

Abstract
The aryl hydrocarbon receptor (AhR) is a DNA binding protein that acts as a nuclear receptor mediating xenobiotic metabolism and environmental responses. Owing to the evolutionary conservation of this gene and its widespread expression in the immune and circulatory systems, AhR has for many years been almost exclusively studied by the pharmacological/toxicological field for its role in contaminant toxicity. More recently, the functions of AhR in environmental adaption have been examined in the context of the occurrence, development, and therapy of cardiovascular diseases. Increasing evidence suggests that AhR is involved in maintaining homeostasis or in triggering pathogenesis by modulating the biological responses of critical cell types in the cardiovascular system. Here, we describe the structure, distribution, and ligands of AhR and the AhR signaling pathway and review the impact of AhR on cardiovascular physiology. We also discuss the potential contribution of AhR as a new potential factor in the targeted treatment of cardiovascular diseases.
1. Introduction
Aryl hydrocarbon receptor (AhR) is a ligand-activated transcriptional factor belonging to the superfamily of basic helix-loop-helix/Per-ARNT-Sim (bHLH/PAS) [1], and is the only member of this family known to bind naturally occurring xenobiotics [2]. Traditionally AhR acts as a crucial regulator mediating xenobiotic metabolism and environmental responses, and it was discovered to bind closely with 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD, dioxin) and then was hyperactivated to release a myriad of toxicologic outcomes, which contribute to the potency of TCDD as an inducer or promoter of some carcinogenesis in 1982 [3, 4]. Thus, for many years, AhR has been almost exclusively studied by the toxicological field for its role in various environmental and food contaminants such as polycyclic aromatic hydrocarbons, polychlorinated biphenyls and dioxins. Numerous studies have found that cytosolic AhR can be activated by many natural and synthetic ligands, and translocated into the nucleus where it complexes with the AhR nuclear translocator (ARNT) [5]. The complex recognizes the specific dioxin-responsive elements (DREs) and modulates subsequent transcription of its downstream target genes including phase I and phase II metabolic enzymes, which can affect the metabolism of environmental toxicants and chemical substances [6].
The increasing deterioration of the natural environment is having serious consequences on human health. The circulation system is the major organ exposed xenobiotics and endobiotics during metabolic homeostasis [7], and long-term exposure to environmental pollutants can drastically alter this system, resulting in cardiovascular diseases such as hypertension, atherosclerosis, and ischemic heart disease [8–12]. Because many environmental pollutants contain exogenous aryl hydrocarbon receptor (AhR) ligands, increasing attention is being given to the relationship between AhR and cardiovascular diseases. Recent evidence from gene knock-out studies and clinical trials suggests that not only does AhR have a major impact on general physiological functions, including immune responses, reproduction, oxidative stress, tumor promotion, the cell cycle, and proliferation [13, 14], but also influences cardiovascular physiological functions [15–18].
In this review, we discuss the progress of AhR biology and toxicology, its pathophysiology roles in the heart and vascular systems, and the prospects as a therapeutic target for cardiovascular diseases, with the aim of providing a potential direction for the prevention and treatment of the diseases.
2. AhR
2.1. The Structure of AhR
Anthropogenic AhR comprises 848 amino acid residues and has three functional domains, including the bHLH domain, Per-ARNT-Sim (PAS) domains (A and B), and the transactivation domain (TAD), that span from the amino (N-) terminal to the carboxy (C-) terminal [1, 19] (Figure 1). The amino acid sequence of the bHLH domain and the PAS domains are both highly conserved among species [20]. The bHLH domain is located at the (N-) terminal and can divide into an HLH domain and a basic domain, which determines dimerization of the protein molecule and the combination of AhR with DNA [21]. The main role of the PAS A and B domains is to participate in binding to ligands, release of heat shock protein 90, and increase the stability of the heterodimer AhR-ARNT complex to further affect conformation of DNA [1, 22]. The TAD domain functions as a mediator in transcriptional activation of downstream genes [23].
Figure 1.

Structure of AhR.
2.2. Distribution of AhR in Fetal and Adult Tissues
AhR is expressed ubiquitously in the fetus and in adults, with the distribution changing significantly with age [24] (Figure 2). In the fetus, there are specific distribution of AhR in the lungs, liver, kidneys, pancreas, testicles, esophagus, thymus glands, retinas, and epithelial cells, and relatively low levels in the heart, brain, choroids, thoracic aorta, and sclera; In adults, AhR is expressed at relatively high levels in the lungs, placenta, spleen, pancreas, and liver, and relatively low levels in the heart, brain, and skeletal muscles [25, 26]. AhR exerts diverse physiological effects depending on where it is located in different tissues.
Figure 2.
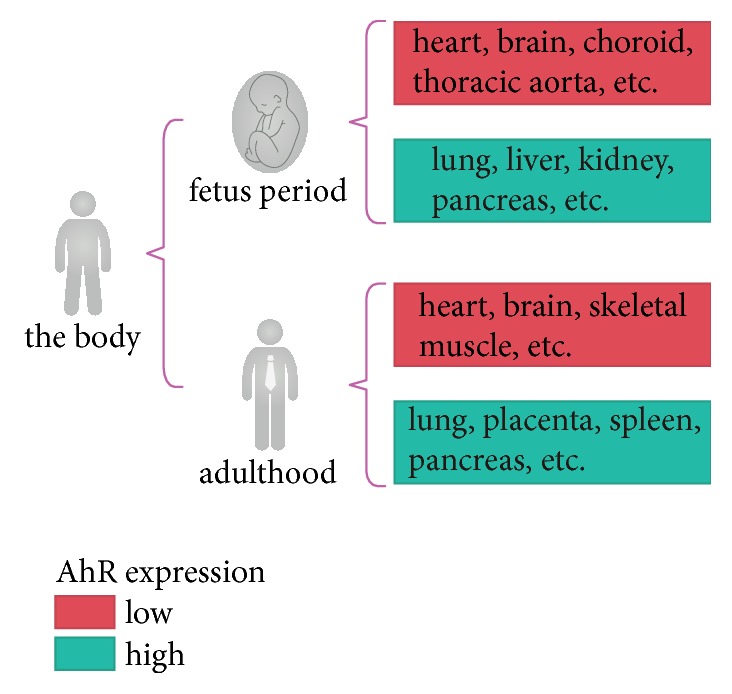
Expression levels of AhR in adult and fetal tissues.
2.3. The Ligands of AhR
AhR ligands can be divided into endogenous ligands and exogenous ligands (Figure 3). Endogenous ligands include indigoids, heme metabolites, eicosanoids, tryptophan derivatives, and equilenin [27]. Exogenous ligands include polycyclic aromatic hydrocarbons, polychlorinated biphenyls, natural compounds, and small molecule compounds [28]. The different structures and properties of AhR ligands mean that when they combine with AhR they have distinct biological effects.
Figure 3.

Exogenous and endogenous ligands of AhR.
2.4. The AhR Signaling Pathway
The AhR signaling pathway involves both classical and non-classical signal transduction mechanisms [2, 24] (Figure 4). In the classical signaling pathway, AhR exists as an AhR molecular chaperone complex comprising an AhR, two heat shock protein 90, and X-associated protein 2 and 23 in the cytosol [24, 29]. Being activated by ligands, AhR is translocated from the cytosol to the nucleus where it disassociates from the complex. The ligand-AhR complex combines with ARNT and binds to a specific DNA promoter sequence called DRE or xenobiotic responsive element (XRE). Ultimately, transcription of a large number of target genes activates and triggers various biological and/or toxicological effects [30].
Figure 4.

Classical and non-classical AhR signaling pathways.
