

Abstract
American Foulbrood is a worldwide distributed, fatal disease of the brood of the Western honey bee (Apis mellifera). The causative agent of this fatal brood disease is the Gram-positive, spore-forming bacterium Paenibacillus larvae, which can be classified into four different genotypes (ERIC I-IV), with ERIC I and II being the ones isolated from contemporary AFB outbreaks. P. larvae is a peritrichously flagellated bacterium and, hence, we hypothesized that P. larvae is capable of coordinated and cooperative multicellular behaviors like swarming motility and biofilm formation. In order to analyze these behaviors of P. larvae, we firstly established appropriate functional assays. Using these assays we demonstrated that P. larvae ERIC II, but not P. larvae ERIC I, was capable of swarming. Swarming motility was hampered in a P. larvae ERIC II-mutant lacking production of paenilarvin, an iturin-like lipopeptide exclusively expressed by this genotype. Both genotypes were able to form free floating biofilm aggregates loosely attached to the walls of the culture wells. Visualizing the biofilms by Congo red and thioflavin S staining suggested structural differences between the biofilms formed. Biofilm formation was shown to be independent from paenilarvin production because the paenilarvin deficient mutant was comparably able to form a biofilm.
Introduction
The Western honey bee Apis mellifera is a generalist pollinator and managed colonies of A. mellifera are widely used in global agriculture for the pollination of many crops and fruit grown in the open field1,2. In addition, A. mellifera also pollinates numerous wild flowers thus contributing to biodiversity in natural ecosystems. Thus, infectious diseases threatening the performance and survival of individual honey bees and honey bee colonies3 are of great concern not only for farmers and beekeepers but also for the general public. Among the pathogens posing the most serious threats to honey bees is Paenibacillus larvae. This Gram-positive, spore-forming bacterium is the causative agent of American Foulbrood (AFB) of honey bees4. P. larvae only infects bee larvae, hence, AFB only affects the bee brood but still the disease is able to kill entire colonies if left untreated5. P. larvae and AFB are globally distributed in A. mellifera populations. AFB is highly contagious and it may spread quite fast within and between honey bee colonies and apiaries. In most countries, it is classified as notifiable disease; control measures are regulated by corresponding laws and often include culling of diseased colonies.
The species P. larvae comprises the four genotypes ERIC I–IV which have been defined by repetitive element PCR (repPCR) performed with primers amplifying enterobacterial repetitive intergenic consensus elements (ERIC primers)4. However, other methods like MALDI-ToF analysis6, multi locus sequence type (MLST)7 or multiple locus variable number of tandem repeat analysis (MLVA)8 are also able to differentiate these four genotypes. This genotype differentiation is practically relevant because the four genotypes differ in several phenotypic features4,9 including virulence at both the level of the individual larva and the colony10,11.
Contemporary outbreaks of AFB all over the world are caused by two of the four genotypes only, by P. larvae ERIC I and ERIC II7. Therefore, these two genotypes have received much attention in recent research. Genome analysis of a non-genotyped strain of P. larvae provided first indications of possible virulence factors12, but it was the comparative genome13,14 and proteome analysis15 that yielded promising candidates, which could then be confirmed experimentally as virulence factors. On the basis of the previous and newly gained knowledge on P. larvae, the following picture emerges of the pathogenesis of P. larvae infections: In individual larvae, the infection process starts with the oral uptake of P. larvae spores during the first 36 hours after egg hatching. After ingestion, the spores germinate in the midgut lumen. The vegetative bacteria proliferate massively until they fill the entire gut lumen16. Eventually, the bacteria start to attack the midgut epithelium with the help of various virulence factors. The chitin-degrading enzyme PlCBP49 is a key virulence factor for both genotypes and is responsible for the degradation of the peritrophic matrix that is supposed to protect the epithelium from attack by pathogens17,18. The surface layer protein SplA is specifically expressed by P. larvae ERIC II and mediates bacterial adhesion to the midgut epithelium, a step that is obviously relevant for the pathogenic strategy of P. larvae ERIC II15,19. The two toxins Plx1 and Plx2 exclusively expressed by P. larvae ERIC I were shown to be relevant for the virulence of this genotype20,21. For the recently described toxin C3larvin22 encoded by the loci Tx7 and TxIII in the genomes of P. larvae ERIC I and ERIC II13, respectively, it has yet to be shown experimentally that it is a virulence factor. The aforementioned virulence factors are involved when P. larvae attacks and breaches the midgut epithelium and invades the hemocoel thereby killing the larva. Subsequently, the larval cadaver is degraded by P. larvae to a ropy mass which dries out to a scale. While the ropy mass can still be removed from the brood cell, the scale tightly adheres to the cell wall and resists the cleaning attempts of the bees.
When searching for virulence factors, the non-ribosomal peptides (NRP) and peptide-polyketide (NRP/PK) hybrids of P. larvae, the biosynthetic machineries of which are encoded by complex giant gene clusters23, attracted special attention. The novel tripeptide sevadicin produced by P. larvae ERIC II has antibacterial activity24. Another novel NRP/PK hybrid molecule is paenilamicin, produced by P. larvae ERIC I and ERIC II, which is active against some bacteria and fungi25 and plays a role in outcompeting microbial competitors of P. larvae in the larval midgut26. Bacillibactin, a catechol-type siderophore known from members of the Bacillus cereus sensu lato group and B. subtilis, is synthetized by both P. larvae genotypes only under iron depletion conditions27. In addition, P. larvae ERIC II non-ribosomally produces a group of lipopeptides belonging to the iturin-family, the so-called paenilarvins28. Paenilarvins have strong antifungal activity28 but lack cytotoxicity29. Hence, although a direct role as virulence factor during the invasive phase of pathogenesis could not be established for the paenilarvins29, they might be involved in outcompeting fungal competitors of P. larvae during the bacterial lifecycle in infected larvae.
P. larvae has been described as peritrichously flagellated and highly motile4 and, indeed, genes for building flagella were found in its genome12 suggesting that P. larvae is capable of coordinated activity like swarming motility and biofilm formation. Swarming motility is defined as a coordinated, flagella-driven movement of a collective group of bacteria across a surface30. A variety of Gram-negative and Gram-positive bacterial species exhibit swarming motility31. Within the genus Paenibacillus, several flagellated species like P. alvei32, P. dendritiformis33, and P. vortex34 show swarming behaviour. Swarming is generally thought to facilitate rapid colonization of nutrient-rich environments and to accelerate biomass production31. Peritrichously arranged flagella and the tightly controlled production of an extracellular matrix consisting of polysaccharides, biosurfactants, peptides, and proteins are required in many cases for swarming motility to occur35.
Many bacterial species are able to switch between planktonic growth and biofilm formation. The broadest definition of a biofilm is that it represents cooperatively acting microorganisms forming cell clusters which are held together and protected from adverse external influence by an extracellular matrix secreted by the members of the biofilm. This matrix is a characteristic hallmark of bacterial biofilms and consists of exopolysaccharides, secreted proteins, amyloid fibres, and sometimes nucleic acids36,37 (and references therein). Sessile biofilms are found attached to both biotic or abiotic surfaces38,39. However, bacteria can also form floating biofilms (‘pellicles’) at air-liquid interfaces38 or even form free floating biofilms40. For the formation of sessile biofilms, different developmental stages are described starting from the initially reversible, later then irreversible surface attachment of planktonic cells followed by the formation of microcolonies until the final macrocolonies are formed. From these macrocolonies, cells are dispersed to become planktonic again or the entire macrocolony can detach or dissolve39. Biofilms are of tremendous medical relevance because they not only protect bacteria from the host’s immune system but also render them refractory against antimicrobial treatment41–44. Not surprisingly though, that biofilms are considered the preferred lifestyle of bacteria when living in a host or on host tissues45.
