

Abstract
We present a model describing how Mg2+ binds and stabilizes specific RNA structures. In this model, RNA stabilization arises from two energetically distinct modes of Mg2+ binding: diffuse- and site-binding. Diffusely bound Mg2+ are electrostatically attracted to the strong anionic field around the RNA and are accurately described by the Poisson–Boltzmann equation as an ensemble distributed according to the electrostatic potentials around the nucleic acid. Site-bound Mg2+ are strongly attracted to specifically arranged electronegative ligands that desolvate the ion and the RNA binding site. Thus, site-binding is a competition between the strong coulombic attraction and the large cost of desolvating the ion and its binding pocket. By using this framework, we analyze three systems where a single site-bound Mg2+ may be important for stability: the P5 helix and the P5b stem loop from the P4-P6 domain of the Tetrahymena thermophila group I intron and a 58-nt fragment of the Escherichia coli 23S ribosomal RNA. Diffusely bound Mg2+ play a dominant role in stabilizing these RNA structures. These ions stabilize the folded structures, in part, by accumulating in regions of high negative electrostatic potential. These regions of Mg2+ localization correspond to ions that are observed in the x-ray crystallographic and NMR structures of the RNA. In contrast, the contribution of site-binding to RNA stability is often quite small because of the large desolvation penalty. However, in special cases, site-binding of partially dehydrated Mg2+ to locations with extraordinarily high electrostatic potential can also help stabilize folded RNA structures.
Divalent cations, like magnesium, stabilize the folded structure of most RNAs (1). The strong anionic field around the nucleic acid governs the interaction of Mg ions with RNA. As a result, Mg ions in solution are attracted to and accumulate around the RNA. The strong attractive interactions between cations and nucleic acids are generally described by the formation of different classes of bound ions in solution (1). The purpose of this work is to present a proper thermodynamic model describing how these different classes of ions together bind and stabilize specific RNA structures.
A Conceptual Framework for Mg+2 Binding
For RNA systems, two distinct classes of bound Mg2+ can be distinguished by the energetic features of their interaction with the RNA, diffusely bound and site-bound. Most of the Mg ions surrounding an RNA behave as a thermally fluctuating ensemble distributed according to a Boltzmann weighted average of the mean electrostatic potential around the RNA (2, 3). These ions can be called diffusely bound. This ensemble of diffusely bound ions preferentially accumulates in deep pockets of negative potential formed by the irregular shape of the molecular surface (2, 4). As a result, high concentrations of these diffusely bound ions may be localized in a relatively small volume around the RNA. Because these ions have a very large residence time in these pockets, they have a high probability of being detected by experimental methods like NMR or x-ray crystallography (2). One view of these diffusely bound ions is that they “dynamically occupy” various regions around the RNA (5). They are bound at specific locations only in the sense that they may be kinetically “trapped” in a region for a relatively long time. An important feature of these ions is that their hydration layer remains intact on average. In other words, any perturbations to the hydration layer at a particular location must be energetically insignificant in terms of the ensemble so that, on average, the chemical potential of these ions is characterized by the bulk aqueous state.
In some cases, however, an individual Mg ion can become fixed to a distinct location on the RNA where it interacts with a specific set of closely spaced electronegative RNA ligands. Importantly, the strong attraction of the ion to its ligands is sufficient to disrupt the hydration layer of the ion. These ions, which are adsorbed to the surface (6), are called site-bound. Energetically significant dehydration is the defining feature of site-binding. Displacing even a single water molecule from around Mg2+ is energetically costly. For example, water molecules outside the inner hydration layer stabilize Mg2+ by more than 200 kcal⋅mol−1 as a whole, and removal of even a single one of these water molecules after site-binding costs up to 10 kcal⋅mol−1 (7). The removal of a single inner-shell water molecule can cost more than 30 kcal⋅mol−1 (7). Because these enormous free energies are not accounted for in the ensemble description, the removal of waters from around the ion must be explicitly treated by using a general model of ionic desolvation.
The nonlinear Poisson–Boltzmann (NLPB) is a rigorous model that can be used to understand the electrostatic properties of RNA in solution (2–4, 8). In fact, the NLPB equation has been shown to provide a very accurate description of diffuse Mg2+ binding to RNA (2, 3). Here, we extend this model to approximate the energetics of Mg2+ site-binding as well (9). In this regard, we presume that the very large differences in solvation and electrostatic free energies are the primary determinants of Mg2+ site-binding (10). These differences allow for the use of the simple Born model for ion hydration to describe site-binding in our calculations.
