

Abstract
Rett syndrome and MECP2 Duplication syndrome are neurodevelopmental disorders attributed to loss-of-function mutations in, or duplication of, the gene encoding methyl-CpG-binding protein 2 (MeCP2), respectively. We recently reported decreased expression and function of the metabotropic glutamate receptor 7 (mGlu7) in a mouse model of Rett syndrome. Positive allosteric modulation of mGlu7 activity was sufficient to improve several disease phenotypes including cognition. Here, we tested the hypothesis that mGlu7 expression would be reciprocally regulated in a mouse model of MECP2 Duplication syndrome, such that negative modulation of mGlu7 activity would exert therapeutic benefit. To the contrary, we report that mGlu7 is not functionally increased in mice overexpressing MeCP2 and that neither genetic nor pharmacological reduction of mGlu7 activity impacts phenotypes that are antiparallel to those observed in Rett syndrome model mice. These data expand our understanding of how mGlu7 expression and function is affected by changes in MeCP2 dosage and have important implications for the therapeutic development of mGlu7 modulators.
Keywords: MeCP2, MECP2 Duplication syndrome, Rett syndrome, mGlu7, ADX71743, negative allosteric modulator
Graphical abstract
Introduction
Loss-of-function mutations in the methyl-CpG-binding protein 2 (MECP2) gene are present in 95% of patients with the neurodevelopmental disorder Rett syndrome (RTT).1,2 Conversely, duplication of the MECP2 locus results in a syndrome characterized by infantile hypotonia, recurrent respiratory infections, limited speech, seizures, intellectual disability, and autism-like behaviors.3,4 While decades of research have yielded significant insights into the pathophysiology of RTT, our understanding of MECP2 Duplication syndrome (MDS) remains limited. Recently, Sztainberg et al. reported that antisense oligonucleotide and/or genetic reduction of MECP2 expression improves phenotypes in MeCP2-Tg1 mice, a model of MDS,5 suggesting that symptoms of MDS may be reversible in patients. These data provide rationale to identify processes downstream of MECP2 dysregulation that can be therapeutically targeted to restore proper neuronal function.
In support of this strategy, we previously identified the metabotropic glutamate receptor 7 (mGlu7) as a potential therapeutic target in a clinical population as well as in a mouse model of RTT.6 mGlu7 is a G protein-coupled receptor expressed widely throughout the brain at glutamatergic and GABAergic presynaptic terminals.7,8 mGlu7 activation inhibits neurotransmitter release and plays important roles in synaptic plasticity in the hippocampus and amygdala.9–13 Additionally, a heterozygous GRM7 mutation has been reported in a patient with idiopathic autism and homozygous mutations have been linked to severe neurological disease.14–16 We have demonstrated that GRM7 expression is activated by MeCP2 binding in vitro and that mGlu7 protein expression is decreased in brain samples from RTT patient autopsies and model mice.6 Furthermore, positive allosteric modulation of mGlu7 activity restores long-term potentiation at Schaffer Collateral CA1 (SCCA1) synapses in the hippocampus and improves cognition, social interaction and respiratory phenotypes in Mecp2+/− female mice.6
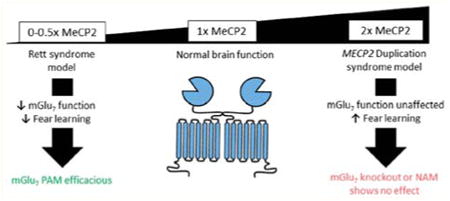
From our findings in RTT model mice, we hypothesized that MeCP2 overexpression would drive increased mGlu7 expression in MeCP2-Tg1 mice. This hypothesis was consistent with previous literature reporting that MeCP2-Tg1 mice display opposing hippocampal phenotypes when compared to RTT model mice. Specifically, slices from MeCP2-Tg1 mice have been shown to display increased paired-pulse ratio and enhanced long-term potentiation at SC-CA1 synapses.5,17 Behaviorally, MeCP2-Tg1 mice exhibit elevated contextual fear freezing at 20 weeks of age.17 Another model of MDS, the Tau-Mecp2 model, also exhibits elevated fear memory, suggesting that overexpression of Mecp2 in the CNS underlies this phenotype.18 Given that positive modulation of mGlu7 activity restores decreased LTP and contextual fear freezing in Mecp2+/− mice to wildtype (WT) levels, we further hypothesized that genetic reduction or negative modulation of mGlu7 activity would normalize hippocampal-dependent phenotypes in MeCP2-Tg1 animals.
Here, we report that mGlu7 protein expression is upregulated in the hippocampus of MeCP2-Tg1 mice, but this increase does not translate to a functional increase in mGlu7 activity at SC-CA1 synapses. In contrast to our hypothesis, genetic reduction or negative modulation of mGlu7 activity shows no efficacy in ameliorating exaggerated contextual fear learning or anxiety-like behavior in this mouse model of MDS.
Results and Discussion
To test the hypothesis that MeCP2 overexpression leads to increased mGlu7 expression, we isolated cortex, hippocampus, and striatum samples from 20-week-old MeCP2-Tg1 mice and WT littermates on a pure FVB/N background for mRNA and protein analysis. Grm7 mRNA expression detected by quantitative real-time polymerase chain reaction (qRT-PCR) was unchanged in all brain regions examined (Figure 1A). Western blotting confirmed global MeCP2 overexpression by 2–3 fold in samples from MeCP2-Tg1 mice (Figure 1B,C); however, elevated mGlu7 protein expression was only detected in hippocampal tissue (Figure 1B,D). Representative full blots can be found in Supplementary Figures 1–2.
Figure 1.
