

Abstract
Neural mechanisms for representing complex communication sounds must solve the problem of encoding multiple and potentially overlapping signals. Birdsong provides an excellent model for such processing, in that many songbird species produce multiple song types. Although auditory song representations in single song type species have been studied, how song is represented in the brains of species that sing multiple song types remains unknown. Here we examine song type representations in swamp sparrows (Melospiza georgiana), a multiple song type species, by making in vivo intracellular recordings from the telencephalic nucleus HVc, the major auditory-vocal interface in the songbird brain. These recordings show that single HVc relay neurons often generate action potentials to playback of only a single song type, even though synaptic inputs on these cells can be activated by playback of other song types in the bird's repertoire and songs of other birds. These subthreshold response patterns suggest that the song evoked action potential discharge of a single relay neuron is more selective than its presynaptic network. One component of this presynaptic network is likely to be in HVc, because multiple recordings from single birds show that different relay neurons can respond best to different song types, whereas single interneurons can generate action potentials to all song types in the bird's repertoire. These results show that single HVc neurons can generate song type-specific action potential responses, a feature that may facilitate the selective auditory encoding of multiple learned vocalizations in a single brain area.
The development and maintenance of learned vocalizations in songbirds and humans requires neural circuits capable of encoding multiple and potentially overlapping signals. In songbirds, the telencephalic nucleus HVc is essential for producing and perceiving learned song (1–4) (Fig. 1a). Beyond their essential role in singing, many HVc neurons show highly selective auditory responses, generating more action potentials to playback of the bird's own song (BOS) than to reverse BOS or other conspecific songs (5, 6). Such highly selective auditory neurons are well suited to their hypothesized roles in facilitating song discrimination and providing auditory feedback important to vocal learning. In the case of birds with multiple song types, these two functions additionally require circuits capable of discriminating among different songs in the bird's own repertoire, rather than simply discriminating the BOS from other conspecific songs. To date, song-selective neurons have been studied exclusively in birds with only a single song type, leaving unclear whether individual HVc neurons can discriminate among different song types produced by an individual bird. Here we address this question by analyzing auditory selectivity of HVc neurons in adult swamp sparrows, a species with multiple song types.
Figure 1.

HVc relay neurons in the swamp sparrow can respond exclusively to playback of a single song type in an individual's repertoire. (a) Schematic view of major brain areas important to birdsong production, learning, and perception. The nucleus HVc (used here as a proper name) receives auditory inputs and contains two distinct relay cells that innervate either the RA, a vocal motor area for song, or area X, part of a pathway essential to audition-guided vocal plasticity. (b) Intracellular recordings made in swamp sparrow brain slices show that different HVc neuron types have distinct intrinsic electrical properties. Membrane potential responses are to a +0.6-nA current pulse (Lower) passed through the recording electrode. To the right is a 5-ms portion of the record to show the action potential shape of each cell type. Note the high firing frequencies and narrow spike widths of the interneuron relative to the two relay cell types. The X-projecting cell and interneuron were identified morphologically via intracellular neurobiotin staining; the third cell type is assumed to be RA-projecting (marked by *) based on its highly refractory firing behavior, a trait displayed by morphologically identified RA-projecting cells in the zebra finch. (c) Action potential responses in this HVc relay neuron were elicited by playback of only one of the bird's three song types. (Upper) A single current clamp record obtained in an HVc relay cell evoked by playback of the preferred song (song type B; shown below as an oscillogram). The response consisted of bursts of 2–4 action potentials throughout the whole song; the membrane potential dwells below resting levels (dashed line) during the stimulus, suggestive of subthreshold inhibition. (Lower) The cell's cumulative action potential responses, shown as peristimulus time histograms (PSTHs; Spikes/bin; bin width 25 ms), to 10 presentations of each of the bird's three song types (oscillograms below each PSTH). Significant action potential responses were evoked only by song type B (two tailed t test: A: P = 0.88; B: P < 0.05; C: P = 0.31). Sonograms (i.e., frequency versus time; darkness corresponds to amplitude) of part of each song (gray box) shows that each type consists of a different repeated multinote syllable (a note is a continuous trace on the sonogram). Vertical scale bars in b and c are 40 mV; horizontal scale bars are 1 s in b and c Lower and 0.5 s in c Upper.