In the non-classical signaling pathway, the AhR signaling pathway can interact with other pathways by competition for transcriptional coactivators or corepressors [30]. For instance, crosstalk between AhR and hypoxia can interact through competition with ARNT [22]. In the nucleus, sustained AhR activation results in G1 phase cell cycle arrest via hyperphosphorylation of retinoblastoma protein and repression of E2F-dependent transcription [31–35]. In macrophages, AhR, in combination with signal transducer and activator of transcription 1 and nuclear factor-κB (NF-κB), inhibit the promoter activity of interleukin-6 (IL-6) induced by lipopolysaccharide [36]. AhR also regulates the development of regulatory type 1 T cells by combining with the transcription factor c-Mcf [37]. Additionally, numerous studies have described interactions between AhR and estrogen receptors, RelB, RelA, β-catenin, and nuclear factor-like 2[38–43].
3. Role of AhR in Cardiovascular Physiology
3.1. AhR in Cardiac Function and Cardiomyogenesis
Despite low expression levels of AhR in the heart, AhR does have noticeable effects on the physiological functioning of the heart. For example, a study has reported obvious cardiac hypertrophy in AhR−/− mice at 5 months, with increasing levels of the β-myosin heavy chain and β-myosin light chain 2V. It was suggested that the underlying mechanism may be associated with the elevated level of vascular endothelial growth factor (VEGF) in AhR−/− mice [44]. In 2003, Vasquez et al. observed that cardiac hypertrophy induced in AhR deficiency showed low indices for contractility, pre-load, afterload, cardiac output, stroke volume, and minimal fibrosis, differing from pressure- or volume overload-related cardiac hypertrophy [15]. The researchers suggested that AhR deficiency mainly lead to cardiomyocyte hypertrophy, resulting in cardiomyopathy and cardiac hypertrophy [45]. Paradoxically, another study reported that cardiac hypertrophy in AhR−/− mice was caused by pressure overload and accompanied by evident fibrosis and elevated expression of plasma endothelin-1 (ET-1) and angiotensin II (Ang II). Captopril, an angiotensin-converting enzyme inhibitor, was used to alleviate the pressure overload, leading to a lowered expression of plasma ET-1 and Ang II and a delay in the increase of mean arterial pressure [16]. Subsequent research found that cardiac function in AhR−/− mice could be completely reversed with BQ-123, an ETA receptor antagonist, indicating that ET-1 could be mediated by AhR and function as the key molecule in the progression of cardiac hypertrophy [46]. A recent study revealed that Vav3, an activator of Rho/Rac GTPases, regulated by AhR, was closely associated with cardiac hypertrophy and fibrosis in AhR−/− mice [47]. Nevertheless, the specific mechanism has yet to be completely determined. Although there are contradictions among studies, it is evident that AhR signaling in cardiac function is important.
The AhR signaling pathway is vital for the development of the heart. When AhR was activated by dioxin, transforming growth factor β (TGF-β)/bone morphogenic protein (BMP) and WNT signaling pathways were disrupted, cardiomyocyte differentiation of enterochromaffin cells stopped, and cardiogenesis was impaired during early differentiation [48]. When AhR was silenced by short hairpin RNA interference in P19 cells, an embryonic carcinoma cell line, expression of the downstream signal molecules of AhR such as ARNT and CYP1A1 and the key molecule in WNT signaling, β-catenin, were suppressed, following by the increase in expression of the cardiomyogenesis-specific GATA4 and Nkx2.5 genes. These results suggest that AhR mediated the differentiation of P19 mouse embryonic carcinoma cells into cardiomyocytes through the AhR and WNT1 signaling pathways [49]. Another study found that activation, inhibition, or knockdown of AhR all could affect cardiomyocyte differentiation of mouse embryonic stem cells; the cause of which was connected with AhR-relating incongruous expression of genes, including genes encoding homeobox transcription factors and polycomb and trithorax group genes [50]. The expression of AhR in the undifferentiated embryonic stem cells impacts their choice of lineage in differentiation, restricting cardiogenesis and commit to a neuroglia cell fate. With regard to self-renewal of embryonic stem cells, a relatively low level of AhR expression was required for cells to retain their stem cell properties. Han et al. propose that after activation by endogenous ligands AhR participates in the coordination of multiple biological processes which define pluripotency and embryonic development, and AhR can regulate cardiogenesis by modulating the cardiac DNA methylome and the expression of imprinting genes [51]. Hence, cardiomyocyte differentiation is a carefully regulated process in which AhR plays a crucial role.
3.2. AhR in the Regulation of Vascular Physiological Functions
Maintenance of the function and structure of blood vessels relies in part on laminar fluid shear stress. Normally, laminar fluid shear stress-activated AhR mediate cell cycle arrest by activating CYP1A1 in human umbilical vein endothelial cells, suggesting the involvement of AhR in the regulation of the vascular microenvironment [52]. A study reported that there exists abnormal vascular structures in the liver, kidneys, and hyaloids in AhR−/− mice [53]. A study of hepatic vascular development revealed that hepatic necrosis and decreased perfusion in the fetal liver was the cause of patent ductus venosus and comparatively smaller livers in adult AhR−/− mice [54]. The results align with that of another study in which mutation of DRE binding sites in AhR affected liver vascular development, suggesting that DNA binding is necessary for AHR-mediated developmental and TCDD-induced toxic signaling [55].
When AhR was activated by 3-methylcholanthrene, an AhR agonist, cell permeability, adhesion, and tube formation of human umbilical vascular endothelial cells was inhibited; but α-naphthoflavone, an AhR antagonist, could reverse the effects of 3-methylcholanthrene [56]. Another study reported that TCDD activated the AhR/CYP1A1 and AhR/CYP1B1 pathways, resulting in suppression of angiogenesis, and angiogenic inhibition was reversed with AhR deficiency [57]. Ichihara et al. used middle cerebral artery occlusion in mice and oxygen-glucose deprivation in rat cortical neurons to define the role of AhR in stroke, and the results found that L-kynurenine is an endogenous ligand that mediates AhR activation in the brain, and demonstrated that an L-kynurenine/AhR pathway mediates acute brain ischemic damage after stroke[58]. Ischemia-induced angiogenesis was observed to significantly increase with AhR deficiency, with the effects being associated with the hypoxia-inducible factor-1α (HIF-1α)-ARNT heterodimer and its downstream gene, VEGF [59]. VEGF is necessary for vascularization. Compared with AhR+/+ transgenic adenocarcinoma of the mouse prostate (TRAMP) mice, AhR−/− TRAMP mice showed a higher incidence of prostate cancer accompanied by an increase in VEGF [60]. Similarly, AhR activation inhibited hypoxia-induced VEGF in prostate bone metastasis cells and endothelial progenitor cells, and was associated with HIF-1α [61]. It is therefore suggested that AhR may prevent interaction of ARNT with HIF-1α and suppress VEGF expression, ultimately blocking angiogenesis.
In summary, AhR participates in the regulation of vascular physiological functions, including vascular development and angiogenesis. Both deficiency and abnormal activation of AhR give rise to vascular dysfunction, and many vascular diseases.
3.3. AhR in Blood Pressure Regulation
It is reported that AhR-deficient mice showed decreased cardiac output and low systolic and diastolic aortic pressure compared with normal mice of the same age [15]. In 2010, Zhang et al. reported that the renin–angiotensin system participated in the regulation of normal blood pressure in AhR heterozygous mice but not AhR−/− mice, confirming the importance of the rennin-angiotensin system in the progression of hypotension in AhR−/− mice. ET-1 signaling was also found to be involved in the mediation of hypotension in AhR−/− mice. Importantly, the study reported that the sympathetic nervous system and nitric oxide (NO) signaling were not involved in the activation of the rennin-angiotensin system and ET-1 [62]. AhR−/− mice tend to develop hypertension at a modest altitude (1632 m), caused by hypoxia [63]. Captopril could alleviate high blood pressure in AhR−/− mice, in part because of the reduction of Ang II [16]. Increasing evidence indicates that vascular α1D-adrenoceptor overexpression is another influential factor of hypertension in AhR−/− mice, and hypertension could be reversed by treatment with captopril [64]. However, inhibition of ET-1 could not only lower mean arterial pressure and the levels of ET-1, but also reduce Ang II expression levels in AhR−/− mice with hypertension, indicating involvement of the regulation of the ET-1-Ang II axis in hypertension in AhR−/− mice induced by hypoxia [65]. Sauzeau et al. found that AhR controlled cardiovascular and respiratory functions by regulating the expression of the Vav3 proto-oncogene, and demonstrated Vav3 to be a bona fide AhR target in charge of a limited subset of the developmental and physiological functions of cardiorespiratory systems [46]. Taken together, the findings suggest that AhR is involved in the complicated networks that regulate blood pressure, and possible mechanisms should be further studied.
