleobactin) plays an important role in iron uptake and regulation in B. pseudomallei. In addition to malleobactin, B. pseudomallei also produces many secondary siderophores, such as pyochelin, pyoverdine, and ornibactin, to control iron uptake [70]. Siderophores have been shown to correlate with the increased virulence of B. pseudomallei [71]. However, the mechanisms of these siderophores are still unclear in B. pseudomallei. In closely related B. cenocepacia, it has been reported that iron uptake via secondary siderophore, ornibactin, depends on the pvdA gene, encoding ornithine N5-oxygenase, and the orbA gene, encoding the outer membrane receptor [72]. An orbA is involved in ferric-ornibactin complex transport. Moreover, pvdA and orbA genes are required for the virulence of B. cenocepacia [72, 73].
Moreover, B. pseudomallei heme uptake (Bhu/Hmu) system encoded by BPSS0240–BPSS0244 genes was found to be upregulated during growth under low-iron conditions [17]. This system requires heme-hemin receptors that are present on the outer membrane of B. pseudomallei. In addition, the heme uptake system requires the action of the cytoplasmic membrane-anchored TonB-ExbB-ExbD complex to energize transport of these iron sources (ATP-binding cassette transporter systems) [70, 74]. The importance of the Bhu/Hmu system was investigated by Kvitko and coworkers [70], who showed that the deletion of the bhu/hmu locus affected the ability to utilize heme or hemoglobin as iron sources.
Under low-iron conditions, B. pseudomallei switched its metabolic pathways by obtaining energy from nitrogen metabolism and electron transport for survival [17]. It was found that BPSS0495, a gene encoding the nitroreductase enzyme responsible for nitrogen compound metabolism, was highly upregulated among B. pseudomallei grown in iron-restricted conditions. B. pseudomallei obtains energy from electron transport with the expression of bacterioferritin-associated ferredoxin genes under low-iron conditions. B. pseudomallei may use ferredoxin as an electron donor [17].
In addition to the iron-acquisition system, B. pseudomallei adapts its virulence-associated phenotypes during survival in low-iron conditions. One study reported that the biofilm formation-associated regulator (bfmR) gene was upregulated under low-iron conditions [38]. It is possible that B. pseudomallei adaptation might employ biofilm formation for survival [61]. This concurs with a previous study that found that the biofilm of B. pseudomallei increased bacterial adherence to host cells [75]. The T6SS genes, which encode proteins that facilitate cell-to-cell spreading, are reportedly induced by iron deprivation [76]. Taken together, it is reasonable to hypothesize that iron-acquisition mechanisms and T6SS might contribute to the control of B. pseudomallei adaptation after exposure to iron-limited conditions.
3.4. Adaptation in Host Cells
During the infection process, B. pseudomallei encounters various stress factors, such as nutrient restriction, oxygen limitations, and host defense mechanisms. Thus, B. pseudomallei must adapt itself to survive in the host using several mechanisms. Successful adaptation results in the survival of B. pseudomallei in a variety of phagocytic and nonphagocytic cells [77]. During B. pseudomallei survival in the host cells, several genes, including virulence factors, are functionally modulated [22, 78]. Several components of T3SS were found to be involved in many stages of B. pseudomallei pathogenesis, including invasion (BopB, BopC, BopE, BipB, BipC, BipD, and BsaZ), phagosome escape (BopC, BipC, BsaM, BsaQ, BsaU, and BsaZ), intracellular survival (BopA, BopB, BopC, BipC, BsaQ, and BsaZ) and cell–cell spreading (BipB, BipC, BsaS, BsaZ, and ChbP) [40, 79–89]. T6SS-1 was shown to modulate the intracellular growth of B. pseudomallei via the sensor regulators, BprC and VirA-VirG (VirAG) [90]. In addition to T3SS, T6SS plays a major role during bacterial transition from the phagosome to the cytosol [90]. Furthermore, the expression of bimA (Burkholderia intracellular motility A), which is translocated by the T5SS, was increased at 2 to 6 h after infection. B. pseudomallei BimA is required for intracellular actin-based motility and cell-to-cell spread [91]. In addition, B. pseudomallei modulates the bacterial surface structures to avoid host immune system recognition by downregulating genes involved in capsular polysaccharide biosynthesis, polysaccharide biosynthesis, LPS biosynthesis, flagella assembly, and chemotaxis during survival inside host cells [23].
σ factor genes were also found to be involved in B. pseudomallei survival in host cells. One of the B. pseudomalleiσ factor genes, rpoS, is reported to be a key regulator for intracellular survival under carbon starvation and oxidative stress [54]. In general, RpoS acts as a positive transcriptional regulator of oxyR and dpsA expression. Under oxidative stress, rpoS upregulated expression of oxyR and the katG–dpsA operon.
