

Abstract
Background
Cancer stem cells (CSCs) or tumor-initiating cells (TICs) represent a small population of cancer cells with self-renewal and tumor-initiating properties. Unlike the bulk of tumor cells, CSCs or TICs are refractory to traditional therapy and are responsible for relapse or disease recurrence in cancer patients. Stem cells have distinct metabolic properties compared to differentiated cells, and metabolic rewiring contributes to self-renewal and stemness maintenance in CSCs.
Main body
Recent advances in metabolomic detection, particularly in hyperspectral-stimulated raman scattering microscopy, have expanded our knowledge of the contribution of lipid metabolism to the generation and maintenance of CSCs. Alterations in lipid uptake, de novo lipogenesis, lipid droplets, lipid desaturation, and fatty acid oxidation are all clearly implicated in CSCs regulation. Alterations on lipid metabolism not only satisfies the energy demands and biomass production of CSCs, but also contributes to the activation of several important oncogenic signaling pathways, including Wnt/β-catenin and Hippo/YAP signaling. In this review, we summarize the current progress in this attractive field and describe some recent therapeutic agents specifically targeting CSCs based on their modulation of lipid metabolism.
Conclusion
Increased reliance on lipid metabolism makes it a promising therapeutic strategy to eliminate CSCs. Targeting key players of fatty acids metabolism shows promising to anti-CSCs and tumor prevention effects.
Keywords: Cancer stem cells, Lipid metabolism, Metabolomics, Lipid droplets, Lipid desaturation, de novo lipogenesis, Fatty acid oxidation
Background
Cancer stem cells (CSCs) or tumor initiating cells (TICs) are small subpopulations (0.001–0.1%) of cancer cells that may account for cancer initiation, metastasis, therapy resistance, and recurrence [1, 2]. CSCs exhibit self-renewal and tumor-initiating properties and are able to recapitulate the bulk hierarchy of cancer tissues [3]. Evidence suggests that epithelial-mesenchymal transition (EMT) is associated with stemness acquisition. Cancer cells after EMT usually exhibit stem cell-like characteristics, and thus CSCs are also believed to act as metastasis-initiating cells [4–8]. The origin of CSCs is still under debate. It has been proposed that CSCs may arise from normal stem cells or tissue progenitor cells as a result of stochastic genetic mutations and epigenetic alterations [9]. Nevertheless, some evidence suggests that differentiated cells might also stochastically dedifferentiate into a more primitive state with tumor-initiating potential [10–12]. Kelly et al. demonstrated that large numbers of leukemia-initiating cells (LICs) can recapitulate the bulk hierarchy of cancer in genetically compatible models [11]. They claimed that the rarity of TICs in xenotransplant experiments is mainly a result of the limited ability of human tumor cells to adapt and survive in an alien (mouse) milieu. However, the hierarchical and stochastic models are not mutual exclusive. Though not all cancers follow the hierarchical model [13, 14], the presence of CSCs is clearly demonstrated in various cancer types using distinct cell surface markers and enzymatic assays, including leukemia [15], breast cancer [16], glioblastoma [17], colorectal cancer [18–20], pancreatic cancer [21–23], liver cancer [24], lung cancer [25], and ovarian cancer [26, 27]. For example, in acute myeloid leukemia (AML), CSCs populations are CD34(+) CD38(−) [15]. In breast cancer, CSCs were identified as CD44(+) CD24(−/low) lineage [16] or ALDH1-positive [28] populations. In glioma, the CD133(+) or CD44(high)/Id1(high) fractions are recognized as CSCs [17, 29]. Overexpression of CD44 variant isoform (CD44v), which is mainly generated by ESRP1 and ESRP2 mediated alternative splicing on CD44 mRNA [30], is observed in various CSCs [31] High level of CD44v8–10 protects CSCs from reactive oxygen species (ROS), which is known to play a “double-edged sword” role in cancer development [31].
Autophagy, Ferroptosis and redox regulation in CSCs
It remains a great challenge to eliminate CSCs and improve the survival of patients, because CSCs are typically quiescent and resistant to conventional radio- and chemotherapy [32, 33]. It was believed that CSCs largely contribute to formation of clinically undetectable minimal residual disease(MRD) after conventional anti-tumor therapies, and therefore are implicated in disease persistence or relapse. Alterations in cellular bioenergetics impart CSCs in MRD to develop adaptive or acquired resistance to anti-tumor therapy, thus leading to tumor recurrence [34]. For example, comprehensive transcriptomic and metabolic analyses of oncogene ablation-resistant pancreatic cancer cells possessing CSCs characteristics revealed enhanced mitochondrial respiration but diminished dependence on the Warburg effect, as well as increased autophagy and lysosome activity, suggesting metabolic alterations and active autophagy are critical features of CSCs [35]. Autophagy primarily acts as a lysosomal dependent metabolic-recycling mechanism which is important for cell survival in stress [36–38]. It has been considered that autophagy may exert a anticarcinogenic role in early stage of cancer development by safeguarding against genomic instability through the clearance the old and dysfunctional mitochondria and protein aggregates [36, 39]. Furthermore, autophagy may exert tumor suppressive function through destabilizing the transcription factor nuclear factor erythroid 2-related factor 2 (NRF2), which imparts tumor cells with resistance to redox stress [40]. Nevertheless, active autophagy is recognized as one of the hallmarkers of cancer [41]. In established cancers, constitutive activation of autophagy contributes to accquired therapeutic resistance. For example, active autophagy protects glioblastoma multiforme (GBM) cells from the unfavorable tumor microenvironment characterized by hyper-oxidative, hypoxic, nutrient-poor conditions [42]. Further more, compelling evidence suggests that autophagic-lysosomal pathway largely contributes to generation, maintenance and differentiation of CSCs [43]. Many studies have shown that CSCs frequently have higher basal level of autophagy than that of non-stem cancer cells. Active autophagy help CSCs to rapidly respond to metabolic stress to maintain their energetic balance. For example, the fusion of lipid droplets with autophagosomes, a process named lipophagy, confer a survival advantage on oncogene ablation-resistant pancreatic cancer cells through increase of fatty acid β-oxidation [35]. A number of studies have shown either chemical or genetical blockade of autophagy impairs self-renewal and tumorigenicity of CSCs [44–46]. Recent studies revealed that some of “conventional” agents used in non-cancerous diseases treatment exert antitumor therapeutic effects by modulating autophagic pathway, suggesting that drug re-positioning targeting autophagy may be a promising therapeutic strategies for human malignancies [40].
Ferroptosis is recognized as an iron-dependent form of nonapoptotic cell death implicated in various human diseases, including ischemic tissue damage and human malignant diseases [47]. Recent study unveiled the crucial role of autophagy in ferroptosis. Pharmacological induction of ferroptosis leads to lysosomal degradation of cellular iron storage proteins ferritin and ferritinophagy cargo receptor NCOA4 in an autophagy dependent manner (a process known as ferritinophagy), suggesting the close relationship between ferroptosis and autophagic cell death [48]. Recently, a synthetic derivative of natural product salinomycin named as ironomycin sequesters lysosomal iron and induces ferroptosis, showing a selective antitumor activity against breast CSCs in vitro and in vivo [49]. In addition, ferroptosis is triggered by iron-dependent excessive lipid hydroperoxides accumulation due to insufficient antioxidant glutathione (GSH) level, the cystine/glutamate antiporter system x(c)(−) is likely to be involved [50]. System x(c)(−) is composed of a light chain, xCT, and a heavy chain, 4F2 heavy chain (4F2hc) [50]. Upregulation of xCT contributes to drug resistance in pancreatic cancers [51]. Up-regulation of xCT also has been demonstrated in other cancers in including lymphomas [52], and gliomas [53, 54]. CD44v has recently been shown to involved in the scavenging of ROS via the stabilization of xCT protein at the cellular membrane, thus activation of CD44v-xCT-GSH axis play a crucial role in redox regulation of CSCs and is likely contribute to the relapse and distant metastasis after repeated radiation therapy [55]. Remarkably, chemotherapy is able to induce ectopic expression of CD44v, which is evidenced in osteosarcoma and hepatic cancer cells of the Li-Fraumeni patient [56]. This is probably due to the selective clonal amplification of undetectable number of CD44v8–10-positive CSCs under the pressure of excessive ROS after radiation and chemotherapy.
Metabolic alterations in human Cancer
Cancer cells exhibit a distinct metabolic profile as compared to it’s normal counterparts. Due to the rapid tumor cells proliferation and inadequate blood vessels formation, tumor microenviroments are characterized by hypoxic, hyper-oxidative, acidic and nutrients-poor conditions, therefore cancer cells must adapt it’s cellular bioenergetics efficiently to deal with this kind of unfavorable microenvironments, a process named metabolic reprogramming. Metabolic reprogramming is essential to sustain cancer cells proliferation and survival when the oncogenic signaling is blocked [35, 57]. Most human cancers show constitutive aerobic glycolysis even in oxygen-rich conditions, a phenomenon called Warburg effect [58, 59]. This kind of metabolic rewiring not only satisfies the energy demands for continuous proliferation, but also provides plenty of building blocks for cellular compartments. Metabolic regulation of stemness is increasingly recognized as fundamental in the control of stem cell fate. In contrast to most differentiated cells, pluripotent stem cells (PSCs) rely primarily on aerobic glycolysis rather than mitochondrial oxidative phosphorylation(OXPHOS) to minimize ROS production, which impairs self-renewal ability [60, 61]. Reduced mitochondrial respiration in quiescent hematopoietic stem cells (HSCs) prevent oxidative damage from ROS, enabling long-term survival because HSCs are sensitive to ROS [62]. Aerobic glycolysis also contributes to acquisition of stemness in CSCs. It has been demonstrated that poorly differentiated cancers show much higher glucose uptake than differentiated cancers, suggesting that a high glycolytic flux in tumor tissues mainly results from a blockade of CSCs differentiation [63]. Conversely, activation of mitochondrial metabolism leads to loss of pluripotent potential and induction of differentiation in P19 embryonal carcinoma stem cells [64]. Recently, Peng et al. demonstrated that breast cancer stem cells (BCSCs) have high levels of pyruvate dehydrogenase kinase 1 (PDK1), which inhibits mitochondrial OXPHOS. Depletion of PDK1 significantly diminishes ALDH1-positive BCSCs, which leads to decreased sphere-formation ability [65], suggesting targeting aerobic glycolysis may be usefull to eliminating CSCs. However, to date, attempts to inhibit glycolysis as cancer therapy remain unsatisfactory [66], which is mainly due to CSCs are very heterogeneous and may thus have divergent metabolic landscapes [67]. In addition to glucose, some cancer cells also use glutamine heavily [68]. However, little is known about the role of glutaminolysis in stem cell homeostasis.
Unlike HSCs, normal neuro stem cells(NSCs) show low levels of glycolysis [69], suggesting that the metabolic phenotype of pluripotent cells is highly plastic and strongly influenced by tissue microenviroments and nutrient availability [70]. A growing body of evidences indicates that CSCs/TICs are more dependent on oxidative metabolism than glycolysis [61, 71]. For example, oncogene ablation resistant pancreatic cells with feature of CSCs strongly rely on mitochondrial respiration rather than Warburg effect [35]. CSCs from ovarian cancer primarily rely on fatty acid β-oxidation (FAO) and are resistant to glucose deprivation [72].
Metabolic and redox cues of Phenoconversion in CSCs
As mentioned above, metabolic cues play a central role in cell fate determination. A growing body of literatures indicates that metabolism reprogramming and CSCs properties are two highly entwined processes during tumor development [73]. On one hand, chronic metabolic stress in premalignant environments may drive the phenoconversion of non-stem cancer cells to a stem-like state in a Wnt-dependent manner [74]. In addition, chronic oxidative stress at non-cytotoxic doses promotes neoplastic transformation and stem cell characteristics in kidney epithelial cells [75], suggesting that ROS may act as a “double-edged sword” in the acquisition of stem cell characteristics in a dose-dependent manner. On the other hand, impairment of mitochondrial metabolism via inhibition of complex I or loss of mitochondrial DNA leads to genetic inactivation of p53 and to a glycolytic switch in neural progenitor/stem cells (NPCs), which result in genomic instability and glioma formation and support the notion that metabolic stress triggers the conversion of normal NSCs to a glioma-initiating NSCs [69]. In the established tumor tissues, tumor cells continually undergo persistent and high level of oxidative stress [76]. In terms of the survival against excessive degree of redox stress, CSCs must adapt it’s cellular bioenergetics efficiently to this kind of unfavorable conditions, a NRF2-dependent anti-ROS signal pathway may be involved [34] . Activation of NRF2 promotes tumor cells resistant to redox stress, whereas inactivation of NRF2 with the flavonoid chrysin effectively sensitizes BEL-7402/ADM tumor cells to doxorubicin by downregulating the PI3K/Akt and ERK pathways [77]. Redox balance may contribute to autophagy associated drug resistance, that why NRF2 inhibitors suppress cancer stemness and sensitize GBM cells to temozolomide(TMZ), an alkylating agent for GBM and anaplastic astrocytoma treatment, which induces autophagy and subsequent therapeutic resistance [42, 78]. Recently, Yoshida et al. demonstrated that CD44v but not the standard CD44, promotes proteasome degradation of c-Myc protein via suppressing redox stress-induced Wnt activation [31]. High amount of CD44v8–10 cooperate with Fbw7, a well-defined ubiquitin ligase of c-myc protein [79], precisely regulate the proliferation and dormancy cycle of CSCs through modulating c-myc protein stability at the invasive front [31, 80].
Studies unveiled that glioma CSCs reside in either perivascular niche or perinecrotic microenviroment [81, 82]. In the perivascular niche, glioma CSCs interact closely with endothelial cells which secrete factors to maintain the self-renewal of CSCs [81]. It has been demonstrated that the CD44 ligand osteopontin enriched in perivascular niche promotes glioma CSCs-like phenotypes and radiation resistance. These effects were mediated by HIF-2α in a cooperative manner with γ-secretase generated CD44 intracellular domain [83]. On the other hand, a hypoxic and perinecrotic microenviroment, which known to stimulate glycolysis and induce autophagy, promote acquisition of a stem-like state and increase the CSCs population through stabilization of both HIF1 and HIF2 [82, 84, 85]. It has been shown that two of pluripotency transcription factors, OCT4 and c-Myc, were directly activated by HIF-2α [86, 87].
Alterations in lipid metabolism in CSCs
Though the bulk tumor cells and CSCs share some common metabolic features as compared to normal cells, it has been proposed that the metabolic state of TICs/CSCs subtlely differs from that of non- stem cancer cells [88]. Fatty acids metabolism not only supports energy production but also plays an important role in biosynthetic pathways and redox homeostasis. Recent advances in proteomics and metabolomics have deepened our knowledge of the role of fatty acids metabolism in determining CSCs fate [89–91]. For example, Chen et al. described that NANOG, a master factor in controlling stem cell fate, stimulates the generation of stem-like TICs and hepatocellular carcinoma (HCC) oncogenesis via metabolic reprogramming from OXPHOS to fatty acids oxidation [60], suggesting lipid metabolism is fundamental for NANOG positive CSCs.
Lipids are essential components of cell and organelle membranes, and fatty acids are required for proliferation of the bulk tumor mass and also for CSCs maintenance [89, 92, 93]. There is a strong contribution from the lipid metabolism, whereas the role of glycolysis in CSCs maintenance may be more tumor-specific. For example, glioma stem cells (GSCs) use less glycolysis than differentiated glioma cells but maintain higher levels of ATP production [94]. Further more, the glycolytic intermediates could be used by CSCs for de novo lipogenesis to increase self-renewal growth [66], suggesting different metabolic pathway could be well coordinated in CSCs to maximize the benefits. Both lipid catabolism and anabolism alterations are associated with acquisition of stemness during cancer development (Fig. 1). For example, BCSCs exhibit elevated long-chain FAO metabolites compared to non-stem cancer cells. Moreover, inhibition of FAO by etomoxir markedly decreases viability and tumorsphere-forming potential of BCSCs but exert little effect on non-stem cancer cells, suggesting that FAO is critical to self-renewal of BCSCs [89].
Fig. 1.
