

Abstract
Nuclear factor Y (NF-Y) is a ubiquitous transcription factor found in eukaryotes. It is composed of three distinct subunits called NF-YA, NF-YB and NF-YC. NF-Ys have been identified as key regulators of multiple pathways in the control of development and tolerance to biotic and abiotic factors. The present study aimed to identify and characterize the complete repertoire of genes coding for NF-Y in citrus, as well as to perform the functional characterization of one of its members, namely CsNFYA5, in transgenic tobacco plants. A total of 22 genes coding for NF-Y were identified in the genomes of sweet orange (Citrus sinensis) and Clementine mandarin (C. clementina), including six CsNF-YAs, 11 CsNF-YBs and five CsNF-YCs. Phylogenetic analyses showed that there is a NF-Y orthologous in the Clementine genome for each sweet orange NF-Y gene; this was not observed when compared to Arabidopsis thaliana. CsNF-Y proteins shared the same conserved domains with their orthologous proteins in other organisms, including mouse. Analysis of gene expression by RNA-seq and EST data demonstrated that CsNF-Ys have a tissue-specific and stress inducible expression profile. qRT-PCR analysis revealed that CsNF-YA5 exhibits differential expression in response to water deficit in leaves and roots of citrus plants. Overexpression of CsNF-YA5 in transgenic tobacco plants contributed to the reduction of H2O2 production under dehydration conditions and increased plant growth and photosynthetic rate under normal conditions and drought stress. These biochemical and physiological responses to drought stress promoted by CsNF-YA5 may confer a productivity advantage in environments with frequent short-term soil water deficit.
Introduction
Nuclear Factor Y (NF-Y), also known as heme activator protein (HAP) or CCAAT-binding factor (CBF), is a heterotrimeric complex transcription factor composed of three distinct subunits called NF-YA (HAP2), NF-YB (HAP3/CBF-A) and NF-YC (HAP5/CBF-C). NF-Ys are evolutionarily conserved in eukaryotes, with each subunit encoded by a single gene in yeasts and animals, and by gene families comprising between eight and 39 members in plants [1]. The current complex assembly model for NF-Y suggests that NF-YB and C initially form a dimer in the cytoplasm, and subsequently translocated to the cell nucleus where they interact with NF-YA to form a heterotrimer complex [2,3]. NF-YA has a characteristically high affinity to and specificity for the CCAAT box, a cis-acting element present in approximately 25% of eukaryotic gene promoters [4].
NF-Ys have emerged as important regulators of various developmental processes and stress tolerance in plants. Arabidopsis NF-YA genes have been shown to regulate gametogenesis, embryogenesis, seed development and flowering [5–7], while in leguminous plants they have been reported as key regulators of organogenesis and development of symbiotic root nodules [8–11]. NF-YB has been implicated in the regulation of embryogenesis, seed and nodule development [9,12–14], flowering time [15–18], chloroplast biogenesis [19], cell proliferation and endosperm development [20], root elongation [21], photosynthesis [22] and photomorphogenesis [23]. NF-YCs have been likewise found to be involved in the control of flowering time [15,17,18,24], symbiotic nodule development [8,25], root growth [25], photosynthesis [26] and photomorphogenesis [27,28].
The overexpression of the drought-induced and guard cell highly expressed Arabidopsis NF-YA5 (AtNF-YA5) gene improved drought tolerance and reduced leaf water loss, while nfya5 knockout mutants and plants overexpressing the microRNA miR169a showed enhanced leaf water loss and increased sensitive to drought stress in comparison with wild-type (WT) plants [4]. Microarray analysis indicated that AtNF-YA5 plays a role in the induction of expression for a number of stress-responsive genes, such as those involved in oxidative stress responses [4]. Arabidopsis transgenic plants overexpressing the soybean NF-YA3 gene have also shown reduced water loss in leaves and enhanced drought tolerance, with an increased expression of ABA-biosynthesis (ABA1 and ABA2), ABA signaling (ABI1 and ABI2) and stress-responsive (RD29A and CBF3) genes [29]. Transgenic rice plants overexpressing the OsHAP2E gene showed tolerance to drought and salt stresses and resistance to Magnaporthe oryzae and Xanthomonas oryzae infections, as well as higher chlorophyll contents, photosynthetic rates and number of tillers than WT plants [30]. Microarray analysis showed that OsHAP2E up-regulated a number of defense (e.g. chitinase, PBZ1, beta-1,3-glucanase and thaumatin-like protein) and oxidative stress-related (glutathione S-transferase) genes [30]. Overexpression of drought-inducible NF-YB and NF-YC genes also has been shown to improve the drought stress tolerance in Arabidopsis, maize, poplar and rice [31–34].
Citrus is an important genus that includes several cultivated species, such as C. sinensis (sweet oranges), C. reticulata (mandarins and tangerines), C. limon (lemons/limes), C. grandis (pummelos) and C. paradisi (grapefruits). Citrus cultivation is mainly located in semiarid regions of the world, where the trees constantly face problems related to water scarcity and drought stress, which are even more pronounced by the use of saline waters for irrigation. In light of this, efforts have been made to identify drought-responsive genes in citrus for use in rootstock breeding programs aiming to obtain drought-tolerant plants. The availability of the reference genome sequences for C. sinensis and C. clementina [35,36], as well as a rich database (HarvEST: Citrus) of expressed sequence tags (ESTs) for different tissues of several Citrus species and the related genus Poncirus under distinct treatments, now provides the possibility to identify and characterize potential candidate genes for drought response and tolerance in citrus and to accelerate the generation of drought-tolerant citrus rootstocks.
Considering the important role of NF-Ys in mediating drought tolerance in plants, in the present study we have characterized the complete set of citrus NF-Y genes. We further characterized functionally a citrus drought-responsive NF-YA family member, with no close homologs in Arabidopsis, through its overexpression in transgenic tobacco plants.
Materials and methods
Plant material and stress treatments
Two-year-old sweet orange (Citrus sinensis L. Osb. var. ‘Westin’) plants grafted on Rangpur lime (C. limonia Osbeck) were used in the drought stress experiment. The experiment was carried out under greenhouse conditions, with the plants grown in plastic pots of 45L containing a mixture of Oxisol and washed sand (3:1 ratio) and subjected to control (leaf predawn water potential between -0.2 to -0.4 MPa) or drought stress (leaf predawn water potential of -1.5 MPa) treatments, as previously described [37].
The plants of Nicotiana tabacum cv. Havana used in Agrobacterium-mediated genetic transformation experiments, were derived from pre-established cultures kept in the Tissue Culture Laboratory of the Center for Biotechnology and Genetics (CBG) of the State University of Santa Cruz (UESC). The stress tolerance of wild-type (WT) and transgenic tobacco plants was examined by the leaf-disc dehydration tolerance assay, in vitro drought stress tolerance assay and soil dry-down experiment under greenhouse conditions, as previously described [38]. In brief, uniform samples of leaf discs were extracted from mature, fully expanded leaves of adult plants of WT and transgenic (T0 generation) tobacco kept in a greenhouse; this was done with the aid of a 1-cm diameter punch, avoiding the region of the central vein. Leaf discs were then subjected to dehydration at room temperature for 210 min, and fresh weight measurements were performed every 30 min using an analytical scale. The rate of dehydration was determined as the percentage of FW loss versus the initial fresh weight. At the end of the dehydration assays (180 min), leaf discs were subjected to histochemical detection of H2O2 according to the previously described methodology [38]. For the in vitro drought stress tolerance assay, seven-day-old WT and transgenic (T2 generation) seedlings were removed from Murashige and Skoog (MS) germination medium for the WT and MS medium + 50 mg L-1 kanamycin (Sigma, St. Louis, MO, USA) for the transgenic lines and transferred to MS medium only (control treatment) or MS medium containing 15% polyethylene glycol 6000 (PEG-6000) (Merck, Darmstadt, Germany). The FW and root length of individual seedlings was measured 30 days after the treatments. At the end of the experiment, lipid peroxidation was determined by measuring the thiobarbituric acid reactive substance (TBARS) content according to the method described by [37]. For the soil dry-down experiment, 30-day-old WT and transgenic (T1 generation) plants, with an average of 10 to 15 leaves, were first transplanted into 20-liter pots. The pots contained Oxisol and washed sand as substrate in a ratio of 2:1. The plants were grown in greenhouse conditions for four weeks under temperature conditions of 25 ± 4°C and 80–90% humidity. The pots were then sealed with aluminum foil and the plants were subjected to water deficiency by the gradual suspension of the irrigation until they reached leaf predawn water potential of -1.5 MPa, or maintained leaf predawn water potential values of -0.2 to -0.4 MPa under control conditions, by means of daily irrigations. The leaf water potential was measured daily in the third completely expanded leaf from the apex of the plants, using a Scholander type pressure chamber (m670, Pms Instrument Co., Albany, USA). Each WT and transgenic line was represented by five biological replicates per treatment, obtained through micropropagation. The physiological analyzes, including the net CO2 assimilation rate (A), stomatal conductance to water vapor (gs) and the leaf transpiration rate (E) were performed in the morning (08:00–09:00 AM) using a portable photosynthesis system LI-6400 (Li-Cor) equipped with an artificial light resource (6400-02B RedBlue), as previously described [38].
Identification and sequence analysis of citrus NF-Ys
We have queried the sweet orange genome data available at the Phytozome database (http://www.phytozome.org/citrus) with the 30 Arabidopsis thaliana NF-Y protein sequences [39] downloaded from TAIR (http://www.arabidopsis.org), using the TBLASTN tool. Statistically significant alignments were carefully inspected for the presence of the characteristic NF-Y motifs. We have carried out the identification of Citrus clementina NF-Ys by searching its genome data, also available at the Phytozome database (http://phytozome.jgi.doe.gov/pz/portal.html-!info?alias=Org_Cclementina), and using the same strategy as outlined for sweet orange.
Information about coding sequences (CDS), exon-intron structure, full-length sequences and predicted amino acid sequences of the sweet orange NF-Ys was obtained from the Phytozome database. The exon/intron gene structures were constructed using the Exon-Intron Graphic Maker (http://wormweb.org/exonintron). The physical locations of citrus NF-Ys were determined by confirming the starting position of all genes in each chromosome, using BLASTN searching against the local database of the Citrus sinensis Annotation Project (CAP; http://citrus.hzau.edu.cn/orange/). MapChart 2.30 software (https://www.wur.nl/en/show/Mapchart-2.30.htm) was used to plot the gene loci on the sweet orange chromosomes. GRAVY (grand average of hydropathy), molecular weight and isoelectric point (pI) of the deduced amino acid sequences were predicted with the PROTPARAM tool, available on the Expert Protein Analysis System (ExPASy) at the proteomics server (http://www.expasy.ch/tools/protparam.html). The subcellular localization was predicted using the WoLF PSORT tool (http://www.genscript.com/psort/wolf_psort.html). The alignments of predicted amino acid sequences were carried out with the ClustalX program [40] and shaded with BOXSHADE 3.21 software (http://www.ch.embnet.org/software/BOX_form.html). Phylogenetic trees were generated using the Neighbor-Joining method [41] with a bootstrap option of 1000 replications. The dendrograms were constructed by the MEGA6 program [42].