B. pseudomallei has various metabolic mechanisms to obtain the available host nutrients for its own proliferation. In a challenge study of oxygen-limited conditions, many genes of B. pseudomallei were induced. Among those were genes encoding proteins in arginine and pyruvate fermentation (aceE, arcD, and tatA), ATP synthases (atpA and atpD), electron transport proteins (aarC, cydA, cydB, mocA, and BPSL1260), flagella-mediated motility (flgA, flgC, flgK, flgM, fliF, fliJ, fliK, and pilT), stress-related proteins (clpB, rpoH, and rpoS), virulence factors (bopE, bipC, bipD, orgA, and pilA), and polyhydroxybutyrate synthase (bdhA-2). These findings suggest that B. pseudomallei presents an excellent transcriptional network that allows it to respond to conditions of limited oxygen [25]. Hypoxic conditions also lead to the repression of genes involved in ribosomal biogenesis, suggesting an overall reduction in protein synthesis during oxygen depletion, which is related to reduced bacterial growth rate [25].
In contrast to the challenge study, during the early stage of macrophage infection, a study has shown that genes involved in metabolism, glycolysis, and oxidative phosphorylation were downregulated while genes responsible for anaerobic metabolism, including pyruvate dehydrogenase, acetate kinase, and alcohol dehydrogenase, were induced [92]. This might be because the bacteria need to adjust their metabolism in response to the hypoxic conditions in the host cells [92]. Genes involved in benzoate degradation were also upregulated, suggesting that intracellular B. pseudomallei utilize aromatic compounds as a carbon source.
These findings demonstrate the importance of environmental or host conditions in the regulation of B. pseudomallei intracellular survival. However, the mechanism of regulation of gene expression requires further investigation.
4. Conclusions and Future Perspectives
This review outlines our current knowledge of the adaptive mechanisms that enable B. pseudomallei to survive and grow under various conditions, such as salinity, oxidative stress, altered iron concentrations, and host-associated conditions. Adaptations allow the organism to tolerate hostile environments and may also provide other advantages, such as increased bacterial virulence, evasion of host defenses, reduction in free radicals, and decreased growth rates for latent infections.
B. pseudomallei possesses several mechanisms by which it senses sources of stress in the environment and in the host, and then, depending on the type of stress, bacterial adaptation leads to the modulation of changes in the expression of the genes and proteins involved in metabolism, ion transport systems, and virulence factors. Increasing evidence strongly supports the adaptation of B. pseudomallei within the host, including pathways involved in environmental survival, which lead to bacterial persistence under adverse conditions. This insight is useful for understanding the underlying mechanisms that are important for the intracellular and extracellular adaptation of B. pseudomallei. This precise knowledge therefore opened the doors for novel targets for the treatment and prevention of melioidosis.
The potential sources of stress encountered by B. pseudomallei are not limited to those reviewed here. Further studies of B. pseudomallei adaptation under other stress conditions, such as acidity, osmotic stress, ammonia accumulation, antibacterial agent exposure, the presence of nitric oxide, and abscess condition, will also contribute to our understanding of bacterial survival and persistence. Other bacterial components that may be altered during B. pseudomallei adaptation following exposure to stress should also be investigated. The adaptation of B. pseudomallei to survival in ecological niches is a complex multifactorial process that depends on more than one environmental factor. However, currently, no reports show that B. pseudomallei can adapt in response to simultaneous exposure to multiple sources of stress. Such studies are needed to reflect actual environmental challenges and to provide a better understanding of B. pseudomallei survival and pathogenesis.
Acknowledgments
This work was supported by Mahidol University. Taksaon Duangurai is a scholar of Ph.D. student research assistantship of tropical medicine, Mahidol University. Pornpan Pumirat is a scholar of the Thailand Research Fund (MRG5580040) and the ICTM grant from the Faculty of Tropical Medicine, Mahidol University. The authors would like to thank Assist. Professor Usa Boonyuen for her insightful advice and Mr. Paul Adams and the Office of Research Services, Faculty of Tropical Medicine, Mahidol University, for proofreading the manuscript.
Conflicts of Interest
The authors of this work have no conflicts of interest to declare regarding the publication of this paper.