A Theoretical Model for Mg+2 Binding
In this section, our conceptual notions of Mg2+ binding presented above are developed in terms of a rigorous physical model for ion–RNA interactions. We begin by defining the chemical potential of a molecule in an aqueous solution containing MgCl2 and NaCl (1, 11) as
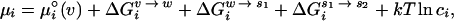 |
1 |
where k is the Boltzmann constant; T is the absolute temperature; ci is the molar concentration of the species, i; the term μ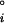 (v) is the vacuum standard state chemical potential; and ΔG
(v) is the vacuum standard state chemical potential; and ΔG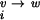 , ΔG
, ΔG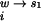 , and ΔG
, and ΔG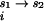 are the free energies of transferring each molecule from vacuum to pure water, from pure water to a univalent salt solution, and from the univalent salt solution to a mixed salt solution (containing both MgCl2 and NaCl), respectively. To focus on the interaction of divalent ions with the RNA, a reference state in univalent salt can be defined as
are the free energies of transferring each molecule from vacuum to pure water, from pure water to a univalent salt solution, and from the univalent salt solution to a mixed salt solution (containing both MgCl2 and NaCl), respectively. To focus on the interaction of divalent ions with the RNA, a reference state in univalent salt can be defined as
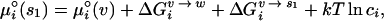 |
2 |
so that the overall chemical potential of the RNA can then be written
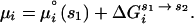 |
3 |
The last term, 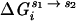 , includes both diffuse- and site-binding contributions to the interaction of divalent ions with the molecular species, i.
, includes both diffuse- and site-binding contributions to the interaction of divalent ions with the molecular species, i.
Fig. 1 shows a thermodynamic pathway for partitioning 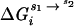 into contributions from both the diffuse- and site-binding of Mg2+ to the RNA. The shorthand notation of ΔGMg will refer to ΔGRNA
into contributions from both the diffuse- and site-binding of Mg2+ to the RNA. The shorthand notation of ΔGMg will refer to ΔGRNA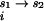 . The initial state is simply the reference state of the RNA in a univalent salt solution with a chemical potential given by μ
. The initial state is simply the reference state of the RNA in a univalent salt solution with a chemical potential given by μ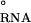 (s1). In the first step, the RNA is transferred into a solution containing the reference state concentration of Na+ along with a fixed concentration of Mg2+. The free energy of this step (ΔGD) is determined by interaction of only diffusely bound Mg2+ with the RNA. In the second step, individual Mg ions site-bind to specific locations on the RNA in the presence of monovalent cations and diffusely bound Mg ions. The free energy of this step is given by ΔGS. Therefore, the overall Mg-binding free energy, ΔGMg, is given by
(s1). In the first step, the RNA is transferred into a solution containing the reference state concentration of Na+ along with a fixed concentration of Mg2+. The free energy of this step (ΔGD) is determined by interaction of only diffusely bound Mg2+ with the RNA. In the second step, individual Mg ions site-bind to specific locations on the RNA in the presence of monovalent cations and diffusely bound Mg ions. The free energy of this step is given by ΔGS. Therefore, the overall Mg-binding free energy, ΔGMg, is given by
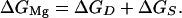 |
4 |
These free energy terms are discussed in more detail below.
Figure 1.

The thermodynamic pathway for calculating the stabilization of RNA by Mg2+; ΔΔGMg. This process consists of the free energy of site-binding, ΔGS, and diffuse-binding, ΔGD. The green spheres represent diffusely bound Mg2+; the yellow spheres represent site-bound Mg2+.
The term ΔGD is the electrostatic free energy of assembling a diffusely bound ensemble of Mg ions around the RNA. In the NLPB formalism, these diffusely bound Mg ions are treated as a cosolvent that affects the equilibrium through its influence on the chemical potential (or activity coefficients) of the RNA (12). Treating Mg2+ as a cosolvent is important because the long-range electrostatic interactions that dominate diffuse Mg2+ binding cannot be described properly by a simple mass action relationship. For RNA systems, ΔGD can be very accurately determined, using the NLPB equation, by calculating the difference in the total electrostatic free energy of the nucleic acid in an Mg-containing solution and an Mg-free solution (2, 3).
The term ΔGS is essentially a binding free energy of a charged ligand (Mg2+) to the RNA. This free energy has been described extensively elsewhere (9, 11, 13). For simplicity, we will consider only the case of a single site-bound ion, although the formalism can be extended easily to treat multiple site-binding. In this model, site-binding of a single Mg ion can be described by the equilibrium reaction:
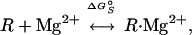 |
5 |
where R is the RNA without any site-bound Mg2+, and R⋅Mg2+ is the RNA with a single site-bound Mg2+. Because site-binding is dominated by local phenomena, the interaction between a site-bound ion and a discrete site can be described formally by an association constant Ka, which obeys the laws of mass action. Thus, ΔG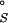 = −RTln(Ka) is the “standard state” binding free energy at given concentrations of NaCl and diffusely bound MgCl2.
= −RTln(Ka) is the “standard state” binding free energy at given concentrations of NaCl and diffusely bound MgCl2.
