

Abstract
Background
Nasopharyngeal carcinoma is a rare form of cancer across the world except in certain areas such as Southern China, Hong Kong and Malaysia. NPC is considered a relatively radiosensitive tumor and patients diagnosed at early stages tend to survive longer compared to those with advanced disease. Given that early symptoms of NPC are non-specific and that the nasopharynx is relatively inaccessible, less invasive screening methods such as biomarker screening might be the key to improve NPC survival and management. A number of genes with their respective polymorphisms have been shown in past studies to be associated with survival of various cancers. hOGG1 and XPD genes encode for a DNA glycosylase and a DNA helicase respectively; both are proteins that are involved in DNA repair. ITGA2 is the alpha subunit of the transmembrane receptor integrin and is mainly responsible for cell-cell and cell-extracellular matrix interaction. TNF-α is a cytokine that is released by immune cells during inflammation.
Methods
Restriction fragment length polymorphism-polymerase chain reaction (RFLP-PCR) was used to genotype all the aforementioned gene polymorphisms. Kaplan-Meier survival function, log-rank test and Cox regression were used to investigate the effect of gene polymorphisms on the all-cause survival of NPC.
Results
NPC cases carrying T/T genotype of ITGA2 C807T have poorer all-cause survival compared to those with C/C genotypes, with an adjusted HR of 2.06 (95% CI = 1.14–3.72) in individual model. The 5-year survival rate of C/C carriers was 55% compared to those with C/T and T/T where the survival rates were 50% and 43%, respectively.
Conclusion
The finding from the present study showed that ITGA2 C807T polymorphism could be potentially useful as a prognostic biomarker for NPC. However, the prognostic value of ITGA2 C807T polymorphism has to be validated by well-designed further studies with larger patient numbers.
Introduction
Nasopharyngeal carcinoma (NPC) is the commonest malignancy that originates from the nasopharynx. NPC is regarded as a rare form of cancer with varying disease prevalence across populations. The annual incidence of NPC in Southeast Asia [1], Southern China [2] and Hong Kong [3] is very high compared to the rest of the world. According to the International Agency for Research on Cancer, 84,400 NPC cases were newly diagnosed in 2008 with 51,600 deaths worldwide [4]. In Malaysia, NPC is the 4th most common cancer [5]. In comparison with other carcinomas, NPC remains a relatively radiosensitive tumor [6]. Despite this, 20–30% of NPC patients still experience distant metastasis after combined intensity-modulated radiotherapy and chemotherapy [7]. Currently, tumor, node, metastasis (TNM) classification is mainly used for determination of prognosis and treatment strategy in NPC [8]. However, TNM staging is inadequate in the prognostic evaluation due to the heterogeneity of NPC, in which patients at the same TNM stage undergoing similar treatment modalities often experience different clinical outcomes [9]. Hence, development of a new prognostic marker is imperative for the improvement of the current management of patients diagnosed with NPC.
Integrins are transmembrane glycoprotein heterodimers that are made up from α- and β-subunits. So far 18 α- and 8 β-subunits have been discovered in mammals and they combine to form at least 25 different heterodimers that are specific for a unique set of ligands [10]. Integrins mainly serve as cell surface adhesive receptors that mediate cell-cell and cell-extracellular matrix interactions [11]. From previous research, integrins are shown to be involved in the regulation of cell proliferation, migration and survival [12–14]. Coupled with the downstream effect resulting from integrin signaling, integrin contributes to tumor progression through mediation of angiogenesis, lymphangiogenesis and inflammation in the tumor microenvironment [15–16]. Integrin α2 (ITGA2) from the α-family is a collagen receptor mainly expressed on platelets and epithelial cells [17]. ITGA2 807C>T (rs1126643) is one of the extensively studied polymorphisms of ITGA2. The expression of α2β1 integrin on the platelet surface has been reported to be lower in subjects carrying 807C compared to their 807T counterpart [18]. ITGA2 807C>T polymorphism was also found to be associated with the risk of developing breast and colorectal cancer [19–20].
Tumor necrosis factor alpha (TNF-α) is one of the earliest known cytokine produced in inflammatory process [21]. High circulating plasma level of TNF-α in patients with malignant lymphomas has been associated with poor disease outcome [22]. Excessive production is linked to weight loss, cachexia, anemia and modification of immune response reducing patients’ tolerance to their disease and treatment [23]. The TNF-α gene is located within class III region of major histocompatibility complex (MHC) [21]. A polymorphism located at position -308 that involves a base change of guanine to adenine was found to influence TNF expression in vitro [24]. Using human B-cell lines in transfection studies, the presence of TNF-α 308A allele was shown to correlate positively with higher inducible levels of TNF expression compared to 308G allele, indicating an important transcriptional regulation site is present in this region [25]. A allele of TNF-α -308G>A polymorphism was found in several studies to be associated with a decrease in survival of various cancers, namely B-cell lymphoma [26], non-Hodgkin lymphoma [27] and chronic lymphocytic lymphoma [28].
DNA repair is one of the important events in human cells that are responsible for maintaining the integrity of the human genome. DNA in human cells is constantly exposed to various endogenous and exogenous carcinogens from the surrounding environment [29]. Polycyclic aromatic hydrocarbons and nitrosamines are examples of exogenous carcinogens human cells are exposed to from ingestion of salted fish and cigarette smoking [30–31]. Normal cellular metabolic processes in the human body produce endogenous carcinogens such as hydroxyl radicals that can cause oxidative damage to DNA [32]. Accumulation of DNA damage, if unrepaired, leads to neoplastic transformation of normal cells [33]. Base excision repair (BER) and nucleotide excision repair (NER) are DNA repair pathways that are responsible of removing carcinogenic metabolites such as 8-oxo-7,8-dihydroguanine (8-oxoG) and bulky DNA adducts respectively [34–35]. Human 8-oxoguanine DNA N-glycosylase I (hOGG1) is the primary enzyme in BER pathway that recognizes 8-oxoG in the damaged DNA site [36]. Besides acting as a glycosylase, hOGG1 protein also function as an apurinic/apyrimidinic (AP) lyase that excises the damaged base from the DNA [37]. A polymorphism in codon 326 of hOGG1 protein has been shown to confer a higher risk of cancers, namely hepatocellular [38], breast [39] and orolaryngeal [40]. Similarly, hOGG1 Ser326Cys polymorphism was found to be significantly associated with survival of lung [41], bladder [42] and pancreatic cancer [43].
Xeroderma pigmentosum group D (XPD) gene on the other hand encodes for a 5’-3’ DNA helicase, which is an essential component in NER pathway [44]. XPD is responsible for unwinding the double helix DNA at the damaged site [45]. NER cannot proceed to the subsequent step of repair without the proper unwinding of DNA [46]. XPD Lys751Gln polymorphism is located within the carboxy-terminal domain (CTD) of XPD protein and is one of the established polymorphisms of the XPD gene [47]. Significant associations have been reported between XPD Lys751Gln polymorphism and several cancers, namely chronic myeloid leukemia [48] and esophageal [49] and nasopharyngeal carcinoma [50]. Studies on head and neck [51], lung [52], breast [53] and colorectal [54] cancers have shown significant association between XPD Lys751Gln polymorphism and survival.