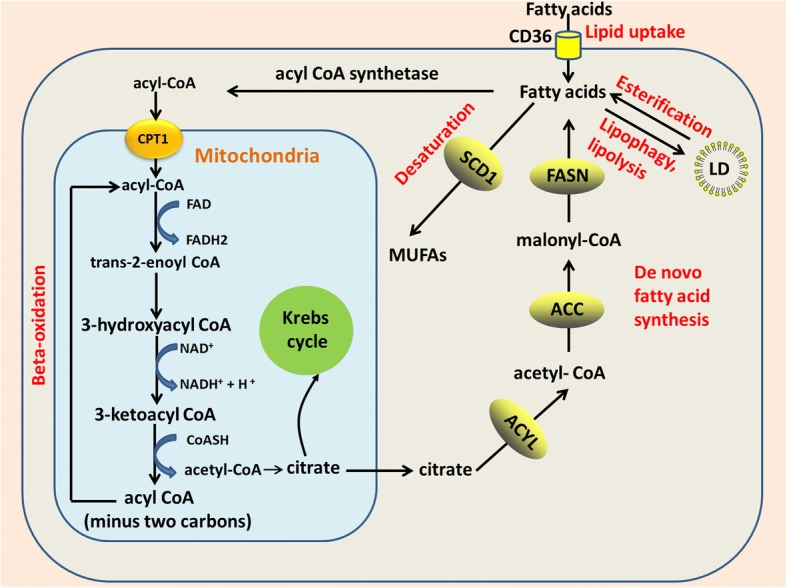
Alterations in lipid metabolism in CSCs. Both lipid catabolism and anabolism alterations contribute to stemness acquisition in CSCs, including lipid uptake, de novo lipogenesis, lipid desaturation, lipolysis, lipophagy, and FAO. Extracellular FFAs are transported into cells via CD36 and then reused via β-oxidation in mitochondria to release acetyl-CoA. Acetyl-CoA is converted to citrate by citrate synthase and then enters the Krebs cycle for complete oxidation. Alternatively, de novo fatty acids synthesis starts with acetyl-CoA and builds up by the addition of two-carbon units. In addition to lipid catabolism, fatty acids are esterified to glycerol and then triglycerides are stored in lipid droplets. Breakdown of lipids droplets via lipolysis or lipophagy enables stored energy mobilization to the mitochondria. Additionally, saturated fatty acids are desaturated into mono-unsaturated fatty acids by SCD1. Alterations in lipid metabolism not only satisfy energy demands for CSCs proliferation, but also provide essential components for biosynthetic pathways and redox homeostasis. ACC, acetyl-CoA carboxylase; ACLY, ATP citrate lyase; FASN, fatty acid synthase; CD36, cluster of differentiation 36; FAs, fatty acids; MUFAs, mono-unsaturated fatty acids; SCD1, stearoyl-CoA desaturase 1; CPT1, carnitine palmitoyltransferase 1; TCA cycle, tricarboxylic acid cycle; FAO, fatty acid oxidation; and LD, lipid droplet
Lipid droplets in CSCs
Lipid droplets (LDs) are intracellular spherical organelles surrounded by a single layer of phospholipids that store lipids [95]. Cancer cells have more LDs compared to normal cells [95]. During metabolic stress resulting from blocked glycolysis, free fatty acids (FFAs) from LDs sustain ATP production though FAO (Fig. 1). Breakdown of LDs in an autophagy-dependent manner, a selective autophagy named lipophagy, enables FFAs mobilization to the mitochondria (Fig. 1), which is pivotal for survival when metabolic restrictions in cancer cells are induced by oncogenic signaling blockade [57, 96]. LDs also protect lipid from peroxidation, as toxic lipid peroxides trigger ferroptosis [47]. In addition, lipid quantification in prostate cancer is associated with the tumor stage, thus serving as a quantitative marker for disease diagnosis [97]. LDs formation is induced by hypoxia via HIF1- and HIF2-mediated repression of carnitine palmitoyltransferase 1A (CPT1A), a key enzyme in involved in mitochondrial FAO [98] (Fig. 2). In addition to de novo lipogenesis, increase in extracellular lipid uptake also contributes to LDs accumulation and tumor initiation capacity in CSCs [99] (Fig. 1).
Fig. 2.
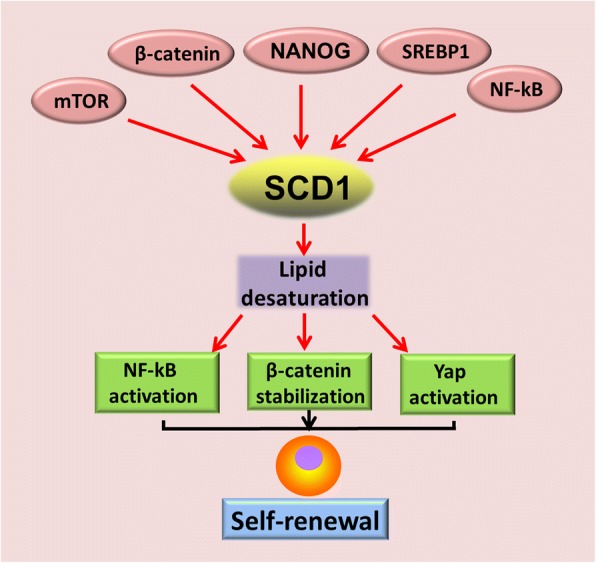
NANOG mediated metabolic reprogramming contributes to CSCs self-renewal and chemoresistance. NANOG binding on FAO genes(Acadvl, Echs1, and Acads) promoters stimulates it’s transcription but exerts opposite effect on OXPHOS genes(Cox6a2 and Cox15) transcription, leading to metabolic switch from OXPHOS to FAO and less ROS production in CSCs/TICs. NANOG also promotes lipid desaturation via up-regulating SCD1 expression. OXPHOS, oxidative phosphorylation; FAO, fatty acid oxidation
CSCs display more LDs compared to the bulk cancer cells in several cancer types. For example, Tirinato et al. demonstrated that colorectal cancer stem cells (CRCSCs) exhibit higher lipid levels compared to normal epithelial colon cells (NECCs), colon carcinoma cells (CCCs), and sphere-derived adherent cancer cells (SDACs). The authors showed that CRCSCs have higher LDs content than SDACs and CCCs, whereas NECCs exhibit the lowest LDs content. Interestingly, lipid content in colorectal cancer cells directly correlates with CD133 and Wnt pathway activity. Furthermore, CRCSCs with high LDs content exhibit higher clonogenic and tumorigenic potential than LDlow CRCSCs [100]. Similarly, ovarian cancer stem cells (OCSCs) (ALDH+/CD133+) isolated from the COV362 cell line have higher LDs content than ALDH−/CD133− cancer cells [90]. Elevated LDs content in CSCs not only provide an alternative energy source when glycolysis is blocked, but also protect fatty acids from harmful peroxidation in the stem cell niche, thus enabling stem cell proliferation [101]. Whereas inhibition of phospholipase A2 leads to reduction in LDs and triggers apoptosis in cancer cells [57].
De novo lipid biosynthesis in CSCs
A metabolic hallmark of cancer is the increase in de novo lipogenesis [102] (Fig. 1). Unlike most non-malignant cells, cancer cells are highly dependent on de novo lipogenesis to satisfy energy demands because of the limited availability of dietary lipids. CSCs may siphon glycolytic metabolic intermediates into de novo lipid biosynthesis to increase self-renewal growth [66]. Yasumoto et al. reported that both 14[C]-glucose and 14[C]-acetate incorporation into lipids is more pronounced in GSCs, indicating that de novo lipogenesis is more active in these cells than in differentiated bulk glioma cells [103]. Intriguingly, fatty acid synthase (FASN), a key lipogenic enzyme, is overexpressed in patients-derived GSCs but is dramatically decreased upon serum-induced differentiation, suggesting that enhanced de novo lipogenesis contributes to maintain the undifferentiated status of GSCs. Inhibition of lipogenesis in GSCs by pharmacologically targeting FASN with cerulenin significantly reduces stemness marker (SOX2, nestin, CD133, and FABP7) expression levels, invasiveness, and sphere formation in GSCs, whereas glial fibrillary acidic protein GFAP levels are increased. Using proteomic and metabolomic analyses, Brandi et al. demonstrated that pancreatic CSCs have higher levels of glycolysis and increased de novo lipogenesis activity compared to bulk parental cancer cells, but reduced mitochondrial OXPHOS levels. The authors showed that FASN is overexpressed and is more sensitive to inhibition by cerulenin in Panc1 CSCs than in the parental non-stem cancer cells. [91]. BCSCs (CD24− CD44+ ESA+) isolated from MCF10DCIS.com cells, which give rise to ductal carcinoma in situ, exhibit higher expression levels of lipogenic genes such as ATP citrate lyase (ACLY), acetyl CoA carboxylase 1 (ACC1), and FASN compared to non-stem cancer cells(Fig. 1). Ectopic expression of sterol regulatory element-binding protein-1 (SREBP1) gene, a master regulator of lipogenesis, upregulates downstream lipogenic genes (ACLY, ACC1, and FASN) concomitant with increased lipogenesis, growth, and mammospheres formation in MCF10A stem-like cells. Upregulation of these lipogenic genes promotes cell viability and proliferation in MCF10A stem-like cells [104]. ACC converts acetyl-CoA to malonyl-CoA during lipid biosynthesis. Inhibition of ACC by soraphen A also notably decreases the number of ALDH1+-CSCs-like cells and impairs mammosphere formation in MCF-7 cells [66].
Lipid desaturation in CSCs
Lipid unsaturation is essential in breast, colon, and prostate cancer cells [105, 106]. Administration of unsaturated fatty acids by gavage to BALB/c mice pre-inoculated with colorectal cancer cells amplify CD133(+) subpopulations, induces stemness and promotes tumor formation and metastasis [107]. Most mono-unsaturated fatty acids (MUFAs) are the catalytic product of stearoyl-CoA desaturase (SCD) (Fig. 1), a rate-limiting enzyme, that adds a cis-double bond at the delta 9 position in acyl-CoA chains [108]. It has been shown that overexpression of SCDs promotes cancer cells proliferation and inhibits cell death [105, 106, 109].
Lipid unsaturation was recently recognized to be a unique metabolic biomarker for ovarian CSCs [90]. Ovarian CSCs harbor more unsaturated lipid-containing LDs than non-stem cancer cells, implying that CSCs rely much more on unsaturated lipids than bulk tumor cells. Blocking lipid desaturase decreases ovarian CSCs marker expression and prevents tumor initiation in vivo [90], which is consistent with the pilot study that SCD1 acts as a stemness regulator in breastcancer [109]. Importantly, these observations indicate that lipid desaturase may be an ideal target for tumor prevention in cancers from various tissue origins.
There are currently several possible mechanisms underlying CSCs regulation by lipd desaturation. Lipids are essential components of cellular membranes, the fluidity of which is determined by the degree of lipid unsaturation. Membrane mechanical properties are critical in cell division, migration, and signal transduction [110]. Reducing membrane fluidity exerts an inhibitory effect on metastatic capacity and stem cell-like properties of breast cancer cells [111]. Increase in polyunsaturated fatty acids prevent lipotoxicity of saturated fatty acid (SFA) to the membrane system, which impair membrane fluidity [112]. Palmitate, a lipotoxic metabolite mainly derived from SFA, drives solid-like domain separation from the ER membrane and thus reduces membrane fluidity [113]. Decrease in palmitic acyl (C16:0)-containing glycerophospholipids promotes HCC cell proliferation and invasiveness, highlighting that excessive palmitic acid impairs HCC development [114]. Compared to non-CSCs, CSCs have lower cell membrane tension and exhibit significant shape deformation in response to stimuli. Increase in membrane tension by immersing CSCs in hypotonic medium leads to a decrease in polarized CSCs [115]. Because cell polarity is essential for asymmetric division and migration [116, 117], CSCs would benefit from increased fatty acid unsaturation in lowering membrane tension and preventing symmetric division or loss of pluripotency. It’s worthy to note that lipotoxic metabolites, including palmitate and stearate, are the preferred substrates of SCDs [108], suggesting SCDs may be crucial to maintain membrane fluidity of CSCs.
Recent studies have further deepened our understanding of how lipid desaturation interplays with oncogenic signaling pathways to generate CSCs. It has been demonstrated that NF-kB signaling activation and ALDH1A1 promotes lipid desaturation [90] (Fig. 2). Reciprocally, inhibition of desaturases with CAY 10566 and SC-26196 dramatically represses NF-kB transcriptional activity and ovarian CSCs characteristics, though the detailed mechanisms underlying NF-kB activation by SCD1 or unsaturated fatty acids still remain unclear. MUFAs produced by SCDs also amplify Wnt signaling via stabilization of β-catenin in rodent hepatic stellate cells (HSCs) and mouse liver TICs [118] (Fig. 2). Furthermore, MUFAs increase cytosolic levels of nuclear import of elav-like protein 1 (HuR), thus promoting HuR-mediated stabilization of Lrp5 and Lrp6 mRNAs [118]. The third oncogenic signaling link to lipid desaturation in CSCs generation is the Hippo/YAP signaling pathway. SCD1 activity promotes nuclear accumulation of YAP and increases transcriptional activity in lung adenocarcinoma CSCs in a Wnt-dependent manner, which is evidenced by Wnt3a rescuing YAP protein from SCD1 inhibition(Fig. 2) [119]. Co-expression of SCD1 with β-catenin and YAP/TAZ transcriptional target birc5 predicts unfavorable clinical outcomes in lung adenocarcinoma patients [119]. In addition to directly promoting stemness in CSCs, unsaturated fatty acids also stimulate mesenchymal stem cells to increase secretion of angiogenic factors such as interleukin-6, VEGF, and nitric oxide [120], which play a crucial role in angiogenesis and metastasis in human cancers.
Elevated lipolysis and extracellular lipid uptake sustain CSCs
FFAs produced by host cell lipolysis also fuel tumor growth [121]. Recently, Singh et al. demonstrated that blocking lipolysis in the digestive system of adult Drosophila melanogaster selectively induces necrotic death in normal and transformed stem cells without affecting differentiated cells [122]. Melanosphere-derived CSCs have increased lipid uptake when compared with differentiating melanosphere-derived cells [123]. Leukemic stem cells (LSCs) residing in gonadal adipose tissue (GAT), which act as a LSC niche to support LSC metabolism, trigger lipolysis to release FFAs through secretion of pro-inflammatory cytokines such as TNF-α, IL-1α, IL-1β, and CSF2. These FFAs are transported into LSCs via CD36(Fig. 1), a fatty acid transporter enriched in a sub-population of LSCs, and then reused via β-oxidation in LSC mitochondria to support LSC survival and evade chemotherapy. Loss of CD36 reduces homing of LSCs to GAT and leukemic burden in mice [124]. Enrichment of CD36 was also observed in glioma CSCs. Uptake of oxidized phospholipids such as oxLDL, a natural ligand of CD36, drives glioma CSCs proliferation but exerts no effect on differentiated glioma cells [125]. In addition to affecting proliferation of CSCs, uptake of palmitic acid via CD36 also specifically activates the metastatic potential of CD44bright oral squamous cell carcinoma (OSCC) metastasis-initiating cells [126], highlighting the central role of lipids uptake in fueling tumor metastasis.
Elevated FAO fuels CSCs
Oncogenic K-Ras mutation contributes to CSCs activation in colorectal cancer tumorigenesis, increased FAO may be involved [127]. Oncogenic K-ras (G12D) activation stimulates mitochondrial FAO to support metabolism and drive non-small cell lung cancer (NSCLC) development via up-regulating autophagy [128]. MYC-driven triple-negative breast cancer (TNBC) has an increased reliance on FAO for uncontrolled tumor growth [129]. Furthermore, mitochondrial FAO also drives triple negative breast cancer cells(TNBC) metastasis [130]. A recent study unveiled that NANOG stimulates mitochondrial FAO gene expression but represses mitochondrial OXPHOS gene expression [60] (Fig. 3). Metabolic reprogramming from OXPHOS to FAO is critical for NANOG-mediated HCC TIC generation [60]. Inhibition of FAO impairs TIC self-renewal and tumorigenicity and sensitizes TICs to sorafenib, which is a broadly used chemotherapy drug against HCC.
Fig. 3.
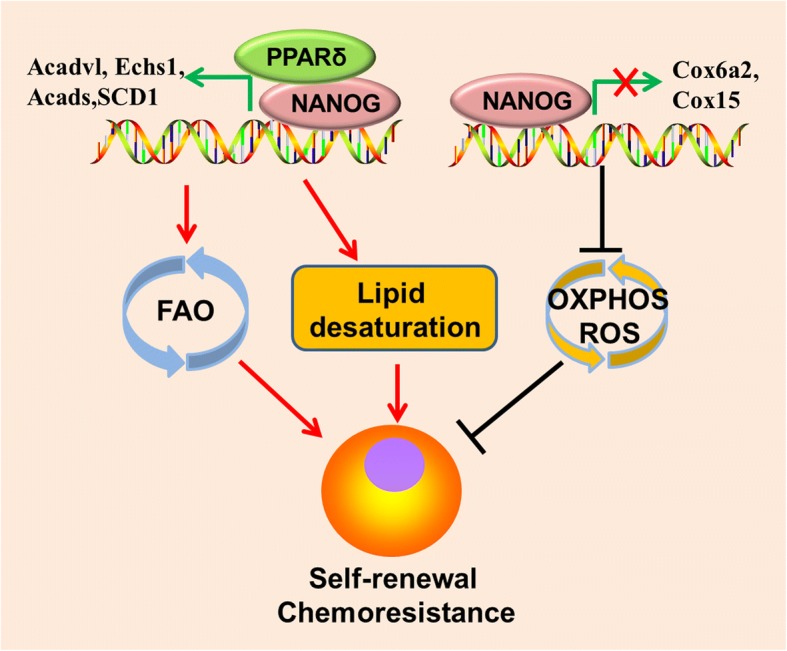
Regulation of SREBP1 and lipid metabolism by oncogenic signaling in CSCs. Oncogenic PI3K (H1047R)- and K-Ras (G12 V) activates SREBP1 and SREBP2 to support de novo lipid synthesis and cell growth. The mTOR signaling regulates SREBP1 level through both transcriptional or translational mechanisms. Activation of PI3K.AKT/mTOR signaling pathway or FGFR3 leads to stabilization of SREBP1 protein and promotes SREBP1 translocation to nucleus. Mitotic kinase Cdk1 and Plk1 physically interact with nuclear SREBP1 protein. Sequentially phosphorylation of SREBP1 by Cdk1 and Plk1 blocks binding between the ubiquitin ligase Fbw7 and SREBP1 and attenuates SREBP1 degradation. Upon EGFR signaling activation, the nuclear form of PKM2 physically interacts with SREBP1, activating SREBP target gene expression and lipid biosynthesis
Mitochondrial FAO plays an important role in satisfying energy requirements in TICs (Fig. 1). Increased FAO supports CSCs survival when glucose metabolism becomes limiting [131, 132]. Increase in FAO is critical to inflammatory signaling-mediated CSCs generation. For example, inhibition of FAO blocks BCSCs self-renewal and increases its chemo-sensitivity [89]. Activation of Src oncoprotein is also associated with CSCs generation [133]. FAO plays a crucial role in Src oncoprotein activation through autophosphorylation at Y419 in TNBC [134]. LSCs lacking CPT1A, a rate-controlling enzyme in FAO, are refractory to avocatin B, a lipid derived from avocado fruit that selectively kills AML stem cells with little effect on its normal counterpart [135], highlighting the importance of FAO in the establishment of chemo-resistance.