References
- 1.Brett P. J., Woods D. E. Pathogenesis of and immunity to melioidosis. Acta Tropica. 2000;74(2-3):201–210. doi: 10.1016/S0001-706X(99)00071-6. [DOI] [PubMed] [Google Scholar]
- 2.Whitmore A. An account of a glanders-like disease occurring in rangoon. Journal of Hygiene. 1913;13(1):1–34. doi: 10.1017/s0022172400005234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Samy R. P., Stiles B. G., Sethi G., Lim L. H. K. Melioidosis: clinical impact and public health threat in the tropics. PLOS Neglected Tropical Diseases. 2017;11(5) doi: 10.1371/journal.pntd.0004738.e0004738 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 4.Inglis T. J., Sousa A. Q. The public health implications of melioidosis. The Brazilian Journal of Infectious Diseases. 2009;13(1):59–66. doi: 10.1590/S1413-86702009000100013. [DOI] [PubMed] [Google Scholar]
- 5.White N. J. Melioidosis. The Lancet. 2003;361(9370):1715–1722. doi: 10.1016/S0140-6736(03)13374-0. [DOI] [PubMed] [Google Scholar]
- 6.Yabuuchi E., Kosako Y., Oyaizu H. Proposal of Burkholderia gen. Anov. and transfer of seven species of the genus Pseudomonas homology group II to the new genus, with the type species Burkholderia cepacia (Palleroni and Holmes 1981) comb: Nov. Microbiology and Immunology. 1992;36(12):1251–1275. doi: 10.1111/j.1348-0421.1992.tb02129.x. [DOI] [PubMed] [Google Scholar]
- 7.Cheng A. C., Currie B. J. Melioidosis: epidemiology, pathophysiology, and management. Clinical Microbiology Reviews. 2005;18(2):383–416. doi: 10.1128/CMR.18.2.383-416.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Inglis T. J. J., Sagripanti J.-L. Environmental factors that affect the survival and persistence of Burkholderia pseudomallei. Applied and Environmental Microbiology. 2006;72(11):6865–6875. doi: 10.1128/AEM.01036-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Liu X., Pang L., Sim S. H., et al. Association of melioidosis incidence with rainfall and humidity, Singapore, 2003–2012. Emerging Infectious Diseases. 2015;21(1):159–162. doi: 10.3201/eid2101.140042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Currie B. J., Jacups S. P. Intensity of rainfall and severity of melioidosis, Australia. Emerging Infectious Diseases. 2003;9(12):1538–1542. doi: 10.3201/eid0912.020750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kaestli M., Grist E. P. M., Ward L., Hill A., Mayo M., Currie B. J. The association of melioidosis with climatic factors in Darwin, Australia: a 23-year time-series analysis. Infection. 2016;72(6):687–697. doi: 10.1016/j.jinf.2016.02.015. [DOI] [PubMed] [Google Scholar]
- 12.Merritt A. J., Inglis T. J. The role of climate in the epidemiology of melioidosis. Current Tropical Medicine Reports. 2017;4(4):185–191. doi: 10.1007/s40475-017-0124-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Manivanh L., Pierret A., Rattanavong S., et al. Burkholderia pseudomallei in a lowland rice paddy: seasonal changes and influence of soil depth and physico-chemical properties. Scientific Reports. 2017;7(1, article no. 3031) doi: 10.1038/s41598-017-02946-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Tong S., Yang S., Lu Z., He W. Laboratory investigation of ecological factors influencing the environmental presence of Burkholderia pseudomallei. Microbiology and Immunology. 1996;40(6):451–453. doi: 10.1111/j.1348-0421.1996.tb01092.x. [DOI] [PubMed] [Google Scholar]
- 15.Hantrakun V., Rongkard P., Oyuchua M., et al. Soil nutrient depletion is associated with the presence of Burkholderia pseudomallei. Applied and Environmental Microbiology. 2016;82(24):7086–7092. doi: 10.1128/AEM.02538-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Musa H. I., Hassan L., Shamsuddin Z. H., Panchadcharam C., Zakaria Z., Aziz S. A. Physicochemical properties influencing presence of Burkholderia pseudomallei in soil from small ruminant farms in peninsular Malaysia. PLoS ONE. 2016;11(9) doi: 10.1371/journal.pone.0162348.e0162348 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Tuanyok A., Kim H. S., Nierman W. C., et al. Genome-wide expression analysis of iron regulation in Burkholderia pseudomallei and Burkholderia mallei using DNA microarrays. FEMS Microbiology Letters. 2005;252(2):327–335. doi: 10.1016/j.femsle.2005.09.043. [DOI] [PubMed] [Google Scholar]