In present study, we investigated the effect of gene polymorphisms on the overall and all-cause survival of NPC patients in Malaysia.
Materials and methods
Ethics statement and study design
This study was conducted with the approval from the Medical Research Ethics Committees of the Ministry of Health, Malaysia (NMRR-08-1572-3115). The present study was funded by Science Fund, Ministry of Science, Technology and Innovation (MOSTI; Project code: 04-11-08-625FR).
Patient selection
NPC cases were selected from the NPC clinic registries in the Departments of Radiology and Oncology of two public hospitals starting in 2011. To minimize the incidence-prevalence bias [55], newly-diagnosed cases and cases diagnosed within two years from the time they were screened, were recruited. The inclusion criteria for NPC cases were NPC patients who were at least 18 years old and who have been confirmed histologically. Controls were age, sex and ethnicity matched to cases. Written informed consent was obtained from eligible cases prior to the enrollment into the study. The information sheet and consent form were available in both English and Malay. We assumed the exposure rate of ITGA2 C807T polymorphism at 49% from Chen et al [56], with two-sided alpha level of 0.05, 256 NPC cases were needed to attain a power of 90% to detect a 50% increase in hazard rate [57]. For the investigation of the all-cause survival, 300 NPC cases were available in the analysis.
Endpoints
The primary endpoint was all-cause survival, defined as an estimate of the probability of surviving all-causes of death in a population. NPC patients who remained alive at the last follow-up were censored. In addition, 5-year overall survival rate was used as secondary endpoint, defined as the percentage of NPC patients who are alive at the end of 60 months from the time the disease was diagnosed.
Sample preparation, DNA extraction and quantification of DNA yield
2.0 ml of fresh whole blood were collected from each study subject and were stored in EDTA tubes for preservation purposes. Cold chain (4°C) was maintained throughout the entire journey from hospitals to the laboratory in the university where the samples were processed. Genomic DNA was extracted from the blood samples using QIAamp® DNA Mini and Blood Mini kit (QIAgen, USA). The extracted DNA was stored at -80°C until further usage. The extracted DNA was quantified using nanophotometer and the absorbance was measured at wavelengths of 260nm and 280nm.
DNA genotyping (RFLP-PCR)
hOGG1 Ser326Cys, ITGA2 C807T, TNF-α -308G>A and XPD Lys751Gln polymorphisms were assessed by using RFLP-PCR. The sequence of forward and reverse primers used for each polymorphism is tabulated in Table 1. For hOGG1 Ser326Cys polymorphism, the outcome of the PCR was a 302bp product. The composition of a total 25μl PCR reaction was 12.5μl of GoTaq® Green Master Mix (Promega, Wisconsin, USA), 0.5μl of each primer (from working concentration of 10 μm), 0.5μl of genomic DNA, and the remaining was topped up with nuclease free water. The PCR thermal profile used was initial denaturation at 95°C for 5 min, 32 cycles of 95°C for 30s, followed by 63°C for 30s and 72°C for 30s, then ended with final extension of 72°C for 5 min. PCR yield from previous PCR was then digested by the restriction enzyme Fnu4HI (New England Biolabs, Ipswich, USA). After the digestion, homozygous Ser/Ser showed only a single 302bp product while homozygous Cys/Cys was fully digested into two different products that were 186bp and 116bp in size. Samples were identified as heterozygous Ser/Cys if 3 products of different sizes appeared in the gel.
Table 1. Sequence of forward and reverse primers used in RFLP-PCR.
| Polymorphisms |
Forward primer (5’-3’) | Reverse primer (5’-3’) |
|---|---|---|
| hOGG1 Ser326Cys (rs1052133) | CTT CCA CCT CCC AAC ACT GTC AC | GTG CCT GGC CTT TGA GGT AGT C |
| ITGA2 C807T (rs1126643) | GTG TTT AAC TTG AAC ACA TAT | ACC TTG CAT ATT GAA TTG CTT |
| TNF-α -308G>A (rs1800629) | AGG CAA TAG GTT TTG AGG GCC AT | ACA CTC CCC ATC CTC CCT GCT C |
| XPD Lys751Gln (rs13181) | CCC CCT CTC CCT TTC CTC TG | AAC CAG GGC CAG GCA AGA C |
For ITGA2 C807T polymorphism, 115bp product was generated at the end of the PCR protocol. The PCR composition used was 25μl PCR reaction consisting of 12.5μl of GoTaq® Green Master Mix (Promega, USA), 1.0μl of each primer (from working concentration of 10μm), 0.5μl of genomic DNA, and the remaining component was nuclease free water. The PCR thermal profile adopted was as follows: 95°C for 5 min, 35 cycles each of 95°C for 30s, followed by 55°C for 30s and 72°C for 30s, with final extension of 72°C for 5 min. The resulting PCR products were digested by restriction enzyme TaqI (New England Biolabs, Ipswich, US). Homozygous CC was fully digested into 2 products that were 92bp and 23bp in size after the excision by the restriction enzyme. For homozygous TT, no digestion occurred and only a single 115bp product was visible. All 3 products of different sizes were observed for heterozygous CT.
For TNF-α -308G>A polymorphism, 117bp PCR product spanning the promoter region of TNF-α from -238 to -308 was generated using the respective forward and reverse primers. The PCR composition used was 25μl PCR reaction consisting of 12.5μl of GoTaq® Green Master Mix (Promega, USA), 0.5μl of each primer (from working concentration of 10μm), 0.5μl of genomic DNA, and the remaining component was nuclease free water. The PCR thermal profile used was as follows: 95°C for 5 min; 95°C for 30s, 62°C for 30s and 72°C for 30s for 35 cycles; followed by 72°C for 5 min. Restriction enzyme NcoI (New England Biolabs, Ipswich, US) was utilized to excise the PCR products from the previous PCR run. Gel image of homozygous G/G showed only 2 products with 95bp and 22bp in size. Homozygous A/A was identified when the gel image showed a single product at 117bp while samples showing all 3 products were identified as heterozygous G/A.
For XPD Lys751Gln polymorphism, PCR product of 184bp in size was generated at the end of the PCR program. The PCR composition used was as follows: 25μl PCR reaction consisting of 12.5μl of GoTaq® Green Master Mix (Promega, USA), 0.5μl of each primer (from working concentration of 10μm), 0.5μl of genomic DNA, and the remaining was nuclease free water. The thermal profile of PCR used was 95°C for 5 min, 35 cycles each of 95°C for 40s, followed by 56°C for 30s and 72°C for 30s, with final extension of 72°C for 5 min. The resulting PCR products were digested by restriction enzyme MboII (New England Biolabs, Ipswich, US). Samples were identified as homozygous Lys/Lys if the results showed full digestion with only 2 products in size of 102bp and 82bp. Homozygous Gln/Gln samples showed only single PCR product that was 184bp in size. All 3 products of different sizes were observed for heterozygous Lys/Gln.