4. AhR as a Therapeutic Target in Cardiovascular Diseases
4.1 AhR and myocarditis. Myocarditis is a significant cause of heart disease, especially in young people[66]. It can lead to dilated cardiomyopathy, a common precursor of heart failure. Myocarditis can be induced by multiple causes, including infection and auto-immune or auto-inflammatory diseases [67, 68]. Infection remains a major factor in myocarditis and is closely associated with the immune and inflammatory responses of the host. Numerous studies have reported that AhR is a crucial factor in the immune system and is involved in the differentiation of antigen-presenting cells and specific T cell subpopulations [69, 70]. AhR participates in the regulation of innate and adaptive immune responses in some models of infection. AhR is also an important protein to limit the inflammatory response. Deletion of AhR exacerbated the inflammatory response to Listeria monocytogenes, Toxoplasma gondii, and Plasmodium falciparum [71–73], and was confirmed in a model of Leishmania major infection [74]. There is strong evidence to suggest that AhR is a pivotal molecule in myocarditis. In 2016, it was first reported that AhR modulated the development of myocarditis during Trypanosoma cruzi infection. When model mice were infected with T. cruzi, parasitemia, inflammation, and fibrosis of the myocardium were significantly reduced in AhR−/− mice compared with wild-type mice through the reduction in reactive oxygen species (ROS) and cytokines [75]. Viral infection is the most common cause of myocarditis. No study has examined the relationship between AhR and viral myocarditis. However, Coogan et al. found that AhR activation increased the number of neutrophils in the lungs, which contributed to poor survival in mice with influenza virus infection [76]. It is possible that AhR modulates the inflammatory response in viral myocarditis. AhR is a promising line of research on myocarditis.
4.1. AhR and Hypertension
It has been reported that exposure to environmental pollutants, particularly traffic-related pollutants, could increase the risk for hypertension [77]; however, causation has not been determined. One possible mechanism is that AhR, as an important regulator of blood pressure, could be activated by abundant exogenous AhR ligands in environment pollutants, such as TCDD [10]. Support for this theory is provided by studies on AhR-/- mice. When AhR was knocked out, mice showed significantly elevated blood pressure as well as elevated Ang II and ET-1 [46]. Another study suggested that 3-methylcholanthrene, an exogenous AhR agonist, can induce high blood pressure associated with endothelial NO synthase (eNOS) inactivation [78]. Endothelial cell-specific AhR-null mice had hypotension, accompanied by increases in eNOS activity and NO production [79]. These findings suggest that AhR could serve as a therapeutic target in hypertension, or other AhR-regulated NO-dependent vascular diseases.
Besides endogenous and exogenous ligands, activation of AhR can also be influenced by genetic polymorphisms. Genetic polymorphisms of the AhR signaling pathway are reported to be closely associated with the pathogenesis of essential hypertension. The majority of single-nucleotide polymorphisms in the AhR pathway, such as rs2228099 (ARNT), rs1048943 (CYP1A1), rs762551 (CYP1A2), and rs1056836 (CYP1B1), are associated with susceptibility to hypertension. The genetic environment and gene–gene interactions in the AhR signaling pathway are reported to determine susceptibility to essential hypertension [80]. Therefore, it is possible that gene therapy targeting AhR signaling could be a potential candidate in the treatment of essential hypertension.
The advent of CRISPR/Cas9, a versatile genome-editing tool, has allowed for precision medicine based on the detection of genetic polymorphisms. CRISPR/Cas9-regulated genome editing is a powerful technology for gene therapy [81]. It is thought that CRISPR/Cas9 will provide great advancements in the potential treatment of hypertension.
It has recently been reported that AhR is expressed in immune cells such as Th17 cells and dendritic cells [70]. Mice lacking T cells exhibited reduced blood pressure increases with Ang II infusion [82]. Whether expression levels of AhR in immune cells influence blood pressure, and possible mechanisms, is an interesting potential area of research.
4.2. AhR and Atherosclerosis
Atherosclerosis mainly occurs in the intimal layer of the blood vessel wall, and is characterized by subendothelial lipid deposits, vascular smooth muscle cell migration and proliferation, and formation of foam cells in the subendothelial space [83]. Risk factors for cardiovascular diseases include vascular senescence and obesity. Chronic vascular inflammation and oxidative stress contribute to atherosclerosis [84, 85], but the molecular mechanisms are not well understood. Exposure to contaminants containing ligands of AhR (dioxins, TCDD, PAH, benzo(α)pyrene) are thought to promote the development and progression of atherosclerosis, indicating that AhR may play a role in the regulation of atherosclerosis [9, 86–88].
Vascular senescence, a risk factor for cardiovascular diseases, is an important factor in the development of atherosclerosis. Studies suggest that senescent vascular cells are present in human atherosclerotic lesions [89–91]. A study in 2014 reported that indoxyl sulfate regulated sirtuin 1 via AhR activation, promoting endothelial senescence [92]. It was suggested that endothelial senescence in atherosclerosis is linked to AhR activation.
Obesity is also a vital contributor to atherosclerosis. In an AhR-directed luciferase-expressing mouse hepatocyte cell line, treatment with oxidized low-density lipoprotein and transforming growth factor-β1 could induce lipid accumulation and luciferase expression, owing to the overexpression of kynurenine, an endogenous AhR ligand, by enhanced indoleamine 2,3-dioxygenase 1 activity. Inhibition of AhR, in turn, prevented obesity [93].
Inflammatory responses contribute to AhR-regulated atherosclerosis. The inflammation-related cytokine monocyte chemoattractant protein-1 (MCP-1), an important endothelium-derived chemokine, was reported to recruit monocytes into the subendothelial space where they differentiated into macrophages, promoting atherosclerotic plaque development [94]. Activation of AhR by TCDD induced the release of a number of inflammatory mediators, including MCP-1, in ApoE-/- mice, leading to the promotion of atherosclerotic lesions and the formation of foam cells [6, 95, 96]. Treatment with CH223191, an AhR antagonist, significantly reduced the development of atherosclerotic lesions induced by TCDD. Expression of MCP-1 triggered by the AhR agonists indoxyl sulfate and coplanar polychlorinated biphenyl 77 could be reversed with the AhR antagonists CH223191 and α-naphthoflavone, respectively[94, 97]. ROS also play an important role in AhR-related atherosclerosis. There is evidence to suggest that ROS are involved in the process of indoxyl sulfate-induced MCP-1 production[98]. Indoxyl sulfate activates AhR and promotes ROS production by enhancing NADPH oxidase 4 expression in human umbilical vein endothelial cells. The AhR antagonist, CH223191, could reverse indoxyl sulfate-induced NADPH oxidase 4 expression [99]. A recent report showed that TCDD increased ROS production in endothelial cells and reduced NO-related vasodilation by AhR-dependent pathway which may be mediated, in part, via induction of cytochrome CYP1A1 [88]. Intercellular adhesion molecule-1 and matrix metalloproteinases, both regulated by AhR, may play a role in atherosclerosis [9, 100]; however, the mechanism remains to be determined. Taken together, these studies suggest that AhR could be a potential drug target to interfere with the development and progression of atherosclerosis.