Swarming and biofilm formation might be important during various stages of the lifecycle of P. larvae in infected larvae. It was recently speculated that exopolymeric substances (EPS) as structural components of the extracellular matrix during biofilm development could possibly be involved in virulence and tolerance to physiological stress in P. larvae46. However, no data actually proving biofilm formation or substantiating the existence and role of P. larvae biofilms in virulence was presented. Therefore, neither swarming nor biofilm formation have been demonstrated for this bacterial species so far. We here present our data showing that P. larvae ERIC II but not P. larvae ERIC I is capable of swarming motility. We provide evidence that paenilarvin, the recently identified iturin-like secondary metabolite of P. larvae ERIC II28,29, is involved in swarming motility. We also demonstrate that both genotypes indeed form biofilms but that their formation is not influenced by paenilarvin. These assays for P. larvae swarming motility and biofilm formation address two hitherto unknown characteristics of this bacterium which might be relevant for P. larvae when it is colonizing the infected or degrading the dead larva. Hence, understanding P. larvae swarming motility and biofilm formation might contribute to a better understanding of the disease processes and pave the way for novel treatment strategies.
Results
Swarming motility of P. larvae
To test the hypothesis that P. larvae is able to perform a multicellular flagellum-driven surface movement known as swarming motility, we first established an appropriate experimental procedure based on soft agar plates (0.5%) made from brain heart infusion broth (BHI) and on inoculating 10 µl of a bacterial suspension with an OD600 of 0.1 centrally onto the agar plate. While P. larvae ERIC I did not show swarming motility (Fig. 1A), P. larvae ERIC II reproducibly exhibited swarming motility under these conditions (Fig. 1A). Time course experiments performed by measuring the swarm radii at different time points (Fig. 1B) revealed that P. larvae ERIC II was able to cover the entire surface of the plate within three days (swarm radius 47 mm). In contrast, the macrocolonies formed by P. larvae ERIC I had reached a size of only 12.91 mm ± 0.29 mm (mean ± SEM) even after seven days. The radii of the area covered by P. larvae ERIC II were significantly different from those covered P. larvae ERIC I for all time points (p < 0.0001, unpaired Student’s t-test). Next, we randomly selected another five field strains of each P. larvae genotype and tested their swarming behaviour. All P. larvae ERIC I strains formed macrocolonies but were not able to swarm while all tested P. larvae ERIC II strains nearly covered the entire surfaces of the plates within two days (Fig. 2). These results suggested that swarming behaviour is another feature distinguishing these two genotypes.
Figure 1.
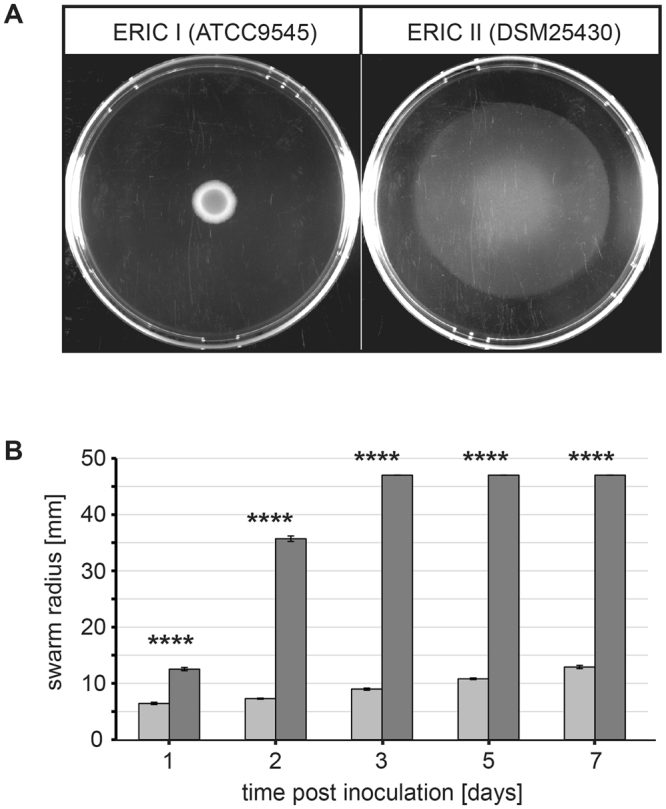
Swarming motility of P. larvae ERIC I (ATCC9545) and P. larvae ERIC II (DSM25430). Bacterial suspensions (10 µl) with an OD600 of 0.1 were inoculated centrally onto brain heart infusion (BHI; 0.5%) agar plates and incubated for two days under aerobic conditions at 37 °C. For both genotypes, three biological replicates with three technical replicates each were performed. (A) A representative plate is shown for each strain. Uncolonized agar is black and bacterial biomass is milky white to white. (B) Swarming motility was monitored over time and swarm radii were measured for each strain at one, two, three, five, and seven days post inoculation. Three biological replicates with three technical replicates each were performed. Values are given as mean ± SEM. Statistical analysis of the difference in swarming motility between ATCC9545 (light gray columns) and DSM25430 (dark gray columns) was performed with an unpaired Student’s t-test for each time point (not significantly different: n.s., p ≥ 0.05; significantly different: *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001).
Figure 2.
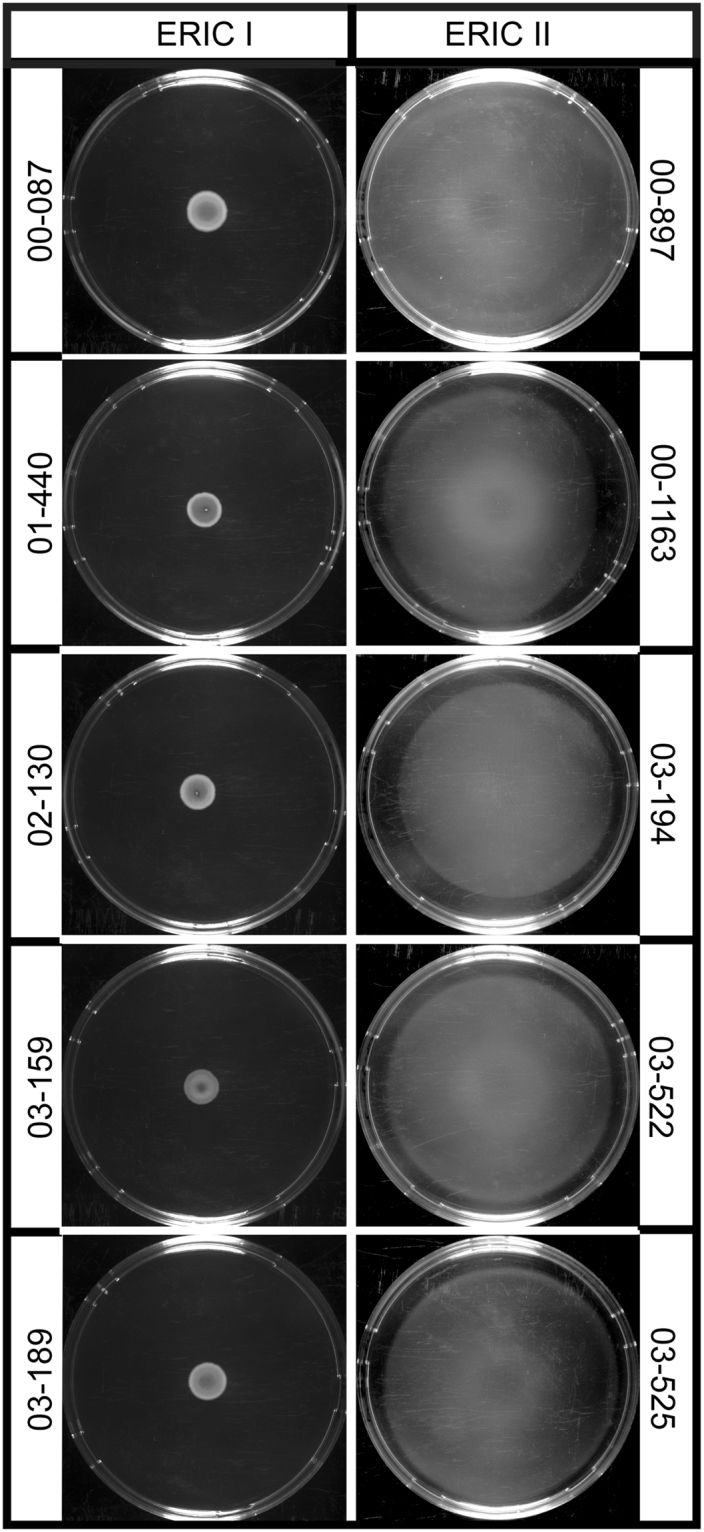
Swarming motility of field isolates of P. larvae ERIC I and P. larvae ERIC II. Swarming assays were performed with five strains each of P. larvae ERIC I and ERIC II which had been isolated from contemporary AFB outbreaks (field isolates). For all strains, three biological replicates with three technical replicates each were performed and a representative plate is shown for each strain. Uncolonized agar is black and bacterial biomass is milky white to white. Swarming was only evident for P. larvae ERIC II strains.