All digested PCR products were visualized under UV light after separation using electrophoresis on 3% ethidium bromide stained agarose gel. For quality control, 10% of the total PCR products were sent for DNA sequencing to confirm the results of RFLP-PCR for every polymorphism.
Statistical analysis
The dependent variable was all-cause survival of NPC patients while independent variables were TNM staging, hOGG1 Ser326Cys, ITGA2 C807T, TNF-α -308G>A and XPD Lys751Gln polymorphisms. Potential confounders controlled in the present study were cigarette smoking, alcohol consumption, salted fish consumption during childhood. Relative frequencies, percentages (%), means and standard deviations were used to describe the characteristics of the studied population. All-cause survival was computed for TNM staging, hOGG1 Ser326Cys, ITGA2 C807T, TNF-α -308G>A and XPD Lys751Gln polymorphisms using Kaplan-Meier method while log-rank test was used to test the differences in survival between genotypes within each SNP. Cox proportional hazard model was used to obtain hazard ratios (HRs) and the respective 95% confidence intervals (CIs) adjusted for the aforementioned confounders, to identify potential role of polymorphism on the all-cause survival of NPC. The threshold for statistical significance was set at 0.05. All of the previously mentioned statistical analyses were performed using SPSS version 21.
Results
Characteristics of study population for NPC survival analysis
The median follow-up duration for all study participants was 46.1 months (range: 1.8–73.0). A total of 109 (36.3%) NPC cases died from all-causes during the present study while the remaining 191 (63.7%) cases were censored. From the 109 deaths, 41 (37.6%) were NPC-specific deaths while the other 68 (62.4%) NPC cases died from other causes. At time of diagnosis, 66 out of 287 (23.0%) NPC cases were primary tumor category T1 while 39.0% were category T2; the number of NPC patients diagnosed at categories T3 and T4 were 54 (18.8%) and 55 (19.2%), respectively. For lymph node category, the number of NPC cases diagnosed with N0 were 47 out of 287 (16.4%), those diagnosed with N1 were 76 (26.4%) in total while 117 (40.8%) NPC patients were diagnosed with N2 and the remaining 47 (16.4%) cases were diagnosed with N3. NPC patients that experienced metastasis comprised of 9 (3.1%) out of 288 cases while 279 (96.9%) did not experience metastasis. The socio-demographic characteristics, the distribution of cigarette smoking, alcohol consumption, salted fish consumption during childhood and genotype frequencies for hOGG1 Ser326Cys, ITGA2 C807T, TNF-α -308G>A and XPD Lys751Gln polymorphisms are shown in Table 2.
Table 2. Characteristics of NPC patients (N = 300).
| Variables | ||
|---|---|---|
| Median follow-up, months (range) | 46.1 (1.8–73.0) | |
| Age (Mean ± SD) | 52.8 ± 10.9 | |
| NPC cases, N = 300 (%) | ||
| Survival (all-cause) | Alive/Censored | 191 (63.7) |
| Death | 109 (36.3) | |
| Cause of death | NPC-specific | 41 (37.6) |
| Other causes | 68 (62.4) | |
| Gender (n = 300) | Male | 232 (77.3) |
| Female | 68 (22.7) | |
| Ethnicity (n = 300) | Malay | 84 (28.0) |
| Chinese | 213 (71.0) | |
| Others | 3 (1.0) | |
| Salted fish consumption during childhood (n = 300) | Never | 103 (34.3) |
| Ever | 197 (65.7) | |
| Cigarette smoking (n = 300) |
Never | 146 (48.7) |
| Ever | 154 (51.3) | |
| Alcohol consumption (n = 300) |
Never | 161 (53.7) |
| Ever | 139 (46.3) | |
| TNM staging | ||
| Primary tumor staging (T) (n = 287) |
T1 | 66 (23.0) |
| T2 | 112 (39.0) | |
| T3 | 54 (18.8) | |
| T4 | 55 (19.2) | |
| Lymph node staging (N) (n = 287) |
N0 | 47 (16.4) |
| N1 | 76 (26.4) | |
| N2 | 117 (40.8) | |
| N3 | 47 (16.4) | |
| Metastasis status (M) (n = 288) |
M0 | 279 (96.9) |
| M1 | 9 (3.1) | |
| hOGG1 Ser326Cys (n = 300) |
Ser/Ser | 47 (15.7) |
| Ser/Cys | 161 (53.7) | |
| Cys/Cys | 92 (30.6) | |
| ITGA2 C807T (n = 300) |
C/C | 171 (57.0) |
| C/T | 103 (34.3) | |
| T/T | 26 (8.7) | |
| TNF-α -308G>A (n = 300) |
G/G | 244 (81.3) |
| G/A+A/A | 56 (18.7) | |
| XPD Lys751Gln (n = 300) |
Lys/Lys | 256 (85.3) |
| Lys/Gln+Gln/Gln | 44 (14.7) |
Effect of gene polymorphisms and TNM staging on survival of NPC patients (univariate analysis)
Most of the aforementioned gene polymorphisms did not significantly influence the all-cause survival of NPC in univariate analysis (data not shown). The only significant difference was observed with ITGA2 C807 polymorphism; NPC patients carrying C/C genotype had a mean survival time of 57.8 months while those carrying C/T and T/T genotypes survived for an average of 53.5 and 46.2 months, respectively (P-value = 0.052, log-rank test).
Results from the univariate analysis showed that only metastasis status was significantly associated with the all-cause survival of NPC (Table 3). NPC cases who were diagnosed without metastasis had a mean survival time of 56.0 months compared with 32.3 months for those diagnosed with metastasis to other organs (P-value<0.01, log-rank test). For primary tumor and node categories, no significant difference in mean survival times was observed (P-value = 0.909 and 0.056, respectively, log-rank test).