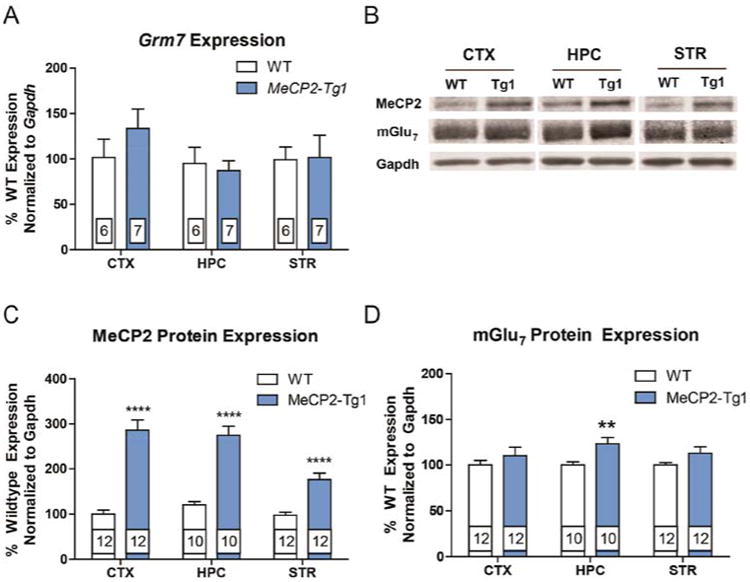
Total mGlu7 protein expression is increased in the hippocampus of MeCP2-Tg1 mice. (A) Grm7 mRNA expression is not changed in samples from MeCP2-Tg1 mice (N = 6–7 samples per genotype). (B) Representative Western blots showing total mGlu7 and MeCP2 protein expression. (C) Quantification of MeCP2 expression from total protein isolates: Cortex (WT 100.1 ± 8.7% (N = 12) vs MeCP2-Tg1 286.6 ± 22.5% (N = 12), ****p < 0.0001), hippocampus (WT 110 ± 6.2% (N = 10) vs MeCP2-Tg1 232.3 ± 17.6% (N = 10), ****p < 0.0001), striatum (WT 97.5 ± 6.7% (N = 12) vs 176.6 ± 13.8% (N = 12), ****p < 0.0001). Student's t tests for each region. (D) Quantification of mGlu7 expression from total protein isolates: Cortex (WT 100 ± 4.8% (N = 12) vs MeCP2-Tg1 110.2 ± 9.2% (N = 12)), hippocampus (WT 100.0 ± 3.5% (N = 10) vs 123.0 ± 7.2% (N = 10), **p < 0.01), striatum (WT 100 ± 2.5% (N = 12) vs MeCP2-Tg1 112.7 ± 7.1% (N = 12)); Student's t tests for each region.
To characterize the functional effects of elevated mGlu7 protein expression within the hippocampus, we performed ex vivo brain slice electrophysiology at SC-CA1 synapses. Similar to previous reports,5,17 slices from 20-week-old MeCP2-Tg1 mice displayed unchanged input–output curves (Figure 2A); however, decreased input-output slope has been reported in younger mice.19 Increased paired-pulse ratio was observed (Figure 2B), which often corresponds with a decrease in presynaptic glutamate release. Since mGlu7 is a presynaptic regulator of neurotransmitter release, this result was consistent with increased mGlu7 protein expression observed in total protein extracted from hippocampal tissue (Figure 1B,D) and directly opposed our findings in Mecp2−/y and Mecp2+/− animals.6
Figure 2.
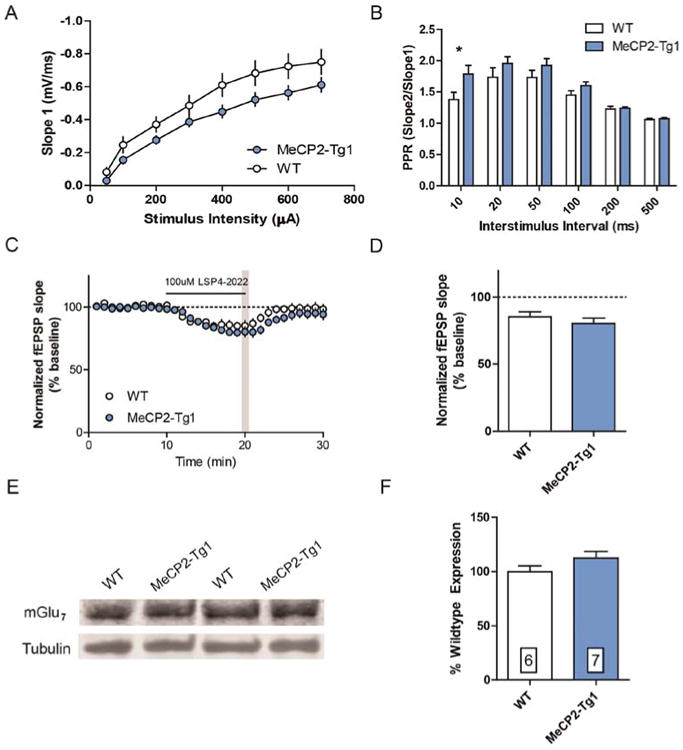
mGlu7 function is not affected by MECP2 duplication at SC-CA1 synapses in mice. (A) Input–output curves were not significantly different between slices derived from 20-week-old WT and MeCP2-Tg1 mice (N = 8 slices/5–6 mice). Two-way ANOVA. (B) Paired pulse ratios were significantly increased at a 10 ms interstimulus interval in MeCP2-Tg1 slices (N = 7 slices/7 mice, *p < 0.05). Two-way ANOVA with Bonferroni post hoc comparisons. (C, D) Depression by 100 μM LSP4–2022 at minute 20 (gray bar) was not significantly different between genotypes. WT 85.4 ± 3.8 (N = 8 slices, 5 mice) vs MeCP2-Tg1 80.2 ± 4.1 (N = 8 slices, 6 mice), Student's t test. (E) Representative mGlu7 Western blots from synaptosomal protein isolates. (F) Quantification of mGlu7 protein expression was not significantly different between genotypes. WT 100 ± 5.2% (N = 6 mice) vs MeCP2-Tg1 112.4 ± 6.2% (N = 7 mice). Student's t test.