The males of many songbird species sing more than one song type, with individuals in some species capable of producing hundreds of distinct song patterns (7). Swamp sparrow song types comprise a single syllable repeated in a 2- to 3-s trill, with different song types characterized by which combinations of the 2–5 species-typical notes form the syllable (8, 9) (Fig. 1c). Because individual male swamp sparrows only sing 2–5 song types and the acoustic structure of these songs is readily quantified (8, 10), this species is especially tractable for studying the neural basis of song type perception. Further, much is known about vocal development and song perception in swamp sparrows (11–13), providing a strong context for relating HVc neuronal responses to the bird's behavior.
Multiple song type species such as the swamp sparrow offer a unique opportunity to clarify some of the neural mechanisms that generate auditory selectivity for learned vocalizations. Prior in vivo electrophysiological recordings made in the HVc of a single song type species, the zebra finch, suggest that heightened selectivity for the BOS emerges via HVc's network activity. Extracellular recordings show a heightened selectivity for BOS in HVc relative to its auditory afferents (14), and intracellular recordings in some HVc neurons detect subthreshold responses to stimuli (e.g., reverse BOS) that fail to evoke action potentials, suggesting synaptic input from less selective afferents (15). Given that HVc neurons of single song type species fire preferentially to the BOS, individual HVc neurons in multiple song type birds may generate action potentials to all songs in the bird's repertoire or, alternatively, may generate action potentials preferentially to only one or a subset of the bird's different song types. In the latter case, subthreshold responses to more of the bird's song types than generate suprathreshold activity could suggest that a single HVc neuron is more selective than its presynaptic partners. Intracellular recordings, which simultaneously detect a cell's subthreshold and suprathreshold responses, provide a means to address these issues in the HVc of a multiple song type species.
HVc has two relay cell types that provide auditory information to brain pathways specialized for the learning, perception and production of song (15, 16) (Fig. 1a). One relay cell type innervates the robust nucleus of the archistriatum (RA), forming a pathway important to producing learned vocalizations (1, 17), whereas the other relay type innervates area X, a basal ganglia homologue within a pathway necessary for audition-dependent vocal plasticity (18, 19); both pathways are implicated in song perception (20, 21). The different relay cell types and interneurons in the HVc of the zebra finch are distinguished by their electrophysiological properties (15, 22, 23), a relationship that we show here also applies in the swamp sparrow HVc. Then, using in vivo intracellular recordings in anesthetized swamp sparrows and song playback, we demonstrate that relay cells show suprathreshold responses to one or at most two song types in the bird's repertoire despite subthreshold responsiveness to all song types.
Materials and Methods
Relevant procedures are given briefly since they already have been published (15, 24); procedural protocol was approved by the Duke University Institutional Animal Care and Use Committee.
Brain Slices and Intracellular Recordings.
Three adult male swamp sparrows (Melospiza georgiana; >1 year) were used for brain slices. After inhalation anesthesia and decapitation, 400-μm-thick sagittal brain slices were cut on a Vibratome, then stored in an interface chamber [room temperature; on artificial cerebrospinal fluid (ACSF) gassed with 95% O2, 5% CO2]. ACSF consisted of 119 mM NaCl, 2.5 mM KCl, 1.3 mM MgCl2, 2.5 mM CaCl2, 1 mM NaH2PO4, 26.2 mM NaHCO3, and 11 mM glucose. Equiosmolar sucrose was substituted for NaCl during the dissection. Intracellular recordings were made on an interface chamber (30°C; Medical Systems, Greenvale, NY) by using sharp borosilicate pipettes (Sutter Instruments, Novato, CA; 80–200 MΩ when filled with 2 M K acetate and 5% neurobiotin). Membrane potentials were amplified by using an Axoclamp 2B amplifier (Axon Instruments, Foster City, CA) in bridge mode, low-pass filtered at 3 kHz, and digitized at 10 kHz. Slices were immersion-fixed in 4% paraformaldehyde in 0.025 M PBS and resectioned (75 μm) on a freezing microtome; neurobiotin was visualized by standard techniques (Vectastain ABC).