- 18.Inglis T. J. J., Rigby P., Robertson T. A., Dutton N. S., Henderson M., Chang B. J. Interaction between Burkholderia pseudomallei and Acanthamoeba species results in coiling phagocytosis, endamebic bacterial survival, and escape. Infection and Immunity. 2000;68(3):1681–1686. doi: 10.1128/IAI.68.3.1681-1686.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Inglis T. J., Rodrigues F., Rigby P., Norton R., Currie B. J. Comparison of the susceptibilities of Burkholderia pseudomallei to meropenem and ceftazidime by conventional and intracellular methods. Antimicrobial Agents and Chemotherapy. 2004;48(8):2999–3005. doi: 10.1128/AAC.48.8.2999-3005.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Noinarin P., Chareonsudjai P., Wangsomnuk P., Wongratanacheewin S., Chareonsudjai S. Environmental free-living amoebae isolated from soil in Khon Kaen, Thailand, antagonize Burkholderia pseudomallei. PLoS ONE. 2016;11(11) doi: 10.1371/journal.pone.0167355.e0167355 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Holden M. T. G., Titball R. W., Peacock S. J., et al. Genomic plasticity of the causative agent of melioidosis, Burkholderia pseudomallei. Proceedings of the National Acadamy of Sciences of the United States of America. 2004;101(39):14240–14245. doi: 10.1073/pnas.0403302101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Willcocks S. J., Denman C. C., Atkins H. S., Wren B. W. Intracellular replication of the well-armed pathogen Burkholderia pseudomallei. Current Opinion in Microbiology. 2016;29:94–103. doi: 10.1016/j.mib.2015.11.007. [DOI] [PubMed] [Google Scholar]
- 23.Sun G. W., Chen Y., Liu Y., et al. Identification of a regulatory cascade controlling Type III Secretion System 3 gene expression in Burkholderia pseudomallei. Molecular Microbiology. 2010;76(3):677–689. doi: 10.1111/j.1365-2958.2010.07124.x. [DOI] [PubMed] [Google Scholar]
- 24.Pumpuang A., Chantratita N., Wikraiphat C., et al. Survival of Burkholderia pseudomallei in distilled water for 16 years. Transactions of the Royal Society of Tropical Medicine and Hygiene. 2011;105(10):598–600. doi: 10.1016/j.trstmh.2011.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hamad M. A., Austin C. R., Stewart A. L., Higgins M., Vázquez-Torres A., Voskuil M. I. Adaptation and antibiotic tolerance of anaerobic Burkholderia pseudomallei. Antimicrobial Agents and Chemotherapy. 2011;55(7):3313–3323. doi: 10.1128/AAC.00953-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.O’Rourke A., Yee N., Nierman W. C., Beyhan S. Environmental and genetic factors controlling Burkholderia pseudomallei persister phenotypes. Current Tropical Medicine Reports. 2017;4(3):111–116. doi: 10.1007/s40475-017-0116-4. [DOI] [Google Scholar]
- 27.Pumirat P., Cuccui J., Stabler R. A., et al. Global transcriptional profiling of Burkholderia pseudomallei under salt stress reveals differential effects on the Bsa type III secretion system. BMC Microbiology. 2010;10, article no. 171 doi: 10.1186/1471-2180-10-171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Pumirat P., Saetun P., Sinchaikul S., Chen S.-T., Korbsrisate S., Thongboonkerd V. Altered secretome of Burkholderia pseudomallei induced by salt stress. Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics. 2009;1794(6):898–904. doi: 10.1016/j.bbapap.2009.01.011. [DOI] [PubMed] [Google Scholar]
- 29.Jitprasutwit S., Ong C., Juntawieng N., et al. Transcriptional profiles of Burkholderia pseudomallei reveal the direct and indirect roles of Sigma E under oxidative stress conditions. BMC Genomics. 2014;15(1, article no. 787) doi: 10.1186/1471-2164-15-787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Korbsrisate S., Vanaporn M., Kerdsuk P., et al. The Burkholderia pseudomallei RpoE (AlgU) operon is involved in environmental stress tolerance and biofilm formation. FEMS Microbiology Letters. 2005;252(2):243–249. doi: 10.1016/j.femsle.2005.09.002. [DOI] [PubMed] [Google Scholar]
