

Abstract
All cellular behaviors arise through the coordinated actions of numerous intracellular biochemical pathways. Over the past twenty years, efforts to probe intracellular biochemical processes have undergone a fundamental transformation brought about by advances in fluorescence imaging, such as the development of genetically encoded fluorescent reporters and new imaging technologies; the impact of these approaches on our understanding of the molecular underpinnings of biological function cannot be understated. In particular, the ability to obtain information on the spatiotemporal regulation of biochemical processes unfolding in real time in the native context of a living cell has crystallized the view, long a matter of speculation, that cells achieve specific biological outcomes through the imposition of spatial control over the distribution of various biomolecules, and their associated biochemical activities, within the cellular environment. Indeed, the compartmentalization of biochemical activities by cells is now known to be pervasive and to span a multitude of spatial scales, from the length of a cell to just a few enzymes. In this Perspective, part of this special issue on “Seeing into cells”, we highlight several recent imaging studies that provide detailed insights into not just where molecules are but where molecules are active within cells, offering a glimpse into the emerging view of biochemical activity architecture as a complement to the physical architecture of a cell.
Graphical Abstract
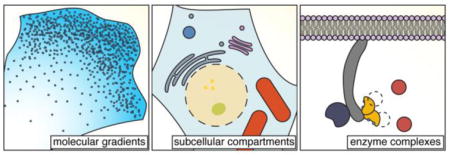
Global biochemical heterogeneity can often arise through local activity changes that are then reinforced by local and long-range feedback mechanisms, yielding biochemical activity gradients1. Approaches to directly visualize these spatial activity gradients have long included small molecule fluorescent indicators2,3 and dye-labeled proteins4,5, and the addition of genetically encoded fluorescent biosensors continues to provide important insights into the activity architecture of biochemical gradients. For example, in a recent study using FRET-based biosensors to image Ras, Rac, RhoA, and Cdc42 activity gradients6, Yang and colleagues identified Cdc42 as the chemotactic “compass” that directs cell migration in neutrophils, with local Cdc42 activity elevations not only predicting cell turning in response to a graded stimulus but also preceding spontaneous symmetry breaking and displaying the properties of an excitable medium7 in response to a uniform stimulus6. Similarly, by imaging cyclic AMP (cAMP) and protein kinase A (PKA) activity in cultured hippocampal neurons, Gorshkov et al observed that both cAMP accumulation and PKA activity were higher towards the distal region of developing axons in neurons grown for 5 days in vitro (DIV), but not in less mature, DIV3 cells8. Notably, disrupting interactions between PKA and A-kinase anchoring proteins (AKAPs) both accelerated axon growth and prompted the formation of an axon-directed cAMP gradient in DIV3 neurons, thus revealing the establishment of a developmentally timed cAMP gradient via AKAP/PKA-dependent negative feedback to control axon growth in these cells8.
Physical compartmentation of the cell imposes additional layers of spatial control over biochemical processes. Membrane-bound organelles, for instance, form functionally specialized subcellular compartments with distinct regulatory landscapes, and imaging studies have revealed that a given biochemical pathway can exhibit distinct activity patterns in association with different organelles. For example, by targeting a novel, genetically encoded mTORC1 activity reporter to different subcellular regions (e.g., plasma membrane, cytosol, lysosome, and nucleus), Zhou and colleagues observed that although growth factor stimulation induced mTORC1 signaling throughout the cell, amino acids specifically stimulated lysosomal and nuclear mTORC1 activity9. A recent study using a FRET-based Rab1 biosensor capable of cycling between the Golgi membrane and cytosol similarly revealed that Rab1 is active in the cytosol but is inactivated at the Gogli10. The ability to visualize compartment-specific biochemical activities has also modified the long-accepted model of canonical G-protein-coupled receptor (GPCR) signaling, revealing that internalized GPCRs continue to engage in G-protein-dependent signaling from endosomes11. In a recent example, Jean-Alphonse et al imaged cAMP and PKA activity using cytosol- and nuclear-targeted FRET sensors and found that β2AR activation promotes sustained cAMP signaling by endosomal parathyroid hormone receptor, leading to rapid and prolonged nuclear cAMP accumulation and PKA activity12. This nuclear PKA activity was not due to translocation of PKA from the cytosol, consistent with previous evidence that sustained, local cAMP production is required to activate nuclear-resident PKA13.