In Vivo Subjects and Song Stimuli.
In vivo experiments used seven adult (>1 year) male swamp sparrows from the field. Their songs were recorded in a sound attenuation chamber (Industrial Acoustics, Bronx, NY) by using a Shure SM-57 microphone and a Marantz PMD 221 tape recorder; sonograms were made with signal software (Engineering Design, Belmont, MA). Analog recordings were digitized at 20 kHz and stored on a hard drive (National Instruments, Austin, TX; software written by M. Rosen, F. Livingston, and R. Balu, Duke University). In all birds, stimuli included forward playback of all of the BOS types; in 6/7 birds, stimuli also included reverse song type playback. Non-BOS stimuli, including other conspecific (swamp sparrow) and heterospecific (song sparrow) songs, were presented to a total of 11 neurons in 6/7 birds.
In Vivo Electrophysiology.
Birds were anesthetized with 20% urethane (120 μl total; i.m.) combined with halothane inhalation. A steel post glued to the skull fixed the head in a stereotaxic device in a sound-attenuating chamber on an air table (TMC, Peabody, MA); the bird was warmed via an electric blanket. A small craniotomy was made over HVc, and the dura was retracted with a fine insect pin.
A hydraulic microdrive (Soma Scientific, Irvine, CA) lowered a sharp glass electrode (100–250 MΩ; 3 M K acetate and 5% neurobiotin) into HVc; ringing the electrode achieved cell entry. Membrane potentials were amplified with an AxoClamp 2B intracellular amplifier (Axon Instruments), low-pass filtered at 3 kHz, digitized at 10 kHz, and stored on a hard drive. HVc neurons were identified online by their action potential responses to injected positive currents (≈+0.5 nA, 1-s duration) (15); recordings usually were too brief (<15 min) to afford sufficient intracellular staining for morphological identification. Three to 30 iterations of each stimulus, delivered every 6–10 s, were played at ≈70 dB (rms, A-weighting) through a speaker 20 cm in front of the bird.
Data Analysis.
A cell's suprathreshold responses to song were assessed with respect to two criteria: whether they varied significantly from the baseline firing rate and whether they were selective for forward over reverse playback of the effective song type or, in three cells, for other BOS types. Peristimulus time histograms were made with a 25-ms bin width. The suprathreshold response (Rsupra) was calculated by Rsupra = SFR − BFR, where SFR and BFR are the firing rates during each stimulus presentation and during a 1.5-s baseline period before each stimulus presentation, respectively. In cells with transient responses, narrower analyses windows (0.2–0.6 s) were centered on the peak response and on the region of highest baseline activity. Response strengths are shown normalized to the maximum response elicited from the given cell. To assess subthreshold responses in spiking and nonspiking cells, raw traces were median-filtered (each point replaced by the median of the surrounding 50 points, or 5 ms at a 10-kHz sampling rate). The subthreshold depolarizing responsiveness (RVm) was measured by RVm = Sarea − Barea, where Sarea and Barea are the integrals of the positive-going deviations in membrane potential during (i.e., Sarea) or prior (i.e., Barea) to the stimulus relative to the mode membrane potential measured during the baseline period. Similar calculations were made for subthreshold hyperpolarizing area. Subthreshold response strengths are shown normalized to the maximum response elicited from that cell. Significance was determined with paired t tests comparing stimulus-evoked suprathreshold, subthreshold depolarizing, or subthreshold hyperpolarizing responses to corresponding baseline measures. Note that some cells that spike to a stimulus may have no significant subthreshold response by using median-filter averaging. This situation can arise when a cell rests very close to spike threshold, or when threshold is actually below the resting membrane potential, as can happen when hyperpolarization deinactivates certain voltage-gated currents.