Mitochondrial FAO also benefits stem cells via several different mechanisms. First, FAO reduces ROS production, which is harmful to stem cells [131], that why disrupting their redox defense capability exerts therapeutic effect against CSCs [136]. Second, mitochondrial FAO is essential for pluripotency maintenance in HSCs and NSCs via controlling the asymmetric division in HSCs [137, 138]. Reduced FAO flux potentiates NSCs symmetric differentiating divisions at the expense of self-renewal [139]. Third, FAO pathway activation by peroxisome proliferator-activated receptor contributes to Tie2+ HSC expansion through induction of mitophagy [140]. In addition to maintaining the redox balance of CSCs, mitochondrial FAO is also important for epigenetic regulation of gene transcription, because acetyl coenzyme A, an intermediate product of FAO, is required for histone acetylation by the histone acetyltransferase p300 [141].
Key modulators of lipid metabolism in CSCs
NANOG
NANOG is a key transcription factor implicated in pluripotency maintenance and self-renewal in ES cells. NANOG is expressed in various human cancers and is associated with poor prognosis [142–144]. Re-activation of NANOG in CSCs is critical for stemness maintenance and self-renewal [145–147], in which it orchestrates mitochondrial metabolic reprogramming [60]. ChIP-seq revealed that NANOG binds to mitochondrial OXPHOS gene (Cox6a2 and Cox15) promoters and represses those genes transcription. In contrast, NANOG binds to FAO genes (Acadvl, Echs1, and Acads) promoters and stimulates it’s transcription, a cooperative interaction with peroxisome proliferator-activated receptor δ(PPARδ) involved(Fig. 3). Thus, overexpression of NANOG represses mitochondrial respiratory activity and ROS production, but stimulates FAO to favor HCC TIC self-renewal and chemo-resistance, whereas silencing NANOG has opposite effects [60]. Either re-expression of OXPHOS genes (Cox6a2 or Cox15) or silencing FAO genes (Echs1 and Acadvl) leads to impairment of TIC self-renewal, suggesting that reprogramming the metabolism from mitochondrial OXPHOS to FAO by NANOG is an intrinsic character of CSCs, not a conditional adaptation. Interestingly, NANOG reduces long-chain FA levels but upregulates SCD1 expression in HCC TICs [60] (Fig. 3). Inhibition of SCD by PluriSin#1, a small-molecule SCD inhibitor, diminishes NANOG-positive stem cells in induced pluripotent stem cells (iPS) and prevents tumorigenicity of iPS derivatives (iPSD) [148], suggesting that lipid desaturation is also required for NANOG-mediated CSCs generation. It is worthy to note that PluriSin#1 preferentially induces NANOG-positive stem cell apoptosis but has little effect on differentiated cardiomyocytes derived from iPS, showing a promising clinical application for cancer therapy.
SREBP1
SREBP1, a master transcriptional regulator of lipogenesis, belongs to the SREBP transcription factor family and plays an important role in fatty acid and cholesterol biosynthesis [149]. SREBP1 is required to maintain lipogenesis under lipid- and oxygen-deprived conditions. Several lipogenesis enzymes are directly regulated by SREBP1, including ACLY, ACC1, and FASN [104] (Fig. 4). SREBP1 overexpression is observed in various human cancers and promotes tumor growth [150–154]. SREBP1 is also upregulated in BCSCs, supporting its stem cell behavior [104]. Activation of SREBP1 and SREBP2 is required for oncogenic PI3K (H1047R)- and K-Ras (G12 V)-stimulated de novo lipid synthesis and breast epithelial cell growth [155] (Fig. 4). In addition to promoting lipogenesis, SREBP1 contributes to generation of mono-unsaturated fatty acids by inducing SCD1 expression [156, 157] (Fig. 4). Inhibition of SREBP1 significantly blocks spheroid growth in glioblastoma [157]. SREBP1 protein is stabilized upon sequential phosphorylation by mitotic kinase Cdk1 and Plk1 during mitosis, blocking binding between the ubiquitin ligase Fbw7 and SREBP1 and attenuating SREBP1 degradation [158–160] (Fig. 4). In addition, nuclear accumulation of mature SREBP1 is promoted by PI3-kinase/Akt/target of rapamycin (mTOR)C1 signaling [161] (Fig. 4). Activation of EGFR signaling induces nuclear translocation of pyruvate kinase M2 (PKM2) [162], a key enzyme in Warburg effect [163]. A latest study unveiled that nuclear PKM2 physically interacts with SREBP1 and stimulates lipid biosynthesis through stabilizing SREBP-1 protein(Fig. 4), providing further evidence to show the crosstalk between glycolysis and fatty acids metabolism [164].
Fig. 4.
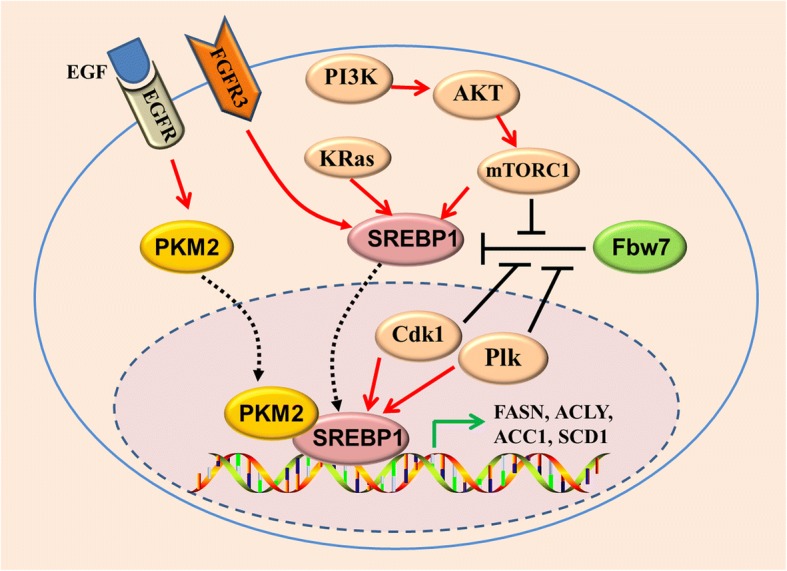
Interaction between oncogenic signaling and lipid desaturation in CSCs. Oncogenic activation of K-RAS, PI3K/AKT/mTOR signaling stimulates de novo lipogenesis via upregulation of SREBP1. Increase of SCD1 expression and lipid desaturation by NANOG or oncogenic signaling in CSCs or TICs reciprocally amplify NF-κB, Wnt/β-catenin, and Yap activation. Activation of JAK/STAT3 promotes CPT1B expression and activates the FAO pathway, which in turn contributes to Src oncoprotein activation. SREBP1, sterol regulatory element-binding protein-1
MYC
MYC is overexpressed in various human cancers [165], and its potential to reprogram the cellular metabolism in cancer is well-recognized [166–168]. Importantly, the role of MYC in metabolic reprogramming has been confirmed in vivo. Though MYC is not necessary for hepatoblastoma development, depletion of MYC delays tumor progression through reducing fatty acid transporter CD36 expression, with a concomitant decrease in LDs accumulation and FAO levels [169]. Inhibition of FAO significantly delays constitutively active MYC-driven lymphoma development in a transgenic model [170, 171], whereas increase in FAO facilitates the tricarboxylic acid (TCA) cycle and ATP production in MYC-overexpressing TNBC [172]. It is worthy to note that inhibition of FAO by etomoxir in a MYChigh TNBC patient-derived xenograft (PDX) reduces the energy metabolism and inhibits tumorigenesis but has little effect on a MYClow TNBC PDX model, suggesting a therapeutic potential in MYC-overexpressing TNBC tumors [129].
SCD
SCD1 and SCD2 are two major isoforms of SCDs. Deletion of SCD1, the most abundant desaturase, decreases cardiac FFA and ceramide content in mice by reducing lipogenesis and activating lipolysis [173]. SCD2 expression is up-regulated to compensate SCD1 deficiency in mice liver [174]. Increased SCD1 expression has been observed in CSCs from ovarian, lung, breast cancer, and HCC [90, 109, 118, 175, 176]. In early stages of lung ADC, SCD1 is co-expressed with CSCs markers (CD133, CD24, CD44, and SOX2) [177]. High expression levels of SCD1 are tightly associated with disease progression and unfavorable clinical outcomes in lung cancer [177], HCC [118, 176], and breast cancer [105]. SCD1 expression is also negatively correlated with tumor differentiation in human HCC [178].
SCD1 is pivotal for CSCs/TIC generation and stemness maintenance [90, 118, 119, 175]. For example, silencing SCD1 in MCF10A cells significantly reduces mammosphere formation and the number of CD44+/CD24− cells [109]. Genetic disruption of either SCD1 or SCD2 similarly inhibits TIC self-renewal and prevents experimental HCC formation induced by chemical carcinogens in mice [118]. However, it’s remain elusive whether SCD1 overexpression enhances stemness in non-stem cancer cells. Surprisingly, SCD1 is decreased in LSCs and plays a tumor-suppressive role in chronic myeloid leukemia [179], indicating that SCD1 function is context-dependent.
SCD1 expression is regulated by distinct oncogenic signaling pathways. SCD1 levels are transcriptionally and translationally controlled by mammalian mTOR signaling [180] (Fig. 2). In rodent HSCs and TICs, SCD1 expression is induced by Wnt-β-catenin signaling and reciprocally stabilizes the β-catenin protein [118] (Fig. 2). Li Junjie et al. demonstrated that NF-kB p65 binds to the SCD1 gene promoter (− 215 to − 206 and + 79 to + 88 bp) and induces its expression in ovarian cancer [90] (Fig. 2). Furthermore, SCD1 is a target of NANOG and is required for NANOG-mediated TIC generation [60] (Fig. 2). In breast cancer cells, SCD1 expression is induced by 17β-estradiol [181]. SCD1 expression is also induced by cancer-associated fibroblasts-released factors and promotes breast cancer cells migration, thus linking tumor microenvironment to metabolic reprogramming of CSCs [182]. Fibroblast growth factor receptor 3 (FGFR3) also stimulates SCD1 expression to accelerate tumor growth via activating SREBP1 in bladder cancers [183] (Fig. 4).
Genetic or pharmacological inhibition of SCD1 exerts a powerful anti-CSCs effect in various cancer types, including lung [175, 177], colon CSCs [184], ovarian [90], breast [109], and liver cancers [118, 178]. Silencing SCD1 or inhibiting its activity with betulinic acid (BetA) leads to rapid cell death in colon CSCs [184]. ROS generation may largely contribute to apoptosis after SCD1 inhibition [185]. Furtehr more, SCD1 activity is required for autophagosome formation [186]. SCD1 inhibition triggers cell death of pancreatic β-cells due to impairment of autophagy [187]. Nevertheless, the role of autophagy in SCD1 inhibition-induced cell death is controversial because pharmacological inhibition of SCD1 induces autophagic cell death via stimulating AMPK signaling [188]. SCD1 inhibition also induces liver TIC differentiation via the ER stress-induced unfolded protein response [176]. Importantly, inhibition of SCD1 overcomes drug resistance of lung adenocarcinoma CSCs to cisplatin [177] and liver TICs to chemotherapeutic drugs [176, 178], making it a desirable approach for novel combined therapeutic strategies.
FASN and ACSVL3
FASN is a key enzyme in lipogenesis. It is highly expressed in patient-derived GSCs but markedly decreased after differentiation. Treatment of GSCs with cerulenin, a pharmacological inhibitor of FASN, leads to reduction of de novo lipogenesis and loss of stemness [103]. Overexpression of very long-chain acyl-CoA synthetase 3 (ACSVL3) has been demonstrated in lung cancer and glioma [189, 190]. Activation of both oncogenic receptor tyrosine kinases (RTK) c-Met and EGFR contributes to increased ACSVL3 levels in glioma cells, whereas silencing ACSVL3 leads to de-activation of Akt signaling [189]. Recently, Sun et al. demonstrated that ACSVL3 is implicated in GSCs maintenance and tumor initiation capacity. In addition, knockdown of ACSVL3 in neurosphere cells impairs its self-renewal and induces differentiation [191].
CD36
CD36 is a scavenging receptor that is enriched in CSCs [124, 125] (Fig. 1). It has been shown that CD36, ITGA6, and CD133 are co-expressed in glioblastoma, and CD36 may be used to functionally distinguish CSCs from non-CSCs [125]. Otherwise, CD36 is highly expressed in metastasis-initiating cells (CD44bright dye+) of OSCC cells compared to its CD44bright dye− counterpart. CD36(−) cells lose the ability to form a single lymph node metastasis, whereas CD36(+) cells develop more lymph node metastases than their parental cells. However, both CD36(+) and CD36(−) cells efficiently form oral lesions and primary tumors when orally inoculated into NSG mice, highlighting it’s distinct role of CD36(+) cells in metastasis. CD36(+)CD44bright cells have higher expression levels of key enzymes involved in FAO (ACADVL, ACADM, and HADHA). Blocking lipid uptake with anti-CD36 antibodies in metastasis-initiating cells inoculated in mice successfully prevents metastasis initiation but not primary tumor formation [126]. In addition, loss of CD36 significantly sensitizes LSCs to chemotherapy and impairs tumorigenicity in mice [124]. However, challenging the accepted notion that chemo-resistant cells are LSCs, Farge et al. demonstrated that cytarabine (AraC)-resistant cells are neither LSCs nor quiescent cells, although AraC-resistant cells exhibit high levels of CD36 expression and FAO activity [192].
CPT1A and CPT1B
Mitochondrial FAO is initiated with the transfer of long-chain fatty acids from the cytosol into the mitochondrial matrix by CPT1 and CPT2. CPT1 (also named CPT1A) is a rate-limiting enzyme in FAO and is located in the outer mitochondrial membrane (Fig. 1), whereas CPT2 is located in the inner mitochondrial membrane. CPT1A is overexpressed in prostate cancer and is associated with a high tumor grade [193, 194]. High expression levels of CPT1A predict unfavorable clinical outcomes in AML [195] and ovarian cancer [196]. Genetic or pharmacological inhibition of CPT1A exerts anti-tumor activity in prostate cancer [193], melanoma [197], breast [198], and ovarian cancer [196]. CPT1A is required for stem cell maintenance in neural stem/progenitor cells (NSPCs) [138]. CPT1A-dependent FAO flux is high in quiescent NSPCs but decreases in proliferating NSPCs. Reduced FAO flux triggers NSPC differentiation and loss of pluripotency [139]. CPT1B is one of the three isoforms of CPT1. Recently, Wang et al. demonstrated the role of JAK/STAT3 signaling in the regulation of BCSCs and cancer chemoresistance through promoting CPT1B expression and FAO in BCSCs. Blocking JAK/STAT3 signaling inhibits the self-renewal of BCSCs and re-sensitizes them to chemotherapy [89].
Targeting lipid metabolism as novel therapeutic strategies against CSCs
CSCs are resistant to most traditional treatments. However, their dependency on lipid metabolism for proliferation and survival offers an Achilles heel for the elimination of these cells. Targeted clearance of CSCs could be achieved by intervening in different aspects of fatty acid metabolism such as lipogenesis, lipid uptake, lipid desaturation, and FAO. Due to the high costs and risk to discover and develop novel therapeutic agents, therapeutic strategies of drug repositioning for difficult-to-cure diseases treatment gain increasing attentions [40]. For instance, terfenadine, a “conventional” agents used to auto-immune disorders such as allergic dermatitis, has been demonstrated to reduce VEGF secretion from mast cells resided in the hypoxic microenvironment, and exerts great potential to kill melanoma cells via ROS-mediated apoptosis and autophagy [199, 200].
Targeting lipogenesis
FASN is the most targetable among lipogenesis genes. Several FASN inhibitors display anti-tumor activity in preclinical cancer models (Table 1). Most importantly, some inhibitors show selective activity against CSCs rather than bulk tumor cells. Strikingly, Orlistat, a FDA-approved anti-obesity drug targeting FASN, exerts a potent anti-tumor activity in various cancers [201, 202]. Remarkably, inhibition of FASN by Orlistat in EGFR mutated NSCLC suppresses tumor growth in vitro and in vivo through reducing EGFR palmitoylation but inducing mutant EGFR ubiquitination and subsequent proteasomal degradation [203].
Table 1.