To test whether mGlu7 receptor activity was increased, we recorded field excitatory postsynaptic potentials (fEPSPs) at SC-CA1 and bath applied the Group III mGlu receptor agonist, LSP4-2022. Application of 100 μM LSP4–2022 depressed synaptic transmission in WT and MeCP2-Tg1 slices, with no difference in the maximal depression of fEPSP slope (Figure 2C,D). While LSP4-2022 also activates mGlu4 and mGlu8, mGlu7 is believed to be the only presynaptic mGlu receptor at SC-CA1 synapses, making this an ideal synapse to probe mGlu7 function.20,21 Since an increase in total mGlu7 protein expression did not translate to an increase in mGlu7 activity in this functional readout, we next tested whether mGlu7 expression was increased in synaptosome fractions. In contrast to the total protein Western blots, we did not observe a significant increase in mGlu7 expression in synaptosome fractions (Figure 2E,F). Taken together, these data suggest that increased cellular mGlu7 does not necessarily lead to increased receptor surface expression and activity at this synapse.
In contrast to Mecp2+/− mice, 20-week-old MeCP2-Tg1 mice have been reported to display increased LTP at SC-CA1 when compared to littermate controls.5,17 We have previously demonstrated that activation of mGlu7 on GABAergic interneurons is required for the induction of LTP at this synapse; mGlu7 activation decreases GABA release and leads to the disinhibition of excitatory neurons.11 We therefore hypothesized that negative modulation of mGlu7 activity might normalize LTP in MeCP2-Tg1 slices despite the lack of increase in receptor function. We induced LTP with high frequency stimulation (two 1s trains at 100 Hz, 20s interstimulus interval) and monitored fEPSPs for 60 min; however, contrary to previous reports,5,17 we observed no change in LTP between WT and MeCP2-Tg1 slices (Figure 3).
Figure 3.
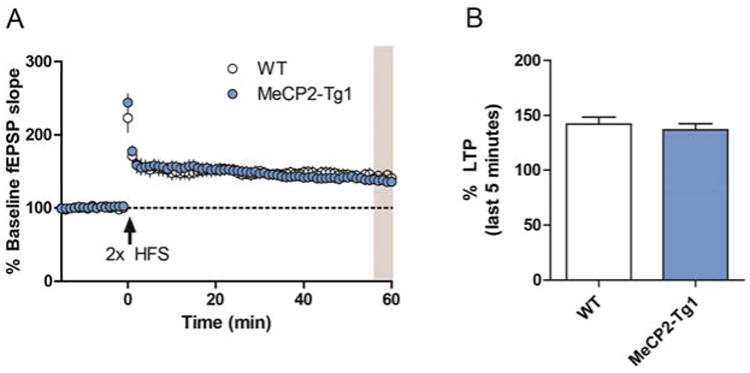
LTP induced by HFS is unchanged at the SC-CA1 synapses in slices from 20-week-old MeCP2-Tg1 mice. (A) After a 15 min baseline recording, LTP was induced by two trains of HFS, and fEPSPs were monitored for 60 min. (B) Quantification of percent LTP during the last 5 min of recording (gray bar). WT 142.1 ± 6.4 (N = 8 slices/9 mice) vs MeCP2-Tg1 137.0 ± 5.4 (N = 13 slices, 9 mice), Student's t test.
MeCP2-Tg1 mice exhibit increased anxiety-like behavior and increased contextual fear learning,17,22 which are phenotypes opposite to those observed in Grm7−/− mice and Mecp2+/− mice.23,24 We previously demonstrated that increasing mGlu7 activity with the positive allosteric modulator (PAM) VU0422288 normalizes anxiolytic behavior and fear learning in Mecp2+/− mice. We therefore hypothesized that either a genetic or pharmacological reduction of mGlu7 function might rescue behavior in MeCP2-Tg1 mice in a similar manner. We crossed female MeCP2-Tg1 mice (FVB/N) with male Grm7+/− mice (C57BL/6J) to genetically reduce mGlu7 expression in the context of MeCP2 overexpression and studied the F1 offspring. We first confirmed that mGlu7 protein levels were significantly reduced in total protein and synaptosomal isolates from these animals (Figure 4A,B). We next performed the elevated plus maze to examine anxiety-like behavior and contextual fear conditioning as a measure of hippocampal-dependent memory. Genetic reduction of mGlu7 expression did not mitigate increased anxiety-like behavior in MeCP2-Tg1 mice as measured by time spent in the open arms of an elevated plus maze (Figure 4C). Furthermore, independent of Grm7 genotype, mice with the MECP2 transgene displayed heightened freezing when re-exposed to the conditioning context 24 h post training along with delayed extinction learning when re-exposed to the conditioning context for 7 subsequent days (Figure 4D).
Figure 4.
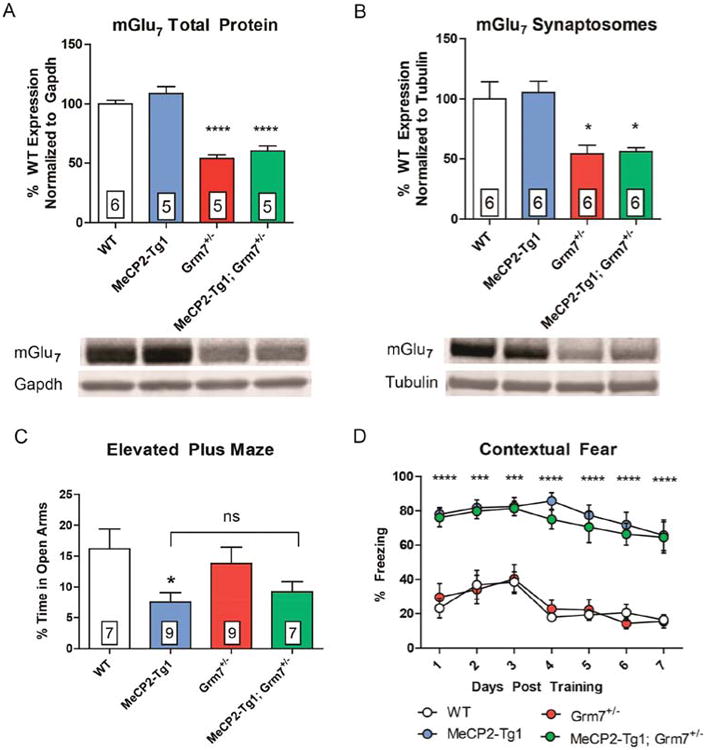
Genetic reduction of mGlu7 does not affect anxiety or fear behavior in MeCP2-Tg1 mice. (A) mGlu7 expression in total protein isolates from hippocampal tissue in MeCP2-Tg1 mice compared relative to WT (WT 100.0 ± 3.1%, MeCP2-Tg1 108.6 ± 5.8%, Grm7+/- 54.0 ± 3.0%, MeCP2-Tg1; Grm7+/− 60.2 ± 4.5%, N = 5–6 mice per genotype, ****p < 0.0001, one-way ANOVA, Bonferroni comparisons relative to WT). (B) mGlu7 expression from synaptosomal isolates from hippocampal tissue (WT 100.0 ± 14.1, MeCP2-Tg1 105.3 ± 9.3, Grm7+/− 54.2 ± 7.3, MeCP2-Tg1; Grm7+/− 55.8 ± 3.7, N = 6 mice per genotype, *p < 0.05, one-way ANOVA, Bonferroni comparisons relative to WT). (C) MeCP2-Tg1 mice spend less time in the open arms of an elevated plus maze relative to WT mice, regardless of Grm7 genotype (N = 7–9 mice per genotype, *p < 0.05, one-way ANOVA, Bonferroni comparisons). (D) MeCP2-Tg1 mice exhibit increased contextual fear freezing compared to those that do not, regardless of Grm7 genotype (N = 7–10 mice per genotype, ***p < 0.001, ****p < 0.0001, two-way ANOVA, Bonferroni comparisons to WT).