Biofilm formation of P. larvae
To test the hypothesis that P. larvae is capable of forming multicellular aggregates known as biofilms we established culture conditions that resulted in the formation of free floating bacterial aggregates that were visible to the naked eye and resembled biofilms (Fig. 3A,B). These aggregates reproducibly formed after five to six days when bacterial suspensions were incubated without agitation (static culture) at 37 °C whereas no such aggregates were visible in the negative controls (Fig. 3C). While the aggregates formed by P. larvae ERIC I appeared “hairy” with protrusions attaching all around to the wall of the well (Fig. 3A), the P. larvae ERIC II aggregates had a rather laminar appearance (Fig. 3B).
Figure 3.

Biofilm formation of P. larvae ERIC I (ATCC9545) and P. larvae ERIC II (DSM25430) cultivated in static liquid. Bacterial suspensions of P. larvae ERIC I (ATCC9545; A,D,G) and P. larvae ERIC II (DSM25430; B,E,H) in brain heart infusion (BHI) broth were incubated without agitation in six-well-plates at 37 °C for five (A,B) and six (D,E) days, or in 96-well-plates for six days (G,H). For each P. larvae genotype, three biological replicates were performed both for obtaining pictures under unstained conditions (A,B), after Congo red staining of the extracellular matrix (D,E), and Crystal violet staining of the bacterial cells adherent to the well walls (G,H). Negative controls for each assay are shown in (C,F, and I). Representative pictures are shown.
A characteristic hallmark of biofilms is that the bacteria are surrounded by an extracellular matrix which contains i.a. exopolysaccharides and amyloid fibers36,37. Hence, dyes specifically staining these extracellular matrix components can be used to specifically visualize biofilms47–49. To determine whether the observed P. larvae aggregates (Fig. 3A,B) indeed represented biofilms, that is that the bacteria were surrounded by an extracellular matrix, we used the amyloidophilic dyes Congo red and thioflavin S. Both dyes stain polysaccharides and amyloid fibers50–53 present in the extracellular matrix produced by bacteria during biofilm formation and for stabilizing the biofilm48,54–58 and, therefore, are routinely used to test for bacterial biofilm formation.
Upon Congo red staining, the multicellular aggregates (Fig. 3A,B) stained red while the surrounding medium remained unstained (Fig. 3D,E) like the negative control which represented CR-stained medium (Fig. 3F). These results indicated that both P. larvae ERIC I and ERIC II aggregates were embedded in extracellular matrix and, hence, that the bacteria formed biofilms under the chosen conditions. The biofilms though floating again appeared to have contact to the walls of the wells. Consequently, bacterial cells adherent to the well walls could be visualized by Crystal violet staining (Fig. 3G,H). No staining was evident in the negative controls (Fig. 3I).
In order to further substantiate that CR specifically stained the extracellular matrix surrounding P. larvae cells organized in a biofilm, we next tried to stain planktonic P. larvae cells, which were obtained by culturing P. larvae under constant agitation thereby preventing biofilm formation. Incubating such a non-static culture of P. larvae, hence, of planktonic P. larvae cells, with CR (Fig. 4A,B) did not reveal any stained aggregates (for comparison see Fig. 3D,E) but instead the wells looked like the negative controls (Figs 3F, 4C,D). These results suggested that the aggregates which formed when P. larvae was cultivated in static liquid and which were visualized by using the biofilm specific dye CR (Fig. 3A–F) were indeed P. larvae biofilms.
Figure 4.

Planktonic cells of P. larvae ERIC I (ATCC9545) and P. larvae ERIC II (DSM25430). Bacterial suspensions of P. larvae ERIC I (ATCC9545; A) and P. larvae ERIC II (DSM25430; B) in brain heart infusion (BHI) broth were incubated under constant agitation to prevent biofilm formation. Cultures of planktonic cells were stained with Congo red (A,B) while BHI medium stained with Congo red (C) or without staining (D) served as negative controls. Representative pictures are shown.
In order to provide further evidence for P. larvae biofilm formation, we next examined the three-dimensional structures of the bacterial aggregates by thioflavin S staining and semi-confocal fluorescence microscopy allowing to generate Z-stacks by incrementally stepping through the samples. Green fluorescence indicative for the presence of amyloid fibers was observed in a defined structured zone of the well as floating mirror-image ellipsoidal clouds each having a more densely fluorescing “axis of reflection” (Fig. 5A,B) which seemed to be more pronounced in DSM25430 than in ATCC9545 (Fig. 5C,D). The P. larvae biofilms neither adhered at the bottom of the well nor formed at the liquid-air interface (pellicle) but were rather levitating in the well (Fig. 5) like free floating biofilm aggregates recently described for Pseudomonas aeruginosa40.
Figure 5.

Fluorescence microscopy of P. larvae floating biofilms. Bacterial suspensions of P. larvae ERIC I (ATCC9545; A,C) and P. larvae ERIC II (DSM25430; B,D) in Sf-900 II SFM medium supplemented with 30 µg/ml thioflavin S were incubated without agitation in 96-well-plates at 37 °C for six days. The thioflavin S-stained extracellular matrix in the floating biofilms was visualized using fluorescence microscopy; Z-stack processing was performed to obtain three-dimensional images of the wells containing the biofilms (A,B) and the region within the wells where the biofilms were located (C,D). Bars represent 20 µm.
Influence of paenilarvin on swarming motility and biofilm formation
Paenilarvins are iturin-like secondary metabolite specifically produced by P. larvae ERIC II28. Because iturin-like secondary metabolites are known to influence the motility of their bacterial producers59,60, we tested whether paenilarvin has an influence on swarming motility or biofilm formation of P. larvae ERIC II. We used the newly established assays for swarming motility and biofilm formation and compared the behaviour of wild-type P. larvae DSM25430 and of an inactivation mutant for the paenilarvin gene cluster, P. larvae DSM25430 Δitu, which is deficient in paenilarvin production29.
We first analyzed the involvement of paenilarvin in swarming motility of P. larvae ERIC II. One day post inoculation, neither the wild-type nor the mutant bacteria had yet started to swarm (Fig. 6). However, after two days, the wild-type bacteria showed the expected swarming behaviour which resulted already after three days in a rather featureless mat covering the entire plate and with the original macrocolony still visible. At day six post inoculation, the inoculation site in the centre of the plate was no longer visible as area of higher cell density and instead a pattern resembling zones of consolidation or terraces (Bull’s eye30;) had developed (Fig. 6A, upper row). In contrast, P. larvae DSM25430 Δitu showed delayed swarming motility: Only after three days, some swarming started at the edge of the central macrocolony and the entire plate was covered not until after six days; the central macrocolony remained clearly visible even after seven days (Fig. 6A, lower row). Measuring the swarming radii at different time points allowed a quantitative analysis of the data and confirmed statistically significant differences (day 2: p value = 0.002; day 3: p < 0.0001) in swarming motility between the wild-type and mutant bacteria (Fig. 6B). These results indicated that in the absence of the iturin-like paenilarvin swarming was significantly delayed suggesting a role of this secondary metabolite in swarming motility of P. larvae ERIC II. Further experiments are necessary to examine the role of paenilarvins in swarming motility in more detail.