- 31.Pumirat P., Boonyuen U., Vanaporn M., et al. The role of short-chain dehydrogenase/oxidoreductase, induced by salt stress, on host interaction of B. pseudomallei. BMC Microbiology. 2014;14(1, article no. 1) doi: 10.1186/1471-2180-14-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Pumirat P., Vanaporn M., Boonyuen U., Indrawattana N., Rungruengkitkun A., Chantratita N. Effects of sodium chloride on heat resistance, oxidative susceptibility, motility, biofilm and plaque formation of Burkholderia pseudomallei. MicrobiologyOpen. 2017;6(4) doi: 10.1002/mbo3.493.e00493 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Jangiama W., Lopraserf S., Tungpradabkula S. Role of Burkholderia pseudomallei RpoS in regulation of catalase activities under hydrogen peroxide induction. ScienceAsia. 2008;34(1):23–29. doi: 10.2306/scienceasia1513-1874.2008.34.023. [DOI] [Google Scholar]
- 34.Chutoam P., Charoensawan V., Wongtrakoongate P., Kum-arth A., Buphamalai P., Tungpradabkul S. RpoS and oxidative stress conditions regulate succinyl-CoA: 3-ketoacid-coenzyme A transferase (SCOT) expression in Burkholderia pseudomallei. Microbiology and Immunology. 2013;57(9):605–615. doi: 10.1111/1348-0421.12077. [DOI] [PubMed] [Google Scholar]
- 35.Lumijiaktase P., Diggle S. P., Loprasert S., et al. Quorum sensing regulates dpsA and the oxidative stress response in Burkholderia pseudomallei. Microbiology. 2006;152(12):3651–3659. doi: 10.1099/mic.0.29226-0. [DOI] [PubMed] [Google Scholar]
- 36.Loprasert S., Sallabhan R., Whangsuk W., Mongkolsuk S. Characterization and mutagenesis of fur gene from Burkholderia pseudomallei. Gene. 2000;254(1-2):129–137. doi: 10.1016/S0378-1119(00)00279-1. [DOI] [PubMed] [Google Scholar]
- 37.Ratledge C., Dover L. G. Iron metabolism in pathogenic bacteria. Annual Review of Microbiology. 2000;54:881–941. doi: 10.1146/annurev.micro.54.1.881. [DOI] [PubMed] [Google Scholar]
- 38.Tabunhan S., Wongratanacheewin S., Wongwajana S., Welbat T. U. M., Faksri K., Namwat W. Characterization of a novel two-component system response regulator involved in biofilm formation and a low-iron response of Burkholderia pseudomallei. The Southeast Asian Journal of Tropical Medicine and Public Health. 2014;45(5):1065–1079. [PubMed] [Google Scholar]
- 39.Felgner P. L., Kayala M. A., Vigil A., et al. A Burkholderia pseudomallei protein microarray reveals serodiagnostic and cross-reactive antigens. Proceedings of the National Acadamy of Sciences of the United States of America. 2009;106(32):13499–13504. doi: 10.1073/pnas.0812080106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Vander Broek C. W., Chalmers K. J., Stevens M. P., Stevens J. M. Quantitative proteomic analysis of Burkholderia pseudomallei Bsa type III secretion system effectors using hypersecreting mutants. Molecular and Cellular Proteomics. 2015;14(4):905–916. doi: 10.1074/mcp.M114.044875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wang L. C., Morgan L. K., Godakumbura P., Kenney L. J., Anand G. S. The inner membrane histidine kinase EnvZ senses osmolality via helix-coil transitions in the cytoplasm. EMBO Journal. 2015;34(19):p. 2481. doi: 10.15252/embj.201592753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kimura Y., Ohtani M., Takegawa K. An adenylyl cyclase, CyaB, acts as an osmosensor in Myxococcus xanthus. Journal of Bacteriology. 2005;187(10):3593–3598. doi: 10.1128/JB.187.10.3593-3598.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kamjumphol W., Chareonsudjai P., Taweechaisupapong S., Chareonsudjai S. Morphological alteration and survival of Burkholderia pseudomallei in soil microcosms. The American Journal of Tropical Medicine and Hygiene. 2015;93(5):1058–1065. doi: 10.4269/ajtmh.15-0177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Zhang Y.-X., Denoya C. D., Skinner D. D., et al. Genes encoding acyl-CoA dehydrogenase (AcdH) homologues from Streptomyces coelicolor and Streptomyces avermitilis provide insights into the metabolism of small branched-chain fatty acids and macrolide antibiotic production. Microbiology. 1999;145(9):2323–2334. doi: 10.1099/00221287-145-9-2323. [DOI] [PubMed] [Google Scholar]