The selectivity of a given neuron for forward over reverse BOS playback was measured by using the psychophysical metric d′ (25). In three relay cells tested, where reverse BOS types were not used, the d′ statistic was calculated against an ineffective BOS type. The d′ value comparing the response to BOS relative to reverse BOS is given by:
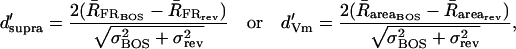 |
where d′supra is suprathreshold selectivity and d′Vm represents subthreshold selectivity. R̄ is the mean value of R (see above), and σ2 is its variance. This measure of selectivity is similar to a ratio measure, but accounts for both the mean and the variance of a cell's responses and can report negative values. A d′ value >0.7 or <−0.7 was used as the criterion for identifying cells as selective.
Results
Establishing Swamp Sparrow HVc Cell Identity.
Prior intracellular studies showed that different morphological cell types in the zebra finch HVc have distinct intrinsic (i.e., dc-evoked) electrical properties (15, 22, 23). To confirm that a similar situation exists in the sparrow HVc, we made swamp sparrow brain slices and recorded intracellularly in HVc (n = 9 slices from three birds). Positive current pulses (0.2–1.0 nA) passed through the electrode were used to evoke action potentials. Three electrically distinct cell types were detected (Fig. 1b), with dc-evoked action potential trains like either X-projecting neurons, RA-projecting neurons, or interneurons described in the finch. Indeed, intracellular staining in the sparrow confirmed that X-projecting neurons (n = 9 cells), with moderate evoked firing rates, and interneurons (n = 2 cells), with high evoked firing rates, were electrically distinct from each other and highly similar to their finch homologues (see figure 1 in refs. 15 and 22). For identified sparrow interneurons, the mean spike frequency to injected current value of 140 Hz/nA approximated that reported for finch interneurons in vivo (≈170 Hz/nA; ref. 15), and was higher than that of sparrow X-projecting neurons (≈28 Hz/nA). We also encountered a third cell type (n = 2 cells) in the sparrow HVc that was electrically distinct from identified interneurons and X-projecting cells (Fig. 1b). Although held too briefly to stain, this third type fired only one or few action potentials when injected with moderate to large positive currents (+0.5 to 1 nA); such refractoriness typifies identified RA-projecting neurons in the finch. Because morphologically identified interneurons and X-projecting cells in the sparrow HVc are electrically distinct from each other yet highly similar to their finch homologues, and because a third cell type in the sparrow HVc has intrinsic properties like finch RA-projecting neurons, we conclude that the intrinsic electrical behavior of HVc neurons is conserved and can be used to sort relay cells and interneurons in both species.
Song Type Specificity.
In most HVc relay cells we examined, only one of the several song types in the bird's repertoire evoked elevated action potential (suprathreshold) discharges [Figs. 1c and 2A; 7/10 relay cells had significant (P < 0.05) excitatory suprathreshold responses to only one song type whereas the other three cells had significant responses to 2/3 song types (n = 7 birds); an additional cell lacked excitatory responses, but showed firing rate suppression to a single song type]. The suprathreshold excitatory responses to the preferred song type were distinguished by phasic and repeated bursts of 2–5 action potentials, which were qualitatively distinct from either onset responses or low-level sustained discharge sometimes elicited by other BOS types or non-BOS song stimuli (Fig. 1c, raw trace Upper, and Fig. 3b). In addition, significant firing rate suppression was elicited in three cells by a single song type in the bird's repertoire; two cells showed suprathreshold responses to another song type in the repertoire, whereas the third cell (cell 847, swred) lacked excitatory suprathreshold responses to all song types (Fig. 2A). All 11 relay cells that showed suprathreshold changes in firing rate to one or more song types also showed temporal sensitivity to the effective stimuli, as these firing rate changes were evoked by forward and not reverse playback of the preferred songs (Fig. 2A; 11/11 cells, d′ > 0.7 or < −0.7). These results demonstrate that swamp sparrow HVc relay neurons often generate action potentials to playback of only a subset of the bird's song types, and most often to only a single song type. Furthermore, these responses reflect sensitivity to temporal features of the preferred stimulus, as shown for the BOS by HVc neurons in birds with a single song type (5, 6, 15, 26).
Figure 2.