4.3. AhR and Ischemic Heart Disease
Ischemic heart disease, including ST-segment elevation myocardial infarction, non-ST-segment elevation myocardial infarction, and stable and unstable angina pectoris, has high global morbidity and mortality. Coronary artery occlusion or stenosis stemming from coronary atherosclerosis is the major cause of ischemic heart disease [12]. Numerous studies have shown that environment pollutants associated with AhR signaling are important factors in atherosclerosis [9, 86–88]. There is evidence to indicate that exposure to exogenous ligands of AhR, such as dioxin, TCDD, and coplanar polychlorinated biphenyls, increase the risk for ischemic heart disease [12]. A study examined the role of AhR in coronary artery disease susceptibility in a Chinese population, and the results suggested that expression of circulating AhR was elevated in patients with coronary artery disease. Further analysis of AhR polymorphisms found that AhR rs2066853 showed a significant correlation with the risk for coronary artery disease [101]. A study at Stanford University reported that the transcription factor TCF21 promoted the expression of inflammation-related genes in human coronary artery smooth muscle cells via interaction with AhR, leading to an increased risk for coronary artery disease [102]. Xue et al. reported that, in myocardial ischemic injury, AhR gave rise to substantial expression of inflammatory cytokines, including high-sensitivity C-reactive protein, interleukin-1β, and interleukin-6. However, baicalin, a flavonoid compound, could attenuate the inflammatory response and myocardial injury via suppression of the expression of AhR [103]. Taken together, the findings suggest that AhR may be an important gene or drug target for the prevention and treatment of ischemic heart disease.
4.4. AhR and Myocardial Ischemia-Reperfusion Injury
Myocardial ischemia-reperfusion injury is a byproduct of reperfusion after acute myocardial infarction (reperfusion being the best treatment for acute myocardial infarction), and results in cardiomyocyte dysfunction and aggravated myocardial tissue injury [104]. Over the past few decades, researchers have found that ischemic post-conditioning (a treatment other than the traditional ischemic preconditioning) has a cardioprotective effect on ischemic reperfusion injury. In 2013, Vilahur et al. demonstrated that ischemic post-conditioning exerted a protective effect on cardiac structure and function via downregulation of the AhR signaling pathway [105]. Ischemic post-conditioning for myocardial ischemic reperfusion injury could reduce the expression of AhR and ARNT, resulting in a decrease in apoptosis. A recent study found that flavonoids capable of inhibiting AhR have a dual character in myocardial ischemia-reperfusion injury, either protection or deterioration [106]. While there are inconsistencies in the studies, AhR appears to be a significant mediator in myocardial ischemia-reperfusion injury.
4.5. AhR and Pulmonary Arterial Hypertension
Pulmonary arterial hypertension, a malignant chronic progressive vascular disease, usually leads to right heart failure and death in the late stage. There are various strategies for the treatment of pulmonary arterial hypertension, such as phosphodiesterase type 5 inhibitor, prostanoids (PGI2), prostacyclin, and endothelin receptor antagonists [107]. However, there is no ideal efficacy among the therapeutic treatments. A new effective therapy is needed, and the role of AhR should be further studied.
A study reported that baicalin, a natural flavone, could attenuate the abnormal proliferation of human pulmonary artery smooth muscle cells induced by TGF-β1 via inhibition of the HIF-1α and AhR pathways, indicating the participation of the AhR pathway in the progression of pulmonary arterial hypertension [108]. In addition, HIF-1α and AhR pathways could interact with each other through ARNT [22]. A study suggested that hypoxic pulmonary hypertension could be attenuated by suppressing HIF-1α triggered by hypoxia via RNA interference [109]. Studies examining the effect of the suppression of AhR in combination with suppression of HIF-1α in pulmonary arterial hypertension could prove to be useful. Therefore, AhR could be a potential drug target for the treatment of pulmonary arterial hypertension.
5. Conclusion
With an increased understanding of the link between environmental pollutants and cardiovascular diseases, the impact of AhR on the cardiovascular system has become evident. AhR plays an important role in maintaining cardiovascular homeostasis. In the cardiovascular system, the absence of AhR can result in abnormal cardiac function, hypertension or hypotension, vascular dysfunction, and cardiovascular disease. The types of cardiovascular disease include myocarditis, hypertension, atherosclerosis, ischemic heart disease, and pulmonary arterial hypertension. The pathogenesis induced by AhR varies among cardiovascular diseases, but includes inflammatory responses, immune responses, oxidative stress, and endothelial dysfunction. The molecular mechanisms behind AhR signaling, crosstalk between AhR signaling and other signaling pathways, and genetic polymorphisms require further study. Genetic polymorphisms of AhR will provide valuable information for guiding targeted gene therapy. Despite progress in our understanding of AhR-relating cardiovascular diseases, crosstalk between the cardiovascular system and the microenvironment is unclear. For targeted medical therapy, an effective dosage is hard to determine. And because most agonists and antagonists of AhR are not tissue-specific, further development is required. Therefore, clinical application of AhR-related therapies still has a long way to go.
AhR is a key bridging molecule in the cardiovascular system. Problems with AhR ligands and AhR transcripts can lead to abnormal activation of AhR, and result in an unbalanced cardiovascular system and cardiovascular diseases. AhR is a potential drug or gene target for the treatment of cardiovascular diseases.
Acknowledgments
This paper was supported by National Natural Science Foundation of China (no. 81300035 and 81741129), Collaborative Innovation and Platform Environment Construction Projects of Guangdong Province (2015A050502049), Natural Science Foundation of Guangdong Province (2015A030313520 and 2016A020214017) and Characteristic Innovation Project of Department of Education of Guangdong Province (2016KTSCX048).
Contributor Information
Yuanlin Ding, Email: gdmcsbd@163.com.
Can Chen, Email: chencan-21@163.com.
Wei Lei, Email: leiwei2006@126.com.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- 1.Nebert D. W. Aryl hydrocarbon receptor (AHR): “pioneer member” of the basic-helix/loop/helix per-Arnt-sim (bHLH/PAS) family of “sensors” of foreign and endogenous signals. Progress in Lipid Research. 2017;67:38–57. doi: 10.1016/j.plipres.2017.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hao N., Whitelaw M. L. The emerging roles of AhR in physiology and immunity. Biochemical Pharmacology. 2013;86(5):561–570. doi: 10.1016/j.bcp.2013.07.004. [DOI] [PubMed] [Google Scholar]
- 3.Poland A., Palen D., Glover E. Tumour promotion by TCDD in skin of HRS/J hairless mice. Nature. 1982;300(5889):271–273. doi: 10.1038/300271a0. [DOI] [PubMed] [Google Scholar]