Table 3. Effect of TNM staging on survival of NPC patients (univariate analysis, N = 300).
| Variables | No of patients (%) | Mean survival time, months (95% CI) | p-valuea | |
|---|---|---|---|---|
| TNM staging | ||||
| Primary tumor staging (T) (n = 287) | T1 | 66 (23.0) | 56.6 (51.0–62.1) | 0.909 |
| T2 | 112 (39.0) | 54.5 (49.9–59.0) | ||
| T3 | 54 (18.8) | 55.2 (49.4–60.9) | ||
| T4 | 55 (19.2) | 53.2 (46.6–59.8) | ||
| Lymph node staging (N) (n = 287) | N0 | 47 (16.4) | 52.9 (46.1–59.7) | 0.056 |
| N1 | 76 (26.4) | 53.6 (48.1–59.0) | ||
| N2 | 117 (40.8) | 59.3 (55.3–63.3) | ||
| N3 | 47 (16.4) | 55.2 (52.4–58.0) | ||
| Metastasis status (M) (n = 288) | M0 | 279 (96.9) | 56.0 (53.2–58.7) | 0.002b |
| M1 | 9 (3.1) | 32.3 (17.6–47.0) |
abased on log-rank test
bp≤0.01
Effect of individual gene polymorphisms and TNM staging on survival of NPC patients controlling for cigarette smoking, alcohol and salted fish consumption during childhood
Most of the variables mentioned above were not significantly associated with all-cause survival of NPC after adjusting for cigarette smoking, alcohol and salted fish consumption during childhood except for ITGA2 C807T polymorphism and metastasis status (Table 4). For ITGA2 C807T polymorphism, the adjusted HR for T/T genotype carriers was 2.06 (95% CI = 1.14–3.72) compared to the reference C/C genotype, while NPC cases with distant metastasis showed an adjusted HR of 3.13 (95% CI = 1.44–6.82) compared to those without metastasis, consistent with results from the log-rank test above. The survival functions for ITGA2 C807T polymorphism and metastasis status (M) are presented in Fig 1. The 5-year overall survival rate for ITGA2 C/C carriers was 55% compared to those with C/T and T/T genotypes in which the survival rates were 50% and 43%, respectively. NPC cases who experience distant metastasis had a 5-year overall survival rate of 22% compared to the survival rate of 53% for those without metastasis.
Table 4. Gene polymorphisms and TNM staging with the all-cause survival (N = 300) analyzed individually using Cox regression.
| Variables | No of patients (%) | All-cause survival adjusteda HR (95% CI) |
p-value | |
|---|---|---|---|---|
| hOGG1 Genotypes (n = 300) |
Ser/Ser | 47 (15.7) | 1.0 | - |
| Ser/Cys | 161 (53.7) | 1.52 (0.84–2.76) | 0.169 | |
| Cys/Cys | 92 (30.6) | 1.22 (0.43–2.33) | 0.548 | |
| ITGA2 Genotypes (n = 300) |
C/C | 171 (57.0) | 1.0 | - |
| C/T | 103 (34.3) | 1.26 (0.84–1.89) | 0.272 | |
| T/T | 26 (8.7) | 2.06 (1.14–3.72) | 0.017b | |
| TNF-α Genotypes (n = 300) |
G/G | 244 (81.3) | 1.0 | - |
| G/A + A/A | 56 (18.7) | 1.01 (0.63–1.64) | 0.957 | |
| XPD Genotypes (n = 300) |
Lys/Gln + Gln/Gln | 44 (14.7) | 1.0 | - |
| Lys/Lys | 256 (85.3) | 1.15 (0.67–1.99) | 0.614 | |
| TNM staging | ||||
| Primary tumor staging (T) (n = 287) | T1 | 66 (23.0) | 1.0 | - |
| T2 | 112 (39.0) | 1.08 (0.64–1.81) | 0.785 | |
| T3 | 54 (18.8) | 0.93 (0.50–1.73) | 0.824 | |
| T4 | 55 (19.2) | 1.20 (0.66–2.16) | 0.557 | |
| Lymph node staging (N) (n = 287) | N0 | 47 (16.4) | 1.0 | - |
| N1 | 76 (26.4) | 1.06 (0.59–1.89) | 0.858 | |
| N2 | 117 (40.8) | 0.69 (0.39–1.22) | 0.199 | |
| N3 | 47 (16.4) | 1.36 (0.73–2.51) | 0.331 | |
| Metastasis status (M) (n = 288) |
M0 | 279 (96.9) | 1.0 | - |
| M1 | 9 (3.1) | 3.13 (1.44–6.82) | 0.004b |
ahazard ratio adjusted for salted fish consumption during childhood, cigarette smoking, alcohol consumption.
bp≤0.05
Fig 1.
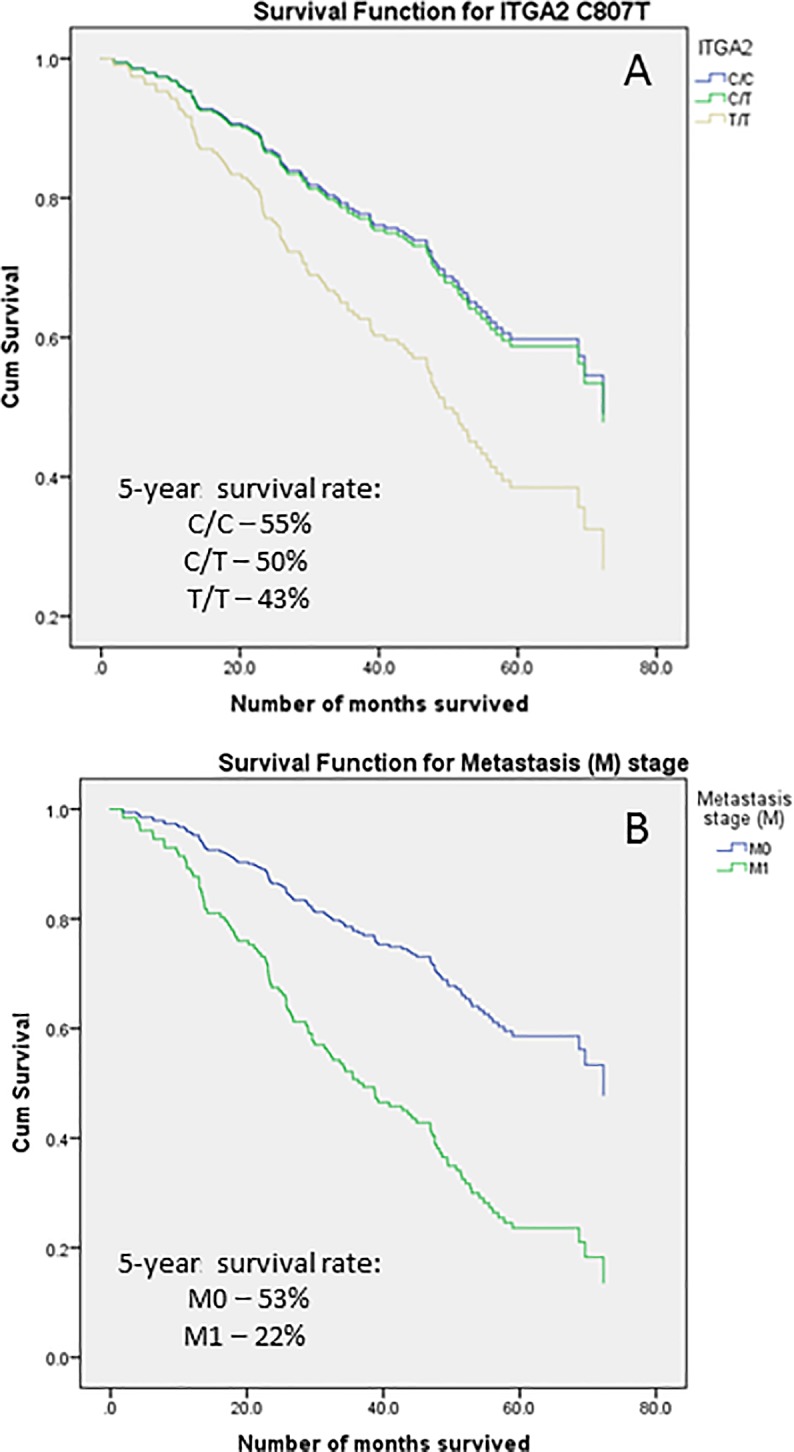
The survival functions for ITGA2 C807T polymorphism (A) and metastasis status (B).