Inhibitors of lipid enzymes involved in CSCS
| Metabolism type | Drug | Targeting enzyme | Cancer type | Stage |
|---|---|---|---|---|
| Lipogenesis | Resveratrol | FASN | Breast cancer CSCs [104, 209]. Glioblastoma CSCs [210, 231]. Pancreatic CSCs [232] |
Clinical Trial |
| Cerulenin | FASN | Glioma CSCs [103], Pancreatic CSCs [91] | Pre-clinical | |
| Orlistat | FASN | NSCLC [203] | FDA-approved anti-obesity drug | |
| Lipid uptake | CD36 antibody | CD36 | OSCC [126] | Pre-clinical |
| MTN | CD36 | Glioblastoma CSCs [125] | Pre-clinical | |
| FAO | Etomoxir | CPT1A | MYC-overexpressing TNBC [129], leukemia [219, 220] | Pre-clinical |
| ST1326 | CPT1A | Lymphoma [170], acute myeloid leukemia [221] | Pre-clinical | |
| Lipid desaturation | SSI-4 | SCD1 | Liver CSCs [176], | Pre-clinical |
| BetA | SCD1 | CRC [184] | Pre-clinical | |
| PluriSin#1 | SCD1 | Teratomas [148] | ||
| MF-438 | SCD1 | Lung cancer CSCs [177] | Pre-clinical | |
| A939572 | SCD1 | CRC [185], clear cell renal cell carcinoma [233] | Pre-clinical | |
| Cay10566 | SCD1 | Breast Carcinoma [216] | Pre-clinical | |
| T-3764518 | SCD1 | CRC [234] | Pre-clinical |
OSCC oral squamous cell carcinomas, CRC colorectal cancer, TNBC triple-negative breast cancer, MTN 2-methylthio-1,4-naphthoquinone, FAO fatty acid β-oxidation
Resveratrol
Resveratrol is a natural phytochemical compound extracted from grapes, red wine, berries, and peanuts(Table 1). Pre-clinical trials revealed that resveratrol exerts chemo-therapeutic and chemo-preventive effects against human cancers [204]. Further more, recent study demonstrated that resveratrol inhibits EMT and overcomes doxorubicin resistance in gastric cancer through modulating PTEN/Akt signaling pathway [205].. It has been shown that resveratrol at low dosage stimulates KEAP1/Nrf2 pathway and protects cells against oxidative agents, which may be the underlying mechanisms for it’s cancer chemoprevention function. However, high concentrations of resveratrol promotes ROS production leading to cell death [206–208]. Resveratrol suppresses lipogenesis and induces apoptosis in BCSCs by suppressing FASN expression [104, 209], making it attractive for clinical utilization. Resveratrol treatment inhibits GSCs proliferation at low doses and induces necrosis at higher doses but has no effect on normal NSCs behavior [210]. SIRT2 activation is required for resveratrol-mediated GSCs proliferation arrest but not for necrosis induced by high doses of resveratrol. Several clinical trials are currently underway [NCT00721877, NCT00920803, NCT00433576, and NCT00578396]. Pterostilbene, a dimethylated derivative of resveratrol with a higher bioavailability and is more lipophilic, exerts more potent suppressive activity against CSCs and cancer metastasis [211, 212].
Cerulenin
Cerulenin, a fungal metabolite, is another natural pharmacological inhibitor of FASN (Table 1). Cerulenin preferentially inhibits pancreatic CSCs proliferation compared to its parental cell [91]. It also inhibits glioma CSCs proliferation, migration and induces CSCs differentiation to glial cells [103]. The anti-tumor activity of cerulenin is enhanced by the combined use of oxaliplatin in human colon cancer cells [213]. Cerulenin also exerts an inhibitory effect on protein palmitoylation, thus affecting CD36 membrane trafficking [214].
Targeting lipid desaturation
It is noteworthy that the SCD requirement for TIC generation was recently confirmed in a genetic mouse model [118]. High dependence on unsaturated fatty acids makes SCD a promising target to eradicate CSCs [215]. Several kinds of SCD1 inhibitors exhibit anti-tumor activity in pre-clinical cancer models (Table 1). SSI-4 is a novel SCD1 inhibitor that reverts sorafenib resistance in liver CSCs. SSI-4 display anti-tumor activity against liver CSCs without serious side effects in pre-clinical animal models [176]. Importantly, SSI-4 in combination with sorafenib show a maximal suppressive effect against tumorigenesis in a sorafenib-resistant PDTX model. Notably, the minimum dose of SSI-4 that has anti-tumor activity is approximately 10 mg/kg, a much lower concentration than that of other inhibitors such as A939572. MF-438 is another SCD1 inhibitor that reverts cisplatin resistance in lung CSCs [176]. A939572 and Cay10566 are also two widely used SCD1 inhibitors [173, 182, 185, 216]. A series of novel and potent SCD inhibitors are currently being developed [217]. Table 1 summarizes the potent SCD1 inhibitors used in pre-clinical stage.
Targeting FAO
The dependency on FAO of CSCs makes it reasonable to target these cells using FAO inhibitors. Etomoxir is a specific inhibitor of mitochondrial CPT1A [218] (Table 1). Camarda et al. used a clinically relevant PDX model and found that etomoxir treatment markedly decreases ATP production and tumor growth in a MYChigh TNBC PDX model [129]. Etomoxir inhibits FAO in leukemia CSCs, thus suppressing cell proliferation and sensitizing human leukemia cells to apoptosis [124, 219, 220]. ST1326 is a CPT1A inhibitor that inhibits FAO and exerts cytotoxic activity against leukemia cell lines but not against normal CD34+ bone marrow cells [221] (Table 1). ST1326 treatment also prevents MYC-driven lymphomagenesis in a Eμ-myc transgenic mice model [170].
Targeting lipid uptake
The scavenger receptor CD36 is enriched in CSCs and is responsible for extracellular lipid uptake [124–126]. Increased lipid uptake is observed in melanoma, glioblastoma, and leukemia CSCs, offering a promising avenue to develop novel therapeutic strategies [99, 123, 124]. Inhibition of CD36 by 2-methylthio-1,4-naphthoquinone treatment decreases self-renewal and induces apoptosis of CSCs in glioblastoma [125] (Table 1). Alternatively, CD36-neutralizing antibodies block extracellular lipid uptake of CSCs in OSCC and markedly inhibits tumor growth without side effects [126] (Table 1). Dependence on extracellular lipids also makes it reasonable to develop lipid nanoparticles as a Trojan horse to deliver drugs into CSCs. Many such strategies are being developed [222, 223]. For example, delivery of lipid nanoparticles containing miR-200c combined with paclitaxel (PTX) into breast CSCs exhibits profound anti-tumor activity [224].
Conclusions
Recent advances in metabolomics have deepened our understanding of the contribution of metabolic reprogramming to tumorigenesis, which is now a well-recognized hallmark of cancer [225]. Alterations in lipid metabolism such as increase in fatty acid uptake, de novo lipogenesis, formation of LDs, FAO, and lipid desaturation are intensively involved in CSCs generation and stemness maintenance. Fatty acid metabolism not only satisfies the energy demands and biomass production of CSCs, but also contributes to the activation of several important oncogenic signaling pathways, including Wnt/β-catenin and Hippo/YAP signaling. Targeting key players of fatty acids metabolism shows promising to anti-CSCs and tumor prevention effects.
Although targeting the cell metabolism provides promising opportunities for eliminating CSCs, we have to face the dilemma of heterogeneity and metabolic plasticity of these cells [226–229]. CSCs and tumor cells may adapt its metabolic profile based on nutrients availability. For example, when the Warburg effect is reversed with cetuximab, HNSCC cells express high levels of ACC, which rewires cancer metabolism from glycolysis to lipogenesis to support energy demands and proliferation [230]. Although increased lipogenesis has been well documented in CSCs from various cancer types, most chemical compounds targeting FASN does not show a therapeutic efficacy in pre-clinical cancer models and only one FASN inhibitor has entered clinical trials. Owing to the metabolic flexibility of CSCs, it is difficult to effectively eliminate these cells by targeting a single metabolic pathway. A great challenge is to develop strategies to synergistically target multiple metabolic pathways in CSCs. An additional challenge comes from the metabolic similarities between CSCs and normal stem cells. For example, mitochondrial FAO is essential for NANOG-driven HCC TIC generation [60], but also contributes to expansion of normal HSCs and NSCs [137, 138]. Thus, the side effects of FAO inhibitors on normal HSCs and NSCs will have to be considered.
Acknowledgements
We apologize to those researchers whose work in the field could not be cited in the present review due to space constrains.
Funding
This study was supported in part by Grants from The National Natural Science Foundation of China (81372304, 81572667, 81772902), the National “111” Project (Project #111–2-12), The Natural Science Foundation of Hunan Province, China (2018JJ1040).
Abbreviations
- ACADM
Acyl-CoA dehydrogenase medium chain
- ALDH1
Aldehyde dehydrogenase isoform 1
- CoA
Acetyl coenzyme A
- CSCs
Cancer stem cells
- EGFR
Epidermal growth factor receptor
- EMT
Epithelial-mesenchymal transition
- FAO
Fatty acid β-oxidation
- HADHA
Hydroxyacyl-CoA dehydrogenase trifunctional multienzyme complex subunit alpha
- HCC
Hepatocellular carcinoma
- HIF1
Hypoxia-inducible factor 1
- HNSCC
Head and neck squamous cell carcinomas
- TBNC
Triple negative breast cancer
- TICs
Tumor initiating cells
- VEGF
Vascular endothelial growth factor
Authors’ contributions
MY, and BX wrote the manuscript. JJL, SNC, JC,YYB, QP, YZ, ZYZ, SPP, XLL, WX, and GYL revised and corrected the manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable
Consent for publication
All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Mei Yi, Email: plumyi@163.com.
Junjun Li, Email: lijunjun@csu.edu.cn.
Shengnan Chen, Email: chenshengnan@csu.edu.cn.
Jing Cai, Email: caijing@csu.edu.cn.
Yuanyuan Ban, Email: banyuanyuan@csu.edu.cn.
Qian Peng, Email: pengqian@csu.edu.cn.
Ying Zhou, Email: zhouying01@csu.edu.cn.
Zhaoyang Zeng, Email: zengzhaoyang@csu.edu.cn.
Shuping Peng, Email: shuping@csu.edu.cn.
Xiaoling Li, Email: lixiaoling@csu.edu.cn.
Wei Xiong, Email: xiongwei@csu.edu.cn.
Guiyuan Li, Email: lgy@csu.edu.cn.
Bo Xiang, Phone: 86-731-8386-7186, Email: xiangbolin@csu.edu.cn.
References
- 1.Carnero A, Garcia-Mayea Y, Mir C, Lorente J, Rubio IT, ME LL. The cancer stem-cell signaling network and resistance to therapy. Cancer Treat Rev. 2016;49:25–36. doi: 10.1016/j.ctrv.2016.07.001. [DOI] [PubMed] [Google Scholar]
- 2.Abbaszadegan MR, Bagheri V, Razavi MS, Momtazi AA, Sahebkar A, Gholamin M. Isolation, identification, and characterization of cancer stem cells: a review. J Cell Physiol. 2017;232(8):2008–2018. doi: 10.1002/jcp.25759. [DOI] [PubMed] [Google Scholar]
- 3.Pattabiraman DR, Weinberg RA. Tackling the cancer stem cells - what challenges do they pose? Nat Rev Drug Discov. 2014;13(7):497–512. doi: 10.1038/nrd4253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Guen VJ, Chavarria TE, Kroger C, Ye X, Weinberg RA, Lees JA. EMT programs promote basal mammary stem cell and tumor-initiating cell stemness by inducing primary ciliogenesis and hedgehog signaling. Proc Natl Acad Sci U S A. 2017;114(49):E10532–E10539. doi: 10.1073/pnas.1711534114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Chen T, You Y, Jiang H, Wang ZZ. Epithelial-mesenchymal transition (EMT): a biological process in the development, stem cell differentiation, and tumorigenesis. J Cell Physiol. 2017;232(12):3261–3272. doi: 10.1002/jcp.25797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Yakisich JS, Azad N, Kaushik V, Iyer AKV. Cancer cell plasticity: rapid reversal of Chemosensitivity and expression of Stemness markers in lung and breast Cancer Tumorspheres. J Cell Physiol. 2017;232(9):2280–2286. doi: 10.1002/jcp.25725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Mladinich M, Ruan D, Chan CH. Tackling Cancer stem cells via inhibition of EMT transcription factors. Stem Cells Int. 2016;2016:5285892. doi: 10.1155/2016/5285892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wang W, Yi M, Zhang R, Li J, Chen S, Cai J, Zeng Z, Li X, Xiong W, Wang L, et al. Vimentin is a crucial target for anti-metastasis therapy of nasopharyngeal carcinoma. Mol Cell Biochem. 2018;438(1–2):47–57. doi: 10.1007/s11010-017-3112-z. [DOI] [PubMed] [Google Scholar]
- 9.Peiris-Pages M, Martinez-Outschoorn UE, Pestell RG, Sotgia F, Lisanti MP. Cancer stem cell metabolism. Breast cancer research : BCR. 2016;18(1):55. doi: 10.1186/s13058-016-0712-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Melzer C, von der Ohe J, Lehnert H, Ungefroren H, Hass R. Cancer stem cell niche models and contribution by mesenchymal stroma/stem cells. Mol Cancer. 2017;16(1):28. doi: 10.1186/s12943-017-0595-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kelly PN, Dakic A, Adams JM, Nutt SL, Strasser A. Tumor growth need not be driven by rare cancer stem cells. Science. 2007;317(5836):337. doi: 10.1126/science.1142596. [DOI] [PubMed] [Google Scholar]
- 12.Chaffer CL, Brueckmann I, Scheel C, Kaestli AJ, Wiggins PA, Rodrigues LO, Brooks M, Reinhardt F, Su Y, Polyak K, et al. Normal and neoplastic nonstem cells can spontaneously convert to a stem-like state. Proc Natl Acad Sci U S A. 2011;108(19):7950–7955. doi: 10.1073/pnas.1102454108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Quintana E, Shackleton M, Foster HR, Fullen DR, Sabel MS, Johnson TM, Morrison SJ. Phenotypic heterogeneity among tumorigenic melanoma cells from patients that is reversible and not hierarchically organized. Cancer Cell. 2010;18(5):510–523. doi: 10.1016/j.ccr.2010.10.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Quintana E, Shackleton M, Sabel MS, Fullen DR, Johnson TM, Morrison SJ. Efficient tumour formation by single human melanoma cells. Nature. 2008;456(7222):593–598. doi: 10.1038/nature07567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lapidot T, Sirard C, Vormoor J, Murdoch B, Hoang T, Caceres-Cortes J, Minden M, Paterson B, Caligiuri MA, Dick JE. A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature. 1994;367(6464):645–648. doi: 10.1038/367645a0. [DOI] [PubMed] [Google Scholar]
- 16.Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF. Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci U S A. 2003;100(7):3983–3988. doi: 10.1073/pnas.0530291100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Singh SK, Hawkins C, Clarke ID, Squire JA, Bayani J, Hide T, Henkelman RM, Cusimano MD, Dirks PB. Identification of human brain tumour initiating cells. Nature. 2004;432(7015):396–401. doi: 10.1038/nature03128. [DOI] [PubMed] [Google Scholar]
- 18.Dalerba P, Dylla SJ, Park IK, Liu R, Wang X, Cho RW, Hoey T, Gurney A, Huang EH, Simeone DM, et al. Phenotypic characterization of human colorectal cancer stem cells. Proc Natl Acad Sci U S A. 2007;104(24):10158–10163. doi: 10.1073/pnas.0703478104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.O'Brien CA, Pollett A, Gallinger S, Dick JE. A human colon cancer cell capable of initiating tumour growth in immunodeficient mice. Nature. 2007;445(7123):106–110. doi: 10.1038/nature05372. [DOI] [PubMed] [Google Scholar]
- 20.Ricci-Vitiani L, Lombardi DG, Pilozzi E, Biffoni M, Todaro M, Peschle C, De Maria R. Identification and expansion of human colon-cancer-initiating cells. Nature. 2007;445(7123):111–115. doi: 10.1038/nature05384. [DOI] [PubMed] [Google Scholar]