Membrane microdomains introduce further spatial differences in biochemical activity that can be visualized using selectively targeted fluorescent biosensors, as was done in a recent study by Seong and colleagues, who used a novel FRET-based biosensor to monitor PDGFR activation. By appending various lipid-modification signals to their biosensor, these authors were able to show that, although PDGF stimulation induced uniform PDGFR activity throughout the plasma membrane, PDGFR activity was uniquely antagonized by integrin-mediated tension sensing within lipid rafts14. These submicron-scale subdivisions within the plasma membrane are thought to result from in-plane, liquid-liquid phase separation by subsets of membrane lipids15 and to serve as organizing centers for various biochemical processes related to intracellular signaling16. However, membrane microdomains have long been a topic of debate, and recent observations of phase separation by membrane proteins add another layer of complexity. For instance, Banjade and Rosen observed phase separation by fluorescently tagged Nephrin inserted into a model membrane17, and Su and colleagues also visualized the formation of phase-separated domains of fluorescently tagged Linker for Activation of T cells (LAT) both in vitro and in live Jurkat cells18. Indeed, by visualizing phase-separated lipid domains in the plasma membranes of mouse B cells, Stone and colleagues recently observed that B cell receptors not only clustered within but also stabilized and expanded lipid raft-like domains19, adding to questions about whether lipid rafts drive receptor clustering or vice versa20.
Growing evidence indicates that liquid-liquid phase separation is not limited to membranes and is in fact responsible for producing discrete spatial domains of biochemical activity – sometimes called “membraneless organelles” – throughout the cell interior21. For example, Brangwynne et al previously used live-cell imaging of fluorescently tagged proteins to reveal that cytosolic P granules behave like phase-separated liquid droplets in C. elegans embryos22, and other well-known ribonucleoprotein domains such as nucleoli and Cajal bodies are also considered to be phase-separated structures21. Zeng and coworkers also recently visualized the formation of phase-separated droplets by the postsynaptic density (PSD) proteins SynGAP and PSD-9523. Such phase separation events appear to be driven by multivalent interactions among the various components21: the polyphosphorylated C-terminal tails of Nephrin24 and LAT18 with cytosolic adaptor proteins; the SynGap homotrimer with multiple copies of PSD-9523; and RNA-binding proteins through intrinsically disordered regions (IDRs), also called low-complexity domains due to their highly repetitive amino acid sequences25. Intrinsic disorder and multivalent binding modalities are common among signaling and regulatory pathways21,26, hinting at a larger role for phase separation, yet just how prevalent phase separation is in regulating intracellular biochemistry remains an open question. Furthermore, the switch-like concentration dependence of phase separation suggests an important role in all-or-none cell fate decisions, as well as other state-switching processes (e.g., actin polymerization18). It remains to be seen how phase separation fits in with other sources of nonlinear signaling dynamics27–29.
Live-cell imaging approaches are also shedding light on the spatial regulation of biochemical activity by the assembly of protein complexes containing multiple enzymes. For example, by expressing fluorescently tagged proteins in living cells, An and colleagues previously found that the six metabolic enzymes that catalyze the ten-step pathway responsible for de novo purine biosynthesis all colocalize within a multi-enzyme complex – the “purinosome” – that dynamically forms visible cytosolic clusters directly associated with purine synthesis30. In a recent study, Kohnhorst et al similarly investigated the behavior of fluorescent protein-tagged glucose metabolic enzymes in living cells and observed the formation of a “glucosome” containing four enzymes that catalyze rate-limiting steps in glycolysis and gluconeogenesis31. These complexes also formed visible clusters that were distinct from other known cytosolic bodies and whose sizes varied dynamically in response to treatments that shifted glucose flux to favor specific pathways (e.g., the pentose phosphate pathway, serine biosynthesis), suggesting a mechanism for compartmentalizing metabolic processes and controlling cellular metabolic states31. Additional imaging studies have revealed that purinosomes localize to the mitochondrial surface32 and are regulated by GPCR33 and mTOR32 signaling; biosensor-based imaging may therefore be useful in revealing additional spatial links between signaling enzyme activity and purinosome function. Purinosomes and glucosomes also bear some resemblance to phase-separated bodies34, yet whether these assemblies are driven by multivalent interactions among their protein components or by some other mechanism remains unclear.