NF-Y proteins were searched against the Protein Data Bank (PDB; http://www.rcsb.org/pdb/) by BLASTP to identify the best template having similar sequence and known three-dimensional structure. The data were run into Phyre2 (Protein Homology/AnalogY Recognition Engine; http://www.sbg.bio.ic.ac.uk/phyre2) for predicting the protein homology modeling under the “intensive” mode. The protein structures of NF-Y were modeled at 90% confidence.
RNA-Seq data were downloaded from the CAP database [43] and used to generate the expression patterns of citrus NF-Ys in different tissues, namely callus, flower, leaf and fruit (flesh tissue). Stress-response expression was inferred from the transcript-per-million (TPM)-converted ratio of the count of CsNF-Y ESTs to the total number of clones present in non-normalized EST libraries (Assembly C52) produced from a range of tissues and stress treatments available in the HarvEST:Citrus 1.32 database (http://harvest-web.org/). The heat maps were generated using the Cluster 3.0 software.
RNA extraction and quantitative real-time RT-PCR (qRT-PCR) analysis
All procedures of RNA extraction, cDNA synthesis and quantitative real-time RT-PCR (qRT-PCR) analysis were performed as previously described [44]. qRT-PCR primers were designed appropriately to avoid the conserved regions. The sequences of the primers used in the qRT-PCR reactions for CsNF-YA5 were either 5'-CATTTCAGAATGGGGAAATCAT-3' and 5'-TTCTTCCTCATCTCACCCAAAG-3' for control and drought-stressed citrus samples, or 5'-TTGCAAGGGCTGGATACCTA-3 ' and 5'-CAACCCCTAGCCCTTCTCAT-3' for the CsNF-YA5 overexpressing transgenic tobacco samples. B-actin (CsACT) or GAPDH (NtGAPDH) genes were amplified together with the target gene (CsNF-YA5) as endogenous controls to normalize expression between the different tissue samples of citrus or tobacco, respectively. The primers CsACT-F (5'-TTAACCCCAAGGCCAACAGA-3'), CsACT-R (5'-TCCCTCATAGATTGGTACAGTATGGAC-3'), NtGAPDHRT-F (5'-TCAAACCCTTCCACCAACTC-3') and NtGAPDHRT (5’-CTAATCGCCCAATTCTTCCA-3') were used in the qRT-PCR reactions. The expression values of the genes were calculated by means of the 2-ΔΔCt method [45]. Confirmation of the amplification of specific products was performed by analyzing the dissociation curves with the Mx3005P qPCR analysis software. Data were obtained from a pool of three biological replicates that were individually validated.
Cloning of CsNF-YA5 and generation of transgenic tobacco plants
The complete coding sequence of CsNF-YA5 was obtained from its amplification in cDNA samples of roots of Rangpur lime (C. limonia Osb.) subjected to water deficit, using the primers 5'-CAAGATGGGATTAATGGACAAGAAC-3 ' and 5'-CCCTCCAACGTCAAGGAACT-3'. The amplified cDNA fragment was inserted into the pGEM-T vector (Promega, USA) by means of the TA cloning system and used in transformation of Escherichia coli, strain TOP 10. Subsequently, the amplified fragment was taken from pGEM-T by means of digestion with the restriction enzyme NotI, and subcloned in sense orientation at the same restriction site of the plasmid pUC118/CaMV 35S, which contains the promoter and terminator sequences of the 35S gene of cauliflower mosaic virus (CaMV 35S). The expression cassette was then excised from pUC118/CaMV 35S, by means of digestion with HindIII and SalI, and inserted into the same restriction sites of pCAMBIA 2301 vector (Cambia, Australia). This vector contains the reporter gene uidA and the selective nptII gene under the control of the constitutive promoter CaMV 35S. The recombinant vector was then introduced into the Agrobacterium tumefaciens strain of EHA-105 and used in genetic transformation experiments of N. tabacum.
The experiments of genetic transformation of tobacco were carried out using the methodology described by Cidade et al. [46]. In brief, leaf segments of ~1 cm2 were excised and placed in contact with bacterial suspension (OD600 = 0.5), for 15 minutes at room temperature and under gentle shaking. The excised explants were subjected to (i) bacterial suspension coculture containing the pCAMBIA 2301 vector with the CsNF-YA5 gene, (ii) bacterial suspension culture containing pCAMBIA 2301 without the insertion of the gene of interest and (iii) non-cocultivated explants with Agrobacterium as control of the experiment. After the development of the aerial parts, the plants were multiplicated by micropropagation, in which the aerial parts were excised with the use of sterile forceps and scalpel and inoculated in MS medium plus 300 mg mL-1 timentin. At least five biological replicates (clones) were obtained for each WT and transgenic lines with empty vector (PC) or overexpressing CsNF-YA5 (NF12, NF15, NF16, NF20 and NF22). The plants were submitted to the acclimation phase, where they remained for one week in distilled and autoclaved water, and covered with plastic bags with cavity at the ends. Afterwards, the plants were transferred to plastic vessels with a capacity of 5-L, containing autoclaved soil substrate and sand in a ratio of 2:1, where they remained for three weeks in a growth room with controlled temperature and luminosity. Screening by PCR and histochemical GUS assays were used to identify the transgenic plants, according to the methodologies described, respectively, by Cidade et al. [46] and Jefferson et al. [47].
Statistical analysis
The statistical analysis was carried out with the SISVAR and BioEstat softwares which tested the experiments as a completely randomized design. Statistical differences were assessed based on the analysis of variance (ANOVA) and averages were separated by means of the Student’s t-test, with a critical value of P≤ 0.05, P≤ 0.01 and P≤ 0.001. The leaf-disc dehydration tolerance assay was composed by three plant replicates per treatment, with five leaf discs per replication. Regression lines were fitted by the method of least squares and the significance of the coefficient of determination (r2) was verified by the F-test. The in vitro drought stress tolerance assay contained five replicate plates per treatment composed of fifteen seedlings for each line. The soil dry-down experiment consisted of five plants per treatment, with each plant considered as an experimental unit.
Results
The citrus NF-Y gene family
BLAST searches using the 30 amino acid sequences of the complete set of A. thaliana NF-Y proteins as query sequences resulted in the identification of a total of 22 different Citrus species NF-Y encoding protein genes (CsNF-Ys) in the sweet orange genome, including six CsNF-YAs, 11 CsNF-YBs and five CsNF-YCs (Table 1). BLAST searches against the genome sequence of Clementine mandarin using the same strategy as outlined for sweet oranges revealed that both Citrus species contain exactly the same number of NF-Y genes (Table 1). Phylogenetic analysis showed that there is a NF-Y ortholog in the genome of Clementine mandarin for each sweet orange NF-Y gene, while two CsNF-YAs (CsNF-YA5 and CsNF-YA6), four CsNF-YBs (CsNF-YB1, CsNF-YB2, CsNF-YB5 and CsNF-YB11) and one CsNF-YC (CsNF-YC4) have no close homologs in A. thaliana (S1–S3 Figs and Table 1).
Table 1. Citrus NF-Y transcription factor encoded genes.