Figure 6.
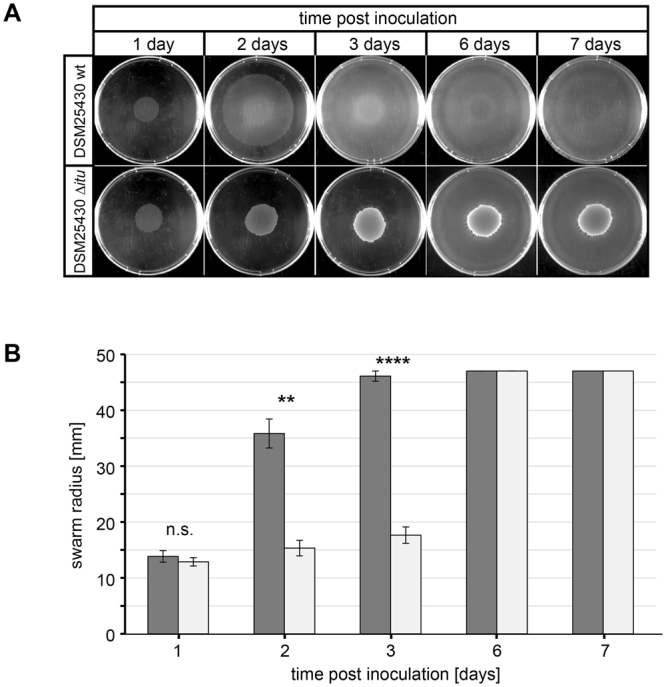
Involvement of paenilarvin in swarming motility of P. larvae ERIC II. Wild-type P. larvae ERIC II (DSM25430 wt) and a corresponding inactivation mutant for the paenilarvin gene cluster (DSM25430 Δitu) were tested in swarming assays. Bacterial suspensions (10 µl) with an OD600 of 0.1 were inoculated centrally onto a brain heart infusion (BHI; 0.5%) agar plate and incubated at 37 °C for seven days. Pictures were taken at one, two, three, six, and seven days post inoculation. Three biological replicates with three technical replicates each were performed. (A) Representative pictures for each day and strain (DSM25430 wt, upper row; DSM25430 Δitu, lower row) are shown. Uncolonized agar is black and bacterial biomass is milky white to white. (B) Swarm radii were measured for each strain and time point. Data represent mean values ± SEM of three independent experiments each with three technical replicates. Statistical analysis of the difference in swarming motility between DSM25430 wt (dark gray columns) and DSM25430 Δitu (off-white columns) was performed with an unpaired Student’s t-test for each time point (not significantly different: n.s., p ≥ 0.05; significantly different: *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001).
When we tested the involvement of paenilarvin in biofilm formation of P. larvae ERIC II, we did not observe any difference between wild-type P. larvae DSM25430 and the paenilarvin gene cluster inactivation mutant P. larvae DSM25430 Δitu (Fig. 7A–D). The biofilms formed by DSM25430 and DSM25430 Δitu contained 8.21 ± 2.15 µg bound CR and 8.22 ± 0.16 µg bound CR, respectively (Fig. 7E). The difference was not significant (p = 0.9889, Student’s t-test). Therefore, biofilm formation was unaffected by the absence of paenilarvin production indicating that the iturin-like lipopeptide has no role in this process.
Figure 7.
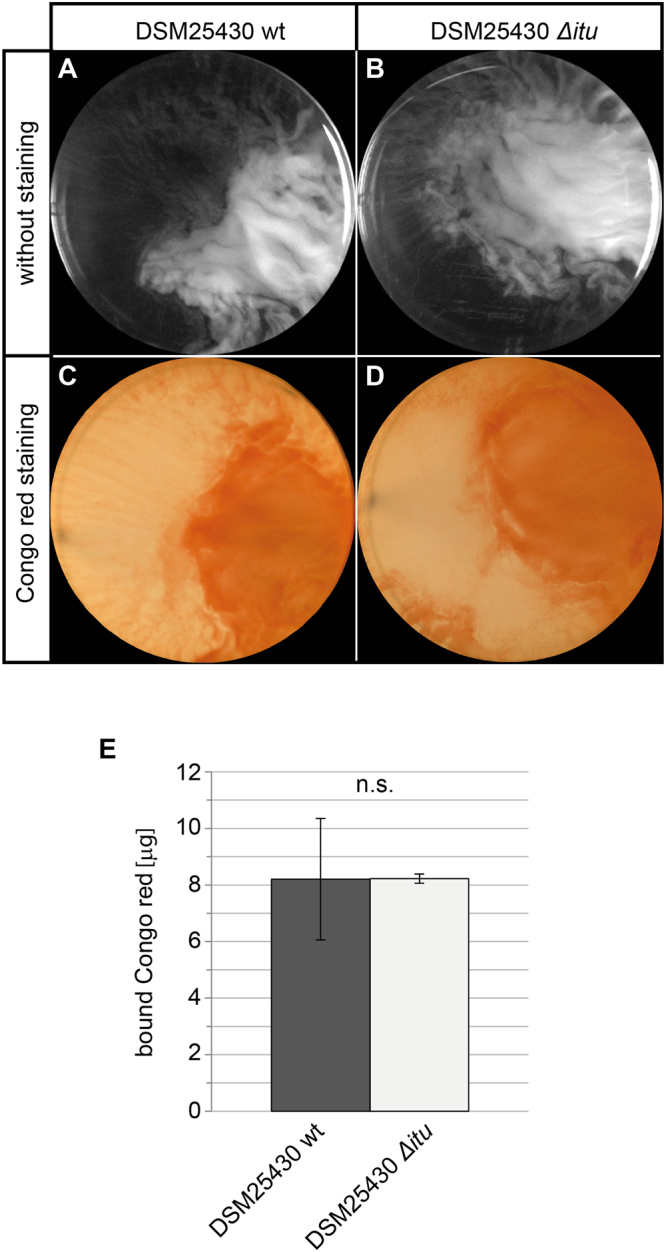
Involvement of paenilarvin in biofilm formation of P. larvae ERIC II. Wild-type P. larvae ERIC II (DSM25430 wt; A,C) and a corresponding inactivation mutant for the paenilarvin gene cluster (DSM25430 Δitu; B,D) were tested in biofilm assays. Bacterial suspensions in brain heart infusion (BHI) broth were incubated without agitation in six-well-plates at 37 °C for five (A,B) and six (C,D) days. For both strains, three biological replicates were performed both for obtaining pictures under unstained conditions (A,B) and after Congo red staining (C,D). Representative pictures are shown. (E) Biofilm formation was quantified via determining the amount of Conge red dye retained in the biofilms upon centrifugation. Data represent mean values ± SD of three independent experiments. The difference between the wild-type and corresponding inactivation mutant was not significant (p = 0.9889, Student’s t-test).
Discussion
The honey bee pathogen P. larvae is the etiological agent of AFB, the most destructive bacterial disease of honey bees4. P. larvae is considered an obligate killer because death of the infected larva and subsequent degradation of the larval cadaver to a slimy mass that dries out to the so-called foulbrood scale consisting of P. larvae spores only are both necessary prerequisites for efficient spore production and transmission (for recent reviews see61,62). Hence, the pathogenic phase (until the death of the larva) and the saprophytic phase (degradation of the larval cadaver) are equally important phases in the P. larvae lifecycle wherein the death of the infected larva does not represent the end of the vegetative growth of P. larvae but rather marks the transition from the pathogenic to the saprophytic phase. In the recent past, the availability of the genome sequences of the P. larvae genotypes ERIC I and ERIC II13 and of protocols for the genetic manipulation of this pathogen19,63 has led to considerable progress in elucidating P. larvae virulence factors. These results improved our understanding of the bacterial strategies employed to kill the larvae, hence, strategies mainly for the pathogenic phase of the lifecycle of P. larvae62. In contrast, the saprophytic phase still remained elusive due to the lack of appropriate functional assays. We are convinced that for degrading the larval cadaver, P. larvae has to be capable of some kind of coordinated or cooperative multicellular behaviour like swarming motility and/or biofilm formation. Hence, the here presented swarming and biofilm assays for P. larvae will allow identifying factors relevant for the success of this bee pathogen after the infected larva has been killed. This will further our understanding of the entire lifecycle of P. larvae, which in turn is a prerequisite for the future development of sustainable treatment strategies.