The term ΔG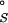 is the difference in chemical potential of the bound (R⋅Mgi2+) and free (R and Mg2+) species (11). As discussed elsewhere, ΔG
is the difference in chemical potential of the bound (R⋅Mgi2+) and free (R and Mg2+) species (11). As discussed elsewhere, ΔG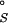 can be partitioned into several terms in the NLPB model (9, 12, 14, 15): the electrostatic attraction of Mg2+ to the site (the “solvent-screened” coulombic interaction free energy), ΔΔGelec; the dehydration of the ion, ΔΔG
can be partitioned into several terms in the NLPB model (9, 12, 14, 15): the electrostatic attraction of Mg2+ to the site (the “solvent-screened” coulombic interaction free energy), ΔΔGelec; the dehydration of the ion, ΔΔG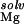 , and the site, ΔΔG
, and the site, ΔΔG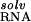 , upon binding; the displacement of univalent salt, ΔΔG
, upon binding; the displacement of univalent salt, ΔΔG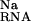 , and (diffusely bound) divalent salt, ΔΔG
, and (diffusely bound) divalent salt, ΔΔG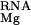 , from around the RNA upon binding; and the loss of translational entropy upon binding, ΔΔG
, from around the RNA upon binding; and the loss of translational entropy upon binding, ΔΔG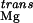 ,
,
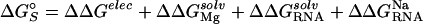 |
6 |
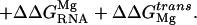 |
The terms ΔΔG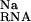 and ΔΔG
and ΔΔG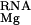 are determined at each set of salt conditions and energetically couple diffuse binding of univalent and divalent ions, respectively, to site-binding. The overall site-binding free energy, ΔGS, is then determined by the standard Langmuir-binding isotherm (16),
are determined at each set of salt conditions and energetically couple diffuse binding of univalent and divalent ions, respectively, to site-binding. The overall site-binding free energy, ΔGS, is then determined by the standard Langmuir-binding isotherm (16),
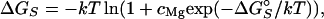 |
7 |
where cMg is the bulk Mg2+ concentration.
The view presented here is based on the premise that the very large differences in solvation and electrostatic free energies are the primary determinants of Mg2+ site-binding (10, 17). As such, the site-bound Mg ions are treated by using the Born model. In this model, the ions are treated as simple spheres defined by their effective radii immersed in a dielectric continuum. The Born radii have a well defined physical meaning related to the size of the solvent cavity formed by the ion (18–20). This simple model is very accurate and avoids the inherent complexity of modeling the hydration layer explicitly (18, 21).
Although other contributions such as van der Waals interactions and changes in the conformational entropy of the RNA certainly play a role, we postulate that they are secondary to the direct electrostatic interaction of Mg2+ with the RNA. By using the NLPB model, we try to provide a reasonable estimate of the electrostatic free energy of site-binding, thereby providing a quantitatively approximate but qualitatively robust description of site-binding.
Methods
Structural Models.
The details of the NLPB model have been reported elsewhere (22, 23). The three-dimensional NMR structures of both the P5 (24) and P5b (25) hairpins were obtained from the atomic coordinates deposited in the Protein Data Bank (26), whereas the refined crystallographic coordinates of the 58-nt domain of large subunit rRNA were provided by G. L. Conn (A. Gittis, E. Lattman, V.K.M., and D.E.D., unpublished data). Protons were added to the crystallographic coordinates and minimized by using the Biopolymer module of the INSIGHT II 97.0 software package (Molecular Simulations, San Diego). A crystallographically observed potassium ion at position 162 was included as part of the rRNA structure. Based on the available thermodynamic data (27), it is expected that this site will be fully occupied at 1.6 M univalent salt concentration used in the calculations.
The RNA was treated as a low dielectric medium (ɛm) within a volume enclosed by its water-accessible surface (probe radius = 1.4 Å). Results are presented for ɛm = 2, a physically reasonable value that accounts for the effects of electronic polarizability (15, 28). The AMBER4.1 parameter set for the nucleic acid was used to define the atomic charges and sizes (29). The surrounding solvent was treated as a continuum with a uniform dielectric constant (ɛs) of 80. The diffusely bound NaCl and MgCl2 were distributed according to a Boltzmann weighted average of the mean electrostatic potential. A 2.0-Å ion-exclusion radius was added to the surface of the macromolecule to account for ion size.
The site-bound Mg ions are treated by using the Born model. In this work, the radius of the Mg ion was set to 1.45 Å to reproduce the experimental hydration free energy of the free ion in pure aqueous solution (30). This value is identical to those found in many prior studies (18, 20, 31). As noted below, we performed additional site-binding calculations in which the Mg2+ was explicitly modeled as its hexahydrated derivative, Mg2+⋅(H2O)6. In these cases, the overall site-binding free energies were reduced by about 5% or less, without accounting for the dynamics of the bound waters. Therefore, given the complexity of modeling the hydration layer explicitly, we have used the more simple and elegant Born representation to describe site-binding in our calculations.
Numerical Methods.
Details of the finite difference procedure to calculate electrostatic potentials and free energies with the NLPB equation in a mixed salt solution have been reported (2, 32). The Na- and Mg-dependent contributions to the chemical potential of each RNA species were calculated by using a 653 lattice. The electrostatic potentials were calculated by using a two-step focusing procedure (33). For each run, potentials were iterated to a convergence of less than 10−4 kT/e. The relevant integrals required to calculate the Na- and Mg-dependent contributions to the binding free energy were evaluated as a summation over discrete lattice points as described (2, 11, 13). In each case, the ion atmosphere was integrated over the lattice generated in each of the three focusing runs and summed together. By using this procedure, charge neutrality was maintained to within 1%, and total free energy values varied by less than 3% for final grid resolutions of 1.0–1.4 grids per angstrom. The local surface potentials and ion concentrations, which do not depend on spatial integration, were calculated at a final resolution of 2.0 grids per angstrom.