| Name | Locus (Phytozome) | Locus (CAP) | C. clementina ortholog | A. thaliana ortholog | Chromosome location | Polypeptide length (MW) | pI | GRAVY | Predicted subcellular location |
|---|---|---|---|---|---|---|---|---|---|
| CsNF-YA1 | orange1.1g019036m | Cs2g30350 | Ciclev10015808m | NF-YA1 NF-YA9 | chr2: 29,712,980..29,717,207 | 347 (37.2kDa) | 8.26 | -0.964 | nucleus |
| CsNF-YA2 | orange1.1g021081m | Cs6g13560 | Ciclev10012232m | NF-YA2 NF-YA10 | chr6: 14,987,576..14,992,571 | 317 (34.6kDa) | 9.27 | -0.597 | nucleus |
| CsNF-YA3 | orange1.1g019782m | Cs1g17780 | Ciclev10026020m | NF-YA3 NF-YA8 NF-YA5 NF-YA6 | chr1: 20,916,460..20,920,299 | 336 (36.7kDa) | 8.67 | -0.617 | nucleus |
| CsNF-YA4 | orange1.1g026474m | Cs2g09780 | Ciclev10016491m | NF-YA4 NF-YA7 | chr2: 7,234,224..7,238,903 | 238 (26.5kDa) | 9.15 | -0.733 | extracellu-lar |
| CsNF-YA5 | orange1.1g019764m | Cs7g01720 | Ciclev10032207m | - | chr7: 461,288..464,375 | 336 (36.8kDa) | 9.54 | -0.749 | nucleus |
| CsNF-YA6 | orange1.1g017825m | Cs9g12370 | Ciclev10005144m | - | chr9: 10,757,085..10,763,538 | 365 (39.9kDa) | 9.44 | -0.723 | nucleus |
| CsNF-YB1 | orange1.1g045289m | orange1.1t03346 | Ciclev10027508m | - | chrUn: 51,410,005..51,410,973 | 71 (8.0kDa) | 4.79 | -0.58 | chloroplast |
| CsNF-YB2 | orange1.1g027605m | orange1.1t03346 | Ciclev10027508m | - | chrUn: 51,410,005..51,410,973 | 221 (24.1kDa) | 6.33 | -0.725 | nucleus |
| CsNF-YB3 | orange1.1g036580m | Cs4g08780 | Ciclev10009623m | NF-YB3 NF-YB2 | chr4: 5,787,274..5,788,494 | 186 (20.2kDa) | 6.53 | -0.788 | nucleus |
| CsNF-YB4 | orange1.1g045194m | - | Ciclev10003157m | NF-YB4 | - | 157 (17.6kDa) | 8.37 | -0.947 | nucleus |
| CsNF-YB5 | orange1.1g044287m | Cs9g04610 | Ciclev10006769m | - | chr9: 2,565,158..2,565,691 | 188 (20.9kDa) | 5.43 | -0.84 | nucleus |
| CsNF-YB6 | orange1.1g026469m | Cs2g01680 | Ciclev10016498m | NF-YB6 | chr2: 563,303..566,657 | 238 (26.5kDa) | 6.72 | -0.668 | mitochon-drion |
| CsNF-YB7 | orange1.1g047516m | Cs1g09850 | Ciclev10003558m | NF-YB7 | chr1: 11,604,325..11,604,969 | 214 (24.1kDa) | 5.80 | -1.014 | nucleus |
| CsNF-YB8 | orange1.1g030647m | Cs6g04240 | Ciclev10012920m | NF-YB8 NF-YB10 | chr6: 4,871,183..4,873,746 | 174 (19.0kDa) | 5.67 | -0.833 | nucleus |
| CsNF-YB9 | orange1.1g038325m | Cs4g08720 | Ciclev10010391m | NF-YB9 | chr4: 5,742,982..5,745,273 | 231 (25.4kDa) | 6.05 | -0.542 | nucleus |
| CsNF-YB10 | orange1.1g030547m | Cs7g27760 | Ciclev10032893m | NF-YB8 NF-YB10 | chr7: 28,445,326..28,456,064 | 175 (18.9kDa) | 5.81 | -0.685 | nucleus |
| CsNF-YB11 | orange1.1g038014m | Cs5g32730 | Ciclev10024294m | - | chr5: 33,945,582..33,946,004 | 140 (15.6kDa) | 5.62 | -0.805 | nucleus |
| CsNF-YC1 | orange1.1g026901m | orange1.1t02576 | Ciclev10032646m | NF-YC1 NF-YC4 | chrUn: 39,384,571..39,387,608 | 231 (25.0kDa) | 5.05 | -0.455 | nucleus |
| CsNF-YC2 | orange1.1g024265m | orange1.1t03257 | Ciclev10007003m | NF-YC2 | chrUn: 50,081,198..50,085,450 | 270 (30.2kDa) | 5.81 | -0.644 | nucleus |
| CsNF-YC3 | orange1.1g019814m | Cs3g13490 | Ciclev10002193m | NF-YC3 NF-YC9 | chr3: 17,935,028..17,937,988 | 335 (37.3kDa) | 9.24 | -0.641 | nucleus |
| CsNF-YC4 | orange1.1g047870m | Cs5g03650 | Ciclev10023545m | - | chr5: 1,967,345..1,967,909 | 102 (11.2kDa) | 9.61 | -0.034 | chloroplast |
| CsNF-YC5 | orange1.1g045847m | Cs6g18220 | Ciclev10013435m | NF-YC12 | chr6: 18,422,446..18,425,136 | 237 (27.1kDa) | 5.24 | -0.526 | nucleus |
Most CsNF-Y genes were precisely mapped on the sweet orange chromosomes by homology searches against the sweet orange genome assembly available at the CAP database. Some CsNF-Ys could not be located on any chromosome (ChrUN) because of an incomplete physical map for sweet orange. CsNF-Y loci were distributed throughout all the sweet orange chromosomes, except for the chromosome 8, at densities varying from one to three genes per chromosome (S4 Fig and Table 1). Two pairs of paralogous CsNF-Y genes were detected based on both phylogenetic analysis and chromosomal location. One of these pairs, CsNF-YB8/10 (82.2% similarity), was putative segmental duplicates according to criteria of Gu et al. [48]: the length of aligned sequence covers >80% of the longer gene and the similarity of the aligned region is >70%. The other gene pair, CsNF-YB3/9 (~40 kb proximity to each other), was putative tandem duplicates according to the criteria of Hanada et al. [49]: tandem duplicates are genes in any gene pair that belong to the same gene family and are located within 100 kb each other.
Analysis of the exon-intron structure of all CsNF-Y genes was carried out using the sweet orange gene models annotated in Phytozome (S5 Fig). In general, there were similarities in the number and length of the exons, but not of the introns, among CsNF-Ys and their putative orthologs from Arabidopsis (AtNF-Ys). All CsNF-YA genes were interrupted by three to four introns in their coding regions. Besides, most CsNF-YA genes contained introns embedded in their 5’- and/or 3’-UTR. No intron was present in 6 members from the CsNF-YB family, while the other members showed variations from 1 to 5 introns in their coding regions. Two CsNF-YB members, CsNF-YB8/10, contained introns in their 5’-UTR. There were 3 CsNF-YC members (CsNF-YC1/2/4) containing no or only one intron, while the others contained 2 or 6 introns in their coding regions. Two CsNF-YC members, CsNF-YC2/3, also contained introns in their 5’- and/or 3’-UTR. Interestingly, the pair of paralogous genes showed some variations in their exon-intron structures, such as different number of introns (CsNF-YB8/10) and length of exons (CsNF-YB3/9). These data suggest that the organization of CsNF-Y genes is dynamic and rapidly evolving.
The CsNF-Y genes encode proteins ranging from 71 (8 kDa) to 365 (39.9 kDa) amino acids in length, and pI values ranging from 4.79 to 9.54 (Table 1). The average protein length of CsNF-YAs, CsNF-YBs and CsNF-YCs were 323 (35.3 kDa), 176.5 (19.5 kDa) and 202.2 (22.5 kDa) amino acids respectively, while their average pI were, respectively, 9.05, 6.01 and 7.07. All the CsNF-Y proteins had a negative GRAVY score, indicating that they are hydrophilic proteins (Table 1). Most CsNF-Ys were predicted to be located in the nucleus, with some of them also targeted to chloroplast and mitochondrion (Table 1).
Multiple alignments showed that CsNF-Y proteins contain the evolutionarily conserved domains that are responsible for heterodimerization, heterotrimerization and DNA interactions at the CCAAT sites (Fig 1). The conserved core region of CsNF-YA is ~53 amino acids in length and composed of two sub-domains, one mediating the NF-YB/C interaction and the other responsible for CCAAT binding (Fig 1A). These sub-domains are separated from each other by a relatively conserved linker. The CsNF-YB subunit core region is ~88 amino acids in length and contains the NFYC interaction domain that extends across two independent regions and partly overlaps with the DNA-binding and NF-YA interaction domains (Fig 1B). This region has structural similarities with the histone-fold motif (HFM) of the core histone H2B [50]. The CsNF-YC subunit core region is ~79 amino acids long and possesses the highly conserved domains for NF-YA and NF-YB interactions, with the former extending across two separate regions (Fig 1C). This region is also characterized by an HFM, but that is more similar to the core histone H2A than H2B [50]. The secondary structures of the conserved core regions of NF-Y proteins were predicted to be composed of α-helices and coils, using the protein homology modeling of Phyr2 (Fig 1D). The NF-YA conserved region was comprised of two α-helices, one in each sub-domain. The NF-YB and NF-YC conserved regions were comprised of four α-helices that are located within all the functionally important domains for NF-Y interaction and DNA binding.
Fig 1. Multiple alignments and predicted structure of conserved regions of CsNF-Y family members.

Sequence alignments among the highly conserved domains of NF-YA (A), NF-YB (B) and NF-YC (C) proteins of C. sinensis (Cs) and Mus musculus (Mouse). The DNA binding domain and the domains required for interaction with the other subunits previously defined in yeast and mammals are indicated. The secondary structures, alpha-helices (rectangles) and coils (black lines), are represented on the bottom of the alignment, based on Romier et al. [55]. Predicted structures of NF-Y conserved regions using the protein homology modeling of Phyr2 (D). Images of the models were coloured in a rainbow from blue to red from the N-terminus to the C-terminus.
Expression analysis of CsNF-Y genes in different tissues and in response to abiotic and biotic stresses
Expression analysis from RNA-seq data showed that, except for CsNF-YB4, all the CsNF-Y genes are preferentially expressed in one of the major sweet orange tissues (Fig 2). We have observed four main clusters of expression. Cluster 1 corresponded to CsNF-Y genes highly expressed in callus, while those from cluster 2 included CsNF-Ys with high expression in fruit. Cluster 3 and Cluster 4 included highly expressed CsNF-Y genes in leaf and flower, respectively. Expression analysis from EST data demonstrated that, except for CsNF-YC4, all the CsNF-Y genes were also expressed in response to one or more stress treatments (Fig 3A). Major clusters of expression responses were associated with the drought, nematode, Citrus tristeza virus (CTV) and X. fastidiosa treatments. Drought induced the expression of one CsNF-YA (CsNF-YA5) and five CsNF-YBs (CsNF-YB1/2/4/5/11) in roots. All these genes, plus CsNF-YA1, were also expressed in roots in response to nematode treatment. CTV induced the expression of four CsNF-YAs (CsNF-YA1/2/4/6), two CsNF-YBs (CsNF-YB7/10) and one CsNF-YC (CsNF-YC1) in barks, and three CsNF-YCs (CsNF-YC2/3/5) in leaves. X. fastidiosa induced the expression of one CsNF-YA (CsNF-YA3) and three CsNF-YBs (CsNF-YB3/6/9) in leaves. Taken together, these data indicated that CsNF-Ys exhibit a tissue-specific and abiotic and/or biotic stress-inducible expression pattern.
Fig 2. Heatmap of expression of CsNF-Ys in different tissues of sweet orange.

The color scale shown represents RPKM-normalized log2-transformed counts.
Fig 3. Expression analysis of CsNF-Ys.