Swamp sparrow HVc relay cells can show suprathreshold selectivity to only a subset of the bird's song types, but often show subthreshold responses to its other song types, and the songs of other birds. (A) The suprathreshold selectivity [Upper; d′ value measuring forward vs. reverse song (* or other song type) playback] and subthreshold responsiveness (Lower) of 11 relay cells (800, etc.; from six birds: sw210, etc.) in response to playback of the bird's different song types (A–E). Song types that failed to elicit a significant suprathreshold response were given an arbitrary score of 0.1. In eight cells, only one song type evoked a selective suprathreshold response. In six cells, song types that failed to evoke significant suprathreshold responses still evoked significant subthreshold responses (normalized to the cell's maximum subthreshold response). Positive values reflect depolarizing responses; negative values mark hyperpolarizing responses. (B) In all 12 nonspiking HVc cells encountered, all song types, as well as conspecific and heterospecific songs, evoked significant subthreshold responses (normalized to the maximum response as in A). (C) In relay cells with significant suprathreshold responses to at least one song type subthreshold and, in one cell, suprathreshold responses to conspecific and heterospecific song playback were also detected (responses normalized to maximum subthreshold or suprathreshold value).
Figure 3.

Individual examples showing contrasting subthreshold and suprathreshold response patterns of different HVc neuron types, and qualitative differences in the responses to BOS and non-BOS responses. (a) In this relay neuron (cell 191 in Fig. 2C), suprathreshold responses were evoked only by song type B, but significant (P < 0.05) subthreshold depolarizations were evoked by all song types of that bird and also by another conspecific song (con). (b) In another relay neuron (cell 850 in Fig. 2C) significant action potential responses were evoked by song type C and by conspecific and heterospecific song (het; song sparrow). The suprathreshold response to song type C was highly phasic, distinct from the low-intensity, sustained action potential discharges elicited by the other birds' songs. Hyperpolarizing responses were evoked by playback of all of the bird's song types, but not by the songs of the other birds (Vm). (c) Certain HVc neurons displayed robust but entirely subthreshold depolarizations to all song types within the bird's repertoire; these were greater for forward than reverse playback (reverse song was delivered alternately over the same period shown for the forward song; all responses were significant at P < 0.05; cell is rm060 in Fig. 2B). (d) In an HVc interneuron, action potential responses were evoked by forward and not reverse playback of all song types (d′song type A = 7.0; d′song type B = 4.9; d′song type C = 2.8; three iterations). Vertical scale bars are 10 mV; horizontal scale bars in a, b, and d equal 1 s and in c are 2 s.
The song type-specific action potential responses seen in the swamp sparrow HVc raise the possibility that different neurons are excited by different song types within a single bird. Indeed, such complementary organization was observed in four animals, where different HVc neurons showed suprathreshold responses to different song types within the bird's repertoire (Fig. 2A). These results suggest that single HVc relay neurons can represent single song types in their suprathreshold activity, but as a population may constitute a network that responds to many or all song types in the bird's repertoire.
Subthreshold Responses.
The song type-specific action potential responses we observed may merely reflect excitatory synaptic inputs with similar tuning features, or instead might occur against a background of more broadly responsive synaptic inputs. Using intracellular methods, which reveal both suprathreshold and subthreshold (i.e., synaptic) responses (15, 27, 28), we found that individual HVc neurons with narrow suprathreshold selectivity for only one or two song types can receive broad synaptic drive activated by these and other song types in the repertoire. Although these HVc relay cells only fired action potentials to one, or at most two, of the bird's song types, other song types outside this subset often elicited subthreshold depolarizing or hyperpolarizing responses that were not accompanied by action potential generation (6/11 cells; Figs. 2A and 3 a and b). Furthermore, 12 HVc neurons that did not generate action potentials to any song type still were strongly depolarized by playback of all song types in the bird's repertoire (Figs. 2B and 3c), and the relative strengths of these depolarizing responses varied among different cells from the same bird. In all such “silent” cells tested with forward and reverse song playback, subthreshold temporal sensitivity was evident (Fig. 3c; 8/10 cells had d′ > 0.7 for all song types, 2/10 cells had d′ > 0.7 for 2/3 song types). Even when subthreshold hyperpolarizations were not distinct, action potential suppression sometimes was observed to forward playback of the nonpreferred song types (two cells; not shown). Along with the broad subthreshold inhibitory responses mentioned above, firing rate suppression suggests broadly tuned, forward song-selective inhibition. In the zebra finch HVc, a close concordance exists between interneuron firing and inhibition in X-projecting cells (15). Consistent with the idea that the inhibition we saw in certain swamp sparrow relay cells also might come from local interneurons, cells with the fast-spiking properties (as observed online) typical of interneurons showed elevated action potential discharge to forward but not reverse playback of all song types in the bird's repertoire (Fig. 3d; n = 2 cells in two birds).
