

Abstract
The mechanistic basis for chloroquine resistance (CQR) in Plasmodium falciparum recently has been linked to the polymorphic gene pfcrt. Alleles associated with CQR in natural parasite isolates harbor threonine (T), as opposed to lysine (K) at amino acid 76. P. falciparum CQR strains of African and Southeast Asian origin carry pfcrt alleles encoding an amino acid haplotype of CVIET (residues 72–76), whereas most South American CQR strains studied carry an allele encoding an SVMNT haplotype; chloroquine-sensitive strains from malarious regions around the world carry a CVMNK haplotype. Upon investigating the origin of pfcrt alleles in Papua New Guinean (PNG) P. falciparum we found either the chloroquine-sensitive-associated CVMNK or CQR-associated SVMNT haplotypes previously seen in Brazilian isolates. Remarkably we did not find the CVIET haplotype observed in CQR strains from Southeast Asian regions more proximal to PNG. Further we found a previously undescribed CQR phenotype to be associated with the SVMNT haplotype from PNG and South America. This CQR phenotype is significantly less responsive to verapamil chemosensitization compared with the effect associated with the CVIET haplotype. Consistent with this, we observed that verapamil treatment of P. falciparum isolates carrying pfcrt SVMNT is associated with an attenuated increase in digestive vacuole pH relative to CVIET pfcrt-carrying isolates. These data suggest a key role for pH-dependent changes in hematin receptor concentration in the P. falciparum CQR mechanism. Our findings also suggest that P. falciparum CQR has arisen through multiple evolutionary pathways associated with pfcrt K76T.
Public health of children in developing tropical countries is severely challenged by malaria (1). Plasmodium falciparum can impose life-threatening elements of disease before birth by compromising fetal development and maternal health (1–5). The threat of malaria continues through at least the first 5 years of life before most children living in endemic regions develop immunity sufficient to suppress severe pathogenesis (6–10). Because antimalarial treatment by chloroquine (CQ) is well tolerated by children and pregnant women (11, 12), the steady spread of CQ resistance (CQR) throughout malaria-endemic regions is a tragic setback.
The molecular details contributing to CQR in P. falciparum are undecided, yet resistant parasites are known to accumulate lower levels of CQ, and diverse agents including verapamil (VPL) can reverse or attenuate the CQR phenotype (13–17). Similar decreased drug accumulation and VPL reversal are features of drug resistance observed in other systems, notably mammalian tumor cells (18). Hypotheses directing investigations of the CQR mechanism include modifications in drug or ion transport through parasitized erythrocyte membranes (19–22), altered binding of CQ to its target (heme), or detoxification of heme/heme-CQ conjugates (23–27). Recent studies also suggest that digestive vacuole (DV) pH influences CQ accumulation, and thereby susceptibility (28, 29). Similar mechanistic factors are central to hypotheses proposed to explain anticancer drug resistance (18).
Analysis of a genetic cross between the CQR (Dd2) and CQ-sensitive (CQS) (HB3) P. falciparum clones (30) has provided evidence indicating that CQR can be localized to a 36-kb segment of the parasite chromosome 7 (31) and culminated in the identification of the pfcrt (P. falciparum CQR transporter) gene, residing in this chromosome 7 segment (32). Eight point mutations in this gene distinguish CQR from CQS progeny of the Dd2 × HB3 cross, including a threonine (T) to lysine (K) substitution at residue 76. Although the K76T polymorphism is observed within different amino acid haplotypes (CVIET, CVMNT, CVMET, or SVMNT residues 72–76), molecular surveys of multiple laboratory-adapted field isolates have found that this K76T mutation is present in all CQR strains, regardless of geographic origin (32).
Recent investigations of pfcrt polymorphisms vs. in vivo CQ susceptibility have shown a 100% prevalence of the pfcrt 76T allele in recurrent or persistent P. falciparum isolates from malaria-endemic regions of Africa and Southeast Asia (SEA) (33–36). This allele also was found in some CQ-treated individuals that were able to clear the parasite infection. These results suggest that while all cases of P. falciparum CQ treatment failure are associated with pfcrt 76T, the in vivo outcome also depends on other factors that may include a patient's acquired immunity and/or additional parasite genetic factors that augment in vivo resistance.
Here we assess the evolution of pfcrt polymorphism in P. falciparum from malaria-holoendemic regions of Papua New Guinea (PNG) where P. falciparum CQR has been reported (37–39). We also examine polymorphisms in pfmdr1 (P. falciparum multidrug resistance 1; refs. 40 and 41), whose product is homologous to mammalian P-glycoproteins implicated in tumor drug resistance (18), as these polymorphisms have been associated with CQR in some but not all studies (42).
Methods
Study Sites.
P. falciparum field samples were obtained from malaria-exposed study subjects living in three different malaria holoendemic regions of PNG. Surveys included 280 individuals from the Dreikikir (rainforest-Prince Alexander foothills) and 432 individuals from the Wosera (grassland/marsh-Screw River flood plain) regions of East Sepik Province, and 190 individuals from the Liksul (coastal rainforest) region of Madang Province. Entomological inoculation rates (the average number of infective bites/person per night; refs. 43–46) were 0.9 for the Dreikikir region, 0.15 for the Wosera region, and 0.7 for the Liksul/Madang region (43–46). The percentage of health centers with essential drugs (including CQ) was 68% for the Dreikikir region, 84% for the Wosera region, and 92% for the Liksul/Madang region and approximates differences in CQ availability/usage in the three study sites (47). Blood samples from all individuals participating in this study were collected under clinical protocols approved by institutional review boards of The University Hospitals of Cleveland and PNG.
P. falciparum Laboratory Strains and Genomic DNAs.
These included 7G8, RCS, S40/88, 306 (Brazil); NIG60, NIG82, 91566, D6 (Africa); K1, TM346, GA3, VS-1 (SEA); W2, Dd2 (Indochina); HB3 (Honduras) (provided by MR4, American Type Culture Collection), and 1775, 1776, 1787, 1904, 1905, 1917, 1933, 1934, and 1935 (Madang, PNG) (provided by Alan Cowman, Walter and Eliza Hall Institute, Melbourne, Australia).