As genetic reduction of mGlu7 could have evoked developmental compensatory mechanisms, we next sought to confirm that acute negative modulation of mGlu7 signaling pharmacologically had no impact on phenotypes in adult MeCP2-Tg1 mice. To assess anxiety phenotypes, WT and MeCP2-Tg1 mice were pretreated with either vehicle or the mGlu7 NAM ADX7174325 and tested in the elevated plus maze assay. Similar to genetic reduction of mGlu7, administration of ADX71743 did not alter anxiety phenotypes in WT and MeCP2-Tg1 mice (Figure 5A). To determine whether mGlu7 negative modulation had any effect on learning and memory phenotypes, WT and MeCP2-Tg1 mice were pretreated with a single dose of ADX71743 prior to fear conditioning, and tested for contextual fear memory 24 h later. Relative to vehicle-treated controls, MeCP2-Tg1 mice treated with vehicle exhibited a significant increase in freezing when re-exposed to the context, and ADX71743 administration had no effect on freezing in MeCP2-Tg1 mice (Figure 5B). Conversely, WT mice treated with ADX71743 also presented with significantly increased freezing relative to vehicle-treated controls, and failed to extinguish the fear memory in a manner similar to MeCP2-Tg1 mice (Figure 5C,D).
Figure 5.
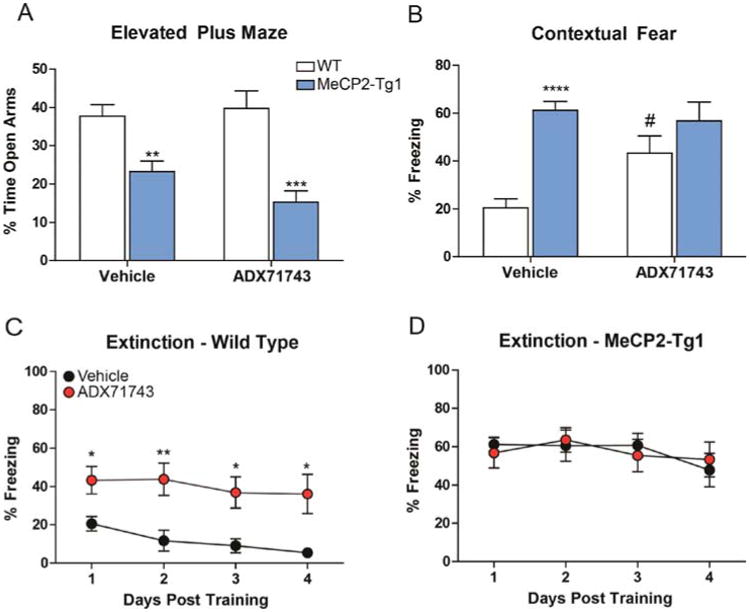
mGlu7 negative allosteric modulation does not affect anxiety or fear behavior in MeCP2-Tg1 mice. (A) Vehicle-treated MeCP2-Tg1 mice spent significantly less time in the open arms of the elevated plus maze relative to WT controls (WT: 37.8 ± 3.0% (N = 16) vs MeCP2-Tg1: 23.2 ± 2.8% (N = 18), **p < 0.01), and ADX71743 (60 mg/kg) administration had no effect on this phenotype (WT: 39.8 ± 4.6% (N = 9) vs MeCP2-Tg1: 15.3 ± 3.0%, (N = 7) ***p < 0.001). Two-way ANOVA with Bonferroni post hoc analysis. (B) Relative to WT mice, MeCP2-Tg1 mice treated with vehicle exhibited an enhanced contextual fear freezing response (WT: 20.5 ± 3.7% (N = 22) vs MeCP2-Tg1: 61.2 ± 3.7% (N = 17), ****p < 0.0001). ADX71743 pretreatment had no effect in MeCP2-Tg1 mice relative to the vehicle-treated mice of the same genotype (MeCP2-Tg1:56.8 ± 7.9% (N = 11)); however, treatment with ADX71743 resulted in a significant increase in freezing in WT mice (WT: 43.3 ± 7.2% (N = 12), #p 0.05, # denotes within genotype comparison. Two-way ANOVA with student's t test post analysis). (C,D) Wild-type mice treated with ADX71743 during training demonstrated a significantly attenuated ability to extinguish fear memory relative to vehicle-treated controls (*p < 0.05, **p < 0.01). ADX71743 administration had no effect on extinction in MeCP2-Tg1 mice. Two-way ANOVA with Bonferroni post hoc analysis.