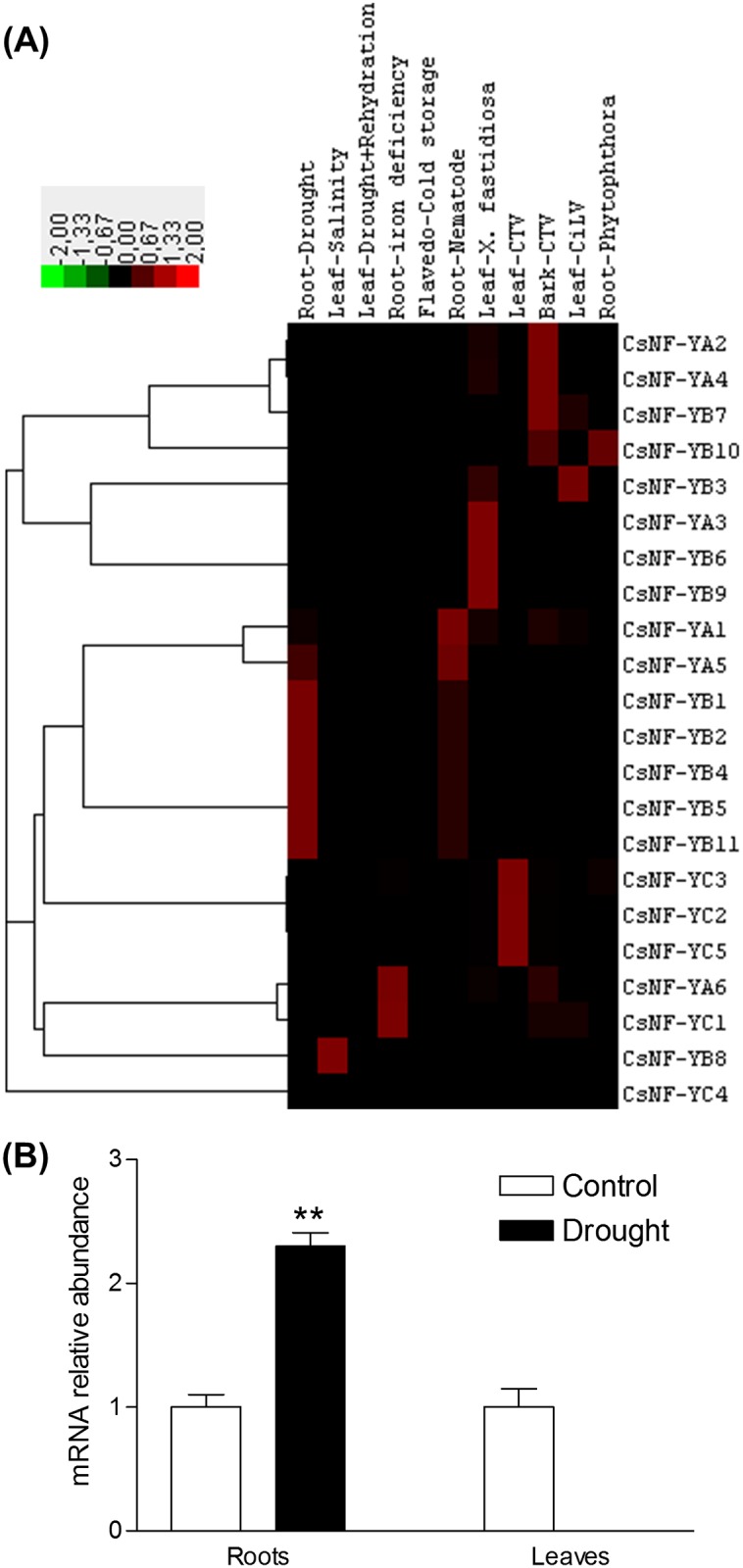
Heatmap of expression of CsNF-Ys in response to different stress treatments (A). The color scale shown represents TPM-normalized log2-transformed counts. qRT-PCR expression analysis of CsNF-YA5 (B). CsNF-YA5 mRNA abundance in leaves and roots of sweet orange plants grafted on Rangpur lime, subjected to control (irrigated) and drought treatments. The data are means ± SE of three biological replicates in which β-actin transcripts were used as internal controls.**Significantly different from control treatment at P ≤ 0.01.
CsNF-YA5 expression analysis in response to drought stress
Our comprehensive analysis has provided a good starting point for the functional characterization of citrus NF-Ys. It helped us to choose CsNF-YA5 as the first citrus NF-Y family member to be functionally characterized because it was the only CsNF-YA gene to be induced by drought treatment. Besides, CsNF-YA5 has no close homologs in Arabidopsis, representing therefore a novel NF-YA gene. It shares the highest amino acid sequence similarity with the predicted nuclear transcription factor Y subunit A-4 from Theobroma cacao (XP_007052521.2; 65% similarity). qRT-PCR expression analysis revealed distinct expression patterns of CsNF-YA5 in leaves and roots of drought-stressed citrus plants (Fig 3B). A significant induction of the expression of CsNF-YA5 (2.5-×) was observed in roots of plants subjected to drought treatment, when compared to that of the control treatment. On the other hand, the expression of CsNF-YA5 was completely repressed in leaves of plants exposed to drought treatment.
CsNF-YA5 overexpression in transgenic tobacco plants
The complete coding sequence of CsNF-YA5 was amplified from root cDNA samples of Rangpur lime (S6 Fig), inserted into pCAMBIA 2301 under the control of the constitutive promoter CaMV 35S and used in the Agrobacterium-mediated genetic transformation of tobacco. Several shoots representing distinct transformation events were obtained from the genetic transformation experiments (S7 Fig). Five independent transgenic lines were analyzed by PCR, all of which showed amplification of a fragment compatible with 800-bp of nptII gene (S8A Fig). Histochemical staining analysis of GUS was also used to confirm the transgenic nature of the plants (S8B Fig). The expression profile of CsNF-YA5 in mature leaves of WT and transgenic lines was examined by qRT-PCR analysis (S8C Fig). All the analyzed CsNF-YA5-overexpressing transgenic lines presented higher levels (2.5–4.0-×) of CsNF-YA5 expression when compared to the WT and empty vector (PC) control lines.
Dehydration tolerance assay of CsNF-YA5-overexpressing tobacco plants
Leaf discs of control (WT and PC) and CsNF-YA5-overexpressing transgenic (NF12, NF15, NF16, NF20, NF22) lines were subjected to dehydration stress for 210 min. The time course of water loss rate showed that the leaf discs of NF16 and NF20 dehydrated slower, NF15 and NF22 faster, and NF12 similarly than those of the WT and PC control plants (Fig 4A).
Fig 4. Leaf-disc dehydration tolerance assay.

Dehydration rate in leaf discs of control (NT and PC) and CsNF-YA5-overexpressing transgenic (NF12, NF15, NF16, NF20, NF22) lines for 210 min at room temperature (A). Measurement by fresh weight reduction were carried out every 30 min. Regression lines were fitted to the data using least-squares regression analysis. Each point represents the mean of three plant replicates per treatment, with five leaf discs per replication. In situ detection of H2O2 in tobacco leaf discs treated with DAB (B). Representative photographs showing staining of H2O2 in leaf disks of WT and transgenic lines with empty vector (PC) or overexpressing CsNF-YA5 (NF12, NF15, NF16, NF20, NF22) after 180 min dehydration. WT leaf discs before dehydration (WT+H2O2) and treated with H2O2 [WT(dehydrated)+H2O2] or only water (WT+H2O), after 180 min dehydration, were used as controls.
H2O2 production was analyzed in leaf discs of tobacco plants subjected to 180 min of dehydration using the DAB-HCL histochemical staining analysis. According to the results, four (NF12, NF16, NF20 and NF22) out of the five tested CsNF-YA5-overexpressing transgenic lines showed significantly clearer staining than those of the control plants (WT and PC), indicating a lower H2O2 production (Fig 4B). NF15 presented a more intense coloration and similar to that of the control plants. H2O2 production was lower in non-dehydrated and H2O2-treated WT than in the WT subjected to dehydration and treated with H2O2. We have not observed, as expected, darkening of the leaf discs of WT treated with water instead of DAB, when compared to the leaf discs immersed in DAB solution (Fig 4B).
In vitro drought stress tolerance assay of CsNF-YA5-overexpressing tobacco plants
In order to evaluate the effects of the CsNF-YA5 overexpression on drought stress tolerance of the transgenic lines at the level of whole plant, control and transgenic plants were subjected to the in vitro drought stress tolerance assay. The representative phenotypes of WT and transgenic plants under control and drought treatments are shown in Fig 5A and 5B. In general, seedlings from the transgenic lines tended to have higher biomass (Fig 5C) and root length (Fig 5D) than the WT under both control and PEG treatment. Analysis of the TBARS content showed that the transgenic line NF16 had a lower index of lipid peroxidation than the WT under PEG treatment (Fig 5E).
Fig 5. In vitro drought stress tolerance assay of CsNFYA5-overexpressing transgenic lines.

Representative phenotypes of control (WT and PC) and CsNFYA5 transgenic (NF12, NF15, NF16, NF20 and NF22) lines grown under control and PEG treatments for 30 days (A, B). Shown from left to right in panel B: WT, PC, NF12, NF15, NF16, NF20 and NF22. Seedling biomass and root length, respectively, of control and transgenic lines under control and PEG treatments for 30 days (C, D). The data are means ± SE of five technical replicates composed of fifteen seedlings for each line. Total thiobarbituric acid reactive substances (TBARS) concentration in seedlings of WT and transgenic line NF16 under control and PEG treatments for 30 days (E). The data are means ± SE of six technical replicates composed of fifteen seedlings for each line. Statistically significant differences at P ≤ 0.01 (**) or P ≤ 0.05 (*) between control (WT) and CsNFYA5 transgenic lines, at the respective treatment, are indicated.
Physiological analysis of CsNF-YA5-overexpressing tobacco plants
To understand how the water deficit influences the responses of the CsNF-YA5-overexpressing transgenic lines at the physiological level, control and transgenic lines were subjected to the soil dry-down experiment under greenhouse conditions. Analysis of the net assimilation rate of CO2 (A), stomatal conductance to water vapor (gs) and the leaf transpiration rate (E) was carried out in plants under control (irrigated) and drought conditions. All the CsNF-YA5-overexpressing transgenic lines showed values of A, gs and E significantly higher than the control lines (WT and PC) in both irrigated and drought treatments (Fig 6). Another interesting observation was that, in contrast to the control lines, three (NF15, NF16 and NF22) out of the five CsNF-YA5-overexpressing transgenic lines exhibited higher rates of A, gs and E in the drought than in the irrigated treatment.
Fig 6. Physiological analysis of CsNFYA5-overexpressing transgenic lines.