Parasite Culture and CQ Response Assays.
Parasite strains (W2, D6, RCS, S40/88, 306, NIG60, NIG82, 91566, TM346, GA3, VS-1, TM90-C235, FCB, and the PNG strains 1905 and 1917) were propagated in vitro in human erythrocytes (48). CQ response assays were performed by monitoring [3H] hypoxanthine uptake (49, 50). Partial reversal of CQR was observed by addition of 0.6 μM VPL to parasite cultures analyzed in parallel. To assess further the effects of VPL on the CQ IC50, parasites were cultured in medium containing CQ alone and CQ plus VPL across a range of VPL concentrations (2-fold dilutions from 1.0 μM to 0.016 μM). The relative CQR reversal effects of VPL were compared among parasite strains expressed as a response modification index = CQ IC50 with VPL/CQ IC50 without VPL (17).
DV pH Analysis.
DV pH was analyzed as described (28, 29). The six strains examined (see Results) were coded (by P.A.Z.), and measurements were performed in double-blind fashion. After data analysis was complete, the code was revealed. Cultures of intraerythrocytic P. falciparum, synchronized at the late trophozoite stage of development, were attached to thin glass substrate via polylysine. These preparations were then perfused with physiologic buffer balanced with 24 mM HCO3−/5% CO2 containing 1 μM acridine orange (AO) on the stage of an epifluorescence microscope used in a customized single cell photometry apparatus (28, 29). Previous studies using this methodology (28) localized AO primarily to the DV. Cells were kept under constant perfusion. Once the DV AO staining had reached a plateau (<10 min), perfusate was rapidly changed to identical perfusate harboring 0.8 μM VPL. DV AO fluorescence was monitored on-line (29), and the percent change in AO signal was calculated as described (29). Experiments were performed for 7–13 glass coverslips for each strain (n = 56–107 parasites), and AO signals were monitored independently for individual intraerythrocytic parasites by using dynamic thresholding methods as described (28).
DNA Template Preparation.
Genomic DNA was extracted from whole blood (200 μl) of PNG study subjects or from P. falciparum cultures, using QIAamp 96 or individual spin blood kits (Qiagen, Valencia, CA). Alternatively, DNA preparations were extracted from agarose gel slices using a Geneclean Spin kit (Bio 101).
PCR Analysis.
PCRs were performed as described (51). For analyses requiring nested PCR amplification, a 3-μl aliquot of the nest 1 reaction was used as template in nest 2 reactions. PCR assays were performed by using a PTC-225 Peltier Thermal Cycler (MJ Research, Watertown, MA). PCR products from nest 1 and nest 2 amplifications were electrophoresed in 2% agarose gels, stained with SYBR Gold (Molecular Probes), and visualized on a Storm 860 imaging system with imagequant software (Molecular Dynamics).
PCR amplification of pfcrt and pfmdr1-specific sequences were performed by using primers and strategies identified in Tables 3 and 4, which are published as supporting information on the PNAS web site, www.pnas.org. Nomenclature for PCR primers is based on nucleotide sequence numbering derived from GenBank accession nos. AF030694 (31) and X56851 (52), respectively.
PCR amplifications for P. falciparum microsatellite loci (Pfg377, Pf-PK2, TA81, and TA109) and the cg2 omega repeat region were performed as described (53, 54) with the modification that all labeled primers for each second-round, seminested amplification reaction were 5′ end-labeled with Cy5 (Research Genetics, Huntsville, AL). P. falciparum strain-specific microsatellite polymorphisms were evaluated after electrophoresis on 6% denaturing polyacrylamide gels and fluorescence imaging on a Storm 860 (Molecular Dynamics).
Direct DNA Sequence Analysis.
The PCR amplification products were purified by using a QIAquick PCR purification kit (Qiagen) and directly sequenced on an ABI377 automated sequencer using fluorescent dye-terminator chemistry.
Sequence-Specific Oligonucleotide Probe (SSOP) Hybridization Assays.
Dot blot preparation and SSOP hybridization were performed as described (55). High stringency washing conditions for pfcrt and pfmdr1 SSOPs are indicated in Tables 3 and 4. Fluorescence hybridization signal was detected by using the Storm 860.
Statistical Analysis.
Statistical analyses were performed by using STATVIEW 5.0.1 (Abacus Concepts, Berkeley, CA).
Results
DNA Sequence Analysis.
Our investigation of the local evolution of CQR in PNG began with direct DNA sequence analysis of a 610-bp pfcrt, exon 2-specific amplicon spanning molecular polymorphisms in codons 72–76. Analysis of five P. falciparum-infected individuals from Dreikikir revealed three sequences predicted to encode amino acids characterizing the CQR-associated, SVMNT (codons 72–76) haplotype, observed previously only in Brazilian P. falciparum strains (32). None of the five sequences were of the CVIET haplotype, observed in CQR African and SEA P. falciparum strains (32). Two sequences represented the CVMNK allele, observed worldwide in CQS P. falciparum strains (32).
P. falciparum Genotyping Studies.
SSOPs designed to hybridize specifically to the SVMNT, CVIET, or CVMNK allelic sequences were used to perform a broad survey of the P. falciparum populations in the three different malaria-endemic PNG study regions. Results of post-PCR SSOP hybridization analysis on 902 field samples showed that among the 434 pfcrt PCR-positive samples 58.3% hybridized to the SVMNT probe only, 24% hybridized to the CVMNK probe only, and 17.7% hybridized to both SVMNT and CVMNK probes; no samples hybridized to the CVIET probe. These results, based on pfcrt sequence polymorphism, suggest greater similarity between PNG and South America (SA) (New World) compared with African/SEA (Old World) parasites.