To complement our mGlu7 studies in RTT model mice, the experiments here aimed to evaluate the therapeutic potential of mGlu7 receptor negative modulation in a mouse model of MDS. We previously reported that MeCP2 binding to the GRM7 promoter activates gene transcription in vitro. In brain tissue samples from Mecp2-/y mice, mGlu7 mRNA and protein expression were decreased in a brain-region specific manner.6 Here, we tested the hypothesis that overexpression of MeCP2 in vivo would drive increased mGlu7 expression. Interestingly, the only significant change observed in samples from MeCP2-Tg1 mice was an increase in total cellular mGlu7 in the hippocampus. These findings suggest that the relationship between MeCP2 dysregulation and mGlu7 expression may be more complex than initially appreciated and dependent on the brain region examined. It is possible that basal expression of mGlu7 is maintained at, or near, maximal levels or that another process independent of MeCP2 is rate-limiting, such that MeCP2 overexpression has no effect on mGlu7 protein levels in some brain regions. Further studies will be needed to fully elucidate the molecular link between MeCP2, GRM7 mRNA, and mGlu7 protein expression.
Independent of receptor expression, we hypothesized that mGlu7 genetic reduction or negative modulation could be an effective approach to reduce symptoms in MeCP2-Tg1 mice due to the established role of mGlu7 activation in LTP induction at SC-CA1 synapses.11 However, despite extensive efforts, we were unable to replicate previous reports that LTP is elevated in slices from MeCP2-Tg1 animals.5,17 This could be due to differences in strain, age, genetic drift or technical/experimental differences. Interestingly, Tau-Mecp2 mice exhibit decreased HFS-induced LTP at SC-CA1 despite exaggerated fear learning.18 Together, these seemingly discordant results highlight that the effect of MeCP2 overexpression on LTP at SC-CA1 synapses appears to be complex and that, at least for this form of LTP, may not correlate with anxiety or fear memory phenotypes in mouse models of MeCP2 over-expression. In our experiments, one single administration of ADX71743 was given prior to fear conditioning to mimic our previous experiments in RTT model mice; however, if the primary deficit in MeCP2-Tg1 mice is memory extinction as opposed to acquisition, we do not exclude the possibility that compound administration on test days may be efficacious.
In summary, we tested the hypothesis that mGlu7 expression is reciprocally regulated by MeCP2 such that negative modulation of mGlu7 would be efficacious in a mouse model of MDS. However, mGlu7 expression was increased only in the hippocampus of MeCP2-Tg1 mice, and behavioral phenotypes were found to be insensitive to changes in mGlu7 expression/activity. Interestingly, Lu et al. recently reported that mouse models of RTT and MDS share similar hippocampal circuit abnormalities including hypersynchronous firing.26 Our findings further support the idea that mouse models of MDS are not simply antiparallel to RTT models, but instead exhibit a unique pathophysiology that warrants further study.
Methods
Chemicals
ADX71743 and LSP4-2022 were synthesized in-house as described in ref 11. For in vivo experiments, ADX71743 was formulated using 10% Tween 20 as vehicle.
Animals
All animals used in the present study were group housed with food and water given ad libitum and maintained on a 12 h light/dark cycle. Animals were cared for in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. All studies were approved by the Institutional Animal Care and Use Committee for Vanderbilt University School of Medicine and took place during the light phase. MeCP2-Tg1 breeders were obtained from The Jackson Laboratory (FVB-Tg(MECP2)1 Hzo/J, stock no. 008679). Male mice were aged until 20–25 weeks of age for all experiments. Grm7 knockout mice were cryorecovered from the Mutant Mouse Regional Resource Center (B6.129P2-Grm7Tm1Dgen/Mmnc) and maintained under identical conditions.
Quantitative Real-Time PCR (QRT-PCR)
Cortex, hippocampus, and striatum were microdissected from 20-week-old MeCP2-Tg1 mice and WT littermates. Total RNA was prepared from tissue samples using TRIzol Reagent (Invitrogen) in accordance with manufacturer's instructions. Total RNA from each brain region was DNase-treated with Roche Turbo DNase kit, and cDNA from 2 μg of total RNA was synthesized using the VILO kit (Invitrogen). RT-qPCR on cDNA from 25 ng of initial RNA template was then run in triplicate using Taqman Fast Reagent Mix (Life Technologies) and Life Technologies gene expression assays for Grm7 (Mm0118924_m1) and Gapdh (Mm03302249_g1). Ct values for each sample were normalized to Gapdh expression and analyzed using the delta–delta Ct method as described in Gogliotti et al.27 Values exceeding two times the standard deviation were classified as outliers. Each value was compared to the average delta-Ct value acquired for wild-type mice and calculated as percent-relative to the average control delta-Ct.
Total and Synaptosomal Protein Preparation
For total protein isolation, tissue samples from 20-week-old MeCP2-Tg1 mice and WT littermates were homogenized using a hand-held motorized mortar and pestle in radioimmunoprecipitation assay buffer (RIPA) containing 10 mM Tris-HCL, 150 mM NaCl, 1 mM ethyl-enediaminetetraacetic acid (EDTA), 0.1% sodium dodecyl sulfate (SDS), 1% Triton X-100, and 1% deoxycholate. After homogenization, samples were spun for 20 min at 15 000g at 4 °C. The supernatant was then transferred to new tubes, and the protein concentration was determined using a bicinchoninic acid (BCA) protein assay (Pierce). For synaptosome preparations, tissue was homogenized in 9 mL of ice-cold sucrose/HEPES (0.32 M sucrose, 4.2 mM HEPES, pH 7.4) using a Teflon-glass homogenizer (Wheaton Science Products). The homogenate was centrifuged at 1000g for 5 min at 4 °C, and the resultant supernatant was centrifuged at 12 000g for 15 min at 4 °C. The final pellets containing synaptosomes were resuspended in RIPA buffer, and protein concentration was determined by BCA assay.
SDS-Page and Western blotting
Proteins (50 μg) were electrophoretically separated using a 4–20% SDS polyacrylamide gel and then transferred onto a nitrocellulose membrane (Bio-Rad). Membranes were blocked with Odyssey blocking buffer (LiCor) for 1 h at room temperature. Membranes were probed with the following primary antibodies: rabbit anti-MeCP2 (1:1000, Millipore 07–013), rabbit anti-mGlu7 antibody (1:1000, Millipore 07–239), mouse anti-Gapdh antibody (1:5000, ThermoFisher MA5-15738), and mouse anti-Tubulin antibody (1:500, Abcam ab44928) overnight at 4 °C. Membranes were washed three times with Tris-buffered saline and Tween 20 (TBS-T, 25 mM Tris, 150 mM NaCl, 0.05% Tween 20) and then incubated with either a goat anti-rabbit fluorescent secondary antibody (800, 1:5000, LiCor) or a goat anti-mouse fluorescent secondary antibody (680, 1:5000, LiCor). Fluorescence was then quantified using Image Studio Light software. Each value for MeCP2 and mGlu7 protein expression was first normalized to the value calculated for Gapdh expression (total protein blots) or Tubulin expression (synaptosome blots).