Discussion
The distribution of TNM staging in the present study was mostly comparable to that of a number of studies from different populations. The only exception was that the proportion of NPC cases diagnosed with T1 staging was higher in the present study compared studies done elsewhere [58–60].
NPC cases carrying T/T genotype of ITGA2 C807T were observed to have a poorer all-cause survival as compared to those with C/C genotypes, with an adjusted HR of 2.06 (95% CI = 1.14–3.72) in individual model. The 5-year survival rate of C/C carriers was 55% compared to those with C/T and T/T where the survival rates were 50% and 43%, respectively. Our results also showed that NPC patients with ITGA2 C807T T/T genotype (3.8% metastasized) was more susceptible to metastasis compared to their C/C counterparts (1.8% metastasized). To the best of our knowledge, this is the first report of a significant association between integrin α2 polymorphism and survival in NPC. Molecular studies on human cancer have shown that tumor cells switch their integrin expression based on the stage of progression of the disease [61–62]. As a consequence of the changes in integrin expression, neoplastic cells tend to lose the integrins that maintain their adhesion to the basement membrane, at the same time, overexpressing the integrins that foster tumor cells’ survival, migration and proliferation [63]. In addition, integrins were found to provide the anchorage necessary for tumor cells’ migration during metastasis through activation of pro-migratory signals. Invasive malignant cells were found to be constantly making and breaking integrin contacts during cell migration [63]. Activation of myosin II by integrin-induced ERK pathway generates the contractile force that pulls the cells forward along the actin cables towards the newly-formed integrin contacts and breaks the adhesion at the trailing edge of the cells [64].
Integrin α2 is a collagen receptor that is mainly expressed on platelets as well as epithelial cells and the polymorphism in this study is located on position 807 was implicated in several cancer studies to be associated with increased cancer risk [19–20, 65]. Loss of ITGA2 expression was also found to be associated with metastasis of breast and colon cancer [17, 66]. Re-expression of α2β1 in breast cancer cells was shown to reverse the tumorigenic properties of the cells [67]. Similar results has been found in an animal study where integrin α2 was observed to be a metastasis suppressor as lack of α2β1 expression resulted in markedly increased cancer metastasis [68]. Deletion of α2β1 expression was also found in the same study to enhance tumor intravasation [68]. Suppression of FAK-CD-mediated loss of adhesion was also observed when integrin α2 was overexpressed due to activated Src in breast cancer cells [69]. In addition to its role in suppressing metastasis, integrin α2 causes apoptotic cell death via α2β1 transducing the necessary signal that promote cell death in matrix deprived cells [70].
However, contradicting results were found as integrin α2 has been linked to increased adhesion and migration facilitated by MAPK in an in-vivo study involving breast cancer cell [71]. In addition, integrin α2β1 was shown to promote prostate cancer cell metastasis to bone [72]. Inconsistent results from various cancer studies on the role of integrin α2 in cancer progression may suggest that integrin’s function is cell type and context dependent. Such a hypothesis is supported by a finding showing that different responses have been observed when integrin α2 knockout mice were challenged with 2 different cancer cells, namely B16F10 melanoma cells and Lewis Lung carcinoma cells [73]. Challenging the integrin α2 knockout mice with the former cell type resulted in increased tumor angiogenesis and yet there was no response from challenge of the latter [73].
Based on the results from the present study, it is postulated that altered expression of ITGA2 arising from the polymorphic difference of C807T resulted in increased tumor cell metastasis and intravasation. Apoptotic cell death resulting from the detachment of cells from the surrounding ECM might be suppressed as signal transduced by α2β1 integrin is required for the process. Thus, the effect of ITGA2 C807T polymorphism on both metastasis and apoptotic cell death might be one of the key contributing factors for shorter survival of NPC cases carrying the susceptible 807 T/T genotype compared to those with C/C genotype.
There is the question regarding the usefulness of ITGA2 C807T polymorphism in prognosticating the survival of NPC patients if only 8.7% of NPC patients carry the genotype associated with better survival. Given the current magnitude of percentage, the application of ITGA2 C807T in a mass screening program for prognostication of NPC patients’ survival would not be feasible. However, if other researchers can replicate and validate findings from the present study, a sizable number of NPC patients would have benefitted from the accurate prognostic prediction. ITGA2 C807T might be more clinically useful if it is used as a supplementary test for targeted populations on a limited basis.
Conclusion
The finding from the present study showed that ITGA2 C807T polymorphism could be potentially useful as a prognostic biomarker for NPC. NPC patients having T/T genotype were found with poorer 5-year survival compared to the wildtype C/C carriers. If ITGA2 C807T polymorphism is verified as a valid prognostic marker for NPC patients, the possibility exists for customizing treatment modalities for individuals with ITGA2 C807 T/T genotype in an attempt to enhance survival. However, the prognostic value of ITGA2 C807T polymorphism has to be validated by well-designed further studies with larger patient numbers.
Supporting information
(TIF)
(TIF)
(TIF)
(TIF)
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
The study was funded solely by the Science Fund from the Ministry of Science, Technology and Innovation, Malaysia as declared in the submission system (04-11-08-625FR). The funder had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Forman D, Bray F, Brewster DH, Gombe Mbalawa C, Kohler B, Pineros M, et al. Cancer Incidence in Five Continents, Vol. X IARC Scientific Publication No. 164. Lyon: International Agency for Research on Cancer; 2014. [Google Scholar]
- 2.Min HQ. The time trends of nasopharyngeal carcinoma: incidence rate and mortality for 20 years [M] In: Min HQ, editor. Study of nasopharyngeal carcinoma. Guangzhou: Science and Technology Press; 1998. pp.6–12. [Google Scholar]
- 3.Chang ET, Adami HO. The Enigmatic Epidemiology of Nasopharyngeal Carcinoma. Cancer Epidemiol Biomarkers Prev 2006; 15:1765 doi: 10.1158/1055-9965.EPI-06-0353 [DOI] [PubMed] [Google Scholar]
- 4.Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin 2011; 61:69–90. doi: 10.3322/caac.20107 [DOI] [PubMed] [Google Scholar]
- 5.Zainal Ariffin O, Nor Saleha IT. National Cancer Report 2007, Ministry of Health, Malaysia. 2011.