As molecular polymorphism in the pfmdr1 gene also has been associated with CQ susceptibility in P. falciparum in some, but not all, studies (42), a PCR-SSOP hybridization assay was developed to genotype single nucleotide polymorphisms encoding amino acid substitutions at codons 86 (N/Y), 184 (Y/F), 1034 (S/C), 1042 (N/D), and 1246 (D/Y). From past studies the 86Y sequence and downstream 1034–1042-1246 haplotype CDY have shown the most frequent association with CQR in Old World as compared with New World parasites, respectively (refs. 31 and 40; Table 5, which is published as supporting information on the PNAS web site); 86N and the 1034–1042-1246 haplotypes SND and SDD have been associated with CQS throughout malaria-endemic regions of the world. PCR-SSOP hybridization analysis (at codons 86, 1034, 1042, and 1246) was successful for 401 samples. The CQR-associated 86Y polymorphism was observed alone in 63.3% of the PCR-positive samples, whereas 86N alone was observed in 18.5%, and a mixture of 86Y and 86N was observed in 18.2% of the PCR-positive samples. The CQS-associated SND or SDD haplotypes accounted for all of the downstream polymorphism (SND alone = 95.1%, SDD alone = 0.3%, SND and SDD = 4.6%); no samples produced PCR-SSOP hybridization data consistent with the CDY CQR-associated haplotype. In contrast to the pfcrt results described above, results based on pfmdr1 sequence polymorphism suggest greater similarity between PNG and African/SEA (Old World) compared with SA (New World) parasites.
Further analysis of the survey results for each of the three PNG study sites showed (Fig. 1 Left) that the prevalence of the CQR-associated, pfcrt SVMNT haplotype (alone; mixed SVMNT + CVMNK assemblage) was highest in Liksul [83.0% (78/94); 10.6% (10/94)] compared with Dreikikir [40.1% (61/152); 34.9% (53/152)] and the Wosera [60.6% (114/188); 7.5% (14/188)] study areas. In contrast, prevalence of the CQS-associated, pfcrt CVMNK haplotype alone was higher in the Wosera [31.9% (60/188)] and Dreikikir [25.0% (38/152)] compared with Liksul [6.4% (16/94)]. A similar distribution pattern was observed for the pfmdr1 CQR- and CQS-associated polymorphisms (Fig. 1 Right), where 86Y (alone; mixed Y + N assemblage) was most prevalent in Liksul [87.9% (80/91); 3.3% (3/91)] compared with Dreikikir [46.9% (61/130); 33.9% (44/130)] and the Wosera [62.8% (113/180); 14.4% (26/180)] whereas 86N alone was more prevalent in the Wosera [22.8% (41/180)] and Dreikikir [19.2% (25/130)] compared with Liksul [8.8% (8/91)].
Figure 1.

Frequency distributions of pfcrt (Left) and pfmdr1 (Right) alleles in P. falciparum-infected individuals from three PNG field sites. Data reflect the results of DNA probe hybridization experiments conducted using methods described. For the pfcrt analyses the number of P. falciparum-infected individuals by village was Liksul = 94, Dreikikir = 152, and Wosera = 188. For the pfmdr1 analyses the number of P. falciparum-infected individuals by village was Liksul = 91, Dreikikir = 130, and Wosera = 180.
Interestingly, although pfcrt is located on P. falciparum chromosome 7 and pfmdr1 is located on chromosome 5, when results of intraisolate associations between pfcrt and pfmdr1 alleles were evaluated for 300 samples producing genotype results for both genetic loci, significant associations were observed between pfcrt SVMNT and pfmdr1 86Y and between pfcrt CVMNK and pfmdr1 86N alleles (Table 1) (χ2, 4 df = 52.3, P < 0.001).
Table 1.
Comparing intra isolate associations of pfcrt and pfmdr1 alleles
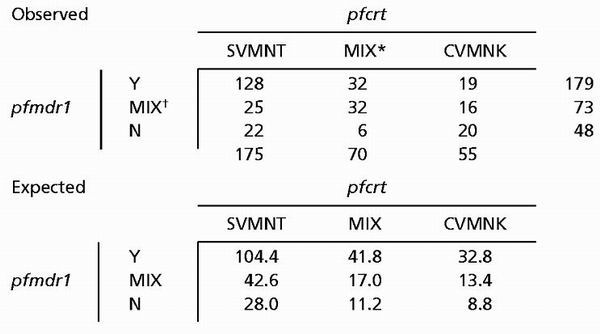 |
MIX = pfcrt SVMNT and CVMNK haplotypes.
MIX = pfmdr1 86Y and 86N polymorphisms.
Correlating pfcrt and pfmdr1 Genotypes and in Vitro CQ Response in PNG P. falciparum Strains.
As the genotyping surveys reported above were performed on archived human blood samples from which live parasites could not be recovered, it was not possible to evaluate CQ IC50 values on those parasites. To test the association between pfcrt and pfmdr1 genotypes and in vitro CQ response in PNG P. falciparum strains, pfcrt genotyping was performed on genomic DNA preparations from nine laboratory culture-adapted PNG strains with reported CQ IC50 values (40). Consistent with our larger population study we detected SVMNT and CVMNK, but not CVIET, haplotypes. Further results showed that the SVMNT haplotype correlated with CQ IC50 values greater than 50 ng/ml in five of the PNG strains (1775, 1776, 1904, 1933, and 1935); pfmdr1 genotyping showed that each of these PNG P. falciparum strains carried the 86Y, CQR-associated polymorphism. The correlation between the pfcrt CVMNK and pfmdr1 86N sequences with a CQ IC50 value of 10 ng/ml was observed in the PNG strain 1787.