Swarming motility is a coordinated bacterial activity and widespread amongst flagellated bacteria64. Several species of the genus Paenibacillus have been shown to be capable of swarming32–34. P. larvae harbors peritrichously arranged flagella, needs to rapidly colonize the larval cadaver which can be considered a nutrient-rich environment, and needs to convert larval to bacterial biomass quite fast. Hence, capability of swarming suggested itself for P. larvae. However, so far we could not demonstrate a general ability to swarm for the species P. larvae. Instead, only the P. larvae genotype ERIC II exhibited swarming motility in our assays. One possible reason for this difference might be the differential expression of paenilarvin, an iturin-like lipopeptide, most probably acting as biosurfactant. It has been described that in addition to flagellae and an extracellular matrix the production of biosurfactants is often required for swarming motility to occur35. Biosurfactants are surface active agents (hence the name) produced by microorganisms. They are amphiphilic and thus lower the surface tension or interfacial tension between either two liquids or a liquid and a solid. On agar plates, they might increase the wettability thereby facilitating spreading behaviour and swarming motility. The role of lipopeptide biosurfactants for the motility of their bacterial producers on agar plates is well characterized and it has been shown for many swarming bacteria that surface motility is lost or reduced in mutants deficient in producing lipopeptides59,60. In accordance with these results, the P. larvae ERIC II Δitu mutant lacking the production of paenilarvins29 exhibited a significantly delayed swarming behaviour. Because this mutant does not differ in growth rate from its corresponding wild type strain29, growth defects cannot explain the delayed swarming phenotype. Our results rather indicate that the paenilarvins are involved in facilitating swarming motility of P. larvae ERIC II. Hence, we propose that the biological roles of paenilarvins comprise not only their anti-fungal activity28 but also their activity as biosurfactant.
It has already been demonstrated that P. larvae ERIC II - but not ERIC I - produces the lipopeptide paenilarvin13,23,28. We here showed that P. larvae ERIC II swarming was hampered in the absence of paenilarvin production. Hence, the inability of P. larvae ERIC I to swarm might be related to the lack of paenilarvin production in this genotype. However, since surface motility was only delayed but not totally lost in the paenilarvin deficient P. larvae ERIC II Δitu mutant, paenilarvin obviously alleviated swarming but was not essential for swarming. Therefore, the lack of paenilarvin production cannot be the only explanation for the lack of swarming of P. larvae ERIC I. Further research is needed to elucidate the regulatory networks that control swarming in P. larvae and to understand the role of swarming during AFB pathogenesis and degradation of the larval cadaver.
Biofilm formation is another coordinated activity of a cooperatively acting group of bacteria. Normally, biofilms are considered sessile communities composed of one or several bacterial species embedded within an extracellular matrix and attached to a solid surface or formed as pellicles at liquid-air interfaces35. Recently, the additional existence of free floating biofilm aggregates has been demonstrated for Pseudomonas aeruginosa40. The ability to form biofilms had not been proposed for P. larvae so far, although P. larvae-specific FISH analysis of diseased larvae had already revealed the formation of bacterial clusters or aggregates akin to floating biofilms in the larval midgut lumen during early stages of infection16. Bacterially produced extracellular matrix is a hallmark and prerequisite of biofilms and, indeed, genes encoding exopolysaccharide biosynthesis proteins have been annotated in the genome of P. larvae (ERIC I: GenBank ETK30098.1; ERIC II: GenBank AHD06625.1)13. Furthermore, the slimy consistence of the bacterial mass which remains after total degradation of larval cadaver points to biofilm formation by P. larvae at least at the end of its lifecycle. Using biofilm specific staining methods we here demonstrated that both genotypes of P. larvae were able to form biofilms when incubated in broth culture without agitation, hence, in static liquid. The biofilms did neither adhere to the bottom of the wells nor did they form pellicles at the air liquid interface but instead they appeared as free floating biofilms as already described for Pseudomonas aeruginosa40. These free floating biofilms seemed to be loosely attached to the walls of the wells. For P. larvae degrading the larval cadaver, this cadaver does neither provide a proper “surface” to adhere to nor a liquid-air interface for the formation of a pellicle. Therefore, a free floating biofilm at the beginning of the saprophytic phase is a likely cooperative activity to ensure optimal colonization of the cadaver and access to all nutrients.
Although lipopeptides have been shown to play an important role in surface attachment and biofilm formation in Pseudomonas and Bacillus59, we could not demonstrate a role for paenilarvin in biofilm formation of P. larvae. Future research is needed to analyze P. larvae biofilm formation in detail. We need to unravel the steps and the regulatory mechanisms in biofilm formation, to identify the components of the extracellular matrix, and to understand the role of biofilm formation for the success of P. larvae as bee pathogen.
Experimental Procedures
Bacterial strains and culture conditions
P. larvae wildtype strains ATCC9545 and DSM25430 representing genotypes ERIC I and ERIC II were used in this study (Genersch et al.4). Both strains have been extensively characterized before; both are virulent and harbor all tested phenotypic features characteristic for the respective genotype4,9–11,14,15. In addition, the recently published mutant strain DSM25430 Δitu was used which carries a paenilarvin gene cluster inactivation29 generated via the TargeTron Gene Knockout System (Sigma-Aldrich,Germany) following a protocol originally established for P. alvei65 and modified for use in P. larvae19. DSM25430 wt and DSM25430 Δitu do neither differ in their growth characteristics nor in virulence for infected larvae29. Furthermore, five field strains of P. larvae ERIC I (00-087, 01-440, 02-130, 03-159, 03-189) and five of P. larvae ERIC II (00-897, 00-1163, 03-194, 03-522, 03-525), all of them isolated from contemporary AFB outbreaks, were employed in this study. These strains have already been extensively characterized and used in other studies4,9,10,66. Wildtype strains as well as mutant strains were cultivated on Columbia sheep blood agar plates (CSA; Oxoid, Hampshire, UK) at 37 °C for 3–5 days or according to the conditions given for each assay (see below).
Swarming assay
Swarming assays were performed with P. larvae ERIC I strains (ATCC9545, 00-087, 01-440, 02-130, 03-159, 03-189) and ERIC II strains (DSM25430, DSM25430 Δitu, 00-897, 00-1163, 03-194, 03-522, 03-525). To obtain pre-cultures, 2.0 ml of BHI (brain heart infusion; Merck KGaA, Darmstadt, Germany) medium were inoculated with one single colony of each P. larvae strain and incubated overnight at 35 °C with agitation. The next day, BHI broth (Merck, Darmstadt, Germany) agar plates (0.5%) were poured from relatively cool (~50 °C30;) agar. The plates were dried with the lid open at room temperature for 15 min. The pre-cultures were diluted to an OD600 of 0.1, and 10 µl of diluted culture were carefully pipetted into the center of each plate. The plates were then dried for another 15 min and subsequently incubated at 37 °C for several days. Pictures of the plates were taken after one, two, three, six, and seven days. The pictures were analyzed via ImageJ 1.44p to measure the radii of the areas covered by swarming P. larvae. Three biological replicates with three technical replicates each were performed. Data represent mean values ± SEM and statistical analysis was performed by the unpaired Student’s t-test with GraphPad Prism 6.