- 21.Li C, Heidt DG, Dalerba P, Burant CF, Zhang L, Adsay V, Wicha M, Clarke MF, Simeone DM. Identification of pancreatic cancer stem cells. Cancer Res. 2007;67(3):1030–1037. doi: 10.1158/0008-5472.CAN-06-2030. [DOI] [PubMed] [Google Scholar]
- 22.Li C, Lee CJ, Simeone DM. Identification of human pancreatic cancer stem cells. Methods Mol Biol. 2009;568:161–173. doi: 10.1007/978-1-59745-280-9_10. [DOI] [PubMed] [Google Scholar]
- 23.Zhu J, He J, Liu Y, Simeone DM, Lubman DM. Identification of glycoprotein markers for pancreatic cancer CD24+CD44+ stem-like cells using nano-LC-MS/MS and tissue microarray. J Proteome Res. 2012;11(4):2272–2281. doi: 10.1021/pr201059g. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Li XF, Chen C, Xiang DM, Qu L, Sun W, Lu XY, Zhou TF, Chen SZ, Ning BF, Cheng Z, et al. Chronic inflammation-elicited liver progenitor cell conversion to liver cancer stem cell with clinical significance. Hepatology. 2017;66(6):1934–1951. doi: 10.1002/hep.29372. [DOI] [PubMed] [Google Scholar]
- 25.Shukla S, Khan S, Sinha S, Meeran SM. Lung cancer stem cells: an epigenetic perspective. Curr Cancer Drug Targets. 2018;18(1):16–31. [DOI] [PubMed]
- 26.Zhang S, Balch C, Chan MW, Lai HC, Matei D, Schilder JM, Yan PS, Huang TH, Nephew KP. Identification and characterization of ovarian cancer-initiating cells from primary human tumors. Cancer Res. 2008;68(11):4311–4320. doi: 10.1158/0008-5472.CAN-08-0364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Curley MD, Therrien VA, Cummings CL, Sergent PA, Koulouris CR, Friel AM, Roberts DJ, Seiden MV, Scadden DT, Rueda BR, et al. CD133 expression defines a tumor initiating cell population in primary human ovarian cancer. Stem Cells. 2009;27(12):2875–2883. doi: 10.1002/stem.236. [DOI] [PubMed] [Google Scholar]
- 28.Charafe-Jauffret E, Ginestier C, Bertucci F, Cabaud O, Wicinski J, Finetti P, Josselin E, Adelaide J, Nguyen TT, Monville F, et al. ALDH1-positive cancer stem cells predict engraftment of primary breast tumors and are governed by a common stem cell program. Cancer Res. 2013;73(24):7290–7300. doi: 10.1158/0008-5472.CAN-12-4704. [DOI] [PubMed] [Google Scholar]
- 29.Anido J, Saez-Borderias A, Gonzalez-Junca A, Rodon L, Folch G, Carmona MA, Prieto-Sanchez RM, Barba I, Martinez-Saez E, Prudkin L, et al. TGF-beta receptor inhibitors target the CD44(high)/Id1(high) glioma-initiating cell population in human glioblastoma. Cancer Cell. 2010;18(6):655–668. doi: 10.1016/j.ccr.2010.10.023. [DOI] [PubMed] [Google Scholar]
- 30.Yae T, Tsuchihashi K, Ishimoto T, Motohara T, Yoshikawa M, Yoshida GJ, Wada T, Masuko T, Mogushi K, Tanaka H, et al. Alternative splicing of CD44 mRNA by ESRP1 enhances lung colonization of metastatic cancer cell. Nat Commun. 2012;3:883. doi: 10.1038/ncomms1892. [DOI] [PubMed] [Google Scholar]
- 31.Yoshida GJ, Saya H. Inversed relationship between CD44 variant and c-Myc due to oxidative stress-induced canonical Wnt activation. Biochem Biophys Res Commun. 2014;443(2):622–627. doi: 10.1016/j.bbrc.2013.12.016. [DOI] [PubMed] [Google Scholar]
- 32.Wang W, Yi M, Chen S, Li J, Zhang H, Xiong W, Li G, Li X, Xiang B. NOR1 suppresses Cancer stem-like cells properties of tumor cells via the inhibition of the AKT-GSK-3beta-Wnt/beta-catenin-ALDH1A1 signal circuit. J Cell Physiol. 2017;232(10):2829–2840. doi: 10.1002/jcp.25706. [DOI] [PubMed] [Google Scholar]
- 33.Yi M, Yang J, Li W, Li X, Xiong W, McCarthy JB, Li G, Xiang B. The NOR1/OSCP1 proteins in cancer: from epigenetic silencing to functional characterization of a novel tumor suppressor. J Cancer. 2017;8(4):626–635. doi: 10.7150/jca.17579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Yoshida GJ. Metabolic reprogramming: the emerging concept and associated therapeutic strategies. Journal of experimental & clinical cancer research : CR. 2015;34:111. doi: 10.1186/s13046-015-0221-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Viale A, Pettazzoni P, Lyssiotis CA, Ying H, Sanchez N, Marchesini M, Carugo A, Green T, Seth S, Giuliani V, et al. Oncogene ablation-resistant pancreatic cancer cells depend on mitochondrial function. Nature. 2014;514(7524):628–632. doi: 10.1038/nature13611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Choi AM, Ryter SW, Levine B. Autophagy in human health and disease. N Engl J Med. 2013;368(7):651–662. doi: 10.1056/NEJMra1205406. [DOI] [PubMed] [Google Scholar]
- 37.Chen S, Zheng P, Wang W, Yi M, Chen P, Cai J, Li J, Peng Q, Ban Y, Zhou Y, et al. Abberent expression of NOR1 protein in tumor associated macrophages contributes to the development of DEN-induced hepatocellular carcinoma. J Cell Physiol. 2018;233(6):5002–5013. doi: 10.1002/jcp.26349. [DOI] [PubMed] [Google Scholar]
- 38.Yi M, Cai J, Li J, Chen S, Zeng Z, Peng Q, Ban Y, Zhou Y, Li X, Xiong W, et al. Rediscovery of NF-κB signaling in nasopharyngeal carcinoma: How genetic defects of NF-κB pathway interplay with EBV in driving oncogenesis? J Cell Physiol. 2018;233(8):5537–49. [DOI] [PubMed]
- 39.White E. The role for autophagy in cancer. J Clin Invest. 2015;125(1):42–46. doi: 10.1172/JCI73941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Yoshida GJ. Therapeutic strategies of drug repositioning targeting autophagy to induce cancer cell death: from pathophysiology to treatment. J Hematol Oncol. 2017;10(1):67. doi: 10.1186/s13045-017-0436-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Galluzzi L, Pietrocola F, Bravo-San Pedro JM, Amaravadi RK, Baehrecke EH, Cecconi F, Codogno P, Debnath J, Gewirtz DA, Karantza V, et al. Autophagy in malignant transformation and cancer progression. EMBO J. 2015;34(7):856–880. doi: 10.15252/embj.201490784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Yan Y, Xu Z, Dai S, Qian L, Sun L, Gong Z. Targeting autophagy to sensitive glioma to temozolomide treatment. Journal of experimental & clinical cancer research : CR. 2016;35:23. doi: 10.1186/s13046-016-0303-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Boya P, Codogno P, Rodriguez-Muela N. Autophagy in stem cells: repair, remodelling and metabolic reprogramming. Development. 2018;145(4) [DOI] [PubMed]
- 44.Yang MC, Wang HC, Hou YC, Tung HL, Chiu TJ, Shan YS. Blockade of autophagy reduces pancreatic cancer stem cell activity and potentiates the tumoricidal effect of gemcitabine. Mol Cancer. 2015;14:179. doi: 10.1186/s12943-015-0449-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Gong C, Bauvy C, Tonelli G, Yue W, Delomenie C, Nicolas V, Zhu Y, Domergue V, Marin-Esteban V, Tharinger H, et al. Beclin 1 and autophagy are required for the tumorigenicity of breast cancer stem-like/progenitor cells. Oncogene. 2013;32(18):2261–2272. doi: 10.1038/onc.2012.252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Yue W, Hamai A, Tonelli G, Bauvy C, Nicolas V, Tharinger H, Codogno P, Mehrpour M. Inhibition of the autophagic flux by salinomycin in breast cancer stem-like/progenitor cells interferes with their maintenance. Autophagy. 2013;9(5):714–729. doi: 10.4161/auto.23997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Dixon SJ, Lemberg KM, Lamprecht MR, Skouta R, Zaitsev EM, Gleason CE, Patel DN, Bauer AJ, Cantley AM, Yang WS, et al. Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell. 2012;149(5):1060–1072. doi: 10.1016/j.cell.2012.03.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Gao M, Monian P, Pan Q, Zhang W, Xiang J, Jiang X. Ferroptosis is an autophagic cell death process. Cell Res. 2016;26(9):1021–1032. doi: 10.1038/cr.2016.95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Mai TT, Hamai A, Hienzsch A, Caneque T, Muller S, Wicinski J, Cabaud O, Leroy C, David A, Acevedo V, et al. Salinomycin kills cancer stem cells by sequestering iron in lysosomes. Nat Chem. 2017;9(10):1025–1033. doi: 10.1038/nchem.2778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Lewerenz J, Hewett SJ, Huang Y, Lambros M, Gout PW, Kalivas PW, Massie A, Smolders I, Methner A, Pergande M, et al. The cystine/glutamate antiporter system x(c)(−) in health and disease: from molecular mechanisms to novel therapeutic opportunities. Antioxid Redox Signal. 2013;18(5):522–555. doi: 10.1089/ars.2011.4391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lo M, Ling V, Wang YZ, Gout PW. The xc- cystine/glutamate antiporter: a mediator of pancreatic cancer growth with a role in drug resistance. Br J Cancer. 2008;99(3):464–472. doi: 10.1038/sj.bjc.6604485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Gout PW, Kang YJ, Buckley DJ, Bruchovsky N, Buckley AR. Increased cystine uptake capability associated with malignant progression of Nb2 lymphoma cells. Leukemia. 1997;11(8):1329–1337. doi: 10.1038/sj.leu.2400739. [DOI] [PubMed] [Google Scholar]
- 53.Savaskan NE, Heckel A, Hahnen E, Engelhorn T, Doerfler A, Ganslandt O, Nimsky C, Buchfelder M, Eyupoglu IY. Small interfering RNA-mediated xCT silencing in gliomas inhibits neurodegeneration and alleviates brain edema. Nat Med. 2008;14(6):629–632. doi: 10.1038/nm1772. [DOI] [PubMed] [Google Scholar]
- 54.Ye ZC, Rothstein JD, Sontheimer H. Compromised glutamate transport in human glioma cells: reduction-mislocalization of sodium-dependent glutamate transporters and enhanced activity of cystine-glutamate exchange. The Journal of neuroscience : the official journal of the Society for Neuroscience. 1999;19(24):10767–10777. doi: 10.1523/JNEUROSCI.19-24-10767.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Ju HQ, Lu YX, Chen DL, Tian T, Mo HY, Wei XL, Liao JW, Wang F, Zeng ZL, Pelicano H, et al. Redox regulation of stem-like cells though the CD44v-xCT Axis in colorectal Cancer: mechanisms and therapeutic implications. Theranostics. 2016;6(8):1160–1175. doi: 10.7150/thno.14848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Yoshida GJ, Fuchimoto Y, Osumi T, Shimada H, Hosaka S, Morioka H, Mukai M, Masugi Y, Sakamoto M, Kuroda T. Li-Fraumeni syndrome with simultaneous osteosarcoma and liver cancer: increased expression of a CD44 variant isoform after chemotherapy. BMC Cancer. 2012;12:444. doi: 10.1186/1471-2407-12-444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lue HW, Podolak J, Kolahi K, Cheng L, Rao S, Garg D, Xue CH, Rantala JK, Tyner JW, Thornburg KL, et al. Metabolic reprogramming ensures cancer cell survival despite oncogenic signaling blockade. Genes Dev. 2017;31(20):2067–2084. doi: 10.1101/gad.305292.117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Strickland M, Stoll EA. Metabolic reprogramming in glioma. Frontiers in cell and developmental biology. 2017;5:43. doi: 10.3389/fcell.2017.00043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Danhier P, Banski P, Payen VL, Grasso D, Ippolito L, Sonveaux P, Porporato PE. Cancer metabolism in space and time: beyond the Warburg effect. Biochim Biophys Acta. 2017;1858(8):556–572. doi: 10.1016/j.bbabio.2017.02.001. [DOI] [PubMed] [Google Scholar]
- 60.Chen CL, Uthaya Kumar DB, Punj V, Xu J, Sher L, Tahara SM, Hess S, Machida K. NANOG metabolically reprograms tumor-initiating stem-like cells through tumorigenic changes in oxidative phosphorylation and fatty acid metabolism. Cell Metab. 2016;23(1):206–219. doi: 10.1016/j.cmet.2015.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Ito K, Suda T. Metabolic requirements for the maintenance of self-renewing stem cells. Nat Rev Mol Cell Biol. 2014;15(4):243–256. doi: 10.1038/nrm3772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Shyh-Chang N, Daley GQ, Cantley LC. Stem cell metabolism in tissue development and aging. Development. 2013;140(12):2535–2547. doi: 10.1242/dev.091777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Riester M, Xu Q, Moreira A, Zheng J, Michor F, Downey RJ. The Warburg effect: persistence of stem cell metabolism in cancers as a failure of differentiation. Ann Oncol. 2018;29(1):264–70. [DOI] [PMC free article] [PubMed]
- 64.Vega-Naredo I, Loureiro R, Mesquita KA, Barbosa IA, Tavares LC, Branco AF, Erickson JR, Holy J, Perkins EL, Carvalho RA, et al. Mitochondrial metabolism directs stemness and differentiation in P19 embryonal carcinoma stem cells. Cell Death Differ. 2014;21(10):1560–1574. doi: 10.1038/cdd.2014.66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Peng F, Wang JH, Fan WJ, Meng YT, Li MM, Li TT, Cui B, Wang HF, Zhao Y, An F, et al. Glycolysis gatekeeper PDK1 reprograms breast cancer stem cells under hypoxia. Oncogene. 2018;37(8):1062–74. [DOI] [PMC free article] [PubMed]
- 66.Corominas-Faja B, Cuyas E, Gumuzio J, Bosch-Barrera J, Leis O, Martin AG, Menendez JA. Chemical inhibition of acetyl-CoA carboxylase suppresses self-renewal growth of cancer stem cells. Oncotarget. 2014;5(18):8306–8316. doi: 10.18632/oncotarget.2059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Valent P, Bonnet D, Wohrer S, Andreeff M, Copland M, Chomienne C, Eaves C. Heterogeneity of neoplastic stem cells: theoretical, functional, and clinical implications. Cancer Res. 2013;73(3):1037–1045. doi: 10.1158/0008-5472.CAN-12-3678. [DOI] [PubMed] [Google Scholar]
- 68.Coles NW, Johnstone RM. Glutamine metabolism in Ehrlich ascites-carcinoma cells. The Biochemical journal. 1962;83:284–291. doi: 10.1042/bj0830284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Bartesaghi S, Graziano V, Galavotti S, Henriquez NV, Betts J, Saxena J, Minieri V, A D, Karlsson A, Martins LM, et al. Inhibition of oxidative metabolism leads to p53 genetic inactivation and transformation in neural stem cells. Proc Natl Acad Sci U S A. 2015;112(4):1059–1064. doi: 10.1073/pnas.1413165112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Zhang H, Badur MG, Divakaruni AS, Parker SJ, Jager C, Hiller K, Murphy AN, Metallo CM. Distinct metabolic states can support self-renewal and lipogenesis in human pluripotent stem cells under different culture conditions. Cell Rep. 2016;16(6):1536–1547. doi: 10.1016/j.celrep.2016.06.102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Alptekin A, Ye B, Ding HF. Transcriptional regulation of stem cell and Cancer stem cell metabolism. Current stem cell reports. 2017;3(1):19–27. doi: 10.1007/s40778-017-0071-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Pasto A, Bellio C, Pilotto G, Ciminale V, Silic-Benussi M, Guzzo G, Rasola A, Frasson C, Nardo G, Zulato E, et al. Cancer stem cells from epithelial ovarian cancer patients privilege oxidative phosphorylation, and resist glucose deprivation. Oncotarget. 2014;5(12):4305–4319. doi: 10.18632/oncotarget.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Di Francesco AM, Toesca A, Cenciarelli C, Giordano A, Gasbarrini A, Puglisi MA. Metabolic modification in gastrointestinal Cancer stem cells: characteristics and therapeutic approaches. J Cell Physiol. 2016;231(10):2081–2087. doi: 10.1002/jcp.25318. [DOI] [PubMed] [Google Scholar]
- 74.Lee E, Yang J, Ku M, Kim NH, Park Y, Park CB, Suh JS, Park ES, Yook JI, Mills GB, et al. Metabolic stress induces a Wnt-dependent cancer stem cell-like state transition. Cell Death Dis. 2015;6:e1805. doi: 10.1038/cddis.2015.171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Mahalingaiah PK, Ponnusamy L, Singh KP. Chronic oxidative stress leads to malignant transformation along with acquisition of stem cell characteristics, and epithelial to mesenchymal transition in human renal epithelial cells. J Cell Physiol. 2015;230(8):1916–1928. doi: 10.1002/jcp.24922. [DOI] [PubMed] [Google Scholar]