Extracellular Field Potential Recordings
Coronal brain slices were prepared from 20-week-old MeCP2-Tg1 mice and WT littermates. Mice were anesthetized with isoflurane and decapitated. Brains were rapidly removed and submerged in ice-cold sucrose cutting buffer containing: 230 mM sucrose, 2.5 mM KCl, 8 mM MgSO4, 0.5 mM CaCl2, 1.25 mM NaH2PO4, 10 mM glucose, and 26 mM NaHCO3 saturated with 95%/5% O2/CO2. A block of tissue containing hippocampus was trimmed, embedded in agarose, and coronal slices 400 μm thick were cut using a Compresstome VF-200 (Precisionary Instruments). Slices were transferred to a holding chamber containing N-methyl-d-glucamine (NMDG)-HEPES recovery solution (in mM, 93 NMDG, 2.5 KCl, 1.2 NaH2PO4, 30 NaHCO3, 20 HEPES, 25 d-glucose, 5 sodium ascorbate, 2 thiourea, 3 sodium pyruvate, 10 MgSO4, 0.5 CaCl2, pH 7.3, 305 mOsm) for 15 min at 32 °C. Slices were then transferred to a room temperature modified artificial cerebral spinal fluid (ACSF) containing (in mM) 126 NaCl, 1.25 NaH2PO4, 2.5 KCl, 10 d-glucose, 26 NaHCO3, 2 CaCl2 and 1 MgSO4, and 600 μM sodium ascorbate for at least 1 h. Subsequently, slices were transferred to a submersion recording chamber and continuously perfused (2 mL/min) with ACSF containing (in mM) 126 NaCl, 1.25 NaH2PO4, 2.5 KCl, 10 d-glucose, 26 NaHCO3, 2 CaCl2, 1 MgSO4 heated to 32 °C. All solutions were continuously bubbled with 95%/5% O2/CO2.
A concentric bipolar stimulating electrode was positioned near the CA3–CA1 border, and paired-pulse field excitatory postsynaptic potentials (fEPSPs) were evoked (100 μs duration, every 20 s) by placing a glass recording electrode in the stratum radiatum of CA1. Input–output curves were generated for each slice, and the stimulation intensity was adjusted to 50% of the maximum response for subsequent experiments. Paired-pulse ratios (PPR) were calculated as the ratio between the slope of the second fEPSP divided by the slope of the first fEPSP. PPRs were calculated at several interstimulus intervals (ISI) ranging from 10 to 500 ms.
For LSP4-2022 and LTP experiments, slopes of three consecutive sweeps were averaged and normalized to the average slope during the baseline period. Data were digitized using a Multiclamp 700B, Digidata 1322A, and pClamp 10 software (Molecular Devices). For LSP4-2022 experiments, paired-pulse fEPSPs were generated with a 20 ms ISI at 0.05 Hz. After 10 min of stable baseline recordings, 100 μM LSP4-2022 was applied for 10 min followed by a 10 min washout period. Long-term potentiation (LTP) was induced by applying two trains of 100 Hz stimulation (HFS, 1 s duration, 20 s intertrain interval (ITI)) after a 15 min baseline. ADX71743 (10 μM) was applied for 10 min prior to and during the application of HFS. fEPSPs were monitored for 60 min after HFS, and percent LTP was quantified as the normalized slope during the last 5 min of recording.
Elevated Plus Maze
Twenty-week-old MeCP2-Tg1 mice and WT littermates were habituated to the testing room for 1 h prior to the elevated plus maze test. Mice were placed on the elevated plus maze and allowed to explore freely for 5 min. Time spent exploring each arm was measured using AnyMaze tracking software. For ADX71743 experiments, mice were dosed intraperitoneally (i.p., 10 mL/kg) with 60 mg/kg ADX71743 or vehicle (10% Tween 80) 15 min prior to being placed in the maze.
Fear Conditioning
Twenty-week-old MeCP2-Tg1 mice and WT littermates were habituated to the testing room for 2 h on the day prior to training and the morning of training. On training day, mice were injected i.p. 15 min prior to conditioning with either vehicle (10% Tween 80) or 60 mg/kg ADX71743. Mice were then placed into an operant chamber with a shock grid (Medassociates Inc.) in the presence of a 10% vanilla odor cue. Following a 3 min habituation period, mice were exposed to two 1 s, 0.7 mA foot shocks spaced 30 s apart. For experiments using the offspring of MeCP2-Tg1 and Grm7+/− mice, only one shock was administered due to higher freezing observed in mice of that genetic background. Mice remained in the context for an additional 30 s after the second foot shock. Mice were placed back into the same shock chamber with a 10% vanilla odor cue and the percent of time spent freezing during a 3 min testing period was assessed for up to 7 days after conditioning.
Supplementary Material
Representative full Western blots for mGlu7 and Mecp2 protein analysis (PDF)
Acknowledgments
Funding: This work was supported by a grant from the National Institute of Mental Health to C.M.N (R21 MH102548) and from the National Institute of Neurological Disorders and Stroke to P.J.C. (R37 NS031373). N.M.F. was supported by NIH training Grants T32 GM007628 and F31 MH113259. R.G.G. received support by a Mentored Training Fellowship from Rettsyndrome.org, a Young Investigator Award from Brain and Behavior Research Foundation, and NIMH K01MH112983. B.J.S received support from F32 MH111124. S.D.V. was supported by T32 GM007628.
Abbreviations
- LTP
long-term potentiation
- MDS
MECP2 Duplication syndrome
- MeCP2
methyl CpG binding protein 2
- mGlu7
metabotropic glutamate receptor 7
- NAM
negative allosteric modulator
- PAM
positive allosteric modulator
- RTT
Rett syndrome
- SC-CA1
Schaffer collateral-CA1
- WT
wild type
Footnotes
Supporting Information: The Supporting Information is available free of charge on the ACS Publications website at DOI: 10.1021/acschemneur-o.7b00414.
