

Abstract
Pannexins belong to a family of ATP-release channels expressed in almost all cell types. An increasing body of literature on pannexins suggests that these channels play dual and sometimes contradictory roles, contributing to normal cell function, as well as to the pathological progression of disease. In this review, we summarize our understanding of pannexin “protective” and “harmful” functions in inflammation, regeneration and mechanical signaling. We also suggest a possible basis for pannexin’s dual roles, related to extracellular ATP and K+ levels and the activation of various types of P2 receptors that are associated with pannexin. Finally, we speculate upon therapeutic strategies related to pannexin using eyes, lacrimal glands, and peripheral nerves as examples of interesting therapeutic targets.
Keywords: pannexin, Panx1, ATP, purinergic signaling, inflammation, regeneration
Introduction
Cell–cell and cell–matrix interactions are fundamental properties of multicellular organisms. Gap junctions, formed by connexins and innexins in vertebrate and invertebrate animals, respectively, allow direct passage of ions and small molecules (<2,000 Da) from cell to cell (Figure 1A).1–3 In addition to gap-junction channels, connexins may form hemichannels (HCs), termed “connexons” (Figure 1B),4,5 which are hexamers of connexin monomers (Figure 1A), each containing four transmembrane domains, two extracellular loops, and cytoplasmic N and C termini (Figure 1C).6 The vertebrate homologues of innexins, called “pannexins”, form mostly HCs, or pannexons (Figure 1B), due to the high level of glycosylation in their extracellular domains (Figure 1D).7–12 Similar to connexins (Figure 1C), pannexins have a cytosolic N-terminal domain, four transmembrane domains with two extracellular loops, and a cytosolic C-terminal domain (Figure 1D).13 However, pannexins have no homology to the vertebrate connexin gap-junction protein,8 and unlike connexins, which have multiple cysteine residues in both extracellular loops, pannexins have only two cysteine residues per loop (Figure 1C and D, black ovals).13
Figure 1.

Connexins and pannexins.
Notes: (A, B) Connexin and pannexin share a similar structure, despite the absence of sequence homology. Connexin and pannexin form functional connexon and pannexon hemichannels, respectively. (C, D) Connexins and pannexins are transmembrane proteins with four transmembrane domains, two extracellular loops, one cytoplasmic loop, and cytoplasmic N- and C-terminal domains. Connexin channels can assemble into a gap junction (A) that mediates intercellular communication, while pannexin’s extracellular loop has a high level of glycosylation in mammalian cells (D), which prevent the formation of gap junctions.
Abbreviation: Glyc, glycosylation.
The pannexin family consists of three proteins, Panx1, Panx2, and Panx3, all of which have been shown to form a single-membrane channel.14,15 Panx1 is ubiquitously expressed in almost all cell types, including those in the nervous and immune systems, eye, muscle, olfactory epithelium, blood vessels, exocrine glands (eg, lacrimal and salivary glands), thyroid, prostate, kidney, and liver (Table 1).16–20 Panx2 transcripts are highly expressed in the central nervous system (CNS).21 Lower levels of Panx2 transcripts have been detected in nonneural tissues, including the testis, kidney, retina, and gastrointestinal tract, while Panx3 mainly localizes in the skin, osteoblasts, and chondrocytes (Table 1).15,20–24 Panx3 has also been found in skeletal muscle,25 lactating mammary glands, sebaceous glands, and the small intestine.15 Interestingly, Panx2 protein appears to be more ubiquitously expressed than initially predicted by mRNA expression21 (Table 1).
Table 1.