Signaling enzyme activities are likewise spatially modulated by assembly into multi-protein complexes via scaffold proteins. PKA is well known to associate with diverse AKAPs35, and ERK/MAPK pathway components also assemble onto various scaffold proteins36. Scaffolds anchor signaling enzymes to different subcellular locations, direct activity towards specific substrates, and recruit various regulatory proteins or other signaling enzymes to produce macromolecular signalosomes with unique signaling behaviors. Direct fusion of fluorescent biosensors to scaffold proteins has been extremely useful in revealing local activity dynamics near signalosomes and continues to offer new insights into how scaffolds modulate enzyme activity37, as illustrated by Greenwald et al using a FRET-based protein kinase C (PKC) biosensor tethered to either the plasma membrane or to the C-terminus of AKAP7α, wherein AKAP-anchored PKC activity was kinetically enhanced compared with unanchored PKC38. Nevertheless, questions remain concerning whether and how AKAPs are able to spatially confine PKA activity given the classical model of PKA catalytic subunit (PKAcat) diffusion. Mo and colleagues were recently able to directly investigate this question in living cells using a novel super-resolution PKA activity reporter39. By targeting this sensor to the plasma membrane, they were able to observe discrete, ~350-nm-diameter puncta of high PKA activity in response to cAMP production. These PKA activity puncta were dependent on AKAP anchoring and also appeared to colocalize with AKAP79 clusters along the membrane, suggesting a mechanism whereby AKAP clusters increase the local concentration of PKA regulatory subunits, thereby enhancing capture of free PKAcat to locally confine PKA activity39.
As alluded to above, all biochemical processes are ultimately governed by events occurring on molecular length-scales, and researchers are increasingly turning towards more advanced imaging modalities that can be used to directly visualize biochemical activity architectures with molecular precision. In their effort to visualize PKA activity microdomains, for instance, Mo and colleagues took advantage of their discovery of a novel fluorescence phenomenon, in which molecular proximity to the green fluorescent protein Dronpa influences the fluorescence fluctuation behavior of the red fluorescent protein TagRFP-T through an effect termed fluorescence fluctuation increase by contact (FLINC), to generate a new class of fluorescent biosensors for super-resolution activity imaging, including probes for PKA activity, ERK activity, and protein-protein interactions39. Targeted biochemical perturbation strategies are also crucial for elucidating the functional importance of spatial organization, with optogenetic systems being well suited for achieving both spatial and temporal precision40. Along these lines, Shin et al have developed an “optoDroplet” system that enables the local and reversible generation of IDR-mediated phase separations in living cells41. Thus, given the success of live-cell fluorescence imaging as a tool to probe the dynamic spatial control of biochemical activities within cells, ongoing technological advances such as these promise to further transform our efforts in bringing the cell’s biochemical activity architecture to light.
Acknowledgments
This work was supported by the National Institutes of Health (R35 CA197622, R01 DK073368, and R01 MH111516 to J.Z.)
References
- 1.Dehmelt L, Bastiaens PIH. Spatial organization of intracellular communication: insights from imaging. Nat Rev Mol Cell Biol. 2010;11:1–13. doi: 10.1038/nrm2903. [DOI] [PubMed] [Google Scholar]
- 2.Brundage RA, Fogarty KE, Tuft RA, Fay FS. Calcium gradients underlying polarization and chemotaxis of eosinophils. Science. 1991;254:703–706. doi: 10.1126/science.1948048. [DOI] [PubMed] [Google Scholar]
- 3.Wei C, Wang X, Chen M, Ouyang K, Song LS, Cheng H. Calcium flickers steer cell migration. Nature. 2009;457:901–905. doi: 10.1038/nature07577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bacskai BJ, Hochner B, Mahaut-Smith M, Adams SR, Kaang BK, Kandel ER, Tsien RY. Spatially resolved dynamics of cAMP and protein kinase A subunits in Aplysia sensory neurons. Science. 1993;260:222–226. doi: 10.1126/science.7682336. [DOI] [PubMed] [Google Scholar]