Biofilm assay
Biofilm assays were performed with the P. larvae ERIC I strain ATCC9545 and the wildtype ERIC II strain DSM25430 and the corresponding mutant DSM25430 Δitu. Pre-cultures of P. larvae were prepared as described above and incubated at 35 °C with agitation until the stationary phase was reached (after about 42 h). The pre-cultures in the stationary phase were diluted to an OD600 of 0.1 and 6.0 ml of diluted culture were transferred to the wells of a Cellstar 6-well tissue culture plate (Greiner Bio-One GmbH, Frickenhausen, Germany). Wells for negative controls contained medium only. The plates were incubated at 37 °C for five days without agitation (static culture). After five days, pictures of the unstained wells containing either cultivated (static culture) P. larvae or medium only (negative controls) were taken. By then, the cultures of ATCC9545 and DSM25430 had reached an OD600 of about 0.4 and 0.5, respectively. For better visualizing the biofilm, Congo red (CR; Merck KGaA, Darmstadt, Germany), a direct dye for staining amyloid fibres and extrapolysaccharides in biofilms47–51 was used. CR was added to all wells to reach a final concentration of 30 µg/ml and the plates were incubated for another 24 h at 37 °C before pictures of all wells (static culture of P. larvae or medium only) were taken.
To compare biofilm formation of P. larvae DSM25430 wildtype and the mutant strain DSM25430 Δitu, binding of CR and, hence, the production of extracellular matrix was quantified. To this end, CR bound by the P. larvae biofilm was removed by centrifugation (5 min, 13,000 g) and unbound CR was determined by measuring the absorbance of the supernatant at 490 nm67,68. BHI medium which was incubated with CR in the same way was used as reference value for calculating the amount of bound CR67,68. The amounts of bound CR can be compared directly since the growth of the mutant does not differ from the wild-type29. Data represent mean values ± SD of three biological replicates and were analyzed by the unpaired Student’s t-test (GraphPad Prism 6).
To further substantiate that CR specifically stains the extracellular matrix and does not stain planktonic bacterial cells, the dilute pre-cultures of P. larvae (OD600 = 0.1) were incubated in the presence of CR (30 µg/ml) for 24 hours at 37 °C with constant agitation to prevent biofilm formation. Under these non-static conditions, the cultures of ATCC9545 and DSM25430 reached an OD600 of about 1.0 and 1.2, respectively. Subsequently, a volume of 6 ml of planktonic cells was transferred to the wells of a 6-well culture plate and pictures were taken.
To demonstrate P. larvae cells adherent to the walls of the culture wells, Crystal violet staining was performed by slightly modifying established protocols69. P. larvae cultures were grown overnight in BHI broth and diluted to an OD600 of 0.1. A volume of each 150 µl of the diluted bacterial culture was pipetted into the wells of a 96-well cell culture plate (Greiner Bio-One GmbH, Frickenhausen, Germany). Negative controls were left uninoculated. Cultures were incubated at 37 °C for six days. The plates were washed three times with MilliQ-water, stained with 0.1% Crystal violet (Merck KGaA, Darmstadt, Germany) and again washed three times before pictures were taken.
For visualizing the three-dimensional structure of the biofilms via fluorescence microscopy, thioflavin S (Sigma-Aldrich Chemie GmbH, Munich, Germany), a fluorescent dye for staining amyloid fibres and extrapolysaccharides in biofilms, was used. Pre-cultures were obtained by inoculating Sf-900 II SFM medium (Thermo Fisher Scientific, Darmstadt, Germany) with one single colony of each P. larvae strain followed by incubation at 37 °C with agitation overnight. The pre-cultures were diluted with SF-900 II SFM medium to an OD600 of 0.01 and transferred to a 96-well tissue culture plate (Greiner Bio-One GmbH, Frickenhausen, Germany). After addition of 30 µg/ml thioflavin S (Sigma-Aldrich Chemie GmbH, Munich, Germany), the plates were sealed with parafilm and incubated at 37 °C for 6 days. Thioflavin S fluorescence organized in a clearly defined 3D-structure was visualized using fluorescence inverted microscopy (Eclipse Ti, Nikon GmbH, Düsseldorf, Germany). Z-stack processing was performed to obtain the three-dimensional images. Biofilms grown in the absence of thioflavin S were included in the experiments as controls in order to determine the background fluorescence.
Data availability
All data generated or analyzed during this study are included in this published article.
Acknowledgements
This work was supported by the Ministries responsible for Agriculture of the German Federal States of Brandenburg (MIL) and Sachsen-Anhalt (MLU), Germany, and by the German Research Foundation (grant # GE1365/1-2 and GE1365/2-1) to EG.
Author Contributions
A.F., E.G. conceived and designed the experiments A.F., J.G., J.E., H.K., E.G.G. performed the experiments J.G., A.F., E.G. analyzed the data E.G., A.F., J.G. wrote the paper; all authors reviewed the manuscript.
Competing Interests
The authors declare no competing interests.
Footnotes
Anne Fünfhaus and Josefine Göbel contributed equally to this work.
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Aizen M, Garibaldi L, Cunningham S, Klein A. Long-term global trends in crop yield and production reveal no current pollination shortage but increasing pollinator dependency. Curr. Biol. 2008;18:1572–1575. doi: 10.1016/j.cub.2008.08.066. [DOI] [PubMed] [Google Scholar]
- 2.Aizen MA, Harder LD. The global stock of domesticated honey bees is growing slower than agricultural demand for pollination. Curr. Biol. 2009;19:915–918. doi: 10.1016/j.cub.2009.03.071. [DOI] [PubMed] [Google Scholar]
- 3.Genersch E. Honey bee pathology: Current threats to honey bees and beekeeping. Appl. Microbiol. Biotechnol. 2010;87:87–97. doi: 10.1007/s00253-010-2573-8. [DOI] [PubMed] [Google Scholar]
- 4.Genersch E, et al. Reclassification of Paenibacillus larvae subsp. pulvifaciens and Paenibacillus larvae subsp. larvae as Paenibacillus larvae without subspecies differentiation. Int. J. Syst. Evol. Microbiol. 2006;56:501–511. doi: 10.1099/ijs.0.63928-0. [DOI] [PubMed] [Google Scholar]
- 5.Genersch E. American Foulbrood in honeybees and its causative agent. Paenibacillus larvae. J. Invertebr. Pathol. 2010;103:S10–S19. doi: 10.1016/j.jip.2009.06.015. [DOI] [PubMed] [Google Scholar]
- 6.Schäfer MO, et al. Rapid identification of differentially virulent genotypes of Paenibacillus larvae, the causative organism of American foulbrood of honey bees, by whole cell MALDI-TOF mass spectrometry. Vet. Microbiol. 2014;170:291–297. doi: 10.1016/j.vetmic.2014.02.006. [DOI] [PubMed] [Google Scholar]
- 7.Morrissey BJ, et al. Biogeography of Paenibacillus larvae, the causative agent of American foulbrood, using a new multilocus sequence typing scheme. Environ. Microbiol. 2015;17:1414–1424. doi: 10.1111/1462-2920.12625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Descamps T, De Smet L, Stragier P, De Vos P, de Graaf DC. Multiple locus variable number of tandem repeat analysis: A molecular genotyping tool for Paenibacillus larvae. Microb. Biotechnol. 2016;9:772–781. doi: 10.1111/1751-7915.12375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Neuendorf S, Hedtke K, Tangen G, Genersch E. Biochemical characterization of different genotypes of Paenibacillus larvae subsp. larvae, a honey bee bacterial pathogen. Microbiology. 2004;150:2381–2390. doi: 10.1099/mic.0.27125-0. [DOI] [PubMed] [Google Scholar]