Responses to Conspecific and Heterospecific Songs.
Playback of conspecific and heterospecific songs was used to further characterize the selective auditory responses in swamp sparrow HVc neurons. In four birds from which cells were tested in this manner, HVc neurons (five relay cells and one interneuron) showed no suprathreshold responses to playback of either conspecific (swamp sparrow) songs or to heterospecific (song sparrow) songs (Figs. 2C and 3a). Despite an absence of suprathreshold responses, non-BOS stimuli could sometimes elicit subthreshold responses, suggesting that cells presynaptic to the impaled neuron were responsive to the playback (non-BOS songs elicited subthreshold responses in 3/5 spiking relay neurons and in four other nonspiking neurons; Figs. 2 B and C and 3a). Even in one relay neuron from a fifth bird in which weak action potential discharge was elicited by non-BOS song playback, the responses were sustained and extremely low in intensity, distinct from the phasic, high-intensity action potential bursts elicited by the preferred BOS song type (cell 850; Figs. 2C and 3b). Furthermore, the non-BOS songs failed to drive the hyperpolarizing responses evoked by all of the BOS types (Fig. 3b).
Discussion
Most of the HVc relay neurons we studied generate action potentials only to a single song type in the bird's repertoire, and thus discriminate between different self-generated vocal signals, not merely the bird's own vocalizations from those of other conspecifics. The capacity for song type-specific firing could facilitate auditory processes important to song learning and adult song maintenance, specifically by activating distinct subpopulations of HVc relay neurons when a bird's vocalization matches one of several memorized song models. Such song type-specific neurons also could facilitate patterns of song use observed in the field, such as vocal matching, where an individual bird selectively sings songs that match those of neighboring conspecifics (7). Furthermore, the finding that HVc relay neurons can generate action potentials to only a single song type despite sustaining marked subthreshold responses to other song types in the bird's repertoire is consistent with the idea that HVc is a site for auditory refinement of learned communication sounds.
In vivo recordings from zebra finches show that different HVc relay cells display distinct subthreshold responses to the BOS, with RA-projecting cells receiving forward song-selective excitatory drive and X-projecting cells receiving a mix of less selective excitation and highly selective inhibition, probably arising from HVc interneurons (15). The subthreshold song-evoked responses of the relay cells studied here, coupled with differences in their suprathreshold excitability to injected currents that we observed online, makes it likely that those cells with exclusively depolarizing (and sometimes entirely subthreshold) responses to song playback were RA-projecting, whereas those showing song-evoked inhibition were area X-projecting. An intriguing aspect of the neuronal behavior we observed is that despite the differing nature of subthreshold responses in these two relay cell types, action potential generation could be limited to a single song type, and did not always require detectable postsynaptic inhibition. Furthermore, these observations indicate that, as a population, neurons presynaptic to many HVc relay cells are driven broadly by all of the bird's song types.