Expression and localization of pannexin mRNA and protein
| Panx1 | Panx2 | Panx3 | |
|---|---|---|---|
| mRNA-expression pattern and levels of expression | Ubiquitously expressed in mammalian211 and chicken tissues212 | Cerebral cortex, cerebellum: neurons and immature astrocytes,21 hippocampus (high);22 kidney tubular cells (low), seminiferous duct cells in the testis (moderate), salivary glands (excretory), and striated ducts (weak)21 | In humans, Panx3 mRNA found in the testis, stomach, spleen, salivary gland, lung, heart, duodenum, and adrenal tissue;213 in mice, Panx3 mRNA found in developing skeletal structures214 |
| Protein expression | Ubiquitously expressed in many mammalian tissues,211,215 with lower levels in the lung, kidney, and heart ventricles13 | Protein is expressed in many tissue types, including gastrointestinal tract glandular and epithelial cells, parietal cells, columnar epithelial cells of the human colon216 (strong), and mouse retina, lung, and skin | Extensive expression throughout the developing skeleton during chondrocyte and osteoblast differentiation,214 skeletal muscle,25 lactating mammary tissue, sebaceous glands, and the small intestine15 |
| Localization within the cell | Mostly membrane,13,26,217 but may form permeable channels in the endoplasmic reticulum27 | Membrane,24,26 predominantly cytoplasmic21 | Membrane, may form channels in the endoplasmic reticulum and gap junctions after overexpression,218,219 diffuse cytoplasmic in the epidermis220 |
| Subunits | 6 | 6–8221 | 6 |
| Key domains (conserved in all isoforms) | Transmembrane domains (×4), extracellular loops (×2), intracellular N-terminus, C-terminus, and intracellular loop | ||
| Glycosylation site | Second extracellular loop | First extracellular loop | First extracellular loop |
Endogenous Panx1 and Panx3 proteins are localized primarily at the plasma membrane,13,26 while Panx2 is highly expressed in the cytoplasmic compartment,21 suggesting a unique intracellular function for Panx2. However, several studies have reported the cytoplasmic localization of Panx1 and Panx3 proteins when these proteins were overexpressed in cells.21,27 For example, Abeele et al27 demonstrated cytoplasmic localization of Panx1 transiently expressed in LNCaP cells, where it formed Ca2+-permeable channels in the endoplasmic reticulum (Table 1). It is quite possible that high levels of pannexin protein expression could lead to both membrane and endoplasmic reticulum-channel formation, thus contributing to sustained increases in intracellular Ca2+.
Pannexins are ATP-release channels that can be activated by caspase cleavage of their pore-associated C-terminal tail, the autoregulatory region controlling channel permeability. The regulated ATP (nucleotide) release through pannexin HCs is implicated in a number of normal physiological functions and in response to stressors or pathological states in cells and tissue.25,28,29 Well-characterized functions of pannexins include regulation of cell differentiation and migration, tissue development and regeneration, inflammation, wound healing and cell death.28 However, mechanistic explanations of how these proteins perform sometimes contradictory roles remain unclear.
In this review, we attempt to clarify existing controversies in the literature on the “protective” and “harmful” roles of pannexin HCs by addressing a question: How do pannexins acquire these different and often opposing roles? We seek to obtain deeper understanding of pannexin signaling, participants in which represent a potential source of novel and promising therapeutic targets in a variety of pathologies. Our focus is entirely on pannexins, with a full understanding that pannexin and connexin HCs have both distinct and complementary but often overlapping functions, particularly in ATP release and inflammation; therefore, we refer the reader to several excellent reviews comparing the roles of these channels.30–33
Pannexins, inflammation, and inflammasome activation
The involvement of pannexins in the induction of inflammation has been reported in multiple publications.28,34–36 Inflammation is the major protective function maintained by the evolutionarily conserved innate immune system in response to harmful stimuli, such as pathogens, stress, injury, or cell death. Acute (short-term) inflammation stimulates a regenerative response, while persistent (chronic) inflammation can cause systemic inflammatory diseases.37 Activation of inflammasomes, facilitating the release of interleukin-1β (IL1β) and IL18 in response to pathogens and tissue injury, is a key function of the innate immune system. The inflammasomes, first characterized in monocytes in 200238 and in neural cells in 2008,39 are multi-protein complexes mediating proteolytic maturation of Casp1, Casp11, IL1β, and IL18. Proteolytic cleavage of IL1β and IL18 precursors is executed by active Casp1 (Figure 2);40,41 and the release of the mature cytokines occurs via megapores, formed by N-terminal domains of the Casp1/11-processed recently identified pore forming protein gasdermin D.42–46 A large body of experimental evidence identifies Panx1 and its associated P2X receptors as essential upstream regulators of inflammasomes and proteolytic activation of Casp1 and Casp11.47–51 Panx1 has been reported to activate inflammasomes in many cell types, including macrophages,52,53 microglia,54 neurons, and astrocytes;49 however, the data on particular cell and inflammasome types remain controversial.55,56 Currently, the bulk of published data support a pivotal role for Panx1 in CNS/ retinal inflammasome regulation.55–58 As such, strong suppression of its major components, including Casp1, Casp11, IL1β, and apoptosis-associate speck-like protein containing a caspase recruitment domain (ASC), is observed in both Panx1−/− mice and wild-type retinas after Panx1 blockade by probenecid.16,54,59
Figure 2.