Two exceptional results involved PNG strains 1905 and 1917. The 1905 strain did not hybridize to any of the pfcrt allele-specific probes previously identified. When DNA sequence analysis was performed on the pfcrt region 1 amplicon of this strain, a di-nucleotide polymorphism was observed in codon 72, changing the sequence from AGT to TCT. When an SSOP, based on the PNG 1905 allele, was used to genotype samples from the Dreikikir (n = 100), Wosera (n = 100), and Liksul (n = 200) areas no samples were observed to hybridize to this probe (data not shown). This DNA sequence polymorphism does not lead to an amino acid substitution (both AGT and TCT encode serine). Interestingly, 1905 has been the only PNG strain observed so far to carry the CQR-associated pfmdr1 CDY haplotype, observed in the Brazilian P. falciparum strain, 7G8 (ref. 40; Table 5). The 1917 strain was previously reported to have a relatively low CQ IC50 of 19 ng/ml. The SSOP hybridization results for this strain identified the pfcrt SVMNT (CQR) and pfmdr1 NSND (CQS; residues 86, 1034, 1042, and 1246) haplotypes.
In Vitro Analysis of the PNG 1905 and 1917 P. falciparum Strains.
Standard drug response assays were performed to assess the CQ IC50s of 1905 and 1917 in comparison to those of W2 (CQR) and D6 (CQS) strains (initial parasitemias for all cultures were between 0.7% and 0.9%). Results of these in vitro assays showed that the CQ IC50s without and with 0.6 μM VPL were 103.1 ng/ml (SE ± 8.2 ng/ml) and 100.5 ng/ml (SE ± 16.9 ng/ml) for strain 1905 and 49.6 ng/ml (SE ± 5.7 ng/ml) and 60.5 ng/ml (SE ± 14.4 ng/ml) for strain 1917; corresponding CQ IC50 values for the P. falciparum reference strains were 132.3 ng/ml (SE ± 12.3 ng/ml) and 84.4 ng/ml (SE ± 15.4 ng/ml) for the CQR strain, W2, and 5.0 ng/ml (SE ± 0.7 ng/ml) and 4.7 ng/ml (SE ± 0.44 ng/ml) for the CQS strain, D6. Results in Fig. 2 show that the PNG 1905 and 1917 strains obtained for further in vitro study produced identical PCR-SSOP hybridization results compared with those performed on archived genomic DNA samples.
Figure 2.

pfcrt DNA sequence-specific probe (SSOP) hybridization results performed on control CQS and CQR P. falciparum strains and two PNG isolates. P. falciparum laboratory strains analyzed include HB3, Dd2, K1, and 7G8. DNA sequence analysis of pfcrt from HB3, Dd2, K1, and 7G8 has shown these strains to carry alleles encoding the CVMNK, CVIET, CVIET, and SVMNT haplotypes, respectively. The SVMNT and SVMNT* SSOPs differ as a result of synonymous DNA sequence polymorphism in the serine codon, 5-′TAAGTGTAATGAATACA-3′ vs. 5′-TATCTGTAATGAATACA-3′, respectively.
The surprising lower sensitivity of the in vitro 1905 and 1917 CQR phenotypes to VPL reversal was reminiscent of previous observations from SA CQR P. falciparum strains (D.E.K., unpublished data). To investigate these potential in vitro similarities, parasites were exposed to CQ alone or CQ and VPL across a 2-fold dilution series of VPL (1.0–0.016 μM). Comparisons of the CQ response modification index were performed for the two PNG strains 1905 and 1917, three CQR P. falciparum SA strains (S40/88, 306, RCS), three CQR P. falciparum African strains (NIG60, NIG82, 91566), and four CQR P. falciparum SEA strains (W2, TM346, GA3, VS-1). Results summarized in Fig. 3 show that the African and SEA P. falciparum strains were significantly more responsive to VPL reversal of CQR than were the SA and PNG strains (ANOVA, 10 residual df, all P < 0.05 at each [VPL]). Moreover, the lower sensitivity to VPL observed for the PNG CQR strains was not significantly different from that observed for the SA CQR strains.
Figure 3.

VPL reversal of CQ IC50s for African, SEA, PNG, and SA CQR P. falciparum strains. CQR P. falciparum strains used in this study included 1905, 1917 (PNG); S40/88, 306, RCS (SA); NIG60, NIG82, 91566 (Africa), and W2, TM346, GA3, and VS-1 (SEA). Parasites were cultured in medium containing CQ alone and CQ plus VPL across a range of VPL concentrations (2-fold dilutions from 1.0 to 0.016 μM). The CQR reversal effects of VPL are expressed based on a response modification index = CQ IC50 with VPL/CQ IC50 without VPL. ANOVA was used to compare PNG/SA strains with African/SEA strains, resulting in the following P values for each respective VPL: 0.047 at 0.0625 ng/ml, 0.0034 at 0.125 ng/ml, <0.0001 at 0.25 ng/ml, 0.5 ng/ml, and 1.0 ng/ml. Application of the Bonferroni correction factor for five comparisons (five [VPL]) required P < 0.01 to achieve statistical significance.
Recent studies suggest that VPL treatment induces an increase in DV pH for CQR Dd2 but not for CQS HB3 (29) that can be monitored via a decrease in DV AO intensity. Given the unique dissociation between CQR and VPL reversal in PNG and SA parasite strains we compared changes in DV AO intensity in response to VPL for CQR P. falciparum strains carrying SVMNT pfcrt [1905, 1917 (PNG); RCS (SA)] vs. CVIET pfcrt (W2, TM90, FCB) haplotypes (Fig. 4 and Table 2). VPL treatment did not significantly change the net DV pH for populations of SVMNT-carrying parasites, whereas populations of CVIET-carrying parasites exhibited a decrease in AO intensity, consistent with alkalinization of the DV and reversal of CQR (28, 29).
Figure 4.