- 45.Wu W., Badrane H., Arora S., Baker H. V., Jin S. MucA-mediated coordination of type III secretion and alginate synthesis in Pseudomonas aeruginosa. Journal of Bacteriology. 2004;186(22):7575–7585. doi: 10.1128/JB.186.22.7575-7585.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Berry A., DeVault J. D., Chakrabarty A. M. High osmolarity is a signal for enhanced algD transcription in mucoid and nonmucoid Pseudomonas aeruginosa strains. Journal of Bacteriology. 1989;171(5):2312–2317. doi: 10.1128/jb.171.5.2312-2317.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kavanagh K. L., Jörnvall H., Persson B., Oppermann U. Medium- and short-chain dehydrogenase/reductase gene and protein families: the SDR superfamily: functional and structural diversity within a family of metabolic and regulatory enzymes. Cellular and Molecular Life Sciences. 2008;65(24):3895–3906. doi: 10.1007/s00018-008-8588-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Oppermann U., Filling C., Hult M., et al. Short-chain dehydrogenases/reductases (SDR): the 2002 update. Chemico-Biological Interactions. 2003;143-144:247–253. doi: 10.1016/S0009-2797(02)00164-3. [DOI] [PubMed] [Google Scholar]
- 49.Vanaporn M., Vattanaviboon P., Thongboonkerd V., Korbsrisate S. The rpoE operon regulates heat stress response in Burkholderia pseudomallei. FEMS Microbiology Letters. 2008;284(2):191–196. doi: 10.1111/j.1574-6968.2008.01216.x. [DOI] [PubMed] [Google Scholar]
- 50.Fu H., Yuan J., Gao H. Microbial oxidative stress response: novel insights from environmental facultative anaerobic bacteria. Archives of Biochemistry and Biophysics. 2015;584, article no. 7047:28–35. doi: 10.1016/j.abb.2015.08.012. [DOI] [PubMed] [Google Scholar]
- 51.Jwa N.-S., Agrawal G. K., Tamogami S., et al. Role of defense/stress-related marker genes, proteins and secondary metabolites in defining rice self-defense mechanisms. Plant Physiology and Biochemistry. 2006;44(5-6):261–273. doi: 10.1016/j.plaphy.2006.06.010. [DOI] [PubMed] [Google Scholar]
- 52.Huang J., Canadien V., Lam G. Y., et al. Activation of antibacterial autophagy by NADPH oxidases. Proceedings of the National Acadamy of Sciences of the United States of America. 2009;106(15):6226–6231. doi: 10.1073/pnas.0811045106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Vatansever F., de Melo W. C. M. A., Avci P., et al. Antimicrobial strategies centered around reactive oxygen species—bactericidal antibiotics, photodynamic therapy, and beyond. FEMS Microbiology Reviews. 2013;37(6):955–989. doi: 10.1111/1574-6976.12026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Subsin B., Thomas M. S., Katzenmeier G., Shaw J. G., Tungpradabkul S., Kunakorn M. role of the stationary growth phase sigma factor RpoS of Burkholderia pseudomallei in response to physiological stress conditions. Journal of Bacteriology. 2003;185(23):7008–7014. doi: 10.1128/JB.185.23.7008-7014.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Davis M. C., Kesthely C. A., Franklin E. A., MacLellan S. R. The essential activities of the bacterial sigma factor. Canadian Journal of Microbiology. 2017;63(2):89–99. doi: 10.1139/cjm-2016-0576. [DOI] [PubMed] [Google Scholar]
- 56.Feklístov A., Sharon B. D., Darst S. A., Gross C. A. Bacterial sigma factors: A historical, structural, and genomic perspective. Annual Review of Microbiology. 2014;68:357–376. doi: 10.1146/annurev-micro-092412-155737. [DOI] [PubMed] [Google Scholar]
- 57.Yam H., Rahim A. A., Mohamad S., et al. The multiple roles of hypothetical gene BPSS1356 in Burkholderia pseudomallei. PLoS ONE. 2014;9(6) doi: 10.1371/journal.pone.0099218.e99218 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Diep D. T., Phuong N. T., Hlaing M. M., Srimanote P., Tungpradabkul S. Role of Burkholderia pseudomallei Sigma N2 in Amino Acids Utilization and in Regulation of Catalase E Expression at the Transcriptional Level. International Journal of Bacteriology. 2015;2015:1–10. doi: 10.1155/2015/623967.623967 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Pegg A. E. Mammalian polyamine metabolism and function. IUBMB Life. 2009;61(9):880–894. doi: 10.1002/iub.230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Loprasert S., Whangsuk W., Sallabhan R., Mongkolsuk S. Regulation of the katG-dpsA operon and the importance of KatG in survival of Burkholderia pseudomallei exposed to oxidative stress. FEBS Letters. 2003;542(1-3):17–21. doi: 10.1016/S0014-5793(03)00328-4. [DOI] [PubMed] [Google Scholar]
- 61.Kamjumphol W., Chareonsudjai S., Chareonsudjai P., Wongratanacheewin S., Taweechaisupapong S. Environmental factors affecting Burkholderia pseudomallei biofilm formation. Southeast Asian Journal of Tropical Medicine and Public Health. 2013;44(1):72–81. [PubMed] [Google Scholar]