- 6.Spano JP, Busson P, Atlan D, Bourhis J, Pignon JP, Esteban C, et al. Nasopharyngeal carcinomas: an update. European Journal of Cancer 2003; 39:2121–35. [DOI] [PubMed] [Google Scholar]
- 7.Kam MK, Teo PM, Chau RM, Cheung KY, Choi PH, Kwan WH, et al. Treatment of nasopharyngeal carcinoma with intensity-modulated radiotherapy: the Hong Kong experience. Int J Radiat Oncol Biol Phys 2004; 60:1440–50. doi: 10.1016/j.ijrobp.2004.05.022 [DOI] [PubMed] [Google Scholar]
- 8.Chen R, Xu Y, Du X, Liu N, Li Y, He Q, et al. CXCL12 genetic variants as prognostic markers in nasopharyngeal carcinoma. Onco Targets Ther 2015; 8:2835–42. doi: 10.2147/OTT.S90430 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Chen L, Mao YP, Xie FY, Liu LZ, Sun Y, Tian L, et al. The seventh edition of the UICC/AJCC staging system for nasopharyngeal carcinoma is prognostically useful for patients treated with intensity-modulated radiotherapy from an endemic area in China. Radiother Oncol 2012; 104:331–7. doi: 10.1016/j.radonc.2011.10.009 [DOI] [PubMed] [Google Scholar]
- 10.Hofmann G, Langsenlehner U, Langsenlehner T, Glehr M, Gerger A, Absenger G, et al. Single nucleotide polymorphisms of integrin alpha-2 and beta-3 genes are not associated with relapse-free and overall survival in colorectal cancer patients. Anticancer research 2011; 31:1373–78. [PubMed] [Google Scholar]
- 11.Yu X, Miyamoto S, Mekada E. Integrin a2b1-dependent EGF receptor activation at cell-cell contact sites. Journal of Cell Science 2000; 113:2139–47. [DOI] [PubMed] [Google Scholar]
- 12.Abram CL, Lowell CA. Convergence of immunoreceptor and integrin signaling. Immunol Rev 2007; 218:29–44. doi: 10.1111/j.1600-065X.2007.00531.x [DOI] [PubMed] [Google Scholar]
- 13.Evans R, Patzak I, Svensson L, De Filippo K, Jones K, McDowall A, et al. Integrins in immunity. J Cell Science 2009; 122:215–25. doi: 10.1242/jcs.019117 [DOI] [PubMed] [Google Scholar]
- 14.Rathinam R, Alahari SK. Important role of integrins in the cancer biology. Cancer Metastasis Rev 2010; 29:223–37. doi: 10.1007/s10555-010-9211-x [DOI] [PubMed] [Google Scholar]
- 15.Streuli CH, Akhtar N. Signal co-operation between integrins and other receptor systems. Biochem J 2009; 418:491–506. doi: 10.1042/BJ20081948 [DOI] [PubMed] [Google Scholar]
- 16.Garmy-Susini B, Varner JA. Roles of integrins in tumor angiogenesis and lymphangiogenesis. Lymphat Res Biol 2008; 6:155–63. doi: 10.1089/lrb.2008.1011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ding W, Fan XL, Xu X, Huang JZ, Xu SH, Geng Q, et al. Epigenetic silencing of ITGA2 by MiR-373 promotes cell migration in breast cancer. PLoS One 2015; 10(8):e0135128 doi: 10.1371/journal.pone.0135128 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kritzik M, Savage B, Nugent DJ, Santoso S, Ruggeri ZM, Kunicki TJ. Nucleotide polymorphisms in the alpha2 gene define multiple alleles that are associated with differences in platelet alpha2 beta1 density. Blood 1998; 92:2382–88. [PubMed] [Google Scholar]
- 19.Gerger A, Hofmann G, Langsenlehner U, Renner W, Weitzer W, Wehrschutz M, et al. Integrin alpha-2 and beta-3 gene polymorphisms and colorectal cancer risk. Int J Colorectal Dis 2009; 24:159–163. doi: 10.1007/s00384-008-0587-9 [DOI] [PubMed] [Google Scholar]
- 20.Langsenlehner U, Renner W, Yazdani-Biuki B, Eder T, Wascher TC, Paulweber B, et al. Integrin alpha-2 and beta-3 gene polymorphisms and breast cancer risk. Breast Cancer Res Treat 2006; 97:67–72. doi: 10.1007/s10549-005-9089-4 [DOI] [PubMed] [Google Scholar]
- 21.Warzocha K, Ribeiro P, Bienvenu J, Roy P, Charlot C, Rigal D, et al. Genetic polymorphisms in the tumor necrosis factor locus influence Non-Hodgkin’s lymphoma outcome. Blood 1998; 91:3574–81. [PubMed] [Google Scholar]
- 22.Salles G, Bienvenu J, Bastion Y, Barbier Y, Doche C, Warzocha K, et al. Elevated circulating levels of TNFa and its p55 soluble receptor are associated with an adverse prognosis in lymphoma patients. Br J Haematol 1996; 93:352 [DOI] [PubMed] [Google Scholar]
- 23.Tracey KJ, Wei H, Manogue KR, Fong Y, Hesse DG, Nguyen HT, et al. Cachectin/tumor necrosis factor induces cachexia, anemia, and inflammation. J Exp Med 1988; 167:1211 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kroeger KM, Abraham LJ. Identification of an AP-2 element in the -323 to -285 region of the TNF-alpha gene. Biochem Mol Biol Int 1996; 40:43 [DOI] [PubMed] [Google Scholar]
- 25.Wilson AG, Symons JA, McDowell TL, McDevitt HO, Duff GW. Effects of a polymorphism in the human tumor necrosis factor alpha promoter on transcriptional activation. Proc Natl Acad Sci USA 1997; 94:3195 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Tarabar O, Cikota-Aleksic B, Tukic L, Milanovic N, Aleksic A, Magic Z. Association of interleukin-10, tumor necrosis factor-α and transforming growth factor-β gene polymorphisms with the outcome of diffuse large B-cell lumphomas. Int J Clin Oncol 2014; 19(1):186–92. doi: 10.1007/s10147-013-0531-z [DOI] [PubMed] [Google Scholar]
- 27.Juszczynski P, Kalinka E, Bienvenu J, Woszczek G, Borowiec M, Robak T, et al. Human leukocyte antigens class II and tumor necrosis factor genetic polymorphisms are independent predictors of non-Hodgkin lymphoma outcome. Blood 2002; 100:3037–40. doi: 10.1182/blood-2002-02-0654 [DOI] [PubMed] [Google Scholar]
- 28.Lech-Maranda E, Mlynarski W, Grzybowska-Izydorczyk O, Borowiec M, Pastorczak A, Cebula-Obrzut B, et al. Polymorphisms of TNF and IL-10 genes and clinical outcome of patients with chronic lymphocytic leukemia. Genes, Chromosomes & Cancer 2013; 52:287–96. [DOI] [PubMed] [Google Scholar]