The subthreshold responses certain HVc relay cells showed to all or most of the bird's song types (and the similarly broad suprathreshold responses of interneurons— although this awaits further characterization of a larger sample) likely reflect local and extrinsic synaptic inputs onto the impaled cell. Local sources of input to HVc relay neurons include interneurons and the local axon collaterals of other relay cells (15, 22, 29). As shown here, single interneurons and populations of relay cells can respond to all song types in the bird's repertoire. Interneurons, which appear to provide inhibition to X-projecting cells in the zebra finch, could account for all of the song-evoked hyperpolarizations seen in swamp sparrow HVc relay cells, whereas excitatory axon collaterals of other relay cells could account for at least some of the observed depolarizing responses. Neurons extrinsic to HVc also could drive the subthreshold responses we observed in HVc relay neurons. A direct source of BOS-biased auditory input to HVc in the zebra finch is the nucleus interfacialis (NIf); other indirect sources are the caudal neostriatum and the caudal hyperstriatum ventrale, which both contain auditory neurons biased toward conspecific songs (30, 31). In the zebra finch, BOS-selective responses persist in HVc even when the local circuit is inactivated, pointing to an extrinsic origin of song-selective input.§ If the broad subthreshold responses in single swamp sparrow relay neurons arise extrinsically, then they may persist upon local inactivation. Conversely, if derived through local interactions, inactivation might abolish responses to certain song types.
The action potential responses of HVc neurons in a bird with multiple song types also yield insight into the functional organization of this nucleus in a fashion not possible in studies of single song type species. Specifically, the units of auditory processing in the swamp sparrow HVc (i.e., song type specific neurons) directly parallel the bird's units of vocal performance (i.e., unique song types). Future work will determine whether the suprathreshold responses we observed also provide a mechanism for categorical perception, in which continuously varying auditory stimuli are partitioned into discrete perceptual categories (32–34). Male swamp sparrows respond vigorously when the duration of the initial note in a song type to which they previously had been habituated is varied across a boundary distinguishing two note categories, but not when the initial note duration is varied by a similar amount within the original note category (13). The fact that swamp sparrows are known to behaviorally discriminate acoustically similar song types categorically, coupled with our finding of song type-specific neuronal responses, should permit further exploration of how perceptual categories are generated by neurophysiological mechanisms in the vertebrate brain.
Acknowledgments
We thank Drs. Lawrence Katz and Erich Jarvis for comments on earlier drafts of the manuscript and Susan Peters for song analysis. This work was supported by grants from the National Institutes of Health and the Klingenstein Fund to R.M. and from the National Science Foundation and the John Simon Guggenheim Foundation to S.N.
Abbreviations
- BOS
bird's own song
- RA
robust nucleus of the archistriatum
Footnotes
Rosen, M. J. & Mooney, R. (2000) Soc. Neurosci. Abstr. 26, 2030.
References
- 1.Nottebohm F, Stokes T M, Leonard C M. J Comp Neurol. 1976;165:457–486. doi: 10.1002/cne.901650405. [DOI] [PubMed] [Google Scholar]
- 2.Vu E T, Mazurek M E, Kuo Y C. J Neurosci. 1994;14:6924–6934. doi: 10.1523/JNEUROSCI.14-11-06924.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Brenowitz E A. Science. 1991;251:303–305. doi: 10.1126/science.1987645. [DOI] [PubMed] [Google Scholar]
- 4.Gentner T Q, Hulse S H, Bentley G E, Ball G F. J Neurobiol. 2000;42:117–133. doi: 10.1002/(sici)1097-4695(200001)42:1<117::aid-neu11>3.0.co;2-m. [DOI] [PubMed] [Google Scholar]