- 4.Murray I. A., Patterson A. D., Perdew G. H. Aryl hydrocarbon receptor ligands in cancer: friend and foe. Nature Reviews Cancer. 2014;14(12):801–814. doi: 10.1038/nrc3846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hoffman E. C., Reyes H., Chu F.-F., et al. Cloning of a factor required for activity of the Ah (dioxin) receptor. Science. 1991;252(5008):954–958. doi: 10.1126/science.1852076. [DOI] [PubMed] [Google Scholar]
- 6.Sherr D. H. Another important biological function for the aryl hydrocarbon receptor. Arteriosclerosis, Thrombosis, and Vascular Biology. 2011;31(6):1247–1248. doi: 10.1161/ATVBAHA.111.227553. [DOI] [PubMed] [Google Scholar]
- 7.Xiao L., Zhang Z., Luo X. Roles of xenobiotic receptors in vascular pathophysiology. Circulation Journal. 2014;78(7):1520–1530. doi: 10.1253/circj.CJ-14-0343. [DOI] [PubMed] [Google Scholar]
- 8.Chuang K.-J., Yan Y.-H., Chiu S.-Y., Cheng T.-J. Long-term air pollution exposure and risk factors for cardiovascular diseases among the elderly in Taiwan. Occupational and Environmental Medicine. 2011;68(1):64–68. doi: 10.1136/oem.2009.052704. [DOI] [PubMed] [Google Scholar]
- 9.Oesterling E., Toborek M., Hennig B. Benzo[a]pyrene induces intercellular adhesion molecule-1 through a caveolae and aryl hydrocarbon receptor mediated pathway. Toxicology and Applied Pharmacology. 2008;232(2):309–316. doi: 10.1016/j.taap.2008.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kopf P. G., Huwe J. K., Walker M. K. Hypertension, cardiac hypertrophy, and impaired vascular relaxation induced by 2,3,7,8-Tetrachlorodibenzo-p-Dioxin are associated with increased superoxide. Cardiovascular Toxicology. 2008;8(4):181–193. doi: 10.1007/s12012-008-9027-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Savouret J.-F., Berdeaux A., Casper R. F. The aryl hydrocarbon receptor and its xenobiotic ligands: A fundamental trigger for cardiovascular diseases. Nutrition, Metabolism & Cardiovascular Diseases. 2003;13(2):104–113. doi: 10.1016/S0939-4753(03)80026-1. [DOI] [PubMed] [Google Scholar]
- 12.Dalton T. P., Kerzee J. K., Wang B., et al. Dioxin exposure is an environmental risk factor for ischemic heart disease. Cardiovascular Toxicology. 2001;1(4):285–298. doi: 10.1385/CT:1:4:285. [DOI] [PubMed] [Google Scholar]
- 13.Hanieh H. Toward understanding the role of aryl hydrocarbon receptor in the immune system: current progress and future trends. BioMed Research International. 2014;2014:14. doi: 10.1155/2014/520763.520763 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hernández-Ochoa I., Karman B. N., Flaws J. A. The role of the aryl hydrocarbon receptor in the female reproductive system. Biochemical Pharmacology. 2009;77(4):547–559. doi: 10.1016/j.bcp.2008.09.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Vasquez A., Atallah-Yunes N., Smith F. C., et al. A role for the aryl hydrocarbon receptor in cardiac physiology and function as demonstrated by AhR knockout mice. Cardiovascular Toxicology. 2003;3(2):153–163. doi: 10.1385/ct:3:2:153. [DOI] [PubMed] [Google Scholar]
- 16.Lund A. K., Goens M. B., Kanagy N. L., Walker M. K. Cardiac hypertrophy in Aryl hydrocarbon receptor null mice is correlated with elevated angiotensin II, endothelin-1, and mean arterial blood pressure. Toxicology and Applied Pharmacology. 2003;193(2):177–187. doi: 10.1016/j.taap.2003.08.008. [DOI] [PubMed] [Google Scholar]
- 17.Pang L.-P., Li Y., Zou Q.-Y., et al. ITE inhibits growth of human pulmonary artery endothelial cells. Experimental Lung Research. 2017;43(8):283–292. doi: 10.1080/01902148.2017.1367868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Pohjanvirta R., editor. The Ah receptor in biology and toxicology. NJ, USA: John Wiley & Sons, Inc; 2012. [Google Scholar]
- 19.Schulte K. W., Green E., Wilz A., Platten M., Daumke O. Structural Basis for Aryl Hydrocarbon Receptor-Mediated Gene Activation. Structure. 2017;25(7):1025–1033.e3. doi: 10.1016/j.str.2017.05.008. [DOI] [PubMed] [Google Scholar]
- 20.Swanson H. I., Bradfield C. A. The AH-receptor: genetics, structure and function. Pharmacogenetics. 1993;3(5):213–230. doi: 10.1097/00008571-199310000-00001. [DOI] [PubMed] [Google Scholar]
- 21.Dolwick K. M., Schmidt J. V., Carver L. A., Swanson H. I., Bradfield C. A. Cloning and expression of a human Ah receptor cDNA. Molecular Pharmacology. 1993;44(5):911–917. [PubMed] [Google Scholar]
- 22.Tsuji N., Fukuda K., Nagata Y., et al. The activation mechanism of the aryl hydrocarbon receptor (AhR) by molecular chaperone HSP90. FEBS Open Bio. 2014;4:796–803. doi: 10.1016/j.fob.2014.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Yamamoto J., Ihara K., Nakayama H., et al. Characteristic expression of aryl hydrocarbon receptor repressor gene in human tissues: Organ-specific distribution and variable induction patterns in mononuclear cells. Life Sciences. 2004;74(8):1039–1049. doi: 10.1016/j.lfs.2003.07.022. [DOI] [PubMed] [Google Scholar]
- 24.Esser C., Rannug A. The aryl hydrocarbon receptor in barrier organ physiology, immunology, and toxicology. Pharmacological Reviews. 2015;67(2):259–279. doi: 10.1124/pr.114.009001. [DOI] [PubMed] [Google Scholar]
- 25.Jiang Y.-Z., Wang K., Fang R., Zheng J. Expression of aryl hydrocarbon receptor in human placentas and fetal tissues. Journal of Histochemistry & Cytochemistry. 2010;58(8):679–685. doi: 10.1369/jhc.2010.955955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Nguyen L. P., Bradfield C. A. The search for endogenous activators of the aryl hydrocarbon receptor. Chemical Research in Toxicology. 2008;21(1):102–116. doi: 10.1021/tx7001965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Stejskalova L., Dvorak Z., Pavek P. Endogenous and exogenous ligands of aryl hydrocarbon receptor: current state of art. Current Drug Metabolism. 2011;12(2):198–212. doi: 10.2174/138920011795016818. [DOI] [PubMed] [Google Scholar]
- 28.Chen H.-S., Perdew G. H. Subunit composition of the heteromeric cytosolic aryl hydrocarbon receptor complex. The Journal of Biological Chemistry. 1994;269(44):27554–27558. [PubMed] [Google Scholar]
- 29.Zhang N., Walker M. K. Crosstalk between the aryl hydrocarbon receptor and hypoxia on the constitutive expression of cytochrome P4501A1 mRNA. Cardiovascular Toxicology. 2007;7(4):282–290. doi: 10.1007/s12012-007-9007-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Puga A., Barnes S. J., Dalton T. P., Chang C.-Y., Knudsen E. S., Maier M. A. Aromatic hydrocarbon receptor interaction with the retinoblastoma protein potentiates repression of E2F-dependent transcription and cell cycle arrest. The Journal of Biological Chemistry. 2000;275(4):2943–2950. doi: 10.1074/jbc.275.4.2943. [DOI] [PubMed] [Google Scholar]
- 31.Barnes-Ellerbe S., Knudsen K. E., Puga A. 2,3,7,8-Tetrachlorodibenzo-p-dioxin blocks androgen-dependent cell proliferation of LNCaP cells through modulation of pRB phosphorylation. Molecular Pharmacology. 2004;66(3):502–511. doi: 10.1124/mol.104.000356. [DOI] [PubMed] [Google Scholar]
- 32.Marlowe J. L., Fan Y., Chang X., et al. The aryl hydrocarbon receptor binds to E2F1 and inhibits E2F1-induced apoptosis. Molecular Biology of the Cell (MBoC) 2008;19(8):3263–3271. doi: 10.1091/mbc.E08-04-0359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Denison M. S., Soshilov A. A., He G., DeGroot D. E., Zhao B. Exactly the same but different: promiscuity and diversity in the molecular mechanisms of action of the aryl hydrocarbon (dioxin) receptor. Toxicological Sciences. 2011;124(1):1–22. doi: 10.1093/toxsci/kfr218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Levine-Fridman A., Chen L., Elferink C. J. Cytochrome P4501A1 Promotes G1 Phase Cell Cycle Progression by Controlling Aryl Hydrocarbon Receptor Activity. Molecular Pharmacology. 2004;65(2):461–469. doi: 10.1124/mol.65.2.461. [DOI] [PubMed] [Google Scholar]