- 5.Yudushkin IA, Schleifenbaum A, Kinkhabwala A, Neel BG, Schultz C, Bastiaens PIH. Live-cell imaging of enzyme-substrate interaction reveals spatial regulation of PTP1B. Science. 2007;315:115–119. doi: 10.1126/science.1134966. [DOI] [PubMed] [Google Scholar]
- 6.Yang HW, Collins SR, Meyer T. Locally excitable Cdc42 signals steer cells during chemotaxis. Nat Cell Biol. 2016;18:191–201. doi: 10.1038/ncb3292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Iglesias PA, Devreotes PN. Biased excitable networks: how cells direct motion in response to gradients. Curr Opin Cell Biol. 2012;24:245–253. doi: 10.1016/j.ceb.2011.11.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gorshkov K, Mehta S, Ramamurthy S, Ronnett GV, Zhou FQ, Zhang J. AKAP-mediated feedback control of cAMP gradients in developing hippocampal neurons. Nat Chem Biol. 2017;13:425–431. doi: 10.1038/nchembio.2298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Zhou X, Clister TL, Lowry PR, Seldin MM, Wong GW, Zhang J. Dynamic Visualization of mTORC1 Activity in Living Cells. Cell Rep. 2015;10:1767–1777. doi: 10.1016/j.celrep.2015.02.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Voss S, Krüger DM, Koch O, Wu YW. Spatiotemporal imaging of small GTPases activity in live cells. Proc Natl Acad Sci USA. 2016;113:14348–14353. doi: 10.1073/pnas.1613999113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Tsvetanova NG, Irannejad R, von Zastrow M. G protein-coupled receptor (GPCR) signaling via heterotrimeric G proteins from endosomes. J Biol Chem. 2015;290:6689–6696. doi: 10.1074/jbc.R114.617951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Jean-Alphonse FG, Wehbi VL, Chen J, Noda M, Taboas JM, Xiao K, Vilardaga JP. β2-adrenergic receptor control of endosomal PTH receptor signaling via Gβγ. Nat Chem Biol. 2017;13:259–261. doi: 10.1038/nchembio.2267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Sample V, DiPilato LM, Yang JH, Ni Q, Saucerman JJ, Zhang J. Regulation of nuclear PKA revealed by spatiotemporal manipulation of cyclic AMP. Nat Chem Biol. 2012;8:375–382. doi: 10.1038/nchembio.799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Seong J, Huang M, Sim KM, Kim H, Wang Y. FRET-based Visualization of PDGF Receptor Activation at Membrane Microdomains. Sci Rep. 2017:1–11. doi: 10.1038/s41598-017-01789-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sezgin E, Levental I, Mayor S, Eggeling C. The mystery of membrane organization: composition, regulation and roles of lipid rafts. Nat Rev Mol Cell Biol. 2017;18:361–374. doi: 10.1038/nrm.2017.16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Simons K, Simons K, Toomre D. Lipid rafts and signal transduction. Nat Rev Mol Cell Biol. 2000;1:31–39. doi: 10.1038/35036052. [DOI] [PubMed] [Google Scholar]
- 17.Banjade S, Rosen MK. Phase transitions of multivalent proteins can promote clustering of membrane receptors. Elife. 2014:3. doi: 10.7554/eLife.04123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Su X, Ditlev JA, Hui E, Xing W, Banjade S, Okrut J, King DS, Taunton J, Rosen MK, Vale RD. Phase separation of signaling molecules promotes T cell receptor signal transduction. Science. 2016;352:595–599. doi: 10.1126/science.aad9964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Stone MB, Shelby SA, Núñez MF, Wisser K, Veatch SL. Protein sorting by lipid phase-like domains supports emergent signaling function in B lymphocyte plasma membranes. Elife. 2017:6. doi: 10.7554/eLife.19891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Edidin M. The state of lipid rafts: from model membranes to cells. Annu Rev Biophys Biomol Struct. 2003;32:257–283. doi: 10.1146/annurev.biophys.32.110601.142439. [DOI] [PubMed] [Google Scholar]
- 21.Banani SF, Lee HO, Hyman AA, Rosen MK. Biomolecular condensates: organizers of cellular biochemistry. Nat Rev Mol Cell Biol. 2017;18:285–298. doi: 10.1038/nrm.2017.7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Brangwynne CP, Eckmann CR, Courson DS, Rybarska A, Hoege C, Gharakhani J, Jülicher F, Hyman AA. Germline P granules are liquid droplets that localize by controlled dissolution/condensation. Science. 2009;324:1729–1732. doi: 10.1126/science.1172046. [DOI] [PubMed] [Google Scholar]