- 29.Swenberg JA, Lu K, Moeller BC, Gao L, Upton PB, Nakamura J, et al. Endogenous versus exogenous DNA adducts: their role in carcinogenesis, epidemiology, and risk assessment. Toxicol Sci 2011; 120:S130–45. doi: 10.1093/toxsci/kfq371 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hecht SS. Tobacco smoke carcinogens and lung cancer. J Natl Cancer Inst 1999; 91(14), 1194–1210. [DOI] [PubMed] [Google Scholar]
- 31.Ward MH, Pan WH, Cheng YJ, Li FH, Brinton LA, Chen CJ, et al. Dietary exposure to nitrite and nitrosamines and risk of nasopharyngeal carcinoma in Taiwan. Int J Cancer 2000; 86(5), 603–609. [DOI] [PubMed] [Google Scholar]
- 32.Demple B, and Harrison L. Repair of oxidative damage to DNA: enzymology and biology. Annu Rev Biochem 1994; 63, 915–948. doi: 10.1146/annurev.bi.63.070194.004411 [DOI] [PubMed] [Google Scholar]
- 33.Coleman WB and Tsongalis GJ. The Molecular Basis of Human Cancer: Springer Science & Business Media; 2001. [Google Scholar]
- 34.Boiteux S and Radicella JP. The human OGG1 gene: structure, functions, and its implication in the process of carcinogenesis. Arch Biochem Biophys 2000; 377(1):1–8. doi: 10.1006/abbi.2000.1773 [DOI] [PubMed] [Google Scholar]
- 35.de Laat WL, Jaspers NG, Hoeijmakers JH. Molecular mechanism of nucleotide excision repair. Genes Dev 1999; 13(7):768–85. [DOI] [PubMed] [Google Scholar]
- 36.Nash HM, Lu R, Lane WS, Verdine GL. The critical active-site amine of the human 8-oxoguanine DNA glycosylase, hOgg1: direct identification, ablation and chemical reconstitution. Chem Biol 1997; 4(9):693–702. [DOI] [PubMed] [Google Scholar]
- 37.Radicella JP, Dherin C, Desmaze C, Fox MS, Boiteux S. Cloning and characterization of hOGG1, a human homolog of the OGG1 gene of Saccharomyces cerevisiae. Proc Natl Acad Sci USA 1997; 94(15):8010–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Yuan T, Wei J, Luo J, Liu M, Deng S and Chen P. Polymorphisms of base-excision repair genes hOGG1 326Cys and XRCC1 280His increase hepatocellular carcinoma risk. Dig Dis Sci 2012; 57:2451–7 doi: 10.1007/s10620-012-2192-6 [published Online First: 8 May 2012]. [DOI] [PubMed] [Google Scholar]
- 39.Xie H, Xia Kai, Rong H and Chen X. Genetic polymorphism in hOGG1 is associated with triple-negative breast cancer risk in Chinese Han women. The Breast 2013; 22(5):707–712 doi: 10.1016/j.breast.2012.12.016 [published Online First: 29 January 2013]. [DOI] [PubMed] [Google Scholar]
- 40.Elahi A, Zheng Z, Park J, Eyring K, McCaffrey T and Lazarus P. The human OGG1 DNA repair enzyme and its association with orolaryngeal cancer risk. Carcinogenesis 2002; 23(7):1229–34 doi: 10.1093/carcin/23.7.1229 [DOI] [PubMed] [Google Scholar]
- 41.Shen J, Deininger P, Hunt JD, Zhao H. 8-hydroxy-2’-deoxyguanosine (8-OH-dG) as a potential survival biomarker in patients with non-small-cell lung cancer. Cancer 2007; 109(3):574–80. doi: 10.1002/cncr.22417 [DOI] [PubMed] [Google Scholar]
- 42.Ha YS, Yan C, Kim IY, Yun SJ, Moon SK, Kim WJ. Tissue hOGG1 genotype predicts bladder cancer prognosis: a novel approach using a peptide nucleic acid clamping method. Ann Surg Oncol 2011; 18(6):1775–81. doi: 10.1245/s10434-010-1500-7 [DOI] [PubMed] [Google Scholar]
- 43.Li D, Li Y, Jiao L, Chang DZ, Beinart G, Wolff RA, et al. Effects of base excision repair gene polymorphisms on pancreatic cancer survival. Int J Cancer 2007; 120(8):1748–54. doi: 10.1002/ijc.22301 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Chen J, Suter B. Xpd, a structural bridge and a functional link. Cell Cycle 2003; 2(6):503–6. doi: 10.4161/cc.2.6.558 [DOI] [PubMed] [Google Scholar]
- 45.Sancar A. Mechanisms of DNA excision repair. Science 1994; 266: 1954–1956. [DOI] [PubMed] [Google Scholar]
- 46.Coin F, Marinoni JC, Rodolfo C, Fribourg S, Pedrini AM, Egly JM. Mutations in the XPD helicase gene result in XP and TTD phenotypes, preventing interaction between XPD and the p44 subunit of TFIIH. Nat Genet 1998; 20(2):184–88. doi: 10.1038/2491 [DOI] [PubMed] [Google Scholar]
- 47.Fan L, Fuss JO, Cheng QJ, Arvai AS, Hammel M, Roberts VA, et al. XPD Helicase Structures and Activities: Insights into the Cancer and Aging Phenotypes from XPD Mutations. Cell 2008; 133(5): 789–800. doi: 10.1016/j.cell.2008.04.030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Banescu C, Trifa AP, Demain S, Lazar EB, Dima D, et al. (2014) Polymorphism of XRCC1, XRCC3, and XPD Genes and Risk of Chronic Myeloid Leukemia. BioMed Research International 2014: 213790 doi: 10.1155/2014/213790 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Huang CG, Liu T, Lv GD, Liu Q, Feng JG, et al. Analysis of XPD genetic polymorphisms of esophageal squamous cell carcinoma in a population of Yili Prefecture, in Xinjiang, China. Mol Biol Rep 2012; 39(1): 709–714. doi: 10.1007/s11033-011-0789-z [DOI] [PubMed] [Google Scholar]
- 50.Lye MS, Visuvanathan S, Chong PP, Yap YY, Lim CC and Ban EZ. Homozygous Wildtype of XPD K751Q Polymorphism Is Associated with Increased Risk of Nasopharyngeal Carcinoma in Malaysian Population. PLoS One 2015; 10(6):e0130530 doi: 10.1371/journal.pone.0130530 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Farnebo L, Stjernstrom A, Fredrikson M, Ansell A, Garvin S, Thunell LK. DNA repair genes XPC, XPD, XRCC1, and XRCC3 are associated with risk and survival of squamous cell carcinoma of the head and neck. DNA Repair (Amst) 2015; 31:64–72. doi: 10.1016/j.dnarep.2015.05.003 [DOI] [PubMed] [Google Scholar]
- 52.Tiseo M, Giovannetti E, Tibaldi C, Camerini A, Di Costanzo F, Barbieri F, et al. Pharmacogenetic study of patients with advanced non-small cell lung cancer (NSCLC) treated with second-line pemetrexed or pemetrexed-carboplatin. Lung Cancer 2012; 78(1):92–9. doi: 10.1016/j.lungcan.2012.07.009 [DOI] [PubMed] [Google Scholar]