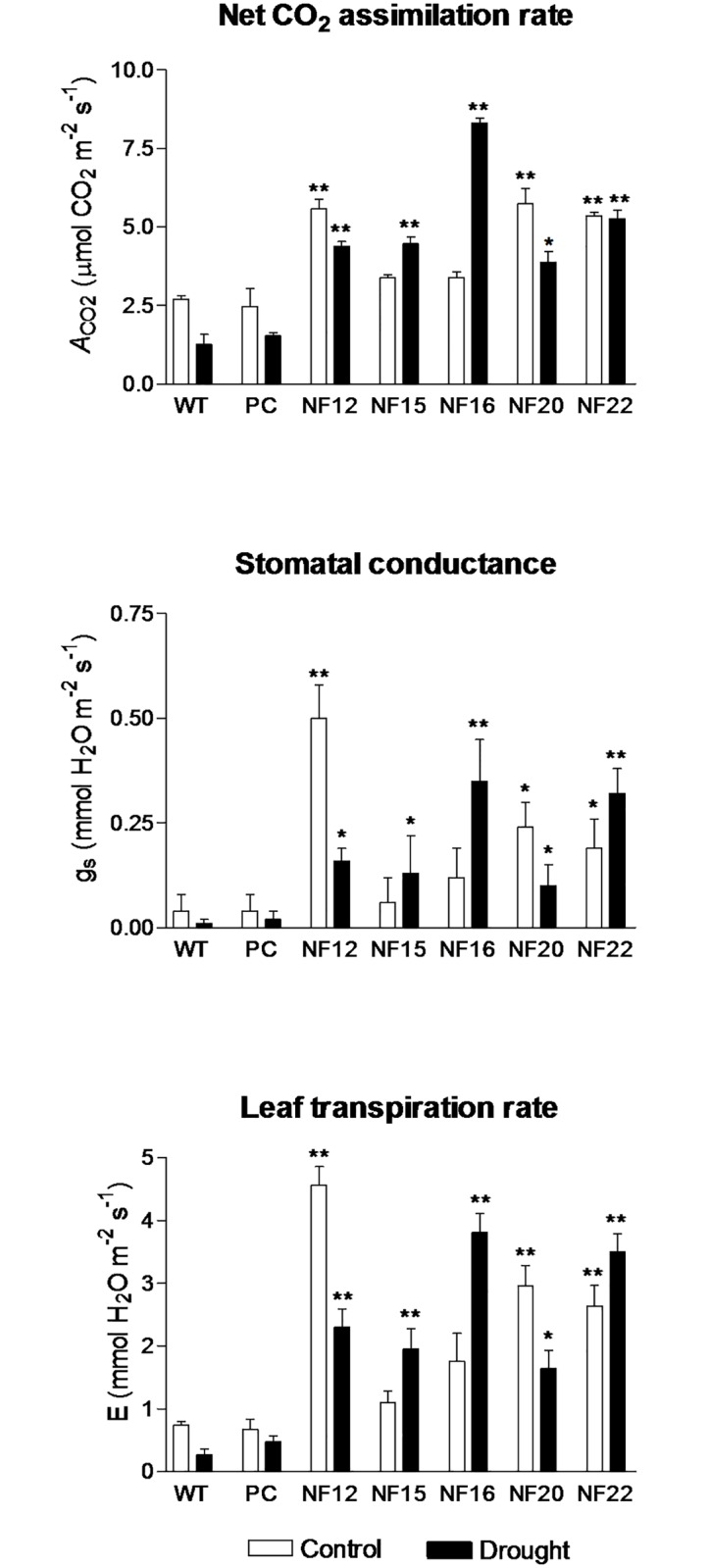
Control (WT and PC) and CsNFYA5 transgenic (NF12, NF15, NF16, NF20 and NF22) lines were subjected to control (irrigated) and drought treatments under greenhouse conditions. The data are means ± SE of five plant replicates per treatment for each line. Statistically significant differences at P ≤ 0.01 (**) or P ≤ 0.05 (*) between control (WT) and CsNFYA5 transgenic lines, at the respective water treatment, are indicated.
Discussion
The number of genes identified for each citrus NF-Y subunit (Table 1) is lower than that reported for A. thaliana [39]. In light of this, the presence of six CsNF-YAs, 11 CsNF-YBs and five CsNF-YCs in citrus could result in the formation of only 330 unique trimetric complexes, a relatively small number when compared to 1,000 unique complexes that could be theoretically combined in A. thaliana (10 NF-YA, 10 NF-YB and 10 NF-YC subunits). However, the number of potential unique NF-Y complexes has been shown to be much smaller than previously anticipated, due to the specificity of expression and interactions among the different NF-Y subunits [1]. The lower number of NF-Y genes found in the citrus genome may be accounted for by the fact that A. thaliana has undergone two additional rounds of recent whole-genome duplication (WGD) when compared to citrus [35]. By the other hand, the lack of close homologs in A. thaliana for some citrus NF-Ys (S1–S3 Figs) suggests that more recent gene duplication events have contributed to the expansion of the family in citrus. The results of phylogenetic analyses (S1–S3 Figs) and chromosome mapping (S4 Fig) suggest that both segmental (e.g. CsNF-YB8 and CsNF-YB10) and tandem (e.g. CsNF-YB3 and CsNF-YB9) duplications have contributed to the CsNF-Y gene expansion in the citrus genome.
Gene-structure analysis of the CsNF-Y gene family revealed that CsNF-YAs have a more complex exon-intron organization than most CsNF-YBs and CsNF-YCs (S5 Fig), consistently with the previously reported organization of NF-Y genes in Arabidopsis and Brassica napus [51], Phaseolus vulgaris [52] and Solanum lycopersicum [53]. Notably, most CsNF-YAs and some CsNF-YBs and CsNF-YCs contained one intron in their 5’- and/or 3’-UTR. In Medicago trunculata, retention of the 5’-UTR intron in alternative spliced variants of NF-YA1 leads to the synthesis of a small peptide that destabilizes the mRNA [54]. In M. trunculata, A. thaliana and Oryza sativa, most NF-YAs have at least one intron in the 5’-UTR [1], suggesting that this post-transcriptional regulatory mechanism might be conserved among plant NF-YA genes. NF-YA transcripts are also regulated at post-transcriptional level by the action of microRNA169 (miR169) that binds to the 3’-UTR and promotes their cleavage by the slicing protein Argonaute 1 [4,8].
Multiple alignments showed that CsNF-Y proteins have the conserved regions involved in protein interaction and DNA binding (Fig 1), which are highly similar from mammals (such as mouse) to plants [50,55]. This high degree of conservation strongly suggests the maintenance of ancestral functions related to the complex formation. From secondary structure prediction analysis, the NF-YA conserved region was divided into an α-helical N-terminal region involved in NF-YB/C interactions and a small coiled C-terminal region involved in DNA binding [55]. For DNA binding, the α1 and α2 of NF-YB and the α1 of NF-YC are also required [50]. The NF-YB helix α2 and NF-YC helices α1 and αC were shown to influence NF-YA binding [50,55]. The NF-YB and NF-YC α3s have been implicated in interactions with TBP, the TATA box-binding subunit of the general initiation factor TFIID [50]. The NF-YC αC region is a target element for gene-specific regulatory proteins, such as SP1, SREBP1, RF-X and C/EBP [55].
Orthology prediction is an important tool for transferring functional information between two species. CsNF-YA1 is orthologous to AtNF-YA1 and AtNF-YA9 (S1 Fig and Table 1) which have been involved, respectively, in flowering time control [15] and fertility [56]. CsNF-YA2 is orthologous to AtNF-YA2 and AtNF-YA10 that have been implicated in the control of root growth and architecture [57]. CsNF-YA3 is orthologous to the AtNF-YA3, which is involved in nitrogen nutrition [58]. CsNF-YA4 is orthologous to the AtNF-YA4 that has been shown to participate in the ER-stress and unfolded protein response (UPR) [3]. The known Arabidopsis regulators of photoperiod dependent flowering time, NF-YB2/3 and NF-YC3/9 [15–18,24], have highly significant orthology matches to CsNF-YB3 and CsNF-YC3, respectively (S2 and S3 Figs and Table 1). Therefore, it is expected that most of these conserved citrus NF-Ys subunits are involved in similar developmental processes and stress responses as reported in A. thaliana.
Expression analysis from RNA-seq data revealed that CsNF-Ys exhibit a tissue-specific expression pattern (Fig 2), as previously reported for NF-Ys of A. thaliana [39], Brachypodium distachyon [59], B. napus [51], Setaria italica [60], Glycine max [61], P. vulgaris [52] and S. lycopersicum [53]. Such specific expression profiles indicate a sub-functionalization of the subunit members and provide certain clues about the potential heterotrimeric NF-Y complexes that may be combined in specific tissues. For example, CsNF-YA3/5, CsNF-YB6/7/9 and CsNF-YC5 could form potential NF-Y complexes in tissue of callus, while CsNF-YA2, CsNF-YB5/11 and CsNF-YC2/3 could do it in fruit tissue. Functional analysis using yeast two- or three-hybrid assays will help to clarify whether such interactions might be detected in vitro. Most CsNF-Y genes were also expressed in response to one or more stress treatments (Fig 3A), suggesting their involvement in the regulation of biotic and/or abiotic stress response. Those subunit members that were co-expressed in the same stress treatments may likewise represent potential NF-Y complexes regulating specific stress responses, such as CsNF-YA5 and CsNF-YB1/2/4/5/11 in drought and CsNF-YA3 and CsNF-YB3/6/9 in X. fastidiosa treatment. Interestingly, CsNF-YA3 is an ortholog of the rice OsHAP2E gene, whose expression was shown to be induced after inoculation with Magnaporthe oryzae or Xanthomonas oryzae pv. oryzae and whose overexpression in transgenic rice resulted in enhanced resistance against these fungal and bacterial pathogens, respectively [30]. The similar expression profiles exhibited by the paralogous CsNF-YB genes in the different tissues (CsNF-YB3/9 and CsNF-YB8/10; Fig 2) or stress treatments (CsNF-YB3/9; Fig 3A) suggest functional redundancy among these genes.
At phenotypic level, it has been well documented that sweet orange scions grafted on Rangpur lime typically exhibit impaired photosynthesis and enhanced root growth under water deficit treatment [62,63]. When these previous findings are considered together with our expression data (Fig 3B), in which CsNFY-A5 is preferentially expressed in callus, a tissue showing high cell division rates, and up-regulated in roots of Rangpur but down-regulated in leaves of sweet orange by drought stress, we can assume its potential role as a candidate gene in the drought-induced regulation of the processes of root growth and photosynthesis. Although a number of NF-YB and NF-YC proteins have been identified as regulators of root growth [21,25,32] and photosynthesis [19,26,31,32] in different plant species, emerging evidences also supporting the role of NF-YA members in these processes have been reported more recently [30,57].
The overexpression of CsNF-YA5 exerted distinct effects on the dehydration tolerance in leaf discs of transgenic tobacco, depending on the transgenic line (Fig 4A). These variations may be presumptively associated with the varying CsNF-YA5 expression levels among the different transgenic lines (S8C Fig). It is possible that CsNF-YA5 has both activating and inhibitory effects on dehydration tolerance, depending on the expression levels of the protein or interacting proteins, or on expression of other intracellular and extracellular factors that have not yet been identified in the present study. On the other hand, the most obvious effects of CsNF-YA5 overexpression were the inhibition of H2O2 production in dehydrated leaf discs as revealed by DAB staining (Fig 4B), improved biomass and root length as revealed by the in vitro drought stress tolerance assay (Fig 5A–5D), and decreased lipid peroxidation under drought treatment as revealed by TBARS analysis (Fig 5E). Such observations suggest a role of CsNF-YA5 in plant growth and activation of the antioxidant system. Connections between NF-YAs and antioxidant defenses have been previously shown. Overexpression of the AtNF-YA5 improved drought resistance in part by activating target antioxidant gene products, such as glutathione S-transferase, peroxidases and an oxidoreductase [4]. Overexpression of the Setaria italica SiNF-YA1 enhanced drought and salt tolerance in transgenic tobacco by increasing the activity of superoxide dismutase (SOD), peroxidase (POD) and catalase (CAT) [60]. In addition, transient luciferase (LUC) expression assays showed that SiNF-YA1 is able to activate the LUC gene driven by the tobacco CAT promoter [60].