- 35.Kimura A., Naka T., Nakahama T., et al. Aryl hydrocarbon receptor in combination with Stat1 regulates LPS-induced inflammatory responses. The Journal of Experimental Medicine. 2009;206(9):2027–2035. doi: 10.1084/jem.20090560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Apetoh L., Quintana F. J., Pot C., et al. The aryl hydrocarbon receptor interacts with c-Maf to promote the differentiation of type 1 regulatory T cells induced by IL-27. Nature Immunology. 2010;11(9):854–861. doi: 10.1038/ni.1912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Helle J., Bader M. I., Keiler A. M., et al. Cross-Talk in the female rat mammary gland: Influence of aryl hydrocarbon receptor on estrogen receptor signaling. Environmental Health Perspectives. 2016;124(5):601–610. doi: 10.1289/ehp.1509680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zhang S., Patel A., Chu C., et al. Aryl hydrocarbon receptor is necessary to protect fetal human pulmonary microvascular endothelial cells against hyperoxic injury: mechanistic roles of antioxidant enzymes and RelB. Toxicology and Applied Pharmacology. 2015;286(2):92–101. doi: 10.1016/j.taap.2015.03.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Braeuning A., Köhle C., Buchmann A., Schwarz M. Coordinate regulation of cytochrome P450 1a1 expression in mouse liver by the aryl hydrocarbon receptor and the β-catenin pathway. Toxicological Sciences. 2011;122(1):16–25. doi: 10.1093/toxsci/kfr080. [DOI] [PubMed] [Google Scholar]
- 40.Procházková J., Kabátková M., Bryja V., et al. The interplay of the aryl hydrocarbon receptor and β-catenin alters both AhR-dependent transcription and wnt/β-catenin signaling in liver progenitors. Toxicological Sciences. 2011;122(2):349–360. doi: 10.1093/toxsci/kfr129. [DOI] [PubMed] [Google Scholar]
- 41.Vogel C. F. A., Sciullo E., Li W., Wong P., Lazennec G., Matsumura F. RelB, a new partner of aryl hydrocarbon receptor-mediated transcription. Molecular Endocrinology. 2007;21(12):2941–2955. doi: 10.1210/me.2007-0211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Miao W., Hu L., Scrivens P. J., Batist G. Transcriptional regulation of NF-E2 p45-related factor (NRF2) expression by the aryl hydrocarbon receptor-xenobiotic response element signaling pathway: direct cross-talk between phase I and II drug-metabolizing enzymes. The Journal of Biological Chemistry. 2005;280(21):20340–20348. doi: 10.1074/jbc.m412081200. [DOI] [PubMed] [Google Scholar]
- 43.Fernandez-Salguero P. M., Ward J. M., Sundberg J. P., Gonzalez F. J. Lesions of aryl-hydrocarbon receptor-deficient mice. Veterinary Pathology. 1997;34(6):605–614. doi: 10.1177/030098589703400609. [DOI] [PubMed] [Google Scholar]
- 44.Kim D. W., Gazourian L., Quadri S. A., Sherr D. H., Sonenshein G. E. The RelA NF-κB subunit and the aryl hydrocarbon receptor (AhR) cooperate to transactivate the c-myc promoter in mammary cells. Oncogene. 2000;19(48):5498–5506. doi: 10.1038/sj.onc.1203945. [DOI] [PubMed] [Google Scholar]
- 45.Lund A. K., Goens M. B., Nuñez B. A., Walker M. K. Characterizing the role of endothelin-1 in the progression of cardiac hypertrophy in aryl hydrocarbon receptor (AhR) null mice. Toxicology and Applied Pharmacology. 2006;212(2):127–135. doi: 10.1016/j.taap.2005.07.005. [DOI] [PubMed] [Google Scholar]
- 46.Sauzeau V., Carvajal-González J. M., Riolobos A. S., et al. Transcriptional factor Aryl hydrocarbon receptor (Ahr) controls cardiovascular and respiratory functions by regulating the expression of the Vav3 proto-oncogene. The Journal of Biological Chemistry. 2011;286(4):2896–2909. doi: 10.1074/jbc.M110.187534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wang Q., Kurita H., Carreira V. Ah receptor activation by dioxin disrupts activin, BMP, and WNT signals during the early differentiation of mouse embryonic stem cells and inhibits cardiomyocyte functions. Toxicological Sciences. 2016;149(2):346–357. doi: 10.1093/toxsci/kfv246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Zhu C., Chen Y.-L., Wang X.-J., Hu X.-S., Yu Z.-B., Han S.-P. ShRNA-mediated gene silencing of AHR promotes the differentiation of P19 mouse embryonic carcinoma cells into cardiomyocytes. Molecular Medicine Reports. 2012;6(3):513–518. doi: 10.3892/mmr.2012.941. [DOI] [PubMed] [Google Scholar]
- 49.Wang Q., Chen J., Ko C.-I., et al. Disruption of aryl hydrocarbon receptor homeostatic levels during embryonic stem cell differentiation alters expression of homeobox transcription factors that control cardiomyogenesis. Environmental Health Perspectives. 2013;121(11-12):1334–1343. doi: 10.1289/ehp.1307297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Ko C.-I., Fan Y., de Gannes M., Wang Q., Xia Y., Puga A. Repression of the Aryl Hydrocarbon Receptor Is Required to Maintain Mitotic Progression and Prevent Loss of Pluripotency of Embryonic Stem Cells. Stem Cells. 2016;34(12):2825–2839. doi: 10.1002/stem.2456. [DOI] [PubMed] [Google Scholar]
- 51.Han Z., Miwa Y., Obikane H., et al. Aryl hydrocarbon receptor mediates laminar fluid shear stress-induced CYP1A1 activation and cell cycle arrest in vascular endothelial cells. Cardiovascular Research. 2008;77(4):809–818. doi: 10.1093/cvr/cvm095. [DOI] [PubMed] [Google Scholar]
- 52.Lahvis G. P., Lindell S. L., Thomas R. S., et al. Portosystemic shunting and persistent fetal vascular structures in aryl hydrocarbon receptor-deficient mice. Proceedings of the National Acadamy of Sciences of the United States of America. 2000;97(19):10442–10447. doi: 10.1073/pnas.190256997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Harstad E. B., Guite C. A., Thomae T. L., Bradfield C. A. Liver deformation in Ahr-null mice: Evidence for aberrant hepatic perfusion in early development. Molecular Pharmacology. 2006;69(5):1534–1541. doi: 10.1124/mol.105.020107. [DOI] [PubMed] [Google Scholar]
- 54.Bunger M. K., Glover E., Moran S. M., et al. Abnormal liver development and resistance to 2,3,7,8-tetrachlorodibenzo-p-dioxin toxicity in mice carrying a mutation in the DNA-Binding domain of the aryl hydrocarbon receptor. Toxicological Sciences. 2008;106(1):83–92. doi: 10.1093/toxsci/kfn149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Juan S.-H., Lee J.-L., Ho P.-Y., Lee Y.-H., Lee W.-S. Antiproliferative and antiangiogenic effects of 3-methylcholanthrene, an aryl-hydrocarbon receptor agonist, in human umbilical vascular endothelial cells. European Journal of Pharmacology. 2006;530(1-2):1–8. doi: 10.1016/j.ejphar.2005.11.023. [DOI] [PubMed] [Google Scholar]
- 56.Li Y., Wang K., Zou Q.-Y., Magness R. R., Zheng J. 2,3,7,8-Tetrachlorodibenzo-p-dioxin differentially suppresses angiogenic responses in human placental vein and artery endothelial cells. Toxicology. 2015;336:70–78. doi: 10.1016/j.tox.2015.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Cuartero M. I., Ballesteros I., De La Parra J., et al. L-kynurenine/aryl hydrocarbon receptor pathway mediates brain damage after experimental stroke. Circulation. 2014;130(23):2040–2051. doi: 10.1161/CIRCULATIONAHA.114.011394. [DOI] [PubMed] [Google Scholar]