Percent change DV AO fluorescence induced by VPL treatment in CQR strains carrying SVMNT vs. CVIET pfcrt haplotypes. Parasites were cultured in medium without VPL, attached to glass substrate, placed within the perfusion cell of a custom single cell photometry apparatus, and continuously perfused with physiologically balanced buffer containing 1 μM AO as described (28, 29). The change in DV AO signal upon changing to a similar perfusate plus 0.8 μM VPL was monitored for individual parasites and is expressed as a percent of the initial signal. For clarity, only data for populations of 1917 (○) and W2 (■) are shown; data for FCB and TM90-C235 were similar to those shown for W2; 1905 and RCS were similar to 1917 (Table 2). Numbers shown next to the symbols are the mean values for all parasites within a given range (0–5%, 5–10%, etc.). The vertical dashed line indicates 0% change. A decrease in net DV AO signal is indicative of DV alkalinization (28, 29). Thus, DV pH changes are not necessarily uniform among populations of parasites (even for these synchronized cultures) but are (on average) more pronounced for CVIET strains than SVMNT strains. The mean % change calculated for DV AO fluorescence for all six strains is presented in Table 2.
Table 2.
Comparing DV AO fluorescence for pfcrt SVMNT and CVIET-carrying CQR P. falciparum strains
| P. falciparum strain | pfcrt haplotype | ΔDV AOF* | No. parasites† |
|---|---|---|---|
| RCS (SA) | SVMNT | −0.14 | 82 |
| 1917 (PNG) | SVMNT | −0.35 | 82 |
| 1905 (PNG) | SVMNT | −0.72 | 93 |
| W2 (Indochina) | CVIET | −9.93 | 107 |
| FCB (SEA) | CVIET | −10.6 | 56 |
| TM90-C235 (Thailand) | CVIET | −6.3 | 93 |
Mean percentage change in DV AO fluorescence.
Number of late-stage trophozoites analyzed for ΔDV AO fluorescence.
As new PfCRT amino acid sequence polymorphisms might be responsible for this unusual dissociation between the in vitro CQR phenotype and VPL reversibility, further DNA sequence analysis was performed. Results showed that all polymorphisms with PNG 1905 and 1917 pfcrt alleles were identical to those recently described for the SA 7G8 CQR P. falciparum strain (72S, 74 M, 75N, 76T, 97H, 220S, 271Q, 326D, and 371R).
Finally, to rule out the possibility that the unique in vitro CQR phenotype characterizing the PNG 1905 and 1917 strains might be based on a mixture of P. falciparum strains, species-specific microsatellite genotyping was performed. Results for five different microsatellite loci (TA81, chromosome 5; TA109, chromosome 6; cg2, chromosome 7; Pfg377, chromosome 12; Pf-PK2, chromosome 12) showed only single and not multiple allelic amplicons, consistent with strain homogeneity (data not shown).
Discussion
Our population genetic survey of three different malaria holoendemic regions of PNG suggests a surprising closer relationship between PNG and SA (specifically Brazilian) CQR P. falciparum isolates carrying the pfcrt SVMNT haplotype than more proximal SEA CQR parasite isolates carrying the CVIET haplotype. In contrast, analysis of pfmdr1 polymorphism suggests that PNG and SEA CQR P. falciparum isolates carrying the YSND haplotype are more closely related compared with SA isolates that carry the NCDY haplotype. Recent multiple locus microsatellite studies by Anderson et al. (56) illustrating greater evolutionary affinity between PNG and SEA as opposed to SA P. falciparum isolates further emphasize the unexpected nature of the findings based on pfcrt polymorphism.
To explain our findings a number of potential mechanisms using recombination or de novo point mutations may be considered. Genetic recombination may have been responsible for introducing the SA pfcrt CQR haplotype onto a PNG CQS haplotype. However, if a recombination model of this nature is used to explain our observations we would have anticipated detecting SA markers at flanking intrachromosomal and/or interchromosomal P. falciparum genetic loci. Our population-based surveys in PNG did not detect the SA pfmdr1 NCDY haplotype in 902 field isolates; PNG 1905 is the only isolate that has been observed to date to carry both pfcrt and pfmdr1 haplotypes characteristic of SA P. falciparum. Alternatively, the PNG pfcrt CQR haplotype could have evolved independently of SA origin. To produce the overall pfcrt haplotype observed in PNG 1917 from the CQS allele would require amino acid substitutions at four positions (C to S at residue 72, K to T at 76, A to S at 220, N to D at 326). Evolution of the SVMNT haplotype by this model would occur in a context exclusive of SA genetic polymorphism and is more consistent with our data as SA pfmdr1 polymorphism was not observed in our field isolates. If this model is shown to be correct, our findings in PNG would emphasize strongly the importance of pfcrt sequence evolution as a major factor contributing to CQR. Finally, it is possible that the SVMNT haplotype distributed throughout the three northern PNG malaria-endemic regions studied here could have arisen through either recombination or de novo point mutations on the SEA CQR (CVIET) pfcrt haplotype. For either of these models to be correct we would expect to observe some evidence of the CVIET haplotype, unless it has been completely replaced by the SVMNT haplotype.
Interestingly, our population genetic observations provide evidence for epistatic interactions between pfcrt and pfmdr1 alleles and CQ susceptibility. Because these genetic loci appear on two different chromosomes (pfcrt, chromosome 7; pfmdr1, chromosome 5), independent assortment of heritable traits is expected (57). With this in mind, it is interesting that P. falciparum isolates emanating from three different PNG locations illustrate significant association between pfcrt and pfmdr1 CQR and between pfcrt and pfmdr1 CQS-associated alleles. Consistent with these observations, an analysis of more than 40 P. falciparum strains characterized from malaria-endemic regions throughout the tropics revealed significant association between pfcrt and pfmdr1 CQR- and CQS-associated alleles, and their in vitro CQ responsiveness [heterogeneity test (2 rows × 4 columns) 3 df = 23.8, P < 0.001; Table 6, which is published as supporting information on the PNAS web site]. Furthermore, a recent study of uncomplicated malaria cases in Gambia (54) observed strong associations between in vitro CQR, pfmdr1 86Y, and the cg2 omega repeat allele from CQR P. falciparum strains (cg2 is located within 10 kb of pfcrt; ref. 31). Overall these observations suggest that CQ selection pressure is likely to have played a significant role in the observed linkage disequilibrium between genetic markers associated with CQR in pfmdr1 and those recently described in pfcrt and nearby cg2 (54).