The hydration-free energies of the RNA and the Mg ion were calculated by using finite difference solutions to the NLPB equation as implemented in the program DELPHI V.3.0 (34). Because the hydration-free energy critically depends on grid resolution, a resolution of at least 3 grids per angstrom was used for these calculations. In each case, calculations showed that the results were stable to within 3% with grid placement.
Results and Discussion
We use the thermodynamic framework, presented above, to explore three RNA systems where a single site-bound Mg2+ is thought to be important for stability: the P5 helix (25) and the P5b stem loop (24) from the P4-P6 domain of the Tetrahymena thermophila group I intron, and a 58-nt fragment (nucleotides 1051–1108) derived from the Escherichia coli 23S rRNA (35). In each case, a high-resolution structure has located an Mg ion (or analogue) in a specific region of the RNA. We will explore the energetics of Mg2+ binding to each of these locations in the context of the overall Mg2+-induced stabilization of the RNA. The P5 helix and P5b stem loop provide models for the interaction of Mg with simple RNA folding motifs, whereas the rRNA fragment provides an example of Mg2+ binding to a more complex RNA tertiary structure (36).
The RNA Hairpins.
Mg2+ binding to the P5 helix and P5b stem loop has been characterized by using proton NMR of cobalt (III) hexammine as an analogue of solvated Mg (24, 25). As found in other RNA molecules (2, 4, 37), the observed metal ion in each of these RNA hairpins is localized to a region of highly negative electrostatic potential in the major groove (Fig. 2a). In Table 1, we list the calculated electrostatic potential at the observed metal binding site along with the energetic contributions to Mg2+ site-binding for each RNA hairpin. For the P5 helix, the calculations were done by using each of the 19 members of the ensemble of structures derived from NMR (25) and then averaged, whereas for the P5b stem loop the calculations were done by using the averaged NMR structure (24). The results for both hairpins are quite similar.
Figure 2.

(Upper) The electrostatic potential mapped onto the solvent-accessible surface of the P5 helix (24) calculated at 0.1 M univalent salt concentration and 0.001 M bulk Mg concentration. The experimentally observed metal ion is shown as a green sphere. (Lower) The 4.5 M three-dimensional isoconcentration contour of diffusely bound Mg2+ around the RNA is shown in green. Their correspondence of the high concentration region to the observed ion is seen. The surface potential scale is in units of kT/e. The figures were rendered by using the program grasp (43).
Table 1.
Energetic contributions to Mg2+ site-binding in kcal/mol
| RNA | φ, kT/e | ΔΔGelec | ΔΔG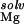
|
ΔΔG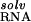
|
ΔΔG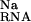
|
ΔΔG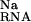 ‡ ‡
|
ΔG
|
|---|---|---|---|---|---|---|---|
| Hairpins* | |||||||
| P5b | −5.9 ± 0.4 | −19.3 ± 2.0 | 25.1 ± 6.3 | 2.4 ± 0.9 | 32.5 ± 5.9 | 0.3 ± 0.1 | 44.0 |
| P5 | −7.2 | −29.3 | 36.5 | 3.0 | 19.7 | 0.5 | 30.4 |
| 58-nt rRNA† | |||||||
| 163 | −39.7 | −135.2 | 102.4 | 10.3 | 44.8 | 0.05 | 25.4 |
| 167 | −57.7 | −113.2 | 61.3 | 3.5 | 40.6 | 0.03 | −4.8 |
Calculated at 0.1 M univalent salt concentration.
Calculated at 1.6 M univalent salt concentration.
Calculated at 0.001 M Mg concentration.
The electrostatic potential at the presumed binding site is between about −6 to −7 kT/e (Table 1). This negative potential provides a strong driving force for the site-binding of Mg2+ to the RNA, as reflected by the large favorable value of ΔΔGelec (Table 1). However, for both hairpins, the enormous cost of dehydrating the Mg2+ (ΔΔG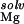 ) itself prevents site-binding (Table 1). It is important to realize that the experimentally observed ion has a completely intact inner hydration layer. The term ΔΔG
) itself prevents site-binding (Table 1). It is important to realize that the experimentally observed ion has a completely intact inner hydration layer. The term ΔΔG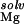 arises primarily from the displacement of water molecules in outer hydration shells. In these calculations, perturbing these outer-shell waters can cost more than 35 kcal⋅mol−1 (Table 1), consistent with experimental measures of Mg2+ hydration. Calculations in which the Mg2+ was explicitly modeled as its hexahydrated derivative, Mg2+⋅(H2O)6, show similarly large dehydration costs (data not shown). The accompanying cost of dehydrating the partially charged groups in the RNA binding site (ΔΔG
arises primarily from the displacement of water molecules in outer hydration shells. In these calculations, perturbing these outer-shell waters can cost more than 35 kcal⋅mol−1 (Table 1), consistent with experimental measures of Mg2+ hydration. Calculations in which the Mg2+ was explicitly modeled as its hexahydrated derivative, Mg2+⋅(H2O)6, show similarly large dehydration costs (data not shown). The accompanying cost of dehydrating the partially charged groups in the RNA binding site (ΔΔG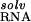 ) is relatively small, but significant (Table 1).
) is relatively small, but significant (Table 1).