- 76.Toyokuni S, Okamoto K, Yodoi J, Hiai H. Persistent oxidative stress in cancer. FEBS Lett. 1995;358(1):1–3. doi: 10.1016/0014-5793(94)01368-B. [DOI] [PubMed] [Google Scholar]
- 77.Gao AM, Ke ZP, Shi F, Sun GC, Chen H. Chrysin enhances sensitivity of BEL-7402/ADM cells to doxorubicin by suppressing PI3K/Akt/Nrf2 and ERK/Nrf2 pathway. Chem Biol Interact. 2013;206(1):100–108. doi: 10.1016/j.cbi.2013.08.008. [DOI] [PubMed] [Google Scholar]
- 78.Ouyang WC, Liao YW, Chen PN, Lu KH, Yu CC, Hsieh PL. Hinokitiol suppresses cancer stemness and oncogenicity in glioma stem cells by Nrf2 regulation. Cancer Chemother Pharmacol. 2017;80(2):411–419. doi: 10.1007/s00280-017-3381-y. [DOI] [PubMed] [Google Scholar]
- 79.Welcker M, Clurman BE. FBW7 ubiquitin ligase: a tumour suppressor at the crossroads of cell division, growth and differentiation. Nat Rev Cancer. 2008;8(2):83–93. doi: 10.1038/nrc2290. [DOI] [PubMed] [Google Scholar]
- 80.Yoshida GJ. The heterogeneity of cancer stem-like cells at the invasive front. Cancer Cell Int. 2017;17:23. doi: 10.1186/s12935-017-0393-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Calabrese C, Poppleton H, Kocak M, Hogg TL, Fuller C, Hamner B, Oh EY, Gaber MW, Finklestein D, Allen M, et al. A perivascular niche for brain tumor stem cells. Cancer Cell. 2007;11(1):69–82. doi: 10.1016/j.ccr.2006.11.020. [DOI] [PubMed] [Google Scholar]
- 82.Heddleston JM, Li Z, Lathia JD, Bao S, Hjelmeland AB, Rich JN. Hypoxia inducible factors in cancer stem cells. Br J Cancer. 2010;102(5):789–795. doi: 10.1038/sj.bjc.6605551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Pietras A, Katz AM, Ekstrom EJ, Wee B, Halliday JJ, Pitter KL, Werbeck JL, Amankulor NM, Huse JT, Holland EC. Osteopontin-CD44 signaling in the glioma perivascular niche enhances cancer stem cell phenotypes and promotes aggressive tumor growth. Cell Stem Cell. 2014;14(3):357–369. doi: 10.1016/j.stem.2014.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Keith B, Simon MC. Hypoxia-inducible factors, stem cells, and cancer. Cell. 2007;129(3):465–472. doi: 10.1016/j.cell.2007.04.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Liang D, Ma Y, Liu J, Trope CG, Holm R, Nesland JM, Suo Z. The hypoxic microenvironment upgrades stem-like properties of ovarian cancer cells. BMC Cancer. 2012;12:201. doi: 10.1186/1471-2407-12-201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Covello KL, Kehler J, Yu H, Gordan JD, Arsham AM, Hu CJ, Labosky PA, Simon MC, Keith B. HIF-2alpha regulates Oct-4: effects of hypoxia on stem cell function, embryonic development, and tumor growth. Genes Dev. 2006;20(5):557–570. doi: 10.1101/gad.1399906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Gordan JD, Bertout JA, Hu CJ, Diehl JA, Simon MC. HIF-2alpha promotes hypoxic cell proliferation by enhancing c-myc transcriptional activity. Cancer Cell. 2007;11(4):335–347. doi: 10.1016/j.ccr.2007.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Liu PP, Liao J, Tang ZJ, Wu WJ, Yang J, Zeng ZL, Hu Y, Wang P, Ju HQ, Xu RH, et al. Metabolic regulation of cancer cell side population by glucose through activation of the Akt pathway. Cell Death Differ. 2014;21(1):124–135. doi: 10.1038/cdd.2013.131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Wang T, Fahrmann JF, Lee H, Li YJ, Tripathi SC, Yue C, Zhang C, Lifshitz V, Song J, Yuan Y, et al. JAK/STAT3-regulated fatty acid beta-oxidation is critical for breast Cancer stem cell self-renewal and Chemoresistance. Cell Metab. 2018;27(1):136–50. [DOI] [PMC free article] [PubMed]
- 90.Li J, Condello S, Thomes-Pepin J, Ma X, Xia Y, Hurley TD, Matei D, Cheng JX. Lipid desaturation is a metabolic marker and therapeutic target of ovarian Cancer stem cells. Cell Stem Cell. 2017;20(3):303–314. doi: 10.1016/j.stem.2016.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Brandi J, Dando I, Pozza ED, Biondani G, Jenkins R, Elliott V, Park K, Fanelli G, Zolla L, Costello E, et al. Proteomic analysis of pancreatic cancer stem cells: functional role of fatty acid synthesis and mevalonate pathways. J Proteome. 2017;150:310–322. doi: 10.1016/j.jprot.2016.10.002. [DOI] [PubMed] [Google Scholar]
- 92.Folmes CD, Park S, Terzic A. Lipid metabolism greases the stem cell engine. Cell Metab. 2013;17(2):153–155. doi: 10.1016/j.cmet.2013.01.010. [DOI] [PubMed] [Google Scholar]
- 93.Knobloch M, Braun SM, Zurkirchen L, von Schoultz C, Zamboni N, Arauzo-Bravo MJ, Kovacs WJ, Karalay O, Suter U, Machado RA, et al. Metabolic control of adult neural stem cell activity by Fasn-dependent lipogenesis. Nature. 2013;493(7431):226–230. doi: 10.1038/nature11689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Vlashi E, Lagadec C, Vergnes L, Matsutani T, Masui K, Poulou M, Popescu R, Della Donna L, Evers P, Dekmezian C, et al. Metabolic state of glioma stem cells and nontumorigenic cells. Proc Natl Acad Sci U S A. 2011;108(38):16062–16067. doi: 10.1073/pnas.1106704108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Tirinato L, Pagliari F, Limongi T, Marini M, Falqui A, Seco J, Candeloro P, Liberale C, Di Fabrizio E. An overview of lipid droplets in Cancer and Cancer stem cells. Stem Cells Int. 2017;2017:1656053. doi: 10.1155/2017/1656053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Maan M, Peters JM, Dutta M, Patterson AD. Lipid metabolism and lipophagy in cancer. Biochem Biophys Res Commun. 2018; S0006-291X(18)30326-7. 10.1016/j.bbrc.2018.02.097. [DOI] [PMC free article] [PubMed]
- 97.O'Malley J, Kumar R, Kuzmin AN, Pliss A, Yadav N, Balachandar S, Wang J, Attwood K, Prasad PN, Chandra D. Lipid quantification by Raman microspectroscopy as a potential biomarker in prostate cancer. Cancer Lett. 2017;397:52–60. doi: 10.1016/j.canlet.2017.03.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Du W, Zhang L, Brett-Morris A, Aguila B, Kerner J, Hoppel CL, Puchowicz M, Serra D, Herrero L, Rini BI, et al. HIF drives lipid deposition and cancer in ccRCC via repression of fatty acid metabolism. Nat Commun. 2017;8(1):1769. doi: 10.1038/s41467-017-01965-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Menard JA, Christianson HC, Kucharzewska P, Bourseau-Guilmain E, Svensson KJ, Lindqvist E, Indira Chandran V, Kjellen L, Welinder C, Bengzon J, et al. Metastasis stimulation by hypoxia and acidosis-induced extracellular lipid uptake is mediated by proteoglycan-dependent endocytosis. Cancer Res. 2016;76(16):4828–4840. doi: 10.1158/0008-5472.CAN-15-2831. [DOI] [PubMed] [Google Scholar]
- 100.Tirinato L, Liberale C, Di Franco S, Candeloro P, Benfante A, La Rocca R, Potze L, Marotta R, Ruffilli R, Rajamanickam VP, et al. Lipid droplets: a new player in colorectal cancer stem cells unveiled by spectroscopic imaging. Stem Cells. 2015;33(1):35–44. doi: 10.1002/stem.1837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Bailey AP, Koster G, Guillermier C, Hirst EM, MacRae JI, Lechene CP, Postle AD, Gould AP. Antioxidant role for lipid droplets in a stem cell niche of Drosophila. Cell. 2015;163(2):340–353. doi: 10.1016/j.cell.2015.09.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Kinlaw WB, Baures PW, Lupien LE, Davis WL, Kuemmerle NB. Fatty acids and breast Cancer: make them on site or have them delivered. J Cell Physiol. 2016;231(10):2128–2141. doi: 10.1002/jcp.25332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Yasumoto Y, Miyazaki H, Vaidyan LK, Kagawa Y, Ebrahimi M, Yamamoto Y, Ogata M, Katsuyama Y, Sadahiro H, Suzuki M, et al. Inhibition of fatty acid synthase decreases expression of Stemness markers in glioma stem cells. PLoS One. 2016;11(1):e0147717. doi: 10.1371/journal.pone.0147717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Pandey PR, Xing F, Sharma S, Watabe M, Pai SK, Iiizumi-Gairani M, Fukuda K, Hirota S, Mo YY, Watabe K. Elevated lipogenesis in epithelial stem-like cell confers survival advantage in ductal carcinoma in situ of breast cancer. Oncogene. 2013;32(42):5111–5122. doi: 10.1038/onc.2012.519. [DOI] [PubMed] [Google Scholar]
- 105.Peck B, Schug ZT, Zhang Q, Dankworth B, Jones DT, Smethurst E, Patel R, Mason S, Jiang M, Saunders R, et al. Inhibition of fatty acid desaturation is detrimental to cancer cell survival in metabolically compromised environments. Cancer & metabolism. 2016;4:6. doi: 10.1186/s40170-016-0146-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Mason P, Liang B, Li L, Fremgen T, Murphy E, Quinn A, Madden SL, Biemann HP, Wang B, Cohen A, et al. SCD1 inhibition causes cancer cell death by depleting mono-unsaturated fatty acids. PLoS One. 2012;7(3):e33823. doi: 10.1371/journal.pone.0033823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Kawahara I, Mori T, Goto K, Fujii K, Ohmori H, Kishi S, Fujiwara-Tani R, Kuniyasu H. Fatty acids induce Stemness in the stromal cells of a CT26 mouse tumor model. Pathobiology. 2017;84(5):237–242. doi: 10.1159/000471854. [DOI] [PubMed] [Google Scholar]
- 108.Kim YC, Ntambi JM. Regulation of stearoyl-CoA desaturase genes: role in cellular metabolism and preadipocyte differentiation. Biochem Biophys Res Commun. 1999;266(1):1–4. doi: 10.1006/bbrc.1999.1704. [DOI] [PubMed] [Google Scholar]
- 109.Colacino JA, McDermott SP, Sartor MA, Wicha MS, Rozek LS. Transcriptomic profiling of curcumin-treated human breast stem cells identifies a role for stearoyl-coa desaturase in breast cancer prevention. Breast Cancer Res Treat. 2016;158(1):29–41. doi: 10.1007/s10549-016-3854-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Taraboletti G, Perin L, Bottazzi B, Mantovani A, Giavazzi R, Salmona M. Membrane fluidity affects tumor-cell motility, invasion and lung-colonizing potential. Int J Cancer. 1989;44(4):707–713. doi: 10.1002/ijc.2910440426. [DOI] [PubMed] [Google Scholar]
- 111.Zhao W, Prijic S, Urban BC, Tisza MJ, Zuo Y, Li L, Tan Z, Chen X, Mani SA, Chang JT. Candidate Antimetastasis drugs suppress the metastatic capacity of breast Cancer cells by reducing membrane fluidity. Cancer Res. 2016;76(7):2037–2049. doi: 10.1158/0008-5472.CAN-15-1970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Pinel A, Rigaudiere JP, Laillet B, Pouyet C, Malpuech-Brugere C, Prip-Buus C, Morio B, Capel F. N-3PUFA differentially modulate palmitate-induced lipotoxicity through alterations of its metabolism in C2C12 muscle cells. Biochim Biophys Acta. 2016;1861(1):12–20. doi: 10.1016/j.bbalip.2015.10.003. [DOI] [PubMed] [Google Scholar]
- 113.Shen Y, Zhao Z, Zhang L, Shi L, Shahriar S, Chan RB, Di Paolo G, Min W. Metabolic activity induces membrane phase separation in endoplasmic reticulum. Proc Natl Acad Sci U S A. 2017;114(51):13394–13399. doi: 10.1073/pnas.1712555114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Lin L, Ding Y, Wang Y, Wang Z, Yin X, Yan G, Zhang L, Yang P, Shen H. Functional lipidomics: palmitic acid impairs hepatocellular carcinoma development by modulating membrane fluidity and glucose metabolism. Hepatology. 2017;66(2):432–448. doi: 10.1002/hep.29033. [DOI] [PubMed] [Google Scholar]
- 115.Wang W, Tao K, Wang J, Yang G, Ouyang Q, Wang Y, Zhang L, Liu F. Exploring the inhibitory effect of membrane tension on cell polarization. PLoS Comput Biol. 2017;13(1):e1005354. doi: 10.1371/journal.pcbi.1005354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Goehring NW, Grill SW. Cell polarity: mechanochemical patterning. Trends Cell Biol. 2013;23(2):72–80. doi: 10.1016/j.tcb.2012.10.009. [DOI] [PubMed] [Google Scholar]
- 117.Chu CW, Xiang B, Ossipova O, Ioannou A, Sokol SY. The Ajuba family protein Wtip regulates actomyosin contractility during vertebrate neural tube closure. J Cell Sci. 2018;131(10):jcs213884. 10.1242/jcs.213884. [DOI] [PMC free article] [PubMed]
- 118.Lai KKY, Kweon SM, Chi F, Hwang E, Kabe Y, Higashiyama R, Qin L, Yan R, Wu RP, Lai K, et al. Stearoyl-CoA desaturase promotes liver fibrosis and tumor development in mice via a Wnt positive-signaling loop by stabilization of low-density lipoprotein-receptor-related proteins 5 and 6. Gastroenterology. 2017;152(6):1477–1491. doi: 10.1053/j.gastro.2017.01.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Noto A, De Vitis C, Pisanu ME, Roscilli G, Ricci G, Catizone A, Sorrentino G, Chianese G, Taglialatela-Scafati O, Trisciuoglio D, et al. Stearoyl-CoA-desaturase 1 regulates lung cancer stemness via stabilization and nuclear localization of YAP/TAZ. Oncogene. 2017;36(32):4573–4584. doi: 10.1038/onc.2017.75. [DOI] [PubMed] [Google Scholar]
- 120.Smith AN, Muffley LA, Bell AN, Numhom S, Hocking AM. Unsaturated fatty acids induce mesenchymal stem cells to increase secretion of angiogenic mediators. J Cell Physiol. 2012;227(9):3225–3233. doi: 10.1002/jcp.24013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Martinez-Outschoorn UE, Pestell RG, Howell A, Tykocinski ML, Nagajyothi F, Machado FS, Tanowitz HB, Sotgia F, Lisanti MP. Energy transfer in "parasitic" cancer metabolism: mitochondria are the powerhouse and Achilles' heel of tumor cells. Cell Cycle. 2011;10(24):4208–4216. doi: 10.4161/cc.10.24.18487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Singh SR, Zeng X, Zhao J, Liu Y, Hou G, Liu H, Hou SX. The lipolysis pathway sustains normal and transformed stem cells in adult Drosophila. Nature. 2016;538(7623):109–113. doi: 10.1038/nature19788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Giampietri C, Petrungaro S, Cordella M, Tabolacci C, Tomaipitinca L, Facchiano A, Eramo A, Filippini A, Facchiano F, Ziparo E. Lipid storage and autophagy in melanoma Cancer cells. Int J Mol Sci. 2017;18(6) [DOI] [PMC free article] [PubMed]
- 124.Ye H, Adane B, Khan N, Sullivan T, Minhajuddin M, Gasparetto M, Stevens B, Pei S, Balys M, Ashton JM, et al. Leukemic stem cells evade chemotherapy by metabolic adaptation to an adipose tissue niche. Cell Stem Cell. 2016;19(1):23–37. doi: 10.1016/j.stem.2016.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Hale JS, Otvos B, Sinyuk M, Alvarado AG, Hitomi M, Stoltz K, Wu Q, Flavahan W, Levison B, Johansen ML, et al. Cancer stem cell-specific scavenger receptor CD36 drives glioblastoma progression. Stem Cells. 2014;32(7):1746–1758. doi: 10.1002/stem.1716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Pascual G, Avgustinova A, Mejetta S, Martin M, Castellanos A, Attolini CS, Berenguer A, Prats N, Toll A, Hueto JA, et al. Targeting metastasis-initiating cells through the fatty acid receptor CD36. Nature. 2017;541(7635):41–45. doi: 10.1038/nature20791. [DOI] [PubMed] [Google Scholar]
- 127.Moon BS, Jeong WJ, Park J, Kim TI, Min do S, Choi KY. Role of oncogenic K-Ras in cancer stem cell activation by aberrant Wnt/beta-catenin signaling. J Natl Cancer Inst. 2014;106(2):djt373. doi: 10.1093/jnci/djt373. [DOI] [PubMed] [Google Scholar]