- 53.Tengstrom M, Mannermaa A, Kosma VM, Soini Y, Hirvonen A, Kataja V. MnSOD rs4880 and XPD rs13181 polymorphisms predict the survival of breast cancer patients treated with adjuvant tamoxifen. Acta Oncol 2014; 53(6):769–75. doi: 10.3109/0284186X.2014.892210 [DOI] [PubMed] [Google Scholar]
- 54.Pare L, Marcuello E, Altes A, del Rio E, Sedano L, Salazar J, et al. Pharmacogenetic prediction of clinical outcome in advanced colorectal cancer patients receiving oxaliplatin/5-fluorouracil as first-line chemotherapy. Br J Cancer 2008; 99(7):1050–55. doi: 10.1038/sj.bjc.6604671 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Tripepi G, Jager KJ, Dekker FW and Zoccali C. Selection Bias and Information Bias in Clinical Research. Nephron Clinical Practice 2010; 115(2), c94–c99. doi: 10.1159/000312871 [DOI] [PubMed] [Google Scholar]
- 56.Chen J, Liu NN, Li JQ, Yang L, Zeng Y, Zhao XM, et al. Association between ITGA2 C807T polymorphism and gastric cancer risk. World J Gastroenterol 2011; 17(23), 2860–2866. doi: 10.3748/wjg.v17.i23.2860 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Schoenfeld DA. Sample-size formula for the proportional-hazards regression model. Biometrics 1983;39:499–503. [PubMed] [Google Scholar]
- 58.Makni L, Stayoussef M, Ghazouani E, Mezlini A, Almawi WY, Yacoubi-Loueslati B. Distinct association of VEGF-A polymorphisms with laryngeal and nasopharyngeal cancer. Meta Gene 2016; 10, 90–94. [Google Scholar]
- 59.Nasr HB, Chahed K, Bouaouina N and Chouchane L. Functional vascular endothelial growth factor -2578 C/A polymorphism in relation to nasopharyngeal carcinoma risk and tumor progression. Clin Chim Acta 2008; 395(1–2), 124–129. doi: 10.1016/j.cca.2008.05.022 [DOI] [PubMed] [Google Scholar]
- 60.Zhu Q, Wang T, Ren J, Hu K, Liu W, Wu G. FAS-670A/G polymorphism: A biomarker for the metastasis of nasopharyngeal carcinoma in a Chinese population. Clin Chim Acta 2010; 411(3–4), 179–183. doi: 10.1016/j.cca.2009.10.024 [DOI] [PubMed] [Google Scholar]
- 61.Albelda SM, Mette SA, Elder DE, Stewart R, Damjanovich L, Herlyn M, Buck CA. Integrin distribution in malignant melanoma: association of the beta 3 subunit with tumor progression. Cancer Res 1990; 50(20), 6757–6764. [PubMed] [Google Scholar]
- 62.Breuss JM, Gallo J, DeLisser HM, Klimanskaya IV, Folkesson HG, Pittet JF, et al. Expression of the beta 6 integrin subunit in development, neoplasia and tissue repair suggests a role in epithelial remodeling. J Cell Sci 1995; 108 (Pt 6), 2241–2251. [DOI] [PubMed] [Google Scholar]
- 63.Guo W, and Giancotti FG. Integrin signalling during tumour progression. Nat Rev Mol Cell Biol 2004; 5(10), 816–826. doi: 10.1038/nrm1490 [DOI] [PubMed] [Google Scholar]
- 64.Conrad PA, Giuliano KA, Fisher G, Collins K, Matsudaira PT, Taylor DL. Relative distribution of actin, myosin I, and myosin II during the wound healing response of fibroblasts. J Cell Biol 1993; 120(6), 1381–1391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.FitzGerald LM, Patterson B, Thomson R, Polanowski A, Quinn S, Brohede J, et al. Identification of a prostate cancer susceptibility gene on chromosome 5p13q12 associated with risk of both familial and sporadic disease. Eur J Hum Genet 2009, 17(3), 368–377. doi: 10.1038/ejhg.2008.171 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Robertson JH, Yang SY, Winslet MC, and Seifalian AM. Functional blocking of specific integrins inhibit colonic cancer migration. Clin Exp Metastasis 2009; 26(7), 769–780. doi: 10.1007/s10585-009-9276-5 [DOI] [PubMed] [Google Scholar]
- 67.Zutter MM, Santoro SA, Staatz WD, and Tsung YL. Re-expression of the alpha 2 beta 1 integrin abrogates the malignant phenotype of breast carcinoma cells. Proc Natl Acad Sci USA 1995; 92(16), 7411–7415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Ramirez NE, Zhang Z, Madamanchi A, Boyd KL, O'Rear LD, Nashabi A, et al. The alpha(2)beta(1) integrin is a metastasis suppressor in mouse models and human cancer. J Clin Invest 2011; 121(1), 226–237. doi: 10.1172/JCI42328 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Park HB, Golubovskaya V, Xu L, Yang X, Lee JW, Scully S, et al. Activated Src increases adhesion, survival and alpha2-integrin expression in human breast cancer cells. Biochem J 2004; 378(Pt 2), 559–567. doi: 10.1042/BJ20031392 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Kozlova NI, Morozevich GE, Ushakova NA, Preobrazhenskaya ME, Shtil AA, Berman AE. The role of α2β1 integrin in anchorage dependent apoptosis of breast carcinoma and hepatoma cells. EXCLI Journal 2007; 6, 145–151. [Google Scholar]
- 71.Chen J, Diacovo TG, Grenache DG, Santoro SA, and Zutter MM. The alpha(2) integrin subunit-deficient mouse: a multifaceted phenotype including defects of branching morphogenesis and hemostasis. Am J Pathol 2002; 161(1), 337–344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Hall CL, Dubyk CW, Riesenberger TA, Shein D, Keller ET, van Golen KL. Type I collagen receptor (alpha2beta1) signaling promotes prostate cancer invasion through RhoC GTPase. Neoplasia 2008; 10(8), 797–803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Zhang Z, Ramirez NE, Yankeelov TE, Li Z, Ford LE, Qi Y, et al. alpha2beta1 integrin expression in the tumor microenvironment enhances tumor angiogenesis in a tumor cell-specific manner. Blood 2008; 111(4), 1980–1988. doi: 10.1182/blood-2007-06-094680 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(TIF)
(TIF)
(TIF)
(TIF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.