Author Contributions: N.M.F, R.G.G, and C.M.N. designed the experiments. N.M.F, R.G.G, S.D.V., and B.J.S. performed the experiments and data analyses. C.M.N. and P.J.C. contributed reagents and other resources. N.M.F, R.G.G, and C.M.N. wrote the manuscript with input from all authors.
Notes: The authors declare no competing financial interest.
References
- 1.Amir RE, Van den Veyver IB, Wan M, Tran CQ, Francke U, Zoghbi HY. Rett syndrome is caused by mutations in X-linked MECP2, encoding methyl-CpG-binding protein 2. Nat Genet. 1999;23:185–188. doi: 10.1038/13810. [DOI] [PubMed] [Google Scholar]
- 2.Neul JL, Fang P, Barrish J, Lane J, Caeg EB, Smith EO, Zoghbi H, Percy A, Glaze DG. Specific mutations in methyl-CpG-binding protein 2 confer different severity in Rett syndrome. Neurology. 2008;70:1313–1321. doi: 10.1212/01.wnl.0000291011.54508.aa. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Van Esch H, Bauters M, Ignatius J, Jansen M, Raynaud M, Hollanders K, Lugtenberg D, Bienvenu T, Jensen LR, Gecz J, Moraine C, Marynen P, Fryns JP, Froyen G. Duplication of the MECP2 region is a frequent cause of severe mental retardation and progressive neurological symptoms in males. Am J Hum Genet. 2005;77:442–453. doi: 10.1086/444549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Del Gaudio D, Fang P, Scaglia F, Ward PA, Craigen WJ, Glaze DG, Neul JL, Patel A, Lee JA, Irons M, Berry SA, Pursley AA, Grebe TA, Freedenberg D, Martin RA, Hsich GE, Khera JR, Friedman NR, Zoghbi HY, Eng CM, Lupski JR, Beaudet AL, Cheung SW, Roa BB. Increased MECP2 gene copy number as the result of genomic duplication in neurodevelopmentally delayed males. Genet Med. 2006;8:784–792. doi: 10.1097/01.gim.0000250502.28516.3c. [DOI] [PubMed] [Google Scholar]
- 5.Sztainberg Y, Chen H, Swann JW, Hao S, Tang B, Wu Z, Tang J, Wan YW, Liu Z, Rigo F, Zoghbi HY. Reversal of phenotypes in MECP2 duplication mice using genetic rescue or antisense oligonucleotides. Nature. 2015;528:123–126. doi: 10.1038/nature16159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gogliotti RG, Senter RK, Fisher NM, Adams J, Zamorano R, Walker AG, Blobaum AL, Engers DW, Hopkins CR, Daniels JS, Jones CK, Lindsley CW, Xiang Z, Conn PJ, Niswender CM. mGlu7 potentiation rescues cognitive, social, and respiratory phenotypes in a mouse model of Rett syndrome. Sci Transl Med. 2017;9:eaai7459. doi: 10.1126/scitranslmed.aai7459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Somogyi P, Dalezios Y, Luján R, Roberts JDB, Watanabe M, Shigemoto R. High level of mGluR7 in the presynaptic active zones of select populations of GABAergic terminals innervating interneurons in the rat hippocampus. Eur J Neurosci. 2003;17:2503–2520. doi: 10.1046/j.1460-9568.2003.02697.x. [DOI] [PubMed] [Google Scholar]
- 8.Bradley SR, Rees HD, Yi H, Levey AI, Conn PJ. Distribution and developmental regulation of metabotropic glutamate receptor 7a in rat brain. J Neurochem. 1998;71:636–645. doi: 10.1046/j.1471-4159.1998.71020636.x. [DOI] [PubMed] [Google Scholar]
- 9.Martín R, Torres M, Sánchez-Prieto J. mGluR7 inhibits glutamate release through a PKC-independent decrease in the activity of P/Q-type Ca2+ channels and by diminishing cAMP in hippocampal nerve terminals. Eur J Neurosci. 2007;26:312–322. doi: 10.1111/j.1460-9568.2007.05660.x. [DOI] [PubMed] [Google Scholar]
- 10.Gee CE, Peterlik D, Neuhäuser C, Bouhelal R, Kaupmann K, Laue G, Uschold-Schmidt N, Feuerbach D, Zimmermann K, Ofner S, Cryan JF, van der Putten H, Fendt M, Vranesic I, Glatthar R, Flor PJ. Blocking metabotropic glutamate receptor subtype 7 (mGlu7) via the Venus flytrap domain (VFTD) inhibits amygdala plasticity, stress, and anxiety-related behavior. J Biol Chem. 2014;289:10975–10987. doi: 10.1074/jbc.M113.542654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Klar R, Walker AG, Ghose D, Grueter BA, Engers DW, Hopkins CR, Lindsley CW, Xiang Z, Conn PJ, Niswender CM. Activation of Metabotropic Glutamate Receptor 7 Is Required for Induction of Long-Term Potentiation at SC-CA1 Synapses in the Hippocampus. J Neurosci. 2015;35:7600–7615. doi: 10.1523/JNEUROSCI.4543-14.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Fendt M, Schmid S, Thakker DR, Jacobson LH, Yamamoto R, Mitsukawa K, Maier R, Natt F, Hüsken D, Kelly PH, McAllister KH, Hoyer D, van der Putten H, Cryan JF, Flor PJ. mGluR7 facilitates extinction of aversive memories and controls amygdala plasticity. Mol Psychiatry. 2008;13:970–979. doi: 10.1038/sj.mp.4002073. [DOI] [PubMed] [Google Scholar]
- 13.Millán C, Luján R, Shigemoto R, Sánchez-Prieto J. The inhibition of glutamate release by metabotropic glutamate receptor 7 affects both [Ca2+]c and cAMP: evidence for a strong reduction of Ca2+ entry in single nerve terminals. J Biol Chem. 2002;277:14092–14101. doi: 10.1074/jbc.M109044200. [DOI] [PubMed] [Google Scholar]