- 5.Margoliash D. J Neurosci. 1986;6:1643–1661. doi: 10.1523/JNEUROSCI.06-06-01643.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Volman S F. J Comp Physiol A. 1996;178:849–862. doi: 10.1007/BF00225832. [DOI] [PubMed] [Google Scholar]
- 7.Catchpole C, Slater P. Bird Song: Biological Themes and Variations. Cambridge, U.K.: Cambridge Univ. Press; 1995. [Google Scholar]
- 8.Marler P, Pickert R. Anim Behav. 1984;32:673–689. [Google Scholar]
- 9.Mowbray T B. In: The Birds of North America. Poole A, Gill F, editors. Vol. 279. Philadelphia: Academy of Natural Sciences; 1997. pp. 1–23. [Google Scholar]
- 10.Clark C, Marler P, Beeman K. Ethology. 1987;76:101–115. [Google Scholar]
- 11.Marler P, Peters S. Science. 1981;213:780–782. doi: 10.1126/science.213.4509.780. [DOI] [PubMed] [Google Scholar]
- 12.Marler P, Peters S. Dev Psychobiol. 1982;15:369–378. doi: 10.1002/dev.420150409. [DOI] [PubMed] [Google Scholar]
- 13.Nelson D A, Marler P. Science. 1989;244:976–978. doi: 10.1126/science.2727689. [DOI] [PubMed] [Google Scholar]
- 14.Lewicki M S, Arthur B J. J Neurosci. 1996;16:6987–6998. doi: 10.1523/JNEUROSCI.16-21-06987.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Mooney R. J Neurosci. 2000;20:5420–5436. doi: 10.1523/JNEUROSCI.20-14-05420.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Vates G E, Broome B M, Mello C V, Nottebohm F. J Comp Neurol. 1996;366:613–642. doi: 10.1002/(SICI)1096-9861(19960318)366:4<613::AID-CNE5>3.0.CO;2-7. [DOI] [PubMed] [Google Scholar]
- 17.Yu A C, Margoliash D. Science. 1996;273:1871–1875. doi: 10.1126/science.273.5283.1871. [DOI] [PubMed] [Google Scholar]
- 18.Bottjer S W, Miesner E A, Arnold A P. Science. 1984;224:901–903. doi: 10.1126/science.6719123. [DOI] [PubMed] [Google Scholar]
- 19.Brainard M, Doupe A. Nature (London) 2000;404:762–766. doi: 10.1038/35008083. [DOI] [PubMed] [Google Scholar]
- 20.Scharff C, Nottebohm F, Cynx J. J Neurobiol. 1998;36:81–90. [PubMed] [Google Scholar]
- 21.Williams H, Nottebohm F. Science. 1985;229:279–282. doi: 10.1126/science.4012321. [DOI] [PubMed] [Google Scholar]
- 22.Dutar P, Vu H M, Perkel D J. J Neurophysiol. 1998;80:1828–1838. doi: 10.1152/jn.1998.80.4.1828. [DOI] [PubMed] [Google Scholar]
- 23.Kubota M, Taniguchi I. J Neurophysiol. 1998;80:914–923. doi: 10.1152/jn.1998.80.2.914. [DOI] [PubMed] [Google Scholar]
- 24.Livingston F S, Mooney R. J Neurosci. 1997;17:8997–9009. doi: 10.1523/JNEUROSCI.17-23-08997.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Green D, Swets J. Signal Detection Theory and Psychophysics. New York: Wiley; 1966. [Google Scholar]
- 26.Theunissen F E, Doupe A J. J Neurosci. 1998;18:3786–3802. doi: 10.1523/JNEUROSCI.18-10-03786.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Jagadeesh B, Wheat H S, Kontsevich L L, Tyler C W, Ferster D. J Neurophysiol. 1997;78:2772–2789. doi: 10.1152/jn.1997.78.5.2772. [DOI] [PubMed] [Google Scholar]
- 28.Rosen M J, Mooney R. J Neurosci. 2000;20:5437–5448. doi: 10.1523/JNEUROSCI.20-14-05437.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Benton S, Cardin J A, Devoogd T J. Brain Res. 1998;799:138–147. doi: 10.1016/s0006-8993(98)00485-5. [DOI] [PubMed] [Google Scholar]
- 30.Chew S J, Mello C, Nottebohm F, Jarvis E-P, Vicario D S. Proc Natl Acad Sci USA. 1995;92:3406–3410. doi: 10.1073/pnas.92.8.3406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Stripling R, Volman S F, Clayton D F. J Neurosci. 1997;17:3883–3893. doi: 10.1523/JNEUROSCI.17-10-03883.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Moody D, Stebbins W, May B. In: Comparative Perception: Complex Signals. Stebbins W, Berkley M, editors. Vol. 2. New York: Wiley; 1990. pp. 279–310. [Google Scholar]
- 33.Liberman A M, Cooper F S, Shankweiller D S, Studdert-Kennedy M. Psychol Rev. 1967;74:431–461. doi: 10.1037/h0020279. [DOI] [PubMed] [Google Scholar]
- 34.Kuhl P, Miller J. Science. 1975;190:69–72. doi: 10.1126/science.1166301. [DOI] [PubMed] [Google Scholar]