- 128.Guo JY, Karsli-Uzunbas G, Mathew R, Aisner SC, Kamphorst JJ, Strohecker AM, Chen G, Price S, Lu W, Teng X, et al. Autophagy suppresses progression of K-ras-induced lung tumors to oncocytomas and maintains lipid homeostasis. Genes Dev. 2013;27(13):1447–1461. doi: 10.1101/gad.219642.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Camarda R, Zhou AY, Kohnz RA, Balakrishnan S, Mahieu C, Anderton B, Eyob H, Kajimura S, Tward A, Krings G, et al. Inhibition of fatty acid oxidation as a therapy for MYC-overexpressing triple-negative breast cancer. Nat Med. 2016;22(4):427–432. doi: 10.1038/nm.4055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Wright HJ, Hou J, Xu B, Cortez M, Potma EO, Tromberg BJ, Razorenova OV. CDCP1 drives triple-negative breast cancer metastasis through reduction of lipid-droplet abundance and stimulation of fatty acid oxidation. Proc Natl Acad Sci U S A. 2017;114(32):E6556–E6565. doi: 10.1073/pnas.1703791114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Carracedo A, Cantley LC, Pandolfi PP. Cancer metabolism: fatty acid oxidation in the limelight. Nat Rev Cancer. 2013;13(4):227–232. doi: 10.1038/nrc3483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Raulien N, Friedrich K, Strobel S, Rubner S, Baumann S, von Bergen M, Korner A, Krueger M, Rossol M, Wagner U. Fatty acid oxidation compensates for lipopolysaccharide-induced Warburg effect in glucose-deprived monocytes. Front Immunol. 2017;8:609. doi: 10.3389/fimmu.2017.00609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Liu C, Li Y, Xing Y, Cao B, Yang F, Yang T, Ai Z, Wei Y, Jiang J. The interaction between Cancer stem cell marker CD133 and Src protein promotes focal adhesion kinase (FAK) phosphorylation and cell migration. J Biol Chem. 2016;291(30):15540–15550. doi: 10.1074/jbc.M115.712976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Park JH, Vithayathil S, Kumar S, Sung PL, Dobrolecki LE, Putluri V, Bhat VB, Bhowmik SK, Gupta V, Arora K, et al. Fatty acid oxidation-driven Src links mitochondrial energy reprogramming and oncogenic properties in triple-negative breast Cancer. Cell Rep. 2016;14(9):2154–2165. doi: 10.1016/j.celrep.2016.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Lee EA, Angka L, Rota SG, Hanlon T, Mitchell A, Hurren R, Wang XM, Gronda M, Boyaci E, Bojko B, et al. Targeting mitochondria with Avocatin B induces selective leukemia cell death. Cancer Res. 2015;75(12):2478–2488. doi: 10.1158/0008-5472.CAN-14-2676. [DOI] [PubMed] [Google Scholar]
- 136.Yoshida GJ, Saya H. Therapeutic strategies targeting cancer stem cells. Cancer Sci. 2016;107(1):5–11. doi: 10.1111/cas.12817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Ito K, Carracedo A, Weiss D, Arai F, Ala U, Avigan DE, Schafer ZT, Evans RM, Suda T, Lee CH, et al. A PML-PPAR-delta pathway for fatty acid oxidation regulates hematopoietic stem cell maintenance. Nat Med. 2012;18(9):1350–1358. doi: 10.1038/nm.2882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Knobloch M, Pilz GA, Ghesquiere B, Kovacs WJ, Wegleiter T, Moore DL, Hruzova M, Zamboni N, Carmeliet P, Jessberger S. A fatty acid oxidation-dependent metabolic shift regulates adult neural stem cell activity. Cell Rep. 2017;20(9):2144–2155. doi: 10.1016/j.celrep.2017.08.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Xie Z, Jones A, Deeney JT, Hur SK, Bankaitis VA. Inborn errors of long-chain fatty acid beta-oxidation link neural stem cell self-renewal to autism. Cell Rep. 2016;14(5):991–999. doi: 10.1016/j.celrep.2016.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Ito K, Turcotte R, Cui J, Zimmerman SE, Pinho S, Mizoguchi T, Arai F, Runnels JM, Alt C, Teruya-Feldstein J, et al. Self-renewal of a purified Tie2+ hematopoietic stem cell population relies on mitochondrial clearance. Science. 2016;354(6316):1156–1160. doi: 10.1126/science.aaf5530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Wong BW, Wang X, Zecchin A, Thienpont B, Cornelissen I, Kalucka J, Garcia-Caballero M, Missiaen R, Huang H, Bruning U, et al. The role of fatty acid beta-oxidation in lymphangiogenesis. Nature. 2017;542(7639):49–54. doi: 10.1038/nature21028. [DOI] [PubMed] [Google Scholar]
- 142.Chang B, Park MJ, Choi SI, In KH, Kim CH, Lee SH. NANOG as an adverse predictive marker in advanced non-small cell lung cancer treated with platinum-based chemotherapy. OncoTargets and therapy. 2017;10:4625–4633. doi: 10.2147/OTT.S144895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Kenda Suster N, Frkovic Grazio S, Virant-Klun I, Verdenik I, Smrkolj S. Cancer stem cell-related marker NANOG expression in ovarian serous tumors: a Clinicopathological study of 159 cases. Int J Gynecol Cancer. 2017;27(9):2006–2013. doi: 10.1097/IGC.0000000000001105. [DOI] [PubMed] [Google Scholar]
- 144.Soni P, Qayoom S, Husain N, Kumar P, Chandra A, Ojha BK, Gupta RK. CD24 and Nanog expression in stem cells in glioblastoma: correlation with response to Chemoradiation and overall survival. Asian Pacific journal of cancer prevention : APJCP. 2017;18(8):2215–2219. doi: 10.22034/APJCP.2017.18.8.2215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Zbinden M, Duquet A, Lorente-Trigos A, Ngwabyt SN, Borges I, Ruiz i, Altaba A. NANOG regulates glioma stem cells and is essential in vivo acting in a cross-functional network with GLI1 and p53. EMBO J. 2010;29(15):2659–2674. doi: 10.1038/emboj.2010.137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Zhang J, Espinoza LA, Kinders RJ, Lawrence SM, Pfister TD, Zhou M, Veenstra TD, Thorgeirsson SS, Jessup JM. NANOG modulates stemness in human colorectal cancer. Oncogene. 2013;32(37):4397–4405. doi: 10.1038/onc.2012.461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Baillie R, Tan ST, Itinteang T. Cancer stem cells in oral cavity squamous cell carcinoma: a review. Front Oncol. 2017;7:112. doi: 10.3389/fonc.2017.00112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Zhang L, Pan Y, Qin G, Chen L, Chatterjee TK, Weintraub NL, Tang Y. Inhibition of stearoyl-coA desaturase selectively eliminates tumorigenic Nanog-positive cells: improving the safety of iPS cell transplantation to myocardium. Cell Cycle. 2014;13(5):762–771. doi: 10.4161/cc.27677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Shimano H, Sato R. SREBP-regulated lipid metabolism: convergent physiology - divergent pathophysiology. Nat Rev Endocrinol. 2017; [DOI] [PubMed]
- 150.Griffiths B, Lewis CA, Bensaad K, Ros S, Zhang Q, Ferber EC, Konisti S, Peck B, Miess H, East P, et al. Sterol regulatory element binding protein-dependent regulation of lipid synthesis supports cell survival and tumor growth. Cancer & metabolism. 2013;1(1):3. doi: 10.1186/2049-3002-1-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Lee JH, Jeon YG, Lee KH, Lee HW, Park J, Jang H, Kang M, Lee HS, Cho HJ, Nam DH, et al. RNF20 suppresses tumorigenesis by inhibiting SREBP1c-PTTG1 Axis in kidney Cancer. Mol Cell Biol. 2017; MCB.00265-17. 10.1128/MCB.00265-17 [DOI] [PMC free article] [PubMed]
- 152.Fan XX, Leung EL, Xie Y, Liu ZQ, Zheng YF, Yao XJ, Lu LL, Wu JL, He JX, Yuan ZW, et al. Suppression of lipogenesis via reactive oxygen species-AMPK signaling for treating malignant and proliferative diseases. Antioxid Redox Signal. 2017. [DOI] [PubMed]
- 153.Luo D, Xiao H, Dong J, Li Y, Feng G, Cui M, Fan S. B7-H3 regulates lipid metabolism of lung cancer through SREBP1-mediated expression of FASN. Biochem Biophys Res Commun. 2017;482(4):1246–1251. doi: 10.1016/j.bbrc.2016.12.021. [DOI] [PubMed] [Google Scholar]
- 154.Li W, Tai Y, Zhou J, Gu W, Bai Z, Zhou T, Zhong Z, McCue PA, Sang N, Ji JY, et al. Repression of endometrial tumor growth by targeting SREBP1 and lipogenesis. Cell Cycle. 2012;11(12):2348–2358. doi: 10.4161/cc.20811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Ricoult SJ, Yecies JL, Ben-Sahra I, Manning BD. Oncogenic PI3K and K-Ras stimulate de novo lipid synthesis through mTORC1 and SREBP. Oncogene. 2016;35(10):1250–1260. doi: 10.1038/onc.2015.179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Sun Y, He W, Luo M, Zhou Y, Chang G, Ren W, Wu K, Li X, Shen J, Zhao X, et al. SREBP1 regulates tumorigenesis and prognosis of pancreatic cancer through targeting lipid metabolism. Tumour Biol. 2015;36(6):4133–4141. doi: 10.1007/s13277-015-3047-5. [DOI] [PubMed] [Google Scholar]
- 157.Lewis CA, Brault C, Peck B, Bensaad K, Griffiths B, Mitter R, Chakravarty P, East P, Dankworth B, Alibhai D, et al. SREBP maintains lipid biosynthesis and viability of cancer cells under lipid- and oxygen-deprived conditions and defines a gene signature associated with poor survival in glioblastoma multiforme. Oncogene. 2015;34(40):5128–5140. doi: 10.1038/onc.2014.439. [DOI] [PubMed] [Google Scholar]
- 158.Bengoechea-Alonso MT, Ericsson J. The phosphorylation-dependent regulation of nuclear SREBP1 during mitosis links lipid metabolism and cell growth. Cell Cycle. 2016;15(20):2753–2765. doi: 10.1080/15384101.2016.1220456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Bengoechea-Alonso MT, Ericsson J. A phosphorylation cascade controls the degradation of active SREBP1. J Biol Chem. 2009;284(9):5885–5895. doi: 10.1074/jbc.M807906200. [DOI] [PubMed] [Google Scholar]
- 160.Bengoechea-Alonso MT, Ericsson J. Cdk1/cyclin B-mediated phosphorylation stabilizes SREBP1 during mitosis. Cell Cycle. 2006;5(15):1708–1718. doi: 10.4161/cc.5.15.3131. [DOI] [PubMed] [Google Scholar]
- 161.Porstmann T, Santos CR, Griffiths B, Cully M, Wu M, Leevers S, Griffiths JR, Chung YL, Schulze A. SREBP activity is regulated by mTORC1 and contributes to Akt-dependent cell growth. Cell Metab. 2008;8(3):224–236. doi: 10.1016/j.cmet.2008.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Yang W, Xia Y, Ji H, Zheng Y, Liang J, Huang W, Gao X, Aldape K, Lu Z. Nuclear PKM2 regulates beta-catenin transactivation upon EGFR activation. Nature. 2011;480(7375):118–122. doi: 10.1038/nature10598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Christofk HR, Vander Heiden MG, Harris MH, Ramanathan A, Gerszten RE, Wei R, Fleming MD, Schreiber SL, Cantley LC. The M2 splice isoform of pyruvate kinase is important for cancer metabolism and tumour growth. Nature. 2008;452(7184):230–233. doi: 10.1038/nature06734. [DOI] [PubMed] [Google Scholar]
- 164.Zhao X, Zhao L, Yang H, Li J, Min X, Yang F, Liu J, Huang G. Pyruvate kinase M2 interacts with nuclear sterol regulatory element-binding protein 1a and thereby activates lipogenesis and cell proliferation in hepatocellular carcinoma. J Biol Chem. 2018;293(17):6623–6634. doi: 10.1074/jbc.RA117.000100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Meyer N, Penn LZ. Reflecting on 25 years with MYC. Nat Rev Cancer. 2008;8(12):976–990. doi: 10.1038/nrc2231. [DOI] [PubMed] [Google Scholar]
- 166.Wahlstrom T, Henriksson MA. Impact of MYC in regulation of tumor cell metabolism. Biochim Biophys Acta. 2015;1849(5):563–569. doi: 10.1016/j.bbagrm.2014.07.004. [DOI] [PubMed] [Google Scholar]
- 167.Stine ZE, Walton ZE, Altman BJ, Hsieh AL, Dang CV. MYC, metabolism, and Cancer. Cancer discovery. 2015;5(10):1024–1039. doi: 10.1158/2159-8290.CD-15-0507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Camarda R, Williams J, Goga A. In vivo reprogramming of Cancer metabolism by MYC. Frontiers in cell and developmental biology. 2017;5:35. doi: 10.3389/fcell.2017.00035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Wang H, Lu J, Edmunds LR, Kulkarni S, Dolezal J, Tao J, Ranganathan S, Jackson L, Fromherz M, Beer-Stolz D, et al. Coordinated activities of multiple Myc-dependent and Myc-independent biosynthetic pathways in Hepatoblastoma. J Biol Chem. 2016;291(51):26241–26251. doi: 10.1074/jbc.M116.754218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Pacilli A, Calienni M, Margarucci S, D'Apolito M, Petillo O, Rocchi L, Pasquinelli G, Nicolai R, Koverech A, Calvani M, et al. Carnitine-acyltransferase system inhibition, cancer cell death, and prevention of myc-induced lymphomagenesis. J Natl Cancer Inst. 2013;105(7):489–498. doi: 10.1093/jnci/djt030. [DOI] [PubMed] [Google Scholar]
- 171.Harris AW, Pinkert CA, Crawford M, Langdon WY, Brinster RL, Adams JM. The E mu-myc transgenic mouse. A model for high-incidence spontaneous lymphoma and leukemia of early B cells. J Exp Med. 1988;167(2):353–371. doi: 10.1084/jem.167.2.353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Pfefferle AD, Herschkowitz JI, Usary J, Harrell JC, Spike BT, Adams JR, Torres-Arzayus MI, Brown M, Egan SE, Wahl GM, et al. Transcriptomic classification of genetically engineered mouse models of breast cancer identifies human subtype counterparts. Genome Biol. 2013;14(11):R125. doi: 10.1186/gb-2013-14-11-r125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Bednarski T, Olichwier A, Opasinska A, Pyrkowska A, Gan AM, Ntambi JM, Dobrzyn P. Stearoyl-CoA desaturase 1 deficiency reduces lipid accumulation in the heart by activating lipolysis independently of peroxisome proliferator-activated receptor alpha. Biochim Biophys Acta. 2016;1861(12 Pt A):2029–2037. doi: 10.1016/j.bbalip.2016.10.005. [DOI] [PubMed] [Google Scholar]
- 174.Li L, Wang C, Calvisi DF, Evert M, Pilo MG, Jiang L, Yuneva M, Chen X. SCD1 expression is dispensable for hepatocarcinogenesis induced by AKT and Ras oncogenes in mice. PLoS One. 2013;8(9):e75104. doi: 10.1371/journal.pone.0075104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Noto A, Raffa S, De Vitis C, Roscilli G, Malpicci D, Coluccia P, Di Napoli A, Ricci A, Giovagnoli MR, Aurisicchio L, et al. Stearoyl-CoA desaturase-1 is a key factor for lung cancer-initiating cells. Cell Death Dis. 2013;4:e947. doi: 10.1038/cddis.2013.444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Ma MKF, Lau EYT, Leung DHW, Lo J, Ho NPY, Cheng LKW, Ma S, Lin CH, Copland JA, Ding J, et al. Stearoyl-CoA desaturase regulates sorafenib resistance via modulation of ER stress-induced differentiation. J Hepatol. 2017;67(5):979–990. doi: 10.1016/j.jhep.2017.06.015. [DOI] [PubMed] [Google Scholar]
- 177.Pisanu ME, Noto A, De Vitis C, Morrone S, Scognamiglio G, Botti G, Venuta F, Diso D, Jakopin Z, Padula F, et al. Blockade of Stearoyl-CoA-desaturase 1 activity reverts resistance to cisplatin in lung cancer stem cells. Cancer Lett. 2017;406:93–104. doi: 10.1016/j.canlet.2017.07.027. [DOI] [PubMed] [Google Scholar]
- 178.Bansal S, Berk M, Alkhouri N, Partrick DA, Fung JJ, Feldstein A. Stearoyl-CoA desaturase plays an important role in proliferation and chemoresistance in human hepatocellular carcinoma. J Surg Res. 2014;186(1):29–38. doi: 10.1016/j.jss.2013.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Zhang H, Li H, Ho N, Li D, Li S. Scd1 plays a tumor-suppressive role in survival of leukemia stem cells and the development of chronic myeloid leukemia. Mol Cell Biol. 2012;32(10):1776–1787. doi: 10.1128/MCB.05672-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Luyimbazi D, Akcakanat A, McAuliffe PF, Zhang L, Singh G, Gonzalez-Angulo AM, Chen H, Do KA, Zheng Y, Hung MC, et al. Rapamycin regulates stearoyl CoA desaturase 1 expression in breast cancer. Mol Cancer Ther. 2010;9(10):2770–2784. doi: 10.1158/1535-7163.MCT-09-0980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Belkaid A, Duguay SR, Ouellette RJ, Surette ME. 17beta-estradiol induces stearoyl-CoA desaturase-1 expression in estrogen receptor-positive breast cancer cells. BMC Cancer. 2015;15:440. doi: 10.1186/s12885-015-1452-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Angelucci C, Maulucci G, Colabianchi A, Iacopino F, D'Alessio A, Maiorana A, Palmieri V, Papi M, De Spirito M, Di Leone A, et al. Stearoyl-CoA desaturase 1 and paracrine diffusible signals have a major role in the promotion of breast cancer cell migration induced by cancer-associated fibroblasts. Br J Cancer. 2015;112(10):1675–1686. doi: 10.1038/bjc.2015.135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Du X, Wang QR, Chan E, Merchant M, Liu J, French D, Ashkenazi A, Qing J. FGFR3 stimulates stearoyl CoA desaturase 1 activity to promote bladder tumor growth. Cancer Res. 2012;72(22):5843–5855. doi: 10.1158/0008-5472.CAN-12-1329. [DOI] [PubMed] [Google Scholar]