- 10.Genersch E, Ashiralieva A, Fries I. Strain- and genotype-specific differences in virulence of Paenibacillus larvae subsp. larvae, the causative agent of American foulbrood disease in honey bees. Appl. Environ. Microbiol. 2005;71:7551–7555. doi: 10.1128/AEM.71.11.7551-7555.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Rauch S, Ashiralieva A, Hedtke K, Genersch E. Negative correlation between individual-insect-level virulence and colony-level virulence of Paenibacillus larvae, the etiological agent of American foulbrood of honeybees. Appl. Environ. Microbiol. 2009;75:3344–3347. doi: 10.1128/AEM.02839-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chan QWT, et al. Updated genome assembly and annotation of Paenibacillus larvae, the agent of American Foulbrood disease of honey bees. BMC Genomics. 2011;12:450. doi: 10.1186/1471-2164-12-450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Djukic M, et al. How to kill the honey bee larva: Genomic potential and virulence mechanisms of Paenibacillus larvae. Plos One. 2014;9:e90914. doi: 10.1371/journal.pone.0090914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Fünfhaus A, Ashiralieva A, Borriss R, Genersch E. Use of suppression subtractive hybridization to identify genetic differences between differentially virulent genotypes of Paenibacillus larvae, the etiological agent of American Foulbrood of honeybees. Environ. Microbiol. Rep. 2009;1:240–250. doi: 10.1111/j.1758-2229.2009.00039.x. [DOI] [PubMed] [Google Scholar]
- 15.Fünfhaus A, Genersch E. Proteome analysis of Paenibacillus larvae reveals the existence of a putative S-layer protein. Environ. Microbiol. Rep. 2012;4:194–202. doi: 10.1111/j.1758-2229.2011.00320.x. [DOI] [PubMed] [Google Scholar]
- 16.Yue D, Nordhoff M, Wieler LH, Genersch E. Fluorescence in situ-hybridization (FISH) analysis of the interactions between honeybee larvae and Paenibacillus larvae, the causative agent of American foulbrood of honeybees (Apis mellifera) Environ. Microbiol. 2008;10:1612–1620. doi: 10.1111/j.1462-2920.2008.01579.x. [DOI] [PubMed] [Google Scholar]
- 17.Garcia-Gonzalez E, Genersch E. Honey bee larval peritrophic matrix degradation during infection with Paenibacillus larvae, the aetiological agent of American foulbrood of honey bees, is a key step in pathogenesis. Environ. Microbiol. 2013;15:2894–2901. doi: 10.1111/1462-2920.12167. [DOI] [PubMed] [Google Scholar]
- 18.Garcia-Gonzalez E, et al. Paenibacillus larvae chitin-degrading protein PlCBP49 is a key virulence factor in American Foulbrood of honey bees. Plos Path. 2014;10:e1004284. doi: 10.1371/journal.ppat.1004284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Poppinga L, et al. Identification and functional analysis of the S-layer protein SplA of Paenibacillus larvae, the causative agent of American Foulbrood of honey bees. Plos Path. 2012;8:e1002716. doi: 10.1371/journal.ppat.1002716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ebeling J, et al. Characterization of the toxin Plx2A, a RhoA-targeting ADP-ribosyltransferase produced by the honey bee pathogen Paenibacillus larvae. Environ. Microbiol. 2017;19:5100–5116. doi: 10.1111/1462-2920.13989. [DOI] [PubMed] [Google Scholar]
- 21.Fünfhaus A, Poppinga L, Genersch E. Identification and characterization of two novel toxins expressed by the lethal honey bee pathogen Paenibacillus larvae, the causative agent of American foulbrood. Environ. Microbiol. 2013;15:2951–2965. doi: 10.1111/1462-2920.12229. [DOI] [PubMed] [Google Scholar]
- 22.Krska D, Ravulapalli R, Fieldhouse RJ, Lugo MR, Merrill AR. C3larvin toxin, an ADP-ribosyltransferase from Paenibacillus larvae. J. Biol. Chem. 2015;290:1639–1653. doi: 10.1074/jbc.M114.589846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Müller S, Garcia-Gonzalez E, Genersch E, Süssmuth R. Involvement of secondary metabolites in the pathogenesis of the American foulbrood of honey bees caused by Paenibacillus larvae. Nat. Prod. Rep. 2015;32:765–778. doi: 10.1039/C4NP00158C. [DOI] [PubMed] [Google Scholar]
- 24.Garcia-Gonzalez E, Müller S, Ensle P, Süssmuth RD, Genersch E. Elucidation of sevadicin, a novel nonribosomal peptide secondary metabolite produced by the honey bee pathogenic bacterium Paenibacillus larvae. Environ. Microbiol. 2014;16:1297–1309. doi: 10.1111/1462-2920.12417. [DOI] [PubMed] [Google Scholar]
- 25.Garcia-Gonzalez E, et al. Biological effects of paenilamicin, a secondary metabolite antibiotic produced by the honey bee pathogenic bacterium Paenibacillus larvae. Microbiology Open. 2014;3:642–656. doi: 10.1002/mbo3.195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Müller S, et al. Paenilamicin - structure and biosynthesis of a hybrid non-ribosomal peptide/ polyketide antibiotic from the bee pathogen Paenibacillus larvae. Angew. Chem. Int. Ed. Engl. 2014;53:10547–10828. doi: 10.1002/anie.201406846. [DOI] [PubMed] [Google Scholar]
- 27.Hertlein G, et al. Production of the catechol type siderophore bacillibactin by the honey bee pathogen Paenibacillus larvae. Plos One. 2014;9:e108272. doi: 10.1371/journal.pone.0108272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sood S, et al. Paenilarvins: Iturin family lipopeptides from the honey bee pathogen Paenibacillus larvae. Chem. Bio Chem. 2014;15:1947–1955. doi: 10.1002/cbic.201402139. [DOI] [PubMed] [Google Scholar]
- 29.Hertlein G, et al. Biological role of paenilarvins, iturin-like lipopeptide secondary metabolites produced by the honey bee pathogen Paenibacillus larvae. Plos One. 2016;11:e0164656. doi: 10.1371/journal.pone.0164656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kearns DB. A field guide to bacterial swarming motility. Nat. Rev. Microbiol. 2010;8:634–644. doi: 10.1038/nrmicro2405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Fraser GM, Hughes C. Swarming motility. Curr. Opin. Microbiol. 1999;2:630–635. doi: 10.1016/S1369-5274(99)00033-8. [DOI] [PubMed] [Google Scholar]
- 32.Thampuran N, Surendran PK. Effect of chemical reagents on swarming of Bacillus species. J. Appl. Bacteriol. 1996;80:296–302. doi: 10.1111/j.1365-2672.1996.tb03223.x. [DOI] [Google Scholar]
- 33.Be’er A, et al. Paenibacillus dendritiformis bacterial colony growth depends on surfactant but not on bacterial motion. J. Bacteriol. 2009;191:5758–5764. doi: 10.1128/JB.00660-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ingham CJ, Ben-Jacob E. Swarming and complex pattern formation in Paenibacillus vortex studied by imaging and tracking cells. BMC Microbiol. 2008;8:36. doi: 10.1186/1471-2180-8-36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Verstraeten N, et al. Living on a surface: swarming and biofilm formation. Trends Microbiol. 2008;16:496–506. doi: 10.1016/j.tim.2008.07.004. [DOI] [PubMed] [Google Scholar]