Overexpression of CsNF-YA5 significantly increased A, gs and E rates (Fig 6), which are important physiological determinants in increasing yields. A significant increase in net photosynthetic rate was also reported in transgenic rice lines overexpressing the OsHAP2E gene, which was correlated with the higher number of tillers and grain yield exhibited by the transgenic plants [30]. Studies have shown that NF-Ys exert a positive regulation on photosynthesis by inducing the expression of several genes related to this process, such as those coding for the small subunit of ribulose-1,5-bisphosphate carboxylase (RBCS) and the light-harvesting chlorophyll a/b-binding protein of photosystem II (CAB) [19], chloroplast ATP synthase [22,64], glutamyl-tRNA reductase (GluTR) and light-harvesting chlorophyll a/b-binding proteins associated with photosystem I (LHCA4 and LHCB4) [22].
In conclusion, the present study demonstrates that citrus NF-Y comprises a relatively small but diverse family of proteins involved in developmental processes and stress response. Functional analysis of CsNFYA5 in transgenic tobacco plants evidenced its involvement in biochemical and physiological mechanisms of adaptation to drought that can contribute to the maintenance of plant growth and productivity in environments with short, frequent and moderate soil water deficit periods alternating with wet periods. The information presented here offers a useful foundation for functional studies of CsNF-Ys and supports the potential application of CsNFYA5 in the improvement of drought tolerance in citrus via biotechnological strategies.
Supporting information
The amino acid sequences were aligned using ClustalX and the Neighbor-Joining method. The tree was built with a bootstrap support of 1000 replications. Numbers at internal nodes denotes the results of bootstrapping analysis (n = 1000).
(TIF)
The amino acid sequences were aligned using ClustalX and the Neighbor-Joining method. The tree was built with a bootstrap of 1,000 replications. Numbers at internal nodes denotes the results of bootstrapping analysis (n = 1000).
(TIF)
The amino acid sequences were aligned using ClustalX and the Neighbor-Joining method. The tree was built with a bootstrap of 1,000 replications. Numbers at internal nodes denotes the results of bootstrapping analysis (n = 1000).
(TIF)
The chromosomal position of each CsNF-Y gene was mapped according to the Citrus sinensis Annotation Project (CAP). The CsNF-YB8/10 pair of segmentally duplicated genes and CsNF-YB3/9 pair of tandemly duplicated genes are indicated. The scale is in Mb.
(TIF)
Open boxes correspond to 5’ and 3’ untranslated regions (UTR) and exons and introns are represented by filled boxes and black lines, respectively. The sizes of 5’ and 3’ UTRs, exons and introns can be estimated using the reference scale bar of 100 bp.
(TIF)
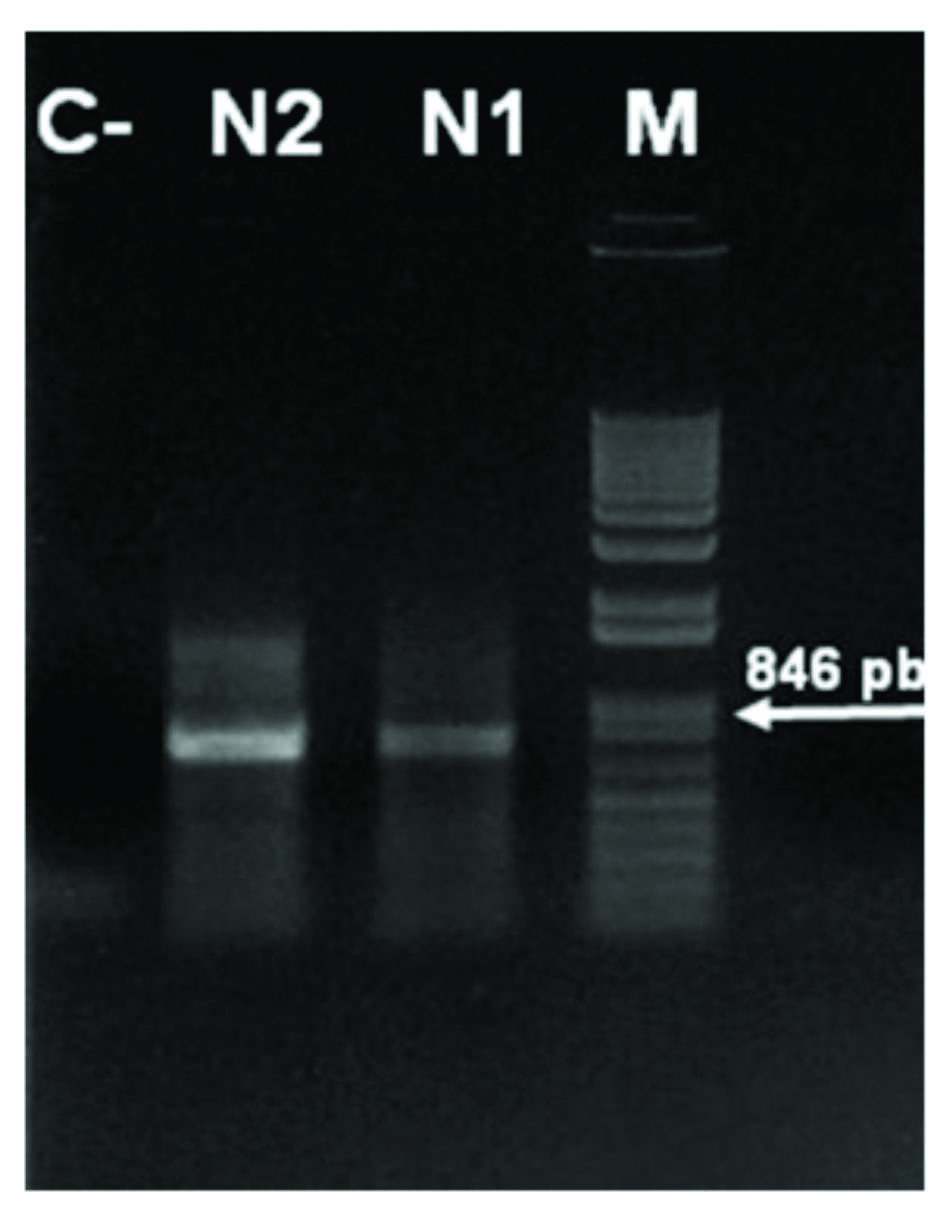
M: 1-kb molecular weight marker; C-: negative control (reaction without template cDNA); C+: positive control (reaction with plasmid DNA); N1 and N2: amplification product in Rangpur lime cDNA containing the expected size of approximately 846-bp.
(JPG)
{kind=link}
Explants of tobacco in MS medium (A). Initiation of the shoot formation in MS medium supplemented with BAP (5.0 mg L-1) and the antibiotics kanamycin (50 mg.L-1) and timentin (300 mg L-1) (B, C). Plants developing in MS medium supplemented with kanamycin (50 mg L-1) and timentin (300 mg L-1) (D). Individualized plants with the presence of roots (E, F).
(JPG)
{kind=link}
Amplification of the nptII gene fragment in transgenic tobacco plants by PCR (A). M: 1-kb marker; C-: negative control (reaction without template DNA); C+: positive control (reaction containing plasmid DNA from pCAMBIA 2301); PC: transgenic line transformed with pCAMBIA 2301 empty vector; NT: non-transformed WT plants; N12-22: CsNF-YA5-overexpressing transgenic lines. PC DNA fragment shifted to an apparently higher molecular size due to the use of GelRed™ to stain the DNA. Histochemical assay for uidA gene expression in transgenic tobacco seedlings (B). qRT-PCR expression analysis of CsNF-YA5 in leaves of control (WT and PC) and CsNFYA5-overexpressing transgenic tobacco lines (C). The data are means ± SE of three biological replicates in which β-actin (citrus) or GAP2C (tobacco) transcripts were used as internal controls. *, **Significantly different from WT at P ≤ 0.05 and P ≤ 0.01, respectively.
(TIF)
Acknowledgments
We appreciate the technical assistance provided by Ms. Junea L. Nascimento.
Data Availability
The Citrus NF-Y genes (CsNF-Ys) were derived from the published genome sequences of C. sinensis (https://phytozome.jgi.doe.gov/pz/portal.html#!info?alias=Org_Csinensis) and C. clementina (https://phytozome.jgi.doe.gov/pz/portal.html#!info?alias=Org_Cclementina), and the accession IDs of all sequences used in the present study are listed in Table 1.