- 14.Liu Y, Zhang Y, Zhao D, Dong R, Yang X, Tammimies K, Uddin M, Scherer SW, Gai Z. Rare de novo deletion of metabotropic glutamate receptor 7 (GRM7) gene in a patient with autism spectrum disorder. Am J Med Genet, Part B. 2015;168B:258–264. doi: 10.1002/ajmg.b.32306. [DOI] [PubMed] [Google Scholar]
- 15.Reuter MS, Tawamie H, Buchert R, Hosny Gebril O, Froukh T, Thiel C, Uebe S, Ekici AB, Krumbiegel M, Zweier C, Hoyer J, Eberlein K, Bauer J, Scheller U, Strom TM, Hoffjan S, Abdelraouf ER, Meguid NA, Abboud A, Al Khateeb MA, Fakher M, Hamdan S, Ismael A, Muhammad S, Abdallah E, Sticht H, Wieczorek D, Reis A, Abou Jamra R. Diagnostic yield and novel candidate genes by exome sequencing in 152 consanguineous families with neurodevelopmental disorders. JAMA Psychiatry. 2017;74:293–299. doi: 10.1001/jamapsychiatry.2016.3798. [DOI] [PubMed] [Google Scholar]
- 16.Charng WL, Karaca E, Coban Akdemir Z, Gambin T, Atik MM, Gu S, Posey JE, Jhangiani SN, Muzny DM, Doddapaneni H, Hu J, Boerwinkle E, Gibbs RA, Rosenfeld JA, Cui H, Xia F, Manickam K, Yang Y, Faqeih EA, Al Asmari A, Saleh MAM, El-Hattab AW, Lupski JR. Exome sequencing in mostly consanguineous Arab families with neurologic disease provides a high potential molecular diagnosis rate. BMC Med Genomics. 2016;9:42. doi: 10.1186/s12920-016-0208-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Collins AL, Levenson JM, Vilaythong AP, Richman R, Armstrong DL, Noebels JL, David Sweatt J, Zoghbi HY. Mild overexpression of MeCP2 causes a progressive neurological disorder in mice. Hum Mol Genet. 2004;13:2679–2689. doi: 10.1093/hmg/ddh282. [DOI] [PubMed] [Google Scholar]
- 18.Na ES, Nelson ED, Adachi M, Autry AE, Mahgoub MA, Kavalali ET, Monteggia LM. A mouse model for MeCP2 duplication syndrome: MeCP2 overexpression impairs learning and memory and synaptic transmission. J Neurosci. 2012;32:3109–3117. doi: 10.1523/JNEUROSCI.6000-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Stansley BJ, Fisher NM, Gogliotti RG, Lindsley CW, Conn PJ, Niswender CM. Contextual fear extinction induces hippocampal metaplasticity mediated by metabotropic glutamate receptor 5. Cereb Cortex. 2017:1–14. doi: 10.1093/cercor/bhx282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Baskys A, Malenka RC. Agonists at metabotropic glutamate receptors presynaptically inhibit EPSCs in neonatal rat hippocampus. J Physiol (Oxford, U K) 1991;444:687–701. doi: 10.1113/jphysiol.1991.sp018901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ayala JE, Niswender CM, Luo Q, Banko JL, Conn PJ. Group III mGluR regulation of synaptic transmission at the SC-CA1 synapse is developmentally regulated. Neuropharmacology. 2008;54:804–814. doi: 10.1016/j.neuropharm.2007.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Samaco RC, Mandel-Brehm C, McGraw CM, Shaw CA, McGill BE, Zoghbi HY. Crh and Oprm1 mediate anxiety-related behavior and social approach in a mouse model of MECP2 duplication syndrome. Nat Genet. 2012;44:206–211. doi: 10.1038/ng.1066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Fendt M, Imobersteg S, Peterlik D, Chaperon F, Mattes C, Wittmann C, Olpe HR, Mosbacher J, Vranesic I, van der Putten H, McAllister KH, Flor PJ, Gee CE. Differential roles of mGlu(7) and mGlu(8) in amygdala-dependent behavior and physiology. Neuropharmacology. 2013;72:215–223. doi: 10.1016/j.neuropharm.2013.04.052. [DOI] [PubMed] [Google Scholar]
- 24.Masugi M, Yokoi M, Shigemoto R, Muguruma K, Watanabe Y, Sansig G, van der Putten H, Nakanishi S. Metabotropic glutamate receptor subtype 7 ablation causes deficit in fear response and conditioned taste aversion. J Neurosci. 1999;19:955–963. doi: 10.1523/JNEUROSCI.19-03-00955.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kalinichev M, Rouillier M, Girard F, Royer-Urios I, Bournique B, Finn T, Charvin D, Campo B, Le Poul E, Mutel V, Poli S, Neale SA, Salt TE, Lütjens R. ADX71743, a potent and selective negative allosteric modulator of metabotropic glutamate receptor 7: in vitro and in vivo characterization. J Pharmacol Exp Ther. 2013;344:624–636. doi: 10.1124/jpet.112.200915. [DOI] [PubMed] [Google Scholar]
- 26.Lu H, Ash RT, He L, Kee SE, Wang W, Yu D, Hao S, Meng X, Ure K, Ito-Ishida A, Tang B, Sun Y, Ji D, Tang J, Arenkiel BR, Smirnakis SM, Zoghbi HY. Loss and Gain of MeCP2 Cause Similar Hippocampal Circuit Dysfunction that Is Rescued by Deep Brain Stimulation in a Rett Syndrome Mouse Model. Neuron. 2016;91:739–747. doi: 10.1016/j.neuron.2016.07.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gogliotti RG, Senter RK, Rook JM, Ghoshal A, Zamorano R, Malosh C, Stauffer SR, Bridges TM, Bartolome JM, Daniels JS, Jones CK, Lindsley CW, Conn PJ, Niswender CM. mGlu5 positive allosteric modulation normalizes synaptic plasticity defects and motor phenotypes in a mouse model of Rett syndrome. Hum Mol Genet. 2016;25:1990–2004. doi: 10.1093/hmg/ddw074. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Representative full Western blots for mGlu7 and Mecp2 protein analysis (PDF)