In addition to our population genetic observations, we have described a unique CQR phenotype exhibiting lower chemosensitivity to VPL associated specifically with the SVMNT haplotype of both PNG and SA P. falciparum isolates (Fig. 3 and D.E.K., L.G., A.O., R.K.M., and N.K.M., unpublished work). Additionally, whereas both pfcrt CQR haplotypes (CVIET and SVMNT) are associated with lower DV pH when compared with the CVMNK haplotype (O.J., L.M.B.U., D.E.K., P.A.Z., and P.D.R., unpublished work), only the CVIET haplotype (but not SVMNT) acquired increased DV pH after VPL treatment (Table 2, Fig. 4). As suggested previously, increased DV pH that occurs with VPL treatment likely increases the solubility of hematin, leading to an increased concentration of toxic hematin-CQ conjugates. Consistent with this model, as the mean DV pH is not observed to increase in SVMNT-carrying parasites, increased production of toxic hematin-CQ conjugates would not be predicted to occur, thus VPL would not attenuate sensitivity to CQ. Although it may be possible in the near future to determine which PfCRT amino acid substitutions contribute to the observed VPL sensitivity differences associated with the CVIET and SVMNT haplotypes upon optimization of full and partial allele replacement strategies, these methodologies were not available to this study.
We have demonstrated that effects of VPL on reversal of drug resistance have been associated with polymorphisms in a gene outside the ABC transporter superfamily (18). Recall that the attenuated VPL reversal of CQR observed here occurred in the PNG 1905 and 1917 strains that carry different pfmdr1 haplotypes, NCDY and NSND, respectively. Therefore, this study suggests that additional proteins relevant to chemoreversal of drug resistance in microbial through mammalian systems have yet to be discovered.
CQR P. falciparum has been documented over the past 40 years (58). The recent discovery of a new genetic marker associated with CQR in P. falciparum has the potential to improve assessment of CQ efficacy in holoendemic regions where malaria is an important cause of childhood mortality (9). Results from this study showing that PfCRT 76T is the predominant allele in three different study sites in PNG provokes obvious concern that CQ is no longer an effective treatment against P. falciparum malaria and concur with recent policy changes by the PNG Ministry of Health recommending that treatment for malaria include CQ in combination with Fansidar. With the uncertainty surrounding malaria vaccine development and growing resistance by the parasite to the small number of effective antimalarial compounds, it becomes increasingly important to define the mechanisms conferring resistance to current drugs. This effort will be strengthened by further clarification of proteins involved in drug action. With this knowledge it may become possible to design future drugs to work despite the acquisition of genetic polymorphism conferring resistance to known antimalarial drugs or to develop drugs that will avoid known resistance mechanisms.
Supplementary Material
Acknowledgments
We thank Drs. J. Reeder, T. J. C. Anderson, L. T. Webster, Jr., M. Jacobs-Lorena, K. P. Day, A. F. Cowman, and T. E. Wellems for discussions leading to the completion of this study and W. Kastens and Dr. C. King for samples collection. We thank Cleveland Genomics and Dr. A. B. S. Sidhu (Einstein) for DNA sequence analysis, L. Gerena (drug susceptibility testing), and D. Tisch (statistical methodology) for outstanding technical input. Finally, we thank all of the study volunteers for their willing participation. Financial support for these studies was provided by the National Institutes of Health (Grants AI-142367–01 and AI-36478–04S1 to J.W.K. and Grant AI 45957–01 to P.D.R.), the Burroughs-Wellcome Fund (to J.W.K. and P.D.R.), the Fogarty International Center (to R.K.M.), and the Department of Defense Global Emerging Infections Surveillance and Response System (to D.E.K.).
Abbreviations
- CQ
chloroquine
- CQR
CQ resistance
- CQS
CQ sensitive
- SEA
Southeast Asia(n)
- SA
South America(n)
- PNG
Papua New Guinea(n)
- VPL
verapamil
- DV
digestive vacuole
- AO
acridine orange
- SSOP
sequence-specific oligonucleotide probe
References
- 1.WHO Expert Committee on Malaria. WHO Technical Report Series 892. Geneva: WHO; 2000. [PubMed] [Google Scholar]
- 2.McGregor I A, Wilson M E, Billewicz W Z. Trans R Soc Trop Med Hyg. 1983;77:232–244. doi: 10.1016/0035-9203(83)90081-0. [DOI] [PubMed] [Google Scholar]
- 3.Steketee R W, Wirima J J, Slutsker L, Haymann D L, Breman J G. Am J Trop Med Hyg. 1996;55, Suppl. 1:2–7. doi: 10.4269/ajtmh.1996.55.2. [DOI] [PubMed] [Google Scholar]