Funding Statement
This work was supported by research grants from CNPq (Process # 478733/2013-5), Instituto Nacional de Ciência e Tecnologia (INCT) de Genômica para Melhoramento de Citros (CNPq Process # 465440/2014-2 and FAPESP Process # 2008/2014/50880-0) and Embrapa (Macroprograma 2). S.L.S. Pereira was supported by a CAPES Foundation Masters Degree scholarship. D.S. Camargo and L.R. Camillo were supported by a CAPES/PNPD postdoctoral fellowship. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Laloum T, De Mita S, Gamas P, Baudin M, Niebel A. CCAAT-box binding transcription factors in plants: Y so many? Trends Plant Sci. 2013;18: 157–166. doi: 10.1016/j.tplants.2012.07.004 [DOI] [PubMed] [Google Scholar]
- 2.Steidl S, Tüncher A, Goda H, Guder C, Papadopoulou N, Kobayashi T, et al. A single subunit of a heterotrimeric CCAAT-binding complex carries a nuclear localization signal: piggy back transport of the pre-assembled complex to the nucleus. J Mol Biol. 2004;342: 515–524. doi: 10.1016/j.jmb.2004.07.011 [DOI] [PubMed] [Google Scholar]
- 3.Liu JX, Howell SH. bZIP28 and NF-Y transcription factors are activated by ER stress and assemble into a transcriptional complex to regulate stress response genes in Arabidopsis. Plant Cell. 2010;22: 782–796. doi: 10.1105/tpc.109.072173 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Li WX, Oono Y, Zhu J, He XJ, Wu JM, Iida K, et al. The Arabidopsis NFYA5 transcription factor is regulated transcriptionally and posttranscriptionally to promote drought resistance. Plant Cell. 2008;20: 2238–2251. doi: 10.1105/tpc.108.059444 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Fornari M, Calvenzani V, Masiero S, Tonelli C, Petroni K. The Arabidopsis NF-YA3 and NF-YA8 genes are functionally redundant and are required in early embryogenesis. PloS One. 2013;8: 82043 doi: 10.1371/journal.pone.0082043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Mu J, Tan H, Hong S, Liang Y, Zuo J. Arabidopsis transcription factor genes NF-YA1, 5, 6, and 9 play redundant roles in male gametogenesis, embryogenesis, and seed development. Mol Plant. 2013;6: 188–201. doi: 10.1093/mp/sss061 [DOI] [PubMed] [Google Scholar]
- 7.Siriwardana CL, Gnesutta N, Kumimoto RW, Jones DS, Myers ZA, Mantovani R, et al. NUCLEAR FACTOR Y, Subunit A (NF-YA) proteins positively regulate flowering and act through FLOWERING LOCUS T. PLoS Genet. 2016;12: e1006496 doi: 10.1371/journal.pgen.1006496 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Combier JP, Frugier F, De Billy F, Boualem A, El-Yahyaoui F, Moreau S, et al. MtHAP2-1 is a key transcriptional regulator of symbiotic nodule development regulated by microRNA169 in Medicago truncatula. Genes Dev. 2006;20: 3084–3088. doi: 10.1101/gad.402806 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Soyano T, Kouchi H, Hirota A, Hayashi M. Nodule inception directly targets NF-Y subunit genes to regulate essential processes of root nodule development in Lotus japonicus. PLoS Genet. 2013;9: 1003352 doi: 10.1371/journal.pgen.1003352 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Laporte P, Lepage A, Fournier J, Catrice O, Moreau S, Jardinaud MF, et al. The CCAAT box-binding transcription factor NF-YA1 controls rhizobial infection. J Exp Bot. 2013;65: 481–494. doi: 10.1093/jxb/ert392 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Laloum T, Baudin M, Frances L, Lepage A, Billault-Penneteau B, Cerri MR, et al. Two CCAAT-box-binding transcription factors redundantly regulate early steps of the legume-rhizobia endosymbiosis. Plant J. 2014;79: 757–768. doi: 10.1111/tpj.12587 [DOI] [PubMed] [Google Scholar]
- 12.West MA, Yee KM, Danao J, Zimmerman JL, Fischer RL, Goldberg RB, et al. LEAFY COTYLEDON1 is an essential regulator of late embryogenesis and cotyledon identity in Arabidopsis. Plant Cell. 1994;6: 1731–1745. doi: 10.1105/tpc.6.12.1731 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Parcy F, Valon C, Kohara A, Miséra S, Giraudat J. The ABSCISIC ACID-INSENSITIVE3, FUSCA3, and LEAFY COTYLEDON1 loci act in concert to control multiple aspects of Arabidopsis seed development. Plant Cell. 1997;9: 1265–1277. doi: 10.1105/tpc.9.8.1265 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kwong RW, Bui AQ, Lee H, Kwong LW, Fischer RL, Goldberg RB, et al. LEAFY COTYLEDON1-LIKE defines a class of regulators essential for embryo development. Plant Cell. 2003;15: 5–18. doi: 10.1105/tpc.006973 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wenkel S, Turck F, Singer K, Gissot L, Le Gourrierec J, Samach A, et al. CONSTANS and the CCAAT box binding complex share a functionally important domain and interact to regulate flowering of Arabidopsis. Plant Cell. 2006;18: 2971–2984. doi: 10.1105/tpc.106.043299 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kumimoto RW, Adam L, Hymus GJ, Repetti PP, Reuber TL, Marion CM, et al. The Nuclear Factor Y subunits NF-YB2 and NF-YB3 play additive roles in the promotion of flowering by inductive long-day photoperiods in Arabidopsis. Planta. 2008;228: 709–723. doi: 10.1007/s00425-008-0773-6 [DOI] [PubMed] [Google Scholar]
- 17.Cao S, Kumimoto RW, Gnesutta N, Calogero AM, Mantovani R, Holt BF. A distal CCAAT/NUCLEAR FACTOR Y complex promotes chromatin looping at the FLOWERING LOCUS T promoter and regulates the timing of flowering in Arabidopsis. Plant Cell. 2014;26: 1009–1017. doi: 10.1105/tpc.113.120352 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gnesutta N, Kumimoto RW, Swain S, Chiara M, Siriwardana C, Horner DS, et al. CONSTANS imparts DNA sequence specificity to the histone fold NF-YB/NF-YC dimer. Plant Cell. 2017;29: 1516–1532. doi: 10.1105/tpc.16.00864 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Miyoshi K, Ito Y, Serizawa A, Kurata N. OsHAP3 genes regulate chloroplast biogenesis in rice. Plant J. 2003;36: 532–540. [DOI] [PubMed] [Google Scholar]
- 20.Sun X, Ling S, Lu Z, Ouyang YD, Liu S, Yao J. OsNF-YB1, a rice endosperm-specific gene, is essential for cell proliferation in endosperm development. Gene. 2014;551: 214–221. doi: 10.1016/j.gene.2014.08.059 [DOI] [PubMed] [Google Scholar]
- 21.Ballif J, Endo S, Kotani M, MacAdama J, Wu Y. Over-expression of HAP3b enhances primary root elongation. Plant Physiol Biochem. 2011;49: 579–583. doi: 10.1016/j.plaphy.2011.01.013 [DOI] [PubMed] [Google Scholar]
- 22.Stephenson TJ, McIntyre CL, Collet C, Xue GP. TaNF-YB3 is involved in the regulation of photosynthesis genes in Triticum aestivum. Funct Integr Genomics. 2011;11: 327–340. doi: 10.1007/s10142-011-0212-9 [DOI] [PubMed] [Google Scholar]
- 23.Huang M, Hu Y, Liu X, Li Y, Hou X. Arabidopsis LEAFY COTYLEDON1 mediates postembryonic development via interacting with PHYTOCHROME-INTERACTING FACTOR4. Plant Cell. 2015;27: 3099–3111. doi: 10.1105/tpc.15.00750 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ben-Naim O, Eshed R, Parnis A, Teper-Bamnolker P, Shalit A, Coupland G, et al. The CCAAT binding factor can mediate interactions between CONSTANS-like proteins and DNA. Plant J. 2006;46: 462–476. doi: 10.1111/j.1365-313X.2006.02706.x [DOI] [PubMed] [Google Scholar]
- 25.Battaglia M, Rípodas C, Clúa J, Baudin M, Aguilar OM, Niebel A, et al. A nuclear factor Y interacting protein of the GRAS family is required for nodule organogenesis, infection thread progression, and lateral root growth. Plant Physiol. 2014;164: 1430–1442. doi: 10.1104/pp.113.230896 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Stephenson TJ, McIntyre CL, Collet C, Xue GP. TaNF-YC11, one of the light-upregulated NF-YC members in Triticum aestivum, is co-regulated with photosynthesis-related genes. Funct Integr Genomics. 2010;10: 265–276. doi: 10.1007/s10142-010-0158-3 [DOI] [PubMed] [Google Scholar]
- 27.Myers ZA, Kumimoto RW, Siriwardana CL, Gayler KK, Risinger JR, Pezzetta D, et al. NUCLEAR FACTOR Y, Subunit C (NF-YC) transcription factors are positive regulators of photomorphogenesis in Arabidopsis thaliana. PLoS Genet. 2016;12: e1006333 doi: 10.1371/journal.pgen.1006333 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Tang Y, Liu X, Liu X, Li Y, Wu K, Hou X. Arabidopsis NF-YCs mediate the light-controlled hypocotyl elongation via modulating histone acetylation. Mol Plant. 2017;10: 260–273. doi: 10.1016/j.molp.2016.11.007 [DOI] [PubMed] [Google Scholar]
- 29.Ni Z, Hu Z, Jiang Q, Zhang H. GmNFYA3, a target gene of miR169, is a positive regulator of plant tolerance to drought stress. Plant Mol Biol. 2013;1: 113–129. doi: 10.1007/s11103-013-0040-5 [DOI] [PubMed] [Google Scholar]
- 30.Alam M, Tanaka T, Nakamura H, Ichikawa H, Kobayashi K, Yaeno T, et al. Overexpression of a rice heme activator protein gene (OsHAP2E) confers resistance to pathogens, salinity and drought, and increases photosynthesis and tiller number. Plant Biotechnol J. 2015;13: 85–96. doi: 10.1111/pbi.12239 [DOI] [PubMed] [Google Scholar]
- 31.Nelson DE, Repetti PP, Adams TR, Creelman RA, Wu J, Warner DC, et al. Plant nuclear factor Y (NF-Y) B subunits confer drought tolerance and lead to improved corn yields on water-limited acres. Proc Natl Acad Sci USA. 2007;104: 16450–16455. doi: 10.1073/pnas.0707193104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Han X, Tang S, An Y, Zheng DC, Xia XL, Yin WL. Overexpression of the poplar NF-YB7 transcription factor confers drought tolerance and improves water-use efficiency in Arabidopsis. J Exp Bot. 2013;64: 4589–4601. doi: 10.1093/jxb/ert262 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Chen M, Zhao Y, Zhuo C, Lu S, Guo Z. Overexpression of a NF-YC transcription factor from bermudagrass confers tolerance to drought and salinity in transgenic rice. Plant Biotechnol J. 2015;13: 482–491. doi: 10.1111/pbi.12270 [DOI] [PubMed] [Google Scholar]
- 34.Zhang T, Zhang D, Liu Y, Luo C, Zhou Y, Zhang L. Overexpression of a NF-YB3 transcription factor from Picea wilsonii confers tolerance to salinity and drought stress in transformed Arabidopsis thaliana. Plant Physiol Biochem. 2015;94: 153–164. doi: 10.1016/j.plaphy.2015.05.001 [DOI] [PubMed] [Google Scholar]
- 35.Xu Q, Chen LL, Ruan X, Chen D, Zhu A, Chen C, et al. The draft genome of sweet orange (Citrus sinensis). Nature Genet. 2013;45: 59–66. doi: 10.1038/ng.2472 [DOI] [PubMed] [Google Scholar]