The formation of the site-bound complex is also strongly opposed by the displacement of Na from around the nucleic acid (ΔΔG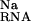 ; Table 1). This effect, generally seen in the interaction of charged ligands with nucleic acids, is the result of the repulsion and displacement of diffusely bound univalent ions by the site-bound Mg2+ (11–14). It is a “desolvation penalty” arising from the loss of favorable electrostatic interactions between the univalent ion and the RNA (11). The NLPB equation provides a very accurate estimate of this term (14). For the P5b stem loop, we note that the magnitude of ΔΔG
; Table 1). This effect, generally seen in the interaction of charged ligands with nucleic acids, is the result of the repulsion and displacement of diffusely bound univalent ions by the site-bound Mg2+ (11–14). It is a “desolvation penalty” arising from the loss of favorable electrostatic interactions between the univalent ion and the RNA (11). The NLPB equation provides a very accurate estimate of this term (14). For the P5b stem loop, we note that the magnitude of ΔΔG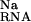 is substantially larger than ΔΔGelec at 0.1 M bulk Na concentration. At 1 mM bulk Mg concentration, the analogous contribution arising from the displacement of diffusely bound Mg2+, ΔΔG
is substantially larger than ΔΔGelec at 0.1 M bulk Na concentration. At 1 mM bulk Mg concentration, the analogous contribution arising from the displacement of diffusely bound Mg2+, ΔΔG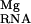 , is quite small (Table 1) because of the relatively high monovalent ion concentrations studied here.
, is quite small (Table 1) because of the relatively high monovalent ion concentrations studied here.
Finally, the loss of translational entropy that accompanies Mg2+ binding, ΔΔG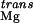 , disfavors site-binding as well (Table 1). This term is estimated to be about 3.0 kcal/mol based on the method of Amzel (38) and is not a dominant contribution to binding. In this formulation, we have neglected any changes in the conformational entropy of the RNA caused by Mg2+ binding, which are extremely difficult to estimate (39). Combining the various free-energy terms, we find that the overall free-energy change, ΔG
, disfavors site-binding as well (Table 1). This term is estimated to be about 3.0 kcal/mol based on the method of Amzel (38) and is not a dominant contribution to binding. In this formulation, we have neglected any changes in the conformational entropy of the RNA caused by Mg2+ binding, which are extremely difficult to estimate (39). Combining the various free-energy terms, we find that the overall free-energy change, ΔG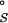 , strongly opposes site-binding to both hairpins (Table 1). The unfavorable free energy occurs because the favorable electrostatic interaction between the site-bound Mg2+ and the RNA (ΔΔGelec) cannot overcome the large cost of desolvating the ion and the nucleic acid (i.e., ΔΔG
, strongly opposes site-binding to both hairpins (Table 1). The unfavorable free energy occurs because the favorable electrostatic interaction between the site-bound Mg2+ and the RNA (ΔΔGelec) cannot overcome the large cost of desolvating the ion and the nucleic acid (i.e., ΔΔG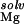 and ΔΔG
and ΔΔG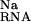 ).
).
In contrast, diffusely bound Mg ions strongly stabilize the folded hairpins at physiological Mg2+ concentrations. Fig. 3 contains a plot of ΔGD as a function of bulk Mg2+ concentration for each hairpin. As shown in the figure, the term ΔGD grows progressively larger with bulk Mg2+ concentration. This effect arises from the progressive accumulation of diffusely bound Mg2+ around the RNA (3). The experimentally observed Mg2+ association constants for each hairpin are at least 1–1.5 × 102 M−1 (25) and are readily explained by ΔGD alone. As found for homopolynucleotides and tRNA (2, 3), the strong coulombic attraction of diffusely bound Mg ions to each hairpin outweighs the accompanying loss of electrostatic interactions between Na and the RNA. As discussed (2, 3), diffuse binding is augmented further by the “entropic release” of Na ions from around the RNA.
Figure 3.

The free-energy contribution of diffuse Mg2+ binding, ΔGD, to the P5 helix (⋅ ⋅ ⋅) and the P5b hairpin (——) calculated at 0.1 M univalent salt concentration.
As shown in Fig. 2 (Lower), the ensemble of diffusely bound ions preferentially accumulates in the major groove of the hairpin and stabilizes the folded hairpin by interacting most strongly with pockets of negative potential formed by the irregular shape of the molecular surface. Comparing the two hairpin representations in Fig. 2, this region of Mg2+ accumulation corresponds to the experimentally observed metal ion, similar to what has been found for yeast tRNAPhe (2). The isoconcentration contour in Fig. 2 (Lower) represents the time-averaged spatial distribution of Mg2+ around the RNA.
The 23S rRNA Fragment.
A 58-nt fragment derived from the E. coli large subunit rRNA adopts a very compact tertiary structure (36) that requires both monovalent and divalent ions to fold (35, 40, 41). The folding of this rRNA fragment in solution has been extensively characterized by using thermal denaturation experiments (35, 40, 42). These studies show that a single highly selective Mg2+ binding site and a second much less selective site stabilize the tertiary structure of the RNA at 1.6 M univalent salt concentration (35).