- 4.Steketee R W, Wirima J J, Hightower A W, Slutsker L, Haymann D L, Breman J G. Am J Trop Med Hyg. 1996;55, Suppl. 1:33–41. doi: 10.4269/ajtmh.1996.55.33. [DOI] [PubMed] [Google Scholar]
- 5.McDermott J M, Wirima J J, Steketee R W, Breman J G, Haymann D L. Am J Trop Med Hyg. 1996;55, Suppl. 1:61–65. doi: 10.4269/ajtmh.1996.55.61. [DOI] [PubMed] [Google Scholar]
- 6.Bruce-Chwatt L J. Ann Trop Med Parasitol. 1952;46:173–200. doi: 10.1080/00034983.1952.11685522. [DOI] [PubMed] [Google Scholar]
- 7.Greenwood B M, Bradley A K, Greenwood A, M, Byass P, Jammeh K, Marsh K, Tulloch S, Oldfield F S, Hayes R. Trans R Soc Trop Med Hyg. 1987;81:478–486. doi: 10.1016/0035-9203(87)90170-2. [DOI] [PubMed] [Google Scholar]
- 8.Genton B, Al-Yaman F, Beck H-P, Hii J, Mellor S, Marara A, Gibson N, Smith T, Alpers M P. Ann Trop Med Parasitol. 1995;89:359–376. doi: 10.1080/00034983.1995.11812965. [DOI] [PubMed] [Google Scholar]
- 9.Genton B, Al-Yaman F, Beck H-P, Hii J, Mellor S, Rare L, Ginny M, Smith T, Alpers M P. Ann Trop Med Parasitol. 1995;89:377–390. doi: 10.1080/00034983.1995.11812966. [DOI] [PubMed] [Google Scholar]
- 10.Baird J K. Ann Trop Med Parasitol. 1998;92:367–390. doi: 10.1080/00034989859366. [DOI] [PubMed] [Google Scholar]
- 11.WHO Expert Committee on Malaria. WHO Technical Report Series 735. Geneva: WHO; 1986. [Google Scholar]
- 12.Wolfe M S, Cordero J F. Br Med J. 1985;290:1466–1467. doi: 10.1136/bmj.290.6480.1466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Fitch C D. Science. 1970;169:289–290. doi: 10.1126/science.169.3942.289. [DOI] [PubMed] [Google Scholar]
- 14.Martin S K, Oduola A M, Milhous W K. Science. 1987;235:899–901. doi: 10.1126/science.3544220. [DOI] [PubMed] [Google Scholar]
- 15.Bitonti A J, Sjoerdsman A, McCann P P, Kyle D E, Oduola A M, Rossan R N, Milhous W K, Davidson D. Science. 1988;242:1301–1303. doi: 10.1126/science.3057629. [DOI] [PubMed] [Google Scholar]
- 16.Peters W, Ekong R, Robinson B L, Warhurst D C. Lancet. 1989;2:334–335. doi: 10.1016/s0140-6736(89)90522-9. [DOI] [PubMed] [Google Scholar]
- 17.Kyle D E, Oduola A M J, Martin S K, Milhous W K. Trans R Soc Trop Med Hyg. 1990;84:474–478. doi: 10.1016/0035-9203(90)90004-x. [DOI] [PubMed] [Google Scholar]
- 18.Roepe P D. Curr Pharm Des. 2000;6:241–260. doi: 10.2174/1381612003401163. [DOI] [PubMed] [Google Scholar]
- 19.Foote S J, Thompson J K, Cowman A F, Kemp D J. Cell. 1989;57:921–930. doi: 10.1016/0092-8674(89)90330-9. [DOI] [PubMed] [Google Scholar]
- 20.Wilson C M, Serrano A E, Wasley A, Bogenschutz M P, Shankar A H, Wirth D F. Science. 1989;244:1184–1186. doi: 10.1126/science.2658061. [DOI] [PubMed] [Google Scholar]
- 21.Sanchez C P, Wunsch S, Lanzer M. J Biol Chem. 1997;272:2652–2658. doi: 10.1074/jbc.272.5.2652. [DOI] [PubMed] [Google Scholar]
- 22.Wunsch S, Sanchez C P, Gekle M, Grosse-Wortmann L, Wiesner J, Lanzer M. J Cell Biol. 1998;140:335–345. doi: 10.1083/jcb.140.2.335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bray P G, Mungthin M, Ridley R G, Ward S A. Mol Pharmacol. 1998;54:170–179. doi: 10.1124/mol.54.1.170. [DOI] [PubMed] [Google Scholar]
- 24.Ginsburg H, Famin O, Zhang J, Krugliak M. Biochem Pharmacol. 1998;56:1305–1313. doi: 10.1016/s0006-2952(98)00184-1. [DOI] [PubMed] [Google Scholar]
- 25.Sullivan D J, Jr, Gluzman I Y, Russell D G, Goldberg D E. Proc Natl Acad Sci USA. 1996;93:11865–11870. doi: 10.1073/pnas.93.21.11865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Goldberg D E, Slater A F, Cerami A, Henderson G B. Proc Natl Acad Sci USA. 1990;87:2931–2935. doi: 10.1073/pnas.87.8.2931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Bray P G, Janneh O, Raynes K J, Mungthin M, Ginsburg H, Ward S A. J Cell Biol. 1999;145:363–376. doi: 10.1083/jcb.145.2.363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Dzekunov S M, Ursos L M B, Roepe P D. Mol Biochem Parasitol. 2000;110:107–124. doi: 10.1016/s0166-6851(00)00261-9. [DOI] [PubMed] [Google Scholar]
- 29.Ursos L M B, Dzekunov S M, Roepe P D. Mol Biochem Parasitol. 2000;110:125–134. doi: 10.1016/s0166-6851(00)00262-0. [DOI] [PubMed] [Google Scholar]
- 30.Wellems T E, Patton L J, Gluzman I Y, Do Rosario V E, Gwadz R W, Walker-Jonah A, Krogstad D J. Nature (London) 1990;345:253–255. doi: 10.1038/345253a0. [DOI] [PubMed] [Google Scholar]
- 31.Su X-Z, Kirkman L S, Fujioka H, Wellems T W. Cell. 1997;91:593–603. doi: 10.1016/s0092-8674(00)80447-x. [DOI] [PubMed] [Google Scholar]