- 36.Wu GA, Prochnik S, Jenkins J, Salse J, Hellsten U, Murat F, et al. Sequencing of diverse mandarin, pummelo and orange genomes reveals complex history of admixture during citrus domestication. Nature Biotechnol. 2014;32: 656–662. doi: 10.1038/nbt.2906 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Gonçalves LP, Alves TF, Martins CP, Sousa AO, Santos IC, Pirovani CP, et al. Rootstock-induced physiological and biochemical mechanisms of drought tolerance in sweet orange. Acta Physiol Plant. 2016;38: 1–12. doi: 10.1007/s11738-016-2198-3 [Google Scholar]
- 38.Martins CPS, Neves DM, Cidade LC, Mendes AFS, Silva DC, Almeida AAF, et al. Expression of the citrus CsTIP2;1 gene improves tobacco plant growth, antioxidant capacity and physiological adaptation under stress conditions. Planta. 2017;245: 951–963. doi: 10.1007/s00425-017-2653-4 [DOI] [PubMed] [Google Scholar]
- 39.Petroni K, Kumimoto RW, Gnesutta N, Calvenzani V, Fornari M, Tonelli C, Holt BF, Mantovani R. The promiscuous life of plant NUCLEAR FACTOR Y transcription factors. Plant Cell. 2012;24: 4777–4792. doi: 10.1105/tpc.112.105734 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Thompson JD, Higgens DG, Gibson TJ. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;22: 4673–4680. doi: 10.1093/nar/22.22.4673 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Saitou N, Nei M. The Neighbor-Joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4: 406–425. doi: 10.1093/oxfordjournals.molbev.a040454 [DOI] [PubMed] [Google Scholar]
- 42.Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 2013;30: 2725–2729. doi: 10.1093/molbev/mst197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Wang J, Chen D, Lei Y, Chang JW, Hao BH, Xing F, et al. Citrus sinensis Annotation Project (CAP): A comprehensive database for sweet orange genome. PloS One. 2014;9: e87723 doi: 10.1371/journal.pone.0087723 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Martins CPS, Pedrosa AM, Du D, Gonçalves LP, Yu Q, Gmitter FG Jr, et al. Genome-wide characterization and expression analysis of major intrinsic proteins during abiotic and biotic stresses in sweet orange (Citrus sinensis L. Osb.). PloS One. 2015;10: e0138786 doi: 10.1371/journal.pone.0138786 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods. 2001;25: 402–408. doi: 10.1006/meth.2001.1262 [DOI] [PubMed] [Google Scholar]
- 46.Cidade LC, de Oliveira TM, Mendes AF, Macedo AF, Floh EI, Gesteira AS, et al. Ectopic expression of a fruit phytoene synthase from Citrus paradisi Macf. promotes abiotic stress tolerance in transgenic tobacco. Mol Biol Rep. 2012;39: 10201–10209. doi: 10.1007/s11033-012-1895-2 [DOI] [PubMed] [Google Scholar]
- 47.Jefferson RA, Kavanagh TA, Bevan MW. GUS fusions: beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987;6: 3901–3907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Gu Z, Cavalcanti A, Chen F-C, Bouman P, Li W-H. Extent of gene duplication in the genomes of Drosophila, nematode, and yeast. Mol Biol Evol. 2002;19: 256–262. doi: 10.1093/oxfordjournals.molbev.a004079 [DOI] [PubMed] [Google Scholar]
- 49.Hanada K, Zou C, Lehti-Shiu MD, Shinozaki K, Shiu SH. Importance of lineage specific expansion of plant tandem duplicates in the adaptive response to environmental stimuli. Plant Physiol. 2008;148: 993–1003. doi: 10.1104/pp.108.122457 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Mantovani R. The molecular biology of the CCAAT-binding factor NF-Y. Gene. 1999;239: 15–27. doi: 10.1016/S0378-1119(99)00368-6 [DOI] [PubMed] [Google Scholar]
- 51.Liang M, Yin X, Lin Z, Zheng Q, Liu G, Zhao G. Identification and characterization of NF-Y transcription factor families in Canola (Brassica napus L.). Planta. 2014;239: 107–126. doi: 10.1007/s00425-013-1964-3 [DOI] [PubMed] [Google Scholar]
- 52.Rípodas C, Castaingts M, Clúa J, Blanco F, Zanetti ME. Annotation, phylogeny and expression analysis of the nuclear factor Y gene families in common bean (Phaseolus vulgaris). Front Plant Sci. 2015;5: 761 doi: 10.3389/fpls.2014.00761 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Li S, Li K, Ju Z, Cao D, Fu D, Zhu H, et al. Genome-wide analysis of tomato NF-Y factors and their role in fruit ripening. BMC Genomics. 2016;17: 36 doi: 10.1186/s12864-015-2334-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Combier JP, De Billy F, Gamas P, Niebel A, Rivas S. Trans-regulation of the expression of the transcription factor MtHAP2-1 by a uORF controls root nodule development. Genes Dev. 2008;22: 1549–1559. doi: 10.1101/gad.461808 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Romier C, Cocchiarella F, Mantovani R, Moras D. The NFYB/NF-YC structure gives insight into DNA binding and transcription regulation by CCAAT factor NF-Y. J Biol Chem. 2003;278: 1336–1345. doi: 10.1074/jbc.M209635200 [DOI] [PubMed] [Google Scholar]
- 56.Levesque-Lemay M, Albani D, Aldcorn D, Hammerlindl J, Keller W, Robert LS. Expression of CCAAT-binding factor antisense transcripts in reproductive tissues affects plant fertility. Plant Cell Rep. 2003;21: 804–808. [DOI] [PubMed] [Google Scholar]
- 57.Sorin C, Declerck M, Christ A, Blein T, Ma L, Lelandais-Brière C, et al. A miR169 isoform regulates specific NF-YA targets and root architecture in Arabidopsis. New Phytol. 2014;202: 1197–1211. doi: 10.1111/nph.12735 [DOI] [PubMed] [Google Scholar]
- 58.Zhao M, Ding H, Zhu JK, Zhang F, Li WX. Involvement of miR169 in the nitrogen-starvation responses in Arabidopsis. New Phytol. 2011;190: 906–915. doi: 10.1111/j.1469-8137.2011.03647.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Cao S, Kumimoto RW, Siriwardana CL, Risinger JR, Holt BF 3rd. Identification and characterization of NF-Y transcription factor families in the monocot model plant Brachypodium distachyon. PLoS One. 2011;6: e21805 doi: 10.1371/journal.pone.0021805 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Feng ZJ, He GH, Zheng WJ, Lu PP, Chen M, Gong YM, et al. Foxtail millet NF-Y families: genome-wide survey and evolution analyses identified two functional genes important in abiotic stresses. Front Plant Sci. 2015;6: 1142 doi: 10.3389/fpls.2015.01142 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Quach TN, Nguyen HT, Valliyodan B, Joshi T, Xu D, Nguyen HT. Genome-wide expression analysis of soybean NF-Y genes reveals potential function in development and drought response. Mol Genet Genomics. 2015;290: 1095–1115. doi: 10.1007/s00438-014-0978-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Erismann NM, Machado EC, Tucci MLS. Photosynthetic limitation by CO2 diffusion in drought stressed orange leaves on three rootstocks. Photosynth Res. 2008;96: 163–172. doi: 10.1007/s11120-008-9297-y [DOI] [PubMed] [Google Scholar]
- 63.Pedroso FKJV, Prudente DA, Bueno ACR, Machado EC, Ribeiro RV. Drought tolerance in citrus trees is enhanced by rootstock-dependent changes in root growth and carbohydrate availability. Environ Exp Bot. 2014;101: 26–35. doi: 10.1016/j.envexpbot.2013.12.024 [Google Scholar]
- 64.Kusnetsov V, Landsberger M, Meurer J, Oelmüller R. The assembly of the CAAT-box binding complex at a photosynthesis gene promoter is regulated by light, cytokinin, and the stage of the plastids. J Biol Chem. 1999;274: 36009–36014. doi: 10.1074/jbc.274.50.36009 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The amino acid sequences were aligned using ClustalX and the Neighbor-Joining method. The tree was built with a bootstrap support of 1000 replications. Numbers at internal nodes denotes the results of bootstrapping analysis (n = 1000).
(TIF)
The amino acid sequences were aligned using ClustalX and the Neighbor-Joining method. The tree was built with a bootstrap of 1,000 replications. Numbers at internal nodes denotes the results of bootstrapping analysis (n = 1000).
(TIF)
The amino acid sequences were aligned using ClustalX and the Neighbor-Joining method. The tree was built with a bootstrap of 1,000 replications. Numbers at internal nodes denotes the results of bootstrapping analysis (n = 1000).
(TIF)
The chromosomal position of each CsNF-Y gene was mapped according to the Citrus sinensis Annotation Project (CAP). The CsNF-YB8/10 pair of segmentally duplicated genes and CsNF-YB3/9 pair of tandemly duplicated genes are indicated. The scale is in Mb.
(TIF)
Open boxes correspond to 5’ and 3’ untranslated regions (UTR) and exons and introns are represented by filled boxes and black lines, respectively. The sizes of 5’ and 3’ UTRs, exons and introns can be estimated using the reference scale bar of 100 bp.
(TIF)
M: 1-kb molecular weight marker; C-: negative control (reaction without template cDNA); C+: positive control (reaction with plasmid DNA); N1 and N2: amplification product in Rangpur lime cDNA containing the expected size of approximately 846-bp.
(JPG)
Explants of tobacco in MS medium (A). Initiation of the shoot formation in MS medium supplemented with BAP (5.0 mg L-1) and the antibiotics kanamycin (50 mg.L-1) and timentin (300 mg L-1) (B, C). Plants developing in MS medium supplemented with kanamycin (50 mg L-1) and timentin (300 mg L-1) (D). Individualized plants with the presence of roots (E, F).
(JPG)
Amplification of the nptII gene fragment in transgenic tobacco plants by PCR (A). M: 1-kb marker; C-: negative control (reaction without template DNA); C+: positive control (reaction containing plasmid DNA from pCAMBIA 2301); PC: transgenic line transformed with pCAMBIA 2301 empty vector; NT: non-transformed WT plants; N12-22: CsNF-YA5-overexpressing transgenic lines. PC DNA fragment shifted to an apparently higher molecular size due to the use of GelRed™ to stain the DNA. Histochemical assay for uidA gene expression in transgenic tobacco seedlings (B). qRT-PCR expression analysis of CsNF-YA5 in leaves of control (WT and PC) and CsNFYA5-overexpressing transgenic tobacco lines (C). The data are means ± SE of three biological replicates in which β-actin (citrus) or GAP2C (tobacco) transcripts were used as internal controls. *, **Significantly different from WT at P ≤ 0.05 and P ≤ 0.01, respectively.
(TIF)
Data Availability Statement
The Citrus NF-Y genes (CsNF-Ys) were derived from the published genome sequences of C. sinensis (https://phytozome.jgi.doe.gov/pz/portal.html#!info?alias=Org_Csinensis) and C. clementina (https://phytozome.jgi.doe.gov/pz/portal.html#!info?alias=Org_Cclementina), and the accession IDs of all sequences used in the present study are listed in Table 1.