- 58.Ichihara S., Yamada Y., Ichihara G., et al. A role for the aryl hydrocarbon receptor in regulation of ischemia-induced angiogenesis. Arteriosclerosis, Thrombosis, and Vascular Biology. 2007;27(6):1297–1304. doi: 10.1161/ATVBAHA.106.138701. [DOI] [PubMed] [Google Scholar]
- 59.Fritz W. A., Lin T.-M., Peterson R. E. The aryl hydrocarbon receptor (AhR) inhibits vanadate-induced vascular endothelial growth factor (VEGF) production in TRAMP prostates. Carcinogenesis. 2008;29(5):1077–1082. doi: 10.1093/carcin/bgn069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Huang S., Guo Y., Jacobi A., et al. Aromatic hydrocarbon receptor suppresses prostate cancer bone metastasis cells-induced vasculogenesis of endothelial progenitor cells under hypoxia. Cellular Physiology and Biochemistry. 2016;39(2):709–720. doi: 10.1159/000445662. [DOI] [PubMed] [Google Scholar]
- 61.Zhang N., Agbor L. N., Scott J. A., et al. An activated renin-angiotensin system maintains normal blood pressure in aryl hydrocarbon receptor heterozygous mice but not in null mice. Biochemical Pharmacology. 2010;80(2):197–204. doi: 10.1016/j.bcp.2010.03.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Lund A. K., Agbor L. N., Zhang N., et al. Loss of the aryl hydrocarbon receptor induces hypoxemia, endothelin-1, and systemic hypertension at modest altitude. Hypertension. 2008;51(3):803–809. doi: 10.1161/HYPERTENSIONAHA.107.100586. [DOI] [PubMed] [Google Scholar]
- 63.Villalobos-Molina R., Vázquez-Cuevas F. G., López-Guerrero J. J., et al. Vascular α1D-adrenoceptors are overexpressed in aorta of the aryl hydrocarbon receptor null mouse: Role of increased angiotensin II. Autonomic and Autacoid Pharmacology. 2008;28(2-3):61–67. doi: 10.1111/j.1474-8673.2008.00418.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Thackaberry E. A., Gabaldon D. M., Walker M. K., Smith S. M. Aryl hydrocarbon receptor null mice develop cardiac hypertrophy and increased hypoxia-inducible factor-1α in the absence of cardiac hypoxia. Cardiovascular Toxicology. 2002;2(4):263–273. doi: 10.1385/ct:2:4:263. [DOI] [PubMed] [Google Scholar]
- 65.Rose N. R. Viral myocarditis. Current Opinion in Rheumatology. 2016;28(4):383–389. doi: 10.1097/bor.0000000000000303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Comarmond C., Cacoub P. Myocarditis in auto-immune or auto-inflammatory diseases. Autoimmunity Reviews. 2017;16(8):811–816. doi: 10.1016/j.autrev.2017.05.021. [DOI] [PubMed] [Google Scholar]
- 67.Fung G., Luo H., Qiu Y., Yang D., McManus B. Myocarditis. Circulation Research. 2016;118(3):496–514. doi: 10.1161/CIRCRESAHA.115.306573. [DOI] [PubMed] [Google Scholar]
- 68.Quintana F. J. The aryl hydrocarbon receptor: a molecular pathway for the environmental control of the immune response. The Journal of Immunology. 2013;138(3):183–189. doi: 10.1111/imm.12046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Esser C., Rannug A., Stockinger B. The aryl hydrocarbon receptor in immunity. Trends in Immunology. 2009;30(9):447–454. doi: 10.1016/j.it.2009.06.005. [DOI] [PubMed] [Google Scholar]
- 70.Brant F., Miranda A. S., Esper L., et al. Role of the aryl hydrocarbon receptor in the immune response profile and development of pathology during Plasmodium berghei Anka infection. Infection and Immunity. 2014;82(8):3127–3140. doi: 10.1128/IAI.01733-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Sanchez Y., De Dios Rosado J., Vega L., et al. The unexpected role for the aryl hydrocarbon receptor on susceptibility to experimental toxoplasmosis. Journal of Biomedicine and Biotechnology. 2010;2010 doi: 10.1155/2010/505694.505694 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Shi L. Z., Faith N. G., Nakayama Y., Suresh M., Steinberg H., Czuprynski C. J. The aryl hydrocarbon receptor is required for optimal resistance to Listeria monocytogenes infection in mice. The Journal of Immunology. 2007;179(10):6952–6962. doi: 10.4049/jimmunol.179.10.6952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Elizondo G., Rodríguez-Sosa M., Estrada-Muñiz E., Gonzalez F. J., Vega L. Deletion of the aryl hydrocarbon receptor enhances the inflammatory response to leishmania major infection. International Journal of Biological Sciences. 2011;7(9):1220–1229. doi: 10.7150/ijbs.7.1220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Barroso A., Gualdrón-López M., Esper L., et al. The aryl hydrocarbon receptor modulates production of cytokines and reactive oxygen species and development of myocarditis during Trypanosoma cruzi infection. Infection and Immunity. 2016;84(10):3071–3082. doi: 10.1128/IAI.00575-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Head Wheeler J. L., Martin K. C., Lawrence B. P. Novel cellular targets of AhR underlie alterations in neutrophilic inflammation and inducible nitric oxide synthase expression during influenza virus infection. The Journal of Immunology. 2013;190(2):659–668. doi: 10.4049/jimmunol.1201341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Coogan P. F., White L. F., Jerrett M., et al. Air pollution and incidence of hypertension and diabetes mellitus in black women living in Los Angeles. Circulation. 2012;125(6):767–772. doi: 10.1161/circulationaha.111.052753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Chang C.-C., Hsu Y.-H., Chou H.-C., Lee Y.-C. G., Juan S.-H. 3-Methylcholanthrene/Aryl-Hydrocarbon Receptor-Mediated Hypertension Through eNOS Inactivation. Journal of Cellular Physiology. 2017;232(5):1020–1029. doi: 10.1002/jcp.25497. [DOI] [PubMed] [Google Scholar]
- 78.Agbor L. N., Elased K. M., Walker M. K. Endothelial cell-specific aryl hydrocarbon receptor knockout mice exhibit hypotension mediated, in part, by an attenuated angiotensin II responsiveness. Biochemical Pharmacology. 2011;82(5):514–523. doi: 10.1016/j.bcp.2011.06.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Polonikov A. V., Bushueva O. Y., Bulgakova I. V., et al. A comprehensive contribution of genes for aryl hydrocarbon receptor signaling pathway to hypertension susceptibility. Pharmacogenetics and Genomics. 2017;27(2):57–69. doi: 10.1097/FPC.0000000000000261. [DOI] [PubMed] [Google Scholar]
- 80.Zhang X., Wang L., Liu M., Li D. CRISPR/Cas9 system: a powerful technology for in vivo and ex vivo gene therapy. SCIENCE CHINA Life Sciences. 2017;60(5):468–475. doi: 10.1007/s11427-017-9057-2. [DOI] [PubMed] [Google Scholar]
- 81.Guzik T. J., Hoch N. E., Brown K. A., et al. Role of the T cell in the genesis of angiotensin II-induced hypertension and vascular dysfunction. The Journal of Experimental Medicine. 2007;204(10):2449–2460. doi: 10.1084/jem.20070657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Bonnefont-Rousselot D. Resveratrol and cardiovascular diseases. Nutrients. 2016;8(5, article no. 250) doi: 10.3390/nu8050250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Xuan Y., Shi Q., Liu G.-J., Luan Q.-X., Cai Y. Porphyromonas gingivalis Infection Accelerates Atherosclerosis Mediated by Oxidative Stress and Inflammatory Responses in ApoE-/- Mice. Clinical Laboratory. 2017;63(10):1627–1637. doi: 10.7754/Clin.Lab.2017.170410. [DOI] [PubMed] [Google Scholar]
- 84.Pernomian L., Da Silva C. H. T. P. Current basis for discovery and development of aryl hydrocarbon receptor antagonists for experimental and therapeutic use in atherosclerosis. European Journal of Pharmacology. 2015;764:118–123. doi: 10.1016/j.ejphar.2015.06.058.70093 [DOI] [PubMed] [Google Scholar]