- 32.Fidock D A, Nomura T, Talley A K, Cooper R A, Dzekunov S M, Ferdig M T, Ursos L M, bir Singh Sidhu A, Naude B, Deitsch K W, et al. Mol Cell. 2000;6:861–871. doi: 10.1016/s1097-2765(05)00077-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Djimde A, Doumbo O K, Cortese J F, Kayentao K, Doumbo S, Diourte Y, Dicko A, Su X-Z, Nomura T, Fidock D A, et al. N Engl J Med. 2001;344:257–263. doi: 10.1056/NEJM200101253440403. [DOI] [PubMed] [Google Scholar]
- 34.Pillai D R, Labbe A C, Vanisaveth V, Hongvangthong B, Pomphida S, Inkathone S, Zhong K, Kain K C. J Infect Dis. 2001;183:789–795. doi: 10.1086/318836. [DOI] [PubMed] [Google Scholar]
- 35.Mayor A G, Gómez-Olivé X, Aponte J J, Casimiro S, Mabunda S, Dgedge M, Barreto A, Alonso P L. J Infect Dis. 2001;183:1413–1416. doi: 10.1086/319856. [DOI] [PubMed] [Google Scholar]
- 36.Dorsey G, Kamya M R, Singh A, Rosenthal P J. J Infect Dis. 2001;183:1417–1420. doi: 10.1086/319865. [DOI] [PubMed] [Google Scholar]
- 37.Han C M, Grimmond T R. Papua New Guinea Med J. 1976;19:236–242. [PubMed] [Google Scholar]
- 38.Cattani J A, Tulloch J L, Vrbova H, Jolley D, Gibson F D, Moir J S, Heywood P F, Alpers M P, Stevenson A, Clancy R. Am J Trop Med Hyg. 1986;35:3–15. doi: 10.4269/ajtmh.1986.35.3. [DOI] [PubMed] [Google Scholar]
- 39.Brabin B J, Ginny M, Alpers M P, Brabin L, Eggelte T, Van der Kaay H J. Ann Trop Med Parasitol. 1990;84:1–9. doi: 10.1080/00034983.1990.11812428. [DOI] [PubMed] [Google Scholar]
- 40.Foote S J, Kyle D E, Martin R K, Oduola A M J, Forsythe K, Kemp D J, Cowman A F. Nature (London) 1990;345:255–258. doi: 10.1038/345255a0. [DOI] [PubMed] [Google Scholar]
- 41.Reed M B, Saliba K J, Caruana S R, Kirk K, Cowman A F. Nature (London) 2000;403:906–909. doi: 10.1038/35002615. [DOI] [PubMed] [Google Scholar]
- 42.Dorsey G, Fidock D A, Wellems T E, Rosenthal P J. In: Antimalarial Chemotherapy. Rosenthal P, editor. Clifton, NJ: Humana; 2001. pp. 153–172. [Google Scholar]
- 43.Bockarie M J, Alexander N, Bockarie F, Ibam E, Barnish G, Alpers M. Trans R Soc Trop Med Hyg. 1996;90:23–25. doi: 10.1016/s0035-9203(96)90465-4. [DOI] [PubMed] [Google Scholar]
- 44.Bockarie M, Kazura J, Alexander N, Dagoro H, Bockarie F, Perry R, Alpers M. Am J Trop Med Hyg. 1996;54:577–581. doi: 10.4269/ajtmh.1996.54.577. [DOI] [PubMed] [Google Scholar]
- 45.Hii J L, Smith T, Mai A, Ibam E, Alpers M P. Bull Entomol Res. 2000;90:211–219. doi: 10.1017/s000748530000033x. [DOI] [PubMed] [Google Scholar]
- 46.Hii J L, Smith T, Vounatsou P, Alexander N, Mai A, Ibam E, Alpers M P. Trans R Soc Trop Med Hyg. 2001;95:7–13. doi: 10.1016/s0035-9203(01)90315-3. [DOI] [PubMed] [Google Scholar]
- 47.Ministry of Health. National Health Plan 2001–2010: Provincial and District Health Profiles. 111, Part 2. Government of Papua New Guinea: Ministry of Health; 2000. [Google Scholar]
- 48.Trager W, Jensen J B. Science. 1976;193:673–675. doi: 10.1126/science.781840. [DOI] [PubMed] [Google Scholar]
- 49.Desjardins R E, Canfield C J, Haynes J D, Chulay J D. Antimicrob Agents Chemother. 1979;16:710–718. doi: 10.1128/aac.16.6.710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Fidock D A, Nomura T, Wellems T E. Mol Pharmacol. 1998;54:1140–1147. doi: 10.1124/mol.54.6.1140. [DOI] [PubMed] [Google Scholar]
- 51.Zimmerman P A, Woolley I, Masinde G L, Miller S M, McNamara D T, Hazlett F, Mgone C S, Alpers M P, Genton B, Boatin B A, Kazura J W. Proc Natl Acad Sci USA. 1999;96:13973–13977. doi: 10.1073/pnas.96.24.13973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Triglia T, Foote S J, Kemp D J, Cowman A F. Mol Cell Biol. 1991;11:5244–5250. doi: 10.1128/mcb.11.10.5244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Su X-Z, Wellems T W. Genomics. 1996;33:430–444. doi: 10.1006/geno.1996.0218. [DOI] [PubMed] [Google Scholar]
- 54.Duraisingh M T, von Seidlein L, Jepson A, Jones P, Sambou I, Pinder M, Warhurst D C. Parasitology. 2000;121:1–8. doi: 10.1017/s0031182099006022. [DOI] [PubMed] [Google Scholar]
- 55.Moulds J M, Zimmerman P A, Doumbo O K, Kassambara L, Sagara I, Diallo D A, Atkinson J P, Krych-Goldberg M, Hauhart R E, Hourcade D E, et al. Blood. 2001;97:2879–2885. doi: 10.1182/blood.v97.9.2879. [DOI] [PubMed] [Google Scholar]
- 56.Anderson T J, Haubold B, Williams J T, Estrada-Franco J G, Richardson L, Mollinedo R, Bockarie M, Mokili J, Mharakurwa S, French N, et al. Mol Biol Evol. 2000;17:1467–1482. doi: 10.1093/oxfordjournals.molbev.a026247. [DOI] [PubMed] [Google Scholar]
- 57.Walliker D, Quakyi I A, Wellems T E, McCutchan T F, Szarfman A, London W T, Corcoran L M, Burkot T R, Carter R. Science. 1987;236:1661–1666. doi: 10.1126/science.3299700. [DOI] [PubMed] [Google Scholar]
- 58.Payne D. Parasitol Today. 1987;3:241–246. doi: 10.1016/0169-4758(87)90147-5. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}