- 36.Mielich-Süss B, Lopez D. Molecular mechanisms involved in Bacillus subtilis biofilm formation. Environ. Microbiol. 2015;17:555–565. doi: 10.1111/1462-2920.12527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Davey ME, O’Toole GA. Microbial biofilms: from ecology to molecular genetics. Microbiol. Mol. Biol. Rev. 2000;64:847–867. doi: 10.1128/MMBR.64.4.847-867.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Nagar E, Schwarz R. To be or not to be planktonic? Self-inhibition of biofilm development. Environ. Entomol. 2015;17:1477–1486. doi: 10.1111/1462-2920.12583. [DOI] [PubMed] [Google Scholar]
- 39.Monds RD, O’Toole GAO. The developmental model of microbial biofilms: ten years of a paradigm up for review. Trends Microbiol. 2008;17:73–87. doi: 10.1016/j.tim.2008.11.001. [DOI] [PubMed] [Google Scholar]
- 40.Kragh KN, et al. Role of multicellular aggregates in biofilm formation. mBio. 2016;7:e00237–00216. doi: 10.1128/mBio.00237-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Algburi A, Comito N, Kashtanov D, Dicks LMT, Chikindas ML. Control of biofilm formation: antibiotics and beyond. Appl. Environ. Microbiol. 2017;83:e02508–02516. doi: 10.1128/AEM.02508-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Stewart PS, Costerton JW. Anitbiotic resistance of bacteria in biofilms. Lancet. 2001;358:135–138. doi: 10.1016/S0140-6736(01)05321-1. [DOI] [PubMed] [Google Scholar]
- 43.Høiby N, Bjarnsholt T, Givskov M, Molin S, Ciofu O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents. 2010;35:322–332. doi: 10.1016/j.ijantimicag.2009.12.011. [DOI] [PubMed] [Google Scholar]
- 44.Beloin C, Renard S, Ghigo J-M, Lebeaux D. Novel approaches to combat bacterial biofilms. Curr. Opin. Pharmacol. 2014;18:61–68. doi: 10.1016/j.coph.2014.09.005. [DOI] [PubMed] [Google Scholar]
- 45.Stoodley P, Sauer K, Davies DG, Costerton JW. Biofilms as complex differentiated communities. Annu. Rev. Microbiol. 2002;56:187–209. doi: 10.1146/annurev.micro.56.012302.160705. [DOI] [PubMed] [Google Scholar]
- 46.Descamps T, De Smet L, De Vos P, de Graaf DC. Unbiased random mutagenesis contributes to a better understanding of the virulent behaviour of Paenibacillus larvae. J. Appl. Microbiol. 2017;124:28–41. doi: 10.1111/jam.13611. [DOI] [PubMed] [Google Scholar]
- 47.Patel CN, et al. Polyamines are essential for the formation of plague biofilm. J. Bacteriol. 2006;188:2355–2363. doi: 10.1128/JB.188.7.2355-2363.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Chimileski S, Franklin MJ, Papke RT. Biofilms formed by the archaeon Haloferax volcanii exhibit cellular differentiation and social motility, and facilitate horizontal gene transfer. BMC Biology. 2014;12:65. doi: 10.1186/s12915-014-0065-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Harrison-Balestra C, Cazzaniga AL, Davis SC, Mertz PM. A wound-isolated Pseudomonas aeruginosa grows a biofilm in vitro within 10 hours and is visualized by light microscopy. Dermatol. Surg. 2003;29:631–635. doi: 10.1046/j.1524-4725.2003.29146.x. [DOI] [PubMed] [Google Scholar]
- 50.Wood PJ. Specificity in the interaction of direct dyes with polysaccharides. Carbohydrate Res. 1980;85:271–287. doi: 10.1016/S0008-6215(00)84676-5. [DOI] [Google Scholar]
- 51.Khurana R, Uversky VN, Nielsen L, Fink AL. Is Congo red an amyloid-specific dye? J. Biol. Chem. 2001;276:22715–22721. doi: 10.1074/jbc.M011499200. [DOI] [PubMed] [Google Scholar]
- 52.Bennhold H. Eine spezifische Amyloidfärbung mit Kongorot. Münch. Med. Wschr. 1922;69:1537–1538. [Google Scholar]
- 53.Nilsson MR. Techniques to study amyloid fibril formation in vitro. Methods. 2004;34:151–160. doi: 10.1016/j.ymeth.2004.03.012. [DOI] [PubMed] [Google Scholar]
- 54.Larsen P, Nielsen J, Otzen D, Nielsen PH. Amyloid-like adhesins produced by floc-forming and filamentous bacteria in activated sludge. Appl. Environ. Microbiol. 2008;74:1517–1526. doi: 10.1128/AEM.02274-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Chapman MR, et al. Role of Escherichia coli curli operons in directing amyloid fiber formation. Science. 2002;295:851–855. doi: 10.1126/science.1067484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Dutta A, Bhattacharyya S, Kundu A, Dutta D, Das AK. Macroscopic amyloid fiber formation by staphylococcal biofilm associated SuhB protein. Biophys. Chem. 2016;217:32–41. doi: 10.1016/j.bpc.2016.07.006. [DOI] [PubMed] [Google Scholar]
- 57.Oli WH, et al. Functional amyloid formation by Streptococcus mutans. Microbiology. 2012;158:2903–2916. doi: 10.1099/mic.0.060855-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Serra DO, Richter AM, Klauck G, Mika F, Hengge R. Microanatomy at cellular resolution and spatial order of physiological differentiation in a bacterial biofilm. mBio. 2013;4:e00103–00113. doi: 10.1128/mBio.00103-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Raaijmakers JM, de Bruijn I, Nybroe O, Ongena M. Natural functions of lipopeptides from Bacillus and Pseudomonas: more than surfactants and antibiotics. FEMS Microbiol. Rev. 2010;34:1037–1062. doi: 10.1111/j.1574-6976.2010.00221.x. [DOI] [PubMed] [Google Scholar]
- 60.Leclère V, Marti R, Béchet M, Fickers P, Jacques P. The lipopeptides mycosubtilin and surfactin enhance spreading of Bacillus subtilis strains by their surface-active properties. Arch. Microbiol. 2006;186:475–483. doi: 10.1007/s00203-006-0163-z. [DOI] [PubMed] [Google Scholar]
- 61.Ebeling J, Knispel H, Hertlein G, Fünfhaus A, Genersch E. Biology of Paenibacillus larvae, a deadly pathogen of honey bee larvae. Appl. Microbiol. Biotechnol. 2016;100:7387–7395. doi: 10.1007/s00253-016-7716-0. [DOI] [PubMed] [Google Scholar]
- 62.Poppinga L, Genersch E. Molecular pathogenesis of American Foulbrood: how Paenibacillus larvae kills honey bee larvae. Curr. Opin. Insect Sci. 2015;10:29–36. doi: 10.1016/j.cois.2015.04.013. [DOI] [PubMed] [Google Scholar]
- 63.Poppinga L, Genersch E. Heterologous expression of green fluorescent protein in Paenibacillus larvae, the causative agent of American Foulbrood of honey bees. J. Appl. Microbiol. 2012;112:430–435. doi: 10.1111/j.1365-2672.2011.05214.x. [DOI] [PubMed] [Google Scholar]
- 64.Sharma M, Anand SK. Swarming: A coordinated bacterial activity. Curr. Science. 2002;83:707–715. [Google Scholar]
- 65.Zarschler K, Janesch B, Zayni S, Schäffer C, Messner P. Construction of a gene knockout system for application in Paenibacillus alvei CCM 2051T, exemplified by the S-layer glycan biosynthesis initiation enzyme WsfP. Appl. Environ. Microbiol. 2009;75:3077–3085. doi: 10.1128/AEM.00087-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Genersch E, Otten C. The use of repetitive element PCR fingerprinting (rep-PCR) for genetic subtyping of German field isolates of Paenibacillus larvae subsp. larvae. Apidologie. 2003;34:195–206. doi: 10.1051/apido:2003025. [DOI] [Google Scholar]
- 67.Janesch B, Koerdt A, Messner P, Schäffer C. The S-layer homology domain-containing protein SlhA from Paenibacillus alvei CCM2051T is important for swarming and biofilm formation. Plos One. 2013;8:e76566. doi: 10.1371/journal.pone.0076566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Ma Q, Wood TK. OmpA influences Escherichia coli biofilm formation by repressing cellulose production through the CpxRA two-component system. Environ. Microbiol. 2009;11:2735–2746. doi: 10.1111/j.1462-2920.2009.02000.x. [DOI] [PubMed] [Google Scholar]
- 69.Merritt, J. H., Kadouri, D. E. & O’Toole, G. A. Growing and analyzing static biofilms. Curr. Protoc. Microbiol. 1B.1.1-1B.1.18 (2005). [DOI] [PMC free article] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.