- 62.Gerhardy B., Simpson G. Melioidosis and idiopathic pulmonary hemosiderosis: a cast-iron case. Respirology Case Reports. 2013;1(2):46–47. doi: 10.1002/rcr2.25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Amornrit W., Muangsombut V., Wangteeraprasert T., Korbsrisate S. Elevated intracellular levels of iron in host cells promotes Burkholderia pseudomallei infection. Asian Biomedicine. 2012;6(3):465–471. doi: 10.5372/1905-7415.0603.078. [DOI] [Google Scholar]
- 64.Suputtamongkol Y., Chaowagul W., Chetchotisakd P., et al. Risk factors for melioidosis and bacteremic melioidosis. Clinical Infectious Diseases. 1999;29(2):408–413. doi: 10.1086/520223. [DOI] [PubMed] [Google Scholar]
- 65.Fong S. M., Wong K. J., Fukushima M., Yeo T. W. Thalassemia major is a major risk factor for pediatric melioidosis in Kota Kinabalu, Sabah, Malaysia. Clinical Infectious Diseases. 2015;60(12):1802–1807. doi: 10.1093/cid/civ189. [DOI] [PubMed] [Google Scholar]
- 66.Schmidt I. H., Gildhorn C., Böning M. A., et al. Burkholderia pseudomallei modulates host iron homeostasis to facilitate iron availability and intracellular survival. PLOS Neglected Tropical Diseases. 2018;12(1):p. e0006096. doi: 10.1371/journal.pntd.0006096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Litwin C. M., Calderwood S. B. Role of iron in regulation of virulence genes. Clinical Microbiology Reviews. 1993;6(2):137–149. doi: 10.1128/CMR.6.2.137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Runyen-Janecky L., Daugherty A., Lloyd B., Wellington C., Eskandarian H., Sagransky M. Role and regulation of iron-sulfur cluster biosynthesis genes in Shigella flexneri virulence. Infection and Immunity. 2008;76(3):1083–1092. doi: 10.1128/IAI.01211-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Dacanay A., Johnson S. C., Bjornsdottir R., et al. Molecular characterization and quantitative analysis of superoxide dismutases in virulent and avirulent strains of Aeromonas salmonicida subsp. salmonicida. Journal of Bacteriology. 2003;185(15):4336–4344. doi: 10.1128/JB.185.15.4336-4344.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Kvitko B. H., Goodyear A., Propst K. L., Dow S. W., Schweizer H. P. Burkholderia pseudomallei known siderophores and hemin uptake are dispensable for lethal murine melioidosis. PLOS Neglected Tropical Diseases. 2012;6(6) doi: 10.1371/journal.pntd.0001715.e1715 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Butt A. T., Thomas M. S. Iron acquisition mechanisms and their role in the virulence of Burkholderia species. Frontiers in Cellular and Infection Microbiology. 2017;7, article no. 460 doi: 10.3389/fcimb.2017.00460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Sokol P. A., Darling P., Woods D. E., Mahenthiralingam E., Kooi C. Role of ornibactin biosynthesis in the virulence of Burkholderia cepacia: characterization of pvdA, the gene encoding L-ornithine N5- oxygenase. Infection and Immunity. 1999;67(9):4443–4455. doi: 10.1128/iai.67.9.4443-4455.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Visser M. B., Majumdar S., Hani E., Sokol P. A. Importance of the ornibactin and pyochelin siderophore transport systems in Burkholderia cenocepacia lung infections. Infection and Immunity. 2004;72(5):2850–2857. doi: 10.1128/IAI.72.5.2850-2857.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Harland D. N., Dassa E., Titball R. W., Brown K. A., Atkins H. S. ATP-binding cassette systems in Burkholderia pseudomallei and Burkholderia mallei. BMC Genomics. 2007;8, article no. 83 doi: 10.1186/1471-2164-8-83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Kunyanee C., Kamjumphol W., Taweechaisupapong S., et al. Burkholderia pseudomallei biofilm promotes adhesion, internalization and stimulates proinflammatory cytokines in human epithelial A549 cells. PLoS ONE. 2016;11(8) doi: 10.1371/journal.pone.0160741.e0160741 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Burtnick M. N., Brett P. J. Burkholderia mallei and Burkholderia pseudomallei cluster 1 type VI secretion system gene expression is negatively regulated by iron and Zinc. PLoS ONE. 2013;8(10) doi: 10.1371/journal.pone.0076767.e76767 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Valvano M. A., Keith K. E., Cardona S. T. Survival and persistence of opportunistic Burkholderia species in host cells. Current Opinion in Microbiology. 2005;8(1):99–105. doi: 10.1016/j.mib.2004.12.002. [DOI] [PubMed] [Google Scholar]