A number of Mg ions have been found in recently refined crystallographic structures of the RNA (G. Conn, A. Gittis, E. Lattman, V.K.M., and D.E.D., unpublished data). Again, each of these metal ions is located in a region of highly negative electrostatic potential. For most of the crystallographically observed sites, the electrostatic potential is between about −2 to −9 kT/e (G. Conn, A. Gittis, E. Lattman, V.K.M., and D.E.D., unpublished data). These sites lie in the grooves of the RNA like those found in the RNA hairpins described previously. The energetics of Mg2+ binding to these sites is qualitatively quite similar to that observed for the hairpin structures. That is to say, ΔΔGelec cannot overcome the large cost associated with ΔΔG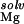 and ΔΔG
and ΔΔG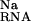 . As a result, site-binding to these specific locations is generally unfavorable. Again, the crystallographically observed metal ions correspond to electronegative pockets where diffusely bound Mg2+ ions accumulate and become trapped (G. Conn, A. Gittis, E. Lattman, V.K.M., and D.E.D., unpublished data).
. As a result, site-binding to these specific locations is generally unfavorable. Again, the crystallographically observed metal ions correspond to electronegative pockets where diffusely bound Mg2+ ions accumulate and become trapped (G. Conn, A. Gittis, E. Lattman, V.K.M., and D.E.D., unpublished data).
However, the electrostatic potentials at two sites, numbers 163 and 167, are an order of magnitude larger than at other locations (Table 1). These two sites are located within an interhelical cleft where the sugar-phosphate backbone is buried within the RNA. In this region, the electrostatic field is focused to form two deep electronegative pockets where the Mg ions are found (Fig. 4). These ions are obvious candidates for the thermodynamically important site-bound ion that is experimentally observed. For each ion, the free-energy analysis again reveals the competing forces that are involved in site-binding. In each case, ΔΔGelec provides a strong driving force for binding. However, at site 163, the large cost of displacing both inner- and outer-layer waters from around the deeply buried Mg2+ (ΔΔG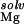 ) together with the repulsion of the monovalent ion atmosphere, ΔΔG
) together with the repulsion of the monovalent ion atmosphere, ΔΔG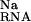 , ultimately prevents binding to the site. In contrast, the ion at site 167 is able to exploit the large electrostatic potential in the cleft by remaining more solvent-exposed, thereby reducing ΔΔG
, ultimately prevents binding to the site. In contrast, the ion at site 167 is able to exploit the large electrostatic potential in the cleft by remaining more solvent-exposed, thereby reducing ΔΔG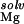 , a strategy that has been noted for other nucleic acid-binding ligands (14). Although both of these observed ions lie in regions where diffusely bound Mg2+ accumulates, our calculations show that site-binding to position 167 is able to displace these ions (with a cost given by ΔΔG
, a strategy that has been noted for other nucleic acid-binding ligands (14). Although both of these observed ions lie in regions where diffusely bound Mg2+ accumulates, our calculations show that site-binding to position 167 is able to displace these ions (with a cost given by ΔΔG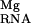 ) and bind to the tertiary structure of the RNA.
) and bind to the tertiary structure of the RNA.
Figure 4.

The electrostatic potential mapped onto the solvent-accessible surface of the RNA calculated at 0.15 M univalent salt concentration and 0.001 M bulk Mg concentration. The two high-potential regions corresponding to positions 163 and 167 are shown. Experimentally observed Mg ions are represented by green spheres The surface potential scale is in units of kT/e. The figures were rendered by using the program grasp (43).
In Fig. 5, the site-binding isotherm is plotted. As expected, the free energy of site-binding, ΔGS, becomes more favorable with increasing Mg2+ concentrations. However, at high Mg2+ concentrations, flattening of the curve results from the anticooperative interaction between site-bound and diffusely bound ions. This interaction is given by the term ΔΔG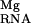 .
.
Figure 5.

The free-energy contributions to Mg2+ binding to the 58-nt rRNA fragment. ΔGD, ⋅ ⋅ ⋅; ΔGS, - - -; and ΔΔGMg, ——, calculated at 1.6 M univalent salt concentration.
Surprisingly, diffusely bound Mg ions also stabilize the tertiary structure of the rRNA fragment at 1.6 M univalent salt concentration. The free energy of diffuse-binding, ΔGD, is also plotted in Fig. 5. Even a few millimolars of Mg2+ effectively compete with high concentrations of univalent ions to interact with the RNA. At low Mg2+ concentrations, the magnitude of ΔGD is of the same order of magnitude as ΔGS. Thus, at 1 mM bulk Mg2+ concentration, the site-bound ion at position 167 binds to the RNA with a free energy of −0.8 kcal⋅mol−1, whereas the ensemble of diffusely bound ions binds with a free energy of about −0.3 kcal⋅mol−1. These results are qualitatively consistent with the two classes of binding sites that are observed in the experimental Mg2+-binding isotherms (35). At higher Mg2+ concentrations, ΔGD is clearly the dominant contribution to RNA stability (Fig. 5).