- 23.Zeng M, Shang Y, Araki Y, Guo T, Huganir RL, Zhang M. Phase Transition in Postsynaptic Densities Underlies Formation of Synaptic Complexes and Synaptic Plasticity. Cell. 2016;166:1163–1175e12. doi: 10.1016/j.cell.2016.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Li P, Banjade S, Cheng HC, Kim S, Chen B, Guo L, Llaguno M, Hollingsworth JV, King DS, Banani SF, Russo PS, Jiang QX, Nixon BT, Rosen MK. Phase transitions in the assembly of multivalent signalling proteins. Nature. 2012;483:336–340. doi: 10.1038/nature10879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lin Y, Protter DSW, Rosen MK, Parker R. Formation and Maturation of Phase-Separated Liquid Droplets by RNA-Binding Proteins. Mol Cell. 2015;60:208–219. doi: 10.1016/j.molcel.2015.08.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Wright PE, Dyson HJ. Intrinsically disordered proteins in cellular signalling and regulation. Nat Rev Mol Cell Biol. 2015;16:18–29. doi: 10.1038/nrm3920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ferrell JE, Ha SH. Ultrasensitivity part I: Michaelian responses and zero-order ultrasensitivity. Trends Biochem Sci. 2014;39:496–503. doi: 10.1016/j.tibs.2014.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ferrell JE, Ha SH. Ultrasensitivity part II: multisite phosphorylation, stoichiometric inhibitors, and positive feedback. Trends Biochem Sci. 2014;39:556–569. doi: 10.1016/j.tibs.2014.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ferrell JE, Ha SH. Ultrasensitivity part III: cascades, bistable switches, and oscillators. Trends Biochem Sci. 2014;39:612–618. doi: 10.1016/j.tibs.2014.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.An S, Kumar R, Sheets ED, Benkovic SJ. Reversible compartmentalization of de novo purine biosynthetic complexes in living cells. Science. 2008;320:103–106. doi: 10.1126/science.1152241. [DOI] [PubMed] [Google Scholar]
- 31.Kohnhorst CL, Kyoung M, Jeon M, Schmitt DL, Kennedy EL, Ramirez J, Bracey SM, Luu BT, Russell SJ, An S. Identification of a Multienzyme Complex for Glucose Metabolism in Living Cells. J Biol Chem. 2017 doi: 10.1074/jbc.M117.783050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.French JB, Jones SA, Deng H, Pedley AM, Kim D, Chan CY, Hu H, Pugh RJ, Zhao H, Zhang Y, Huang TJ, Fang Y, Zhuang X, Benkovic SJ. Spatial colocalization and functional link of purinosomes with mitochondria. Science. 2016;351:733–737. doi: 10.1126/science.aac6054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Verrier F, An S, Ferrie AM, Sun H, Kyoung M, Deng H, Fang Y, Benkovic SJ. GPCRs regulate the assembly of a multienzyme complex for purine biosynthesis. Nat Chem Biol. 2011;7:909–915. doi: 10.1038/nchembio.690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Brangwynne CP. Phase transitions and size scaling of membrane-less organelles. J Cell Biol. 2013;203:875–881. doi: 10.1083/jcb.201308087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wong W, Scott JD. AKAP signalling complexes: focal points in space and time. Nat Rev Mol Cell Biol. 2004;5:959–970. doi: 10.1038/nrm1527. [DOI] [PubMed] [Google Scholar]
- 36.Kolch W. Coordinating ERK/MAPK signalling through scaffolds and inhibitors. Nat Rev Mol Cell Biol. 2005;6:827–837. doi: 10.1038/nrm1743. [DOI] [PubMed] [Google Scholar]
- 37.Tenner B, Mehta S, Zhang J. Optical sensors to gain mechanistic insights into signaling assemblies. Curr Opin Struct Biol. 2016;41:203–210. doi: 10.1016/j.sbi.2016.07.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Greenwald EC, Redden JM, Dodge-Kafka KL, Saucerman JJ. Scaffold state switching amplifies, accelerates, and insulates protein kinase C signaling. J Biol Chem. 2014;289:2353–2360. doi: 10.1074/jbc.M113.497941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Mo GCH, Ross B, Hertel F, Manna P, Yang X, Greenwald E, Booth C, Plummer AM, Tenner B, Chen Z, Wang Y, Kennedy EJ, Cole PA, Fleming KG, Palmer A, Jimenez R, Xiao J, Dedecker P, Zhang J. Genetically encoded biosensors for visualizing live-cell biochemical activity at super-resolution. Nat Methods. 2017;14:427–434. doi: 10.1038/nmeth.4221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zhang K, Cui B. Optogenetic control of intracellular signaling pathways. Trends in Biotechnology. 2015;33:92–100. doi: 10.1016/j.tibtech.2014.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Shin Y, Berry J, Pannucci N, Haataja MP, Toettcher JE, Brangwynne CP. Spatiotemporal Control of Intracellular Phase Transitions Using Light-Activated optoDroplets. Cell. 2017;168:159–171e14. doi: 10.1016/j.cell.2016.11.054. [DOI] [PMC free article] [PubMed] [Google Scholar]