- 78.Moule M. G., Spink N., Willcocks S., et al. Characterization of new virulence factors involved in the intracellular growth and survival of Burkholderia pseudomallei. Infection and Immunity. 2016;84(3):701–710. doi: 10.1128/IAI.01102-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Stevens M. P., Haque A., Atkins T., et al. Attenuated virulence and protective efficacy of a Burkholderia pseudomallei bsa type III secretion mutant in murine models of melioidosis. Microbiology. 2004;150(8):2669–2676. doi: 10.1099/mic.0.27146-0. [DOI] [PubMed] [Google Scholar]
- 80.Pumirat P., Broek C. V., Juntawieng N., et al. Analysis of the prevalence, secretion and function of a cell cycle-inhibiting factor in the melioidosis pathogen Burkholderia pseudomallei. PLoS ONE. 2014;9(5) doi: 10.1371/journal.pone.0096298.e96298 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 81.Stevens M. P., Wood M. W., Taylor L. A., et al. An Inv/Mxi-Spa-like type III protein secretion system in Burkholderia pseudomallei modulates intracellular behaviour of the pathogen. Molecular Microbiology. 2002;46(3):649–659. doi: 10.1046/j.1365-2958.2002.03190.x. [DOI] [PubMed] [Google Scholar]
- 82.Burtnick M. N., Brett P. J., Nair V., Warawa J. M., Woods D. E., Gherardini F. C. Burkholderia pseudomallei type III secretion system mutants exhibit delayed vacuolar escape phenotypes in RAW 264.7 murine macrophages. Infection and Immunity. 2008;76(7):2991–3000. doi: 10.1128/IAI.00263-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Teh B. E., French C. T., Chen Y., et al. Type three secretion system-mediated escape of Burkholderia pseudomallei into the host cytosol is critical for the activation of NFκB. BMC Microbiology. 2014;14(1, article no. 115) doi: 10.1186/1471-2180-14-115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Pilatz S., Breitbach K., Hein N., et al. Identification of Burkholderia pseudomallei genes required for the intracellular life cycle and in vivo virulence. Infection and Immunity. 2006;74(6):3576–3586. doi: 10.1128/IAI.01262-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.French C. T., Toesca I. J., Wu T.-H., et al. Dissection of the Burkholderia intracellular life cycle using a photothermal nanoblade. Proceedings of the National Acadamy of Sciences of the United States of America. 2011;108(29):12095–12100. doi: 10.1073/pnas.1107183108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Muangsombut V., Suparak S., Pumirat P., et al. Inactivation of Burkholderia pseudomallei bsaQ results in decreased invasion efficiency and delayed escape of bacteria from endocytic vesicles. Archives of Microbiology. 2008;190(6):623–631. doi: 10.1007/s00203-008-0413-3. [DOI] [PubMed] [Google Scholar]
- 87.Chen Y., Schröder I., French C. T., et al. Characterization and analysis of the Burkholderia pseudomallei BsaN virulence regulon. BMC Microbiology. 2014;14(1, article no. 206) doi: 10.1186/s12866-014-0206-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Srinon V., Muangman S., Imyaem N., et al. Comparative assessment of the intracellular survival of the Burkholderia pseudomallei bopC mutant. Journal of Microbiology. 2013;51(4):522–526. doi: 10.1007/s12275-013-2557-3. [DOI] [PubMed] [Google Scholar]
- 89.Cullinane M., Gong L., Li X., et al. Stimulation of autophagy suppresses the intracellular survival of Burkholderia pseudomallei in mammalian cell lines. Autophagy. 2008;4(6):744–753. doi: 10.4161/auto.6246. [DOI] [PubMed] [Google Scholar]
- 90.Chen Y., Wong J., Sun G. W., Liu Y., Tan G.-Y. G., Gan Y.-H. Regulation of type VI secretion system during Burkholderia pseudomallei infection. Infection and Immunity. 2011;79(8):3064–3073. doi: 10.1128/IAI.05148-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Sitthidet C., Korbsrisate S., Layton A. N., Field T. R., Stevens M. P., Stevens J. M. Identification of motifs of Burkholderia pseudomallei BimA required for intracellular motility, actin binding, and actin polymerization. Journal of Bacteriology. 2011;193(8):1901–1910. doi: 10.1128/JB.01455-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Chieng S., Carreto L., Nathan S. Burkholderia pseudomallei transcriptional adaptation in macrophages. BMC Genomics. 2012;13, article 328 doi: 10.1186/1471-2164-13-328. [DOI] [PMC free article] [PubMed] [Google Scholar]