Conclusions
We have presented a thermodynamic framework to describe how Mg2+ binds and stabilizes specific RNA structures. In this model, the overall stabilization of RNA by Mg2+ arises from two distinct modes of binding, diffuse- and site-binding, distinguished by their energetic properties. Diffusely bound Mg2+ interact with the strong anionic field around the nucleic acid by means of long-range coulombic interactions described by the NLPB equation. As such, these ions behave as a thermally fluctuating ensemble distributed according to a Boltzmann weighted average of the mean electrostatic potential around the RNA (2, 3). Thus, there is a high probability of finding these ions in pockets of high negative electrostatic potential on the surface of the RNA, where they can be detected by experimental methods like NMR or x-ray crystallography. Because diffuse Mg2+ binding is dominated by long-range electrostatic interactions, these ions cannot be described properly by simple mass action equilibria. In our framework, diffusely bound ions act as a cosolvent that alters the activity of the RNA (Eq. 2; ref. 11).
It is important to remember that all diffusely bound ions within an ensemble are identical from an energetic perspective (i.e., at equilibrium, the chemical potential of each ion is uniform throughout the solution), although they are not all identical on a molecular level. This means that any perturbations to the hydration layer of an ion at a particular location (an ionic “microstate”) must be energetically insignificant in terms of the ensemble so that, on average, the chemical potential of any ion within the ensemble is characterized by the fully hydrated bulk state. This distinction is especially important for the highly populated microstates of the ions that are located in potential wells near the RNA surface. The hydration layer of the ions in such microstates may have significant fluctuations from the mean, but these ions still behave as though they are fully hydrated, on average.
Site-bound Mg ions also interact with specifically arranged anionic ligands on the RNA through strong coulombic forces. However, the accompanying disruption of the hydration of the ion has a substantial energetic cost and must be explicitly treated. As such, site-binding is described as a competition between the strong electrostatic attraction and the large cost of desolvating the ion and the nucleic acid. In our framework, site-binding is treated as a simple mass-action equilibrium between bound and free states (Eq. 5). We have tried to provide a reasonable estimate of the electrostatic free energy of site-binding by using the NLPB model, thereby providing a quantitatively approximate but qualitatively robust description of site-binding.
We used the thermodynamic framework, presented above, to explore three RNA systems where a single site-bound Mg2+ is thought to be important for stability. In these systems, we find that diffusely bound Mg ions play a dominant role in stabilizing the folded structures of RNA. These ions stabilize the folded structure, in part, by accumulating in regions of high negative electrostatic potential. These regions of Mg2+ localization correspond to bound ions that are observed in the x-ray crystallographic and NMR structures of the RNA. In contrast, we find that the overall contribution of site-binding to RNA stability is generally quite small because of the large desolvation penalty. However, binding of a single partially dehydrated Mg2+ to a site with extraordinarily high electrostatic potential helps stabilize the tertiary structure of the rRNA. Although such sites are likely to be relatively uncommon, they are very important in stabilizing complex RNA folds.
Acknowledgments
V.K.M. is a Howard Hughes Medical Institute Physician Postdoctoral Fellow. This work was supported by National Institutes of Health Grant GM58545.
Abbreviation
- NLPB
nonlinear Poisson–Boltzmann
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
References
- 1.Misra V K, Draper D E. Biopolymers. 1998;48:113–135. doi: 10.1002/(SICI)1097-0282(1998)48:2<113::AID-BIP3>3.0.CO;2-Y. [DOI] [PubMed] [Google Scholar]
- 2.Misra V K, Draper D E. J Mol Biol. 2000;299:813–825. doi: 10.1006/jmbi.2000.3769. [DOI] [PubMed] [Google Scholar]
- 3.Misra V K, Draper D E. J Mol Biol. 1999;294:1135–1147. doi: 10.1006/jmbi.1999.3334. [DOI] [PubMed] [Google Scholar]
- 4.Chin K, Sharp K A, Honig B, Pyle A M. Nat Struct Biol. 1999;6:1055–1061. doi: 10.1038/14940. [DOI] [PubMed] [Google Scholar]