- 85.Cole J. W., Xu H. Aryl Hydrocarbon Receptor Repressor Methylation: A Link between Smoking and Atherosclerosis. Circulation: Cardiovascular Genetics. 2015;8(5):640–642. doi: 10.1161/CIRCGENETICS.115.001243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Reynolds L. M., Wan M., Ding J., et al. DNA methylation of the aryl hydrocarbon receptor repressor associations with cigarette smoking and subclinical atherosclerosis. Circulation-Cardiovascular Genetics. 2015;8(5):707–716. doi: 10.1161/CIRCGENETICS.115.001097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Kopf P. G., Walker M. K. 2,3,7,8-Tetrachlorodibenzo-p-dioxin increases reactive oxygen species production in human endothelial cells via induction of cytochrome P4501A1. Toxicology and Applied Pharmacology. 2010;245(1):91–99. doi: 10.1016/j.taap.2010.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Tsirpanlis G. Cellular Senescence, Cardiovascular Risk, and CKD: A Review of Established and Hypothetical Interconnections. American Journal of Kidney Diseases. 2008;51(1):131–144. doi: 10.1053/j.ajkd.2007.07.035. [DOI] [PubMed] [Google Scholar]
- 89.Minamino T., Miyauchi H., Yoshida T., Tateno K., Kunieda T., Komuro I. Vascular cell senescence and vascular aging. Journal of Molecular and Cellular Cardiology. 2004;36(2):175–183. doi: 10.1016/j.yjmcc.2003.11.010. [DOI] [PubMed] [Google Scholar]
- 90.Minamino T., Miyauchi H., Yoshida T., Ishida Y., Yoshida H., Komuro I. Endothelial cell senescence in human atherosclerosis: role of telomere in endothelial dysfunction. Circulation. 2002;105(13):1541–1544. doi: 10.1161/01.CIR.0000013836.85741.17. [DOI] [PubMed] [Google Scholar]
- 91.Koizumi M., Tatebe J., Watanabe I., Yamazaki U., Ikeda T., Morita T. Aryl hydrocarbon receptor mediates indoxyl sulfate-induced cellular senescence in human umbilical vein endothelial cells. Journal of Atherosclerosis and Thrombosis. 2014;21(9):904–916. doi: 10.5551/jat.23663. [DOI] [PubMed] [Google Scholar]
- 92.Moyer B. J., Rojas I. Y., Kerley-Hamilton J. S., et al. Inhibition of the aryl hydrocarbon receptor prevents Western diet-induced obesity. Model for AHR activation by kynurenine via oxidized-LDL, TLR2/4, TGFβ, and IDO1. Toxicology and Applied Pharmacology. 2016;300:13–24. doi: 10.1016/j.taap.2016.03.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Majkova Z., Smart E., Toborek M., Hennig B. Up-regulation of endothelial monocyte chemoattractant protein-1 by coplanar PCB77 is caveolin-1-dependent. Toxicology and Applied Pharmacology. 2009;237(1):1–7. doi: 10.1016/j.taap.2009.02.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Vogel C. F. A., Sciullo E., Matsumura F. Activation of inflammatory mediators and potential role of Ah-receptor ligands in foam cell formation. Cardiovascular Toxicology. 2004;4(4):363–373. doi: 10.1385/CT:4:4:363. [DOI] [PubMed] [Google Scholar]
- 95.Wu D., Nishimura N., Kuo V., et al. Activation of aryl hydrocarbon receptor induces vascular inflammation and promotes atherosclerosis in apolipoprotein E-/- mice. Arteriosclerosis, Thrombosis, and Vascular Biology. 2011;31(6):1260–1267. doi: 10.1161/ATVBAHA.110.220202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Watanabe I., Tatebe J., Namba S., Koizumi M., Yamazaki J., Morita T. Activation of aryl hydrocarbon receptor mediates indoxyl sulfate-induced monocyte chemoattractant protein-1 expression in human umbilical vein endothelial cells. Circulation Journal. 2013;77(1):224–230. doi: 10.1253/circj.CJ-12-0647. [DOI] [PubMed] [Google Scholar]
- 97.Tumur Z., Shimizu H., Enomoto A., Miyazaki H., Niwa T. Indoxyl sulfate upregulates expression of ICAM-1 and MCP-1 by oxidative stress-induced NF-κB activation. American Journal of Nephrology. 2010;31(5):435–441. doi: 10.1159/000299798. [DOI] [PubMed] [Google Scholar]
- 98.Masai N., Tatebe J., Yoshino G., Morita T. Indoxyl sulfate stimulates monocyte chemoattractant protein-1 expression in human umbilical vein endothelial cells by inducing oxidative stress through activation of the NADPH oxidase-nuclear factor-κB pathway. Circulation Journal. 2010;74(10):2216–2224. doi: 10.1253/circj.CJ-10-0117. [DOI] [PubMed] [Google Scholar]
- 99.Meng D., Lv D.-D., Zhuang X., et al. Benzo[a]pyrene induces expression of matrix metalloproteinases and cell migration and invasion of vascular smooth muscle cells. Toxicology Letters. 2009;184(1):44–49. doi: 10.1016/j.toxlet.2008.10.016. [DOI] [PubMed] [Google Scholar]
- 100.Naito R., Miyauchi K. Coronary artery disease and type 2 diabetes mellitus current treatment strategies and future perspective. International Heart Journal. 2017;58(4):475–480. doi: 10.1536/ihj.17-191. [DOI] [PubMed] [Google Scholar]
- 101.Huang S., Shui X., He Y., et al. AhR expression and polymorphisms are associated with risk of coronary arterial disease in Chinese population. Scientific Reports. 2015;5:p. 8022. doi: 10.1038/srep08022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Kim J. B., Pjanic M., Nguyen T., et al. TCF21 and the environmental sensor aryl-hydrocarbon receptor cooperate to activate a pro-inflammatory gene expression program in coronary artery smooth muscle cells. PLoS Genetics. 2017;13(5) doi: 10.1371/journal.pgen.1006750.e1006750 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Xue Y., Shui X., Su W., et al. Baicalin inhibits inflammation and attenuates myocardial ischaemic injury by aryl hydrocarbon receptor. Journal of Pharmacy and Pharmacology. 2015;67(12):1756–1764. doi: 10.1111/jphp.12484. [DOI] [PubMed] [Google Scholar]
- 104.Neri M., Riezzo I., Pascale N., Pomara C., Turillazzi E. Ischemia/reperfusion injury following acute myocardial infarction: A critical issue for clinicians and forensic pathologists. Mediators of Inflammation. 2017;2017 doi: 10.1155/2017/7018393.7018393 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Vilahur G., Cubedo J., Casani L., et al. Reperfusion-triggered stress protein response in the myocardium is blocked by post-conditioning. Systems biology pathway analysis highlights the key role of the canonical aryl-hydrocarbon receptor pathway. European Heart Journal. 2013;34(27):2082–2093. doi: 10.1093/eurheartj/ehs211. [DOI] [PubMed] [Google Scholar]
- 106.Li W., Li Y., Sun R., et al. Dual character of flavonoids in attenuating and aggravating ischemia-reperfusion-induced myocardial injury. Experimental and Therapeutic Medicine. 2017;14(2):1307–1314. doi: 10.3892/etm.2017.4670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Jasinska-Stroschein M., Orszulak-Michalak D. Novel strategies for treatment of pulmonary arterial hypertension. Postepy Higieny I Medycyny Doswiadczalnej. 2017;71:577–588. doi: 10.5604/01.3001.0010.3838. [DOI] [PubMed] [Google Scholar]
- 108.Huang S. A., Chen P., Shui X., et al. Baicalin attenuates transforming growth factor-β1-induced human pulmonary artery smooth muscle cell proliferation and phenotypic switch by inhibiting hypoxia inducible factor-1α and aryl hydrocarbon receptor expression. Journal of Pharmacy and Pharmacology. 2014;66(10):1469–1477. doi: 10.1111/jphp.12273. [DOI] [PubMed] [Google Scholar]
- 109.Li Y., Shi B., Huang L., et al. Suppression of the expression of hypoxia-inducible factor-1α by RNA interference alleviates hypoxia-induced pulmonary hypertension in adult rats. International Journal of Molecular Medicine. 2016;38(6):1786–1794. doi: 10.3892/ijmm.2016.2773. [DOI] [PMC free article] [PubMed] [Google Scholar]