- 184.Potze L, di Franco S, Kessler JH, Stassi G, Medema JP. Betulinic acid kills Colon Cancer stem cells. Current stem cell research & therapy. 2016;11(5):427–433. doi: 10.2174/1574888X11666151203223512. [DOI] [PubMed] [Google Scholar]
- 185.Chen L, Ren J, Yang L, Li Y, Fu J, Li Y, Tian Y, Qiu F, Liu Z, Qiu Y. Stearoyl-CoA desaturase-1 mediated cell apoptosis in colorectal cancer by promoting ceramide synthesis. Sci Rep. 2016;6:19665. doi: 10.1038/srep19665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Ogasawara Y, Itakura E, Kono N, Mizushima N, Arai H, Nara A, Mizukami T, Yamamoto A. Stearoyl-CoA desaturase 1 activity is required for autophagosome formation. J Biol Chem. 2014;289(34):23938–23950. doi: 10.1074/jbc.M114.591065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Janikiewicz J, Hanzelka K, Dziewulska A, Kozinski K, Dobrzyn P, Bernas T, Dobrzyn A. Inhibition of SCD1 impairs palmitate-derived autophagy at the step of autophagosome-lysosome fusion in pancreatic beta-cells. J Lipid Res. 2015;56(10):1901–1911. doi: 10.1194/jlr.M059980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Huang GM, Jiang QH, Cai C, Qu M, Shen W. SCD1 negatively regulates autophagy-induced cell death in human hepatocellular carcinoma through inactivation of the AMPK signaling pathway. Cancer Lett. 2015;358(2):180–190. doi: 10.1016/j.canlet.2014.12.036. [DOI] [PubMed] [Google Scholar]
- 189.Pei Z, Sun P, Huang P, Lal B, Laterra J, Watkins PA. Acyl-CoA synthetase VL3 knockdown inhibits human glioma cell proliferation and tumorigenicity. Cancer Res. 2009;69(24):9175–9182. doi: 10.1158/0008-5472.CAN-08-4689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Pei Z, Fraisl P, Shi X, Gabrielson E, Forss-Petter S, Berger J, Watkins PA. Very long-chain acyl-CoA synthetase 3: overexpression and growth dependence in lung cancer. PLoS One. 2013;8(7):e69392. doi: 10.1371/journal.pone.0069392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Sun P, Xia S, Lal B, Shi X, Yang KS, Watkins PA, Laterra J. Lipid metabolism enzyme ACSVL3 supports glioblastoma stem cell maintenance and tumorigenicity. BMC Cancer. 2014;14:401. doi: 10.1186/1471-2407-14-401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Farge T, Saland E, de Toni F, Aroua N, Hosseini M, Perry R, Bosc C, Sugita M, Stuani L, Fraisse M, et al. Chemotherapy-resistant human acute myeloid leukemia cells are not enriched for leukemic stem cells but require oxidative metabolism. Cancer discovery. 2017;7(7):716–735. doi: 10.1158/2159-8290.CD-16-0441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Flaig TW, Salzmann-Sullivan M, Su LJ, Zhang Z, Joshi M, Gijon MA, Kim J, Arcaroli JJ, Van Bokhoven A, Lucia MS, et al. Lipid catabolism inhibition sensitizes prostate cancer cells to antiandrogen blockade. Oncotarget. 2017;8(34):56051–56065. doi: 10.18632/oncotarget.17359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Schlaepfer IR, Rider L, Rodrigues LU, Gijon MA, Pac CT, Romero L, Cimic A, Sirintrapun SJ, Glode LM, Eckel RH, et al. Lipid catabolism via CPT1 as a therapeutic target for prostate cancer. Mol Cancer Ther. 2014;13(10):2361–2371. doi: 10.1158/1535-7163.MCT-14-0183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Shi J, Fu H, Jia Z, He K, Fu L, Wang W. High expression of CPT1A predicts adverse outcomes: a potential therapeutic target for acute myeloid leukemia. EBioMedicine. 2016;14:55–64. doi: 10.1016/j.ebiom.2016.11.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Shao H, Mohamed EM, Xu GG, Waters M, Jing K, Ma Y, Zhang Y, Spiegel S, Idowu MO, Fang X. Carnitine palmitoyltransferase 1A functions to repress FoxO transcription factors to allow cell cycle progression in ovarian cancer. Oncotarget. 2016;7(4):3832–3846. doi: 10.18632/oncotarget.6757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Sung GJ, Choi HK, Kwak S, Song JH, Ko H, Yoon HG, Kang HB, Choi KC: Targeting CPT1A enhances metabolic therapy in human melanoma cells with the BRAF V600E mutation. Int J Biochem Cell Biol 2016, 81(Pt A):76–81. [DOI] [PubMed]
- 198.Pucci S, Zonetti MJ, Fisco T, Polidoro C, Bocchinfuso G, Palleschi A, Novelli G, Spagnoli LG, Mazzarelli P. Carnitine palmitoyl transferase-1A (CPT1A): a new tumor specific target in human breast cancer. Oncotarget. 2016;7(15):19982–19996. doi: 10.18632/oncotarget.6964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Jeong HJ, Oh HA, Nam SY, Han NR, Kim YS, Kim JH, Lee SJ, Kim MH, Moon PD, Kim HM, et al. The critical role of mast cell-derived hypoxia-inducible factor-1alpha in human and mice melanoma growth. Int J Cancer. 2013;132(11):2492–2501. doi: 10.1002/ijc.27937. [DOI] [PubMed] [Google Scholar]
- 200.Nicolau-Galmes F, Asumendi A, Alonso-Tejerina E, Perez-Yarza G, Jangi SM, Gardeazabal J, Arroyo-Berdugo Y, Careaga JM, Diaz-Ramon JL, Apraiz A, et al. Terfenadine induces apoptosis and autophagy in melanoma cells through ROS-dependent and -independent mechanisms. Apoptosis : an international journal on programmed cell death. 2011;16(12):1253–1267. doi: 10.1007/s10495-011-0640-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Sokolowska E, Presler M, Goyke E, Milczarek R, Swierczynski J, Sledzinski T. Orlistat reduces proliferation and enhances apoptosis in human pancreatic Cancer cells (PANC-1) Anticancer Res. 2017;37(11):6321–6327. doi: 10.21873/anticanres.12083. [DOI] [PubMed] [Google Scholar]
- 202.Papaevangelou E, Almeida GS, Box C, deSouza NM, Chung YL. The effect of FASN inhibition on the growth and metabolism of a cisplatin-resistant ovarian carcinoma model. Int J Cancer. 2018;. 10.1002/ijc.31392. [DOI] [PMC free article] [PubMed]
- 203.Ali A, Levantini E, Teo JT, Goggi J, Clohessy JG, Wu CS, Chen L, Yang H, Krishnan I, Kocher O, et al. Fatty acid synthase mediates EGFR palmitoylation in EGFR mutated non-small cell lung cancer. EMBO Mol Med. 2018;. 10.15252/emmm.201708313. [DOI] [PMC free article] [PubMed]
- 204.Baur JA, Sinclair DA. Therapeutic potential of resveratrol: the in vivo evidence. Nat Rev Drug Discov. 2006;5(6):493–506. doi: 10.1038/nrd2060. [DOI] [PubMed] [Google Scholar]
- 205.Xu J, Liu D, Niu H, Zhu G, Xu Y, Ye D, Li J, Zhang Q. Resveratrol reverses doxorubicin resistance by inhibiting epithelial-mesenchymal transition (EMT) through modulating PTEN/Akt signaling pathway in gastric cancer. Journal of experimental & clinical cancer research : CR. 2017;36(1):19. doi: 10.1186/s13046-016-0487-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Plauth A, Geikowski A, Cichon S, Wowro SJ, Liedgens L, Rousseau M, Weidner C, Fuhr L, Kliem M, Jenkins G, et al. Hormetic shifting of redox environment by pro-oxidative resveratrol protects cells against stress. Free Radic Biol Med. 2016;99:608–622. doi: 10.1016/j.freeradbiomed.2016.08.006. [DOI] [PubMed] [Google Scholar]
- 207.Catanzaro E, Calcabrini C, Turrini E, Sestili P, Fimognari C. Nrf2: a potential therapeutic target for naturally occurring anticancer drugs? Expert Opin Ther Targets. 2017;21(8):781–793. doi: 10.1080/14728222.2017.1351549. [DOI] [PubMed] [Google Scholar]
- 208.Li X, Fang P, Mai J, Choi ET, Wang H, Yang XF. Targeting mitochondrial reactive oxygen species as novel therapy for inflammatory diseases and cancers. J Hematol Oncol. 2013;6:19. doi: 10.1186/1756-8722-6-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Pandey PR, Okuda H, Watabe M, Pai SK, Liu W, Kobayashi A, Xing F, Fukuda K, Hirota S, Sugai T, et al. Resveratrol suppresses growth of cancer stem-like cells by inhibiting fatty acid synthase. Breast Cancer Res Treat. 2011;130(2):387–398. doi: 10.1007/s10549-010-1300-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Sayd S, Thirant C, El-Habr EA, Lipecka J, Dubois LG, Bogeas A, Tahiri-Jouti N, Chneiweiss H, Junier MP. Sirtuin-2 activity is required for glioma stem cell proliferation arrest but not necrosis induced by resveratrol. Stem Cell Rev. 2014;10(1):103–113. doi: 10.1007/s12015-013-9465-0. [DOI] [PubMed] [Google Scholar]
- 211.Wang YJ, Lin JF, Cheng LH, Chang WT, Kao YH, Chang MM, Wang BJ, Cheng HC. Pterostilbene prevents AKT-ERK axis-mediated polymerization of surface fibronectin on suspended lung cancer cells independently of apoptosis and suppresses metastasis. J Hematol Oncol. 2017;10(1):72. doi: 10.1186/s13045-017-0441-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Zhang L, Wen X, Li M, Li S, Zhao H. Targeting cancer stem cells and signaling pathways by resveratrol and pterostilbene. BioFactors. 2018;44(1):61–68. doi: 10.1002/biof.1398. [DOI] [PubMed] [Google Scholar]
- 213.Shiragami R, Murata S, Kosugi C, Tezuka T, Yamazaki M, Hirano A, Yoshimura Y, Suzuki M, Shuto K, Koda K. Enhanced antitumor activity of cerulenin combined with oxaliplatin in human colon cancer cells. Int J Oncol. 2013;43(2):431–438. doi: 10.3892/ijo.2013.1978. [DOI] [PubMed] [Google Scholar]
- 214.Jochen AL, Hays J, Mick G. Inhibitory effects of cerulenin on protein palmitoylation and insulin internalization in rat adipocytes. Biochim Biophys Acta. 1995;1259(1):65–72. doi: 10.1016/0005-2760(95)00147-5. [DOI] [PubMed] [Google Scholar]
- 215.Peck B, Schulze A. Lipid desaturation - the next step in targeting lipogenesis in cancer? FEBS J. 2016;283(15):2767–2778. doi: 10.1111/febs.13681. [DOI] [PubMed] [Google Scholar]
- 216.Mohammadzadeh F, Mosayebi G, Montazeri V, Darabi M, Fayezi S, Shaaker M, Rahmati M, Baradaran B, Mehdizadeh A, Darabi M. Fatty acid composition of tissue cultured breast carcinoma and the effect of Stearoyl-CoA desaturase 1 inhibition. J Breast Cancer. 2014;17(2):136–142. doi: 10.4048/jbc.2014.17.2.136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Imamura K, Tomita N, Kawakita Y, Ito Y, Ono K, Nii N, Miyazaki T, Yonemori K, Tawada M, Sumi H, et al. Discovery of novel and potent Stearoyl coenzyme a desaturase 1 (SCD1) inhibitors as anticancer agents. Bioorg Med Chem. 2017;25(14):3768–3779. doi: 10.1016/j.bmc.2017.05.016. [DOI] [PubMed] [Google Scholar]
- 218.Holubarsch CJ, Rohrbach M, Karrasch M, Boehm E, Polonski L, Ponikowski P, Rhein S. A double-blind randomized multicentre clinical trial to evaluate the efficacy and safety of two doses of etomoxir in comparison with placebo in patients with moderate congestive heart failure: the ERGO (etomoxir for the recovery of glucose oxidation) study. Clin Sci. 2007;113(4):205–212. doi: 10.1042/CS20060307. [DOI] [PubMed] [Google Scholar]
- 219.Samudio I, Harmancey R, Fiegl M, Kantarjian H, Konopleva M, Korchin B, Kaluarachchi K, Bornmann W, Duvvuri S, Taegtmeyer H, et al. Pharmacologic inhibition of fatty acid oxidation sensitizes human leukemia cells to apoptosis induction. J Clin Invest. 2010;120(1):142–156. doi: 10.1172/JCI38942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Estan MC, Calvino E, Calvo S, Guillen-Guio B, Boyano-Adanez Mdel C, de Blas E, Rial E, Aller P. Apoptotic efficacy of etomoxir in human acute myeloid leukemia cells. Cooperation with arsenic trioxide and glycolytic inhibitors, and regulation by oxidative stress and protein kinase activities. PLoS One. 2014;9(12):e115250. doi: 10.1371/journal.pone.0115250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Ricciardi MR, Mirabilii S, Allegretti M, Licchetta R, Calarco A, Torrisi MR, Foa R, Nicolai R, Peluso G, Tafuri A. Targeting the leukemia cell metabolism by the CPT1a inhibition: functional preclinical effects in leukemias. Blood. 2015;126(16):1925–1929. doi: 10.1182/blood-2014-12-617498. [DOI] [PubMed] [Google Scholar]
- 222.Shi Y, Su Z, Li S, Chen Y, Chen X, Xiao Y, Sun M, Ping Q, Zong L. Multistep targeted nano drug delivery system aiming at leukemic stem cells and minimal residual disease. Mol Pharm. 2013;10(6):2479–2489. doi: 10.1021/mp4001266. [DOI] [PubMed] [Google Scholar]
- 223.Wu K, Xing F, Wu SY, Watabe K. Extracellular vesicles as emerging targets in cancer: recent development from bench to bedside. Biochim Biophys Acta. 2017;1868(2):538–563. doi: 10.1016/j.bbcan.2017.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Liu J, Meng T, Yuan M, Wen L, Cheng B, Liu N, Huang X, Hong Y, Yuan H, Hu F. MicroRNA-200c delivered by solid lipid nanoparticles enhances the effect of paclitaxel on breast cancer stem cell. Int J Nanomedicine. 2016;11:6713–6725. doi: 10.2147/IJN.S111647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–674. doi: 10.1016/j.cell.2011.02.013. [DOI] [PubMed] [Google Scholar]
- 226.Thomas D, Majeti R. Burning Fat Fuels Leukemic Stem Cell Heterogeneity. Cell Stem Cell. 2016;19(1):1–2. doi: 10.1016/j.stem.2016.06.014. [DOI] [PubMed] [Google Scholar]
- 227.Choi SA, Wang KC, Phi JH, Lee JY, Park CK, Park SH, Kim SK. A distinct subpopulation within CD133 positive brain tumor cells shares characteristics with endothelial progenitor cells. Cancer Lett. 2012;324(2):221–230. doi: 10.1016/j.canlet.2012.05.026. [DOI] [PubMed] [Google Scholar]
- 228.Wang D, Huang J, Wang X, Yu Y, Zhang H, Chen Y, Liu J, Sun Z, Zou H, Sun D, et al. The eradication of breast cancer cells and stem cells by 8-hydroxyquinoline-loaded hyaluronan modified mesoporous silica nanoparticle-supported lipid bilayers containing docetaxel. Biomaterials. 2013;34(31):7662–7673. doi: 10.1016/j.biomaterials.2013.06.042. [DOI] [PubMed] [Google Scholar]
- 229.Krstic J, Trivanovic D, Jaukovic A, Santibanez JF, Bugarski D. Metabolic plasticity of stem cells and macrophages in Cancer. Front Immunol. 2017;8:939. doi: 10.3389/fimmu.2017.00939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Luo J, Hong Y, Lu Y, Qiu S, Chaganty BK, Zhang L, Wang X, Li Q, Fan Z. Acetyl-CoA carboxylase rewires cancer metabolism to allow cancer cells to survive inhibition of the Warburg effect by cetuximab. Cancer Lett. 2017;384:39–49. doi: 10.1016/j.canlet.2016.09.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Cilibrasi C, Riva G, Romano G, Cadamuro M, Bazzoni R, Butta V, Paoletta L, Dalpra L, Strazzabosco M, Lavitrano M, et al. Resveratrol impairs glioma stem cells proliferation and motility by modulating the Wnt signaling pathway. PLoS One. 2017;12(1):e0169854. doi: 10.1371/journal.pone.0169854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Subramaniam D, Kaushik G, Dandawate P, Anant S. Targeting cancer stem cells for chemoprevention of pancreatic cancer. Curr Med Chem. 2017; [Epub ahead of print] [DOI] [PMC free article] [PubMed]
- 233.von Roemeling CA, Marlow LA, Wei JJ, Cooper SJ, Caulfield TR, Wu K, Tan WW, Tun HW, Copland JA. Stearoyl-CoA desaturase 1 is a novel molecular therapeutic target for clear cell renal cell carcinoma. Clin Cancer Res. 2013;19(9):2368–2380. doi: 10.1158/1078-0432.CCR-12-3249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Nishizawa S, Sumi H, Satoh Y, Yamamoto Y, Kitazawa S, Honda K, Araki H, Kakoi K, Imamura K, Sasaki M, et al. In vitro and in vivo antitumor activities of T-3764518, a novel and orally available small molecule stearoyl-CoA desaturase 1 inhibitor. Eur J Pharmacol. 2017;807:21–31. doi: 10.1016/j.ejphar.2017.03.064. [DOI] [PubMed] [Google Scholar]