- 5.Robinson H, Gao Y, Sanishvili R, Joachimiak A, Wang A H-J. Nucleic Acids Res. 2000;28:1760–1766. doi: 10.1093/nar/28.8.1760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bockris J O M, Reddy A K N, Gamboa-Aldeco M. Modern Electrochemistry: Fundamentals of Electrodics. New York: Academic; 2000. [Google Scholar]
- 7.Peschke M, Blades A T, Kebarle P. J Phys Chem A. 1998;102:9978–9985. [Google Scholar]
- 8.Sharp K A, Honig B, Harvey S C. Biochemistry. 1990;29:340–346. doi: 10.1021/bi00454a006. [DOI] [PubMed] [Google Scholar]
- 9.Gilson M K, Honig B. Proteins Struct Funct Genet. 1988;4:7–18. doi: 10.1002/prot.340040104. [DOI] [PubMed] [Google Scholar]
- 10.Eisenman G, Horn R. J Membr Biol. 1983;76:197–225. doi: 10.1007/BF01870364. [DOI] [PubMed] [Google Scholar]
- 11.Sharp K A, Friedman R A, Misra V, Hecht J, Honig B. Biopolymers. 1995;36:245–262. doi: 10.1002/bip.360360211. [DOI] [PubMed] [Google Scholar]
- 12.Sharp K A. Biopolymers. 1995;36:227–243. doi: 10.1002/bip.360360211. [DOI] [PubMed] [Google Scholar]
- 13.Misra V K, Sharp K A, Friedman R A, Honig B. J Mol Biol. 1994;238:245–263. doi: 10.1006/jmbi.1994.1285. [DOI] [PubMed] [Google Scholar]
- 14.Misra V K, Honig B. Proc Natl Acad Sci USA. 1995;92:4691–4695. doi: 10.1073/pnas.92.10.4691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Misra V K, Hecht J L, Yang A-S, Honig B. Biophys J. 1998;75:2262–2273. doi: 10.1016/S0006-3495(98)77671-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hill T L. Cooperativity Theory in Biochemistry. Steady State and Equilibrium Systems. New York: Springer; 1985. [Google Scholar]
- 17.Eisenman G. Biophys J. 1962;2:259–323. doi: 10.1016/s0006-3495(62)86959-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Rashin A A, Honig B. J Phys Chem. 1985;89:5588–5593. [Google Scholar]
- 19.Roux B, Yu H A, Karplus M. J Phys Chem. 1990;94:4683–4688. [Google Scholar]
- 20.Babu C S, Lim C. Chem Phys Lett. 1999;310:225–228. [Google Scholar]
- 21.Jayaram B, Fine R, Sharp K, Honig B. J Phys Chem. 1989;93:4320–4327. [Google Scholar]
- 22.Sharp K A, Honig B. Annu Rev Biophys Biomol Struct. 1990;19:301–332. doi: 10.1146/annurev.bb.19.060190.001505. [DOI] [PubMed] [Google Scholar]
- 23.Honig B, Nicholls A. Science. 1995;268:1144–1149. doi: 10.1126/science.7761829. [DOI] [PubMed] [Google Scholar]
- 24.Kieft J S, Tinoco I J. Structure (London) 1997;5:713–721. doi: 10.1016/s0969-2126(97)00225-6. [DOI] [PubMed] [Google Scholar]
- 25.Colmenarejo G, Tinoco I. J Mol Biol. 1999;290:119–135. doi: 10.1006/jmbi.1999.2867. [DOI] [PubMed] [Google Scholar]
- 26.Berman H M, Westbrook J, Feng Z, Gilliland G, Bhat T N, Weissig H, Shindyalov I N, Bourne P E. Nucleic Acids Res. 2000;28:235–242. doi: 10.1093/nar/28.1.235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Shiman R, Draper D E. J Mol Biol. 2000;302:79–91. doi: 10.1006/jmbi.2000.4031. [DOI] [PubMed] [Google Scholar]
- 28.Gilson M K, Honig B H. Biopolymers. 1986;25:2097–2119. doi: 10.1002/bip.360251106. [DOI] [PubMed] [Google Scholar]
- 29.Cornell W D, Cieplak P, Bayly C I, Gould I R, Merz K M, Ferguson D M, Spellmeyer D C, Fox T, Caldwell J W, Kollman P A. J Am Chem Soc. 1995;117:5179–5197. [Google Scholar]
- 30.Marcus Y. Ion Properties. New York: Dekker; 1997. [Google Scholar]
- 31.Dudev T, Lim C. J Phys Chem. 2000;104:3692–3694. [Google Scholar]
- 32.Chen S W, Honig B. J Phys Chem B. 1997;101:9113–9118. [Google Scholar]
- 33.Gilson M K, Sharp K A, Honig B H. J Comput Chem. 1987;9:327–335. [Google Scholar]
- 34.Nicholls A, Honig B. J Comp Chem. 1991;12:435–445. [Google Scholar]
- 35.Bukhman Y V, Draper D E. J Mol Biol. 1997;273:1020–1031. doi: 10.1006/jmbi.1997.1383. [DOI] [PubMed] [Google Scholar]
- 36.Conn G L, Draper D E, Lattman E E, Gittis A G. Science. 1999;284:1171–1174. doi: 10.1126/science.284.5417.1171. [DOI] [PubMed] [Google Scholar]
- 37.Hermann T, Westhof E. Structure (London) 1998;6:1303–1314. doi: 10.1016/s0969-2126(98)00130-0. [DOI] [PubMed] [Google Scholar]
- 38.Amzel L M. Proteins. 1997;28:144–149. [PubMed] [Google Scholar]
- 39.Reyes C M, Kollman P A. J Mol Biol. 2000;297:1145–1158. doi: 10.1006/jmbi.2000.3629. [DOI] [PubMed] [Google Scholar]
- 40.Laing L G, Gluick T C, Draper D E. J Mol Biol. 1994;237:577–587. doi: 10.1006/jmbi.1994.1256. [DOI] [PubMed] [Google Scholar]
- 41.Laing L G, Draper D E. J Mol Biol. 1994;237:560–576. doi: 10.1006/jmbi.1994.1255. [DOI] [PubMed] [Google Scholar]
- 42.Wang Y-X, Lu M, Draper D E. Biochemistry. 1993;32:12279–12282. doi: 10.1021/bi00097a002. [DOI] [PubMed] [Google Scholar]
- 43.Nicholls A, Sharp K A, Honig B. Proteins Struct Funct Genet. 1991;11:281–296. doi: 10.1002/prot.340110407. [DOI] [PubMed] [Google Scholar]