The two signaling arms of the inflammasome-activation cascade.
Notes: Signal 1 pathways sense environmental signals via surface Tumor necrosis factor (TNF), Toll-like (TLR) and IL-1 receptors and facilitates transcriptional priming of inflammasome components via the NFκB pathway and upregulates the expression of precursor proteins of IL1β, caspases 1/11(also known as caspase 4), and pro-Nod-like receptors (NLR). Signal 2 facilitates activation of the complex via proteolytic processing and assembly. This arm responds to mechanical stress, activation of a ligand-sensing system within the cytosol or extracellular ATP sensing via Panx1–P2X receptor signalosomes. Upon activation, protease activity of caspases regulates the maturation and release of IL1β and IL18. Recent studies showed that Gasdermin D (GSDMD) is a novel membrane pore-forming protein. Cleaved by inflammatory caspases Casp1 or Casp11(4), GSDMD binds to phosphoinositides in the plasma membrane and oligomerizes to generate membrane pores of ~10–14 nm in diameter.222 This pore size can allow the passage of mature IL1β, IL18, and caspase 1. The formation of the GSDMD pores also disrupts osmotic potential, resulting in an inflammatory form of cell death known as pyroptosis.
Abbreviation: ASC, apoptosis-associate speck-like protein containing a caspase recruitment domain.
There are two major regulatory arms for inflammasome activation (Figure 2): signal 1 pathways sense environmental signals via surface TNF, Toll-like, and IL1 receptors and facilitate inflammasome “priming”, ie, transcriptional activation via MyD88–NFκB-mediated pathways;60,61 and signal 2 pathways regulate inflammasome assembly and processing of Casp1/11, IL1β, and IL18 precursors. This arm is regulated via the Panx1–P2X signalosome to facilitate ATP and K+ release, as well as uptake of extracellular Ca2+ and danger/pathogen-signaling patterns.62,63
Though a role for Panx1 in the inflammasome regulatory cascade appears to be generally conserved across cell types, Qu et al55 suggested that pannexin is “dispensable” for inflammasome formation. In particular, LPS-primed bone marrow-derived macrophages were successfully able to activate Casp1 and secrete its associated inflammatory cytokines (IL1β and IL18) in response to a number of stimuli in the absence of Panx1. Moreover, the authors also concluded that P2X7 and Panx1 can function independently and may be involved in distinct signaling pathways.55 These controversial views on Panx1 function could be explained by cell-type-specific differences and potential variation in culture conditions, and need to be resolved.
Mechanisms of pannexin-channel activation
Several diverse mechanisms regulating pannexin-channel function have been proposed to date. Pannexin channels have been posited to be activated by caspase-mediated channel cleavage in apoptotic immune cells, G-protein-coupled receptors in vascular smooth muscle,64,65 low oxygen tension in erythrocytes and neurons,66 high extracellular K+ in various cell types,49,67 and mechanical stretch.68,69 Progressive Panx1-channel opening is directly linked to ion- and large-molecule transport, and occurs during both irreversible (caspase-mediated cleavage)70 and reversible G-protein-coupled receptor (including α1-adrenoceptor-mediated) forms of channel activation.71 Panx1 activation by caspase-mediated cleavage enables the release of ATP as a “find me” signal that recruits phagocytizing macrophages to apoptotic T lymphocytes.65,70 This mechanism is critical for the fast clearance of apoptotic and dead cells during acute inflammation.28,55,65,72 Cleavage activation of Panx1 is also involved in pyroptotic cell death (Figure 2).73 A recent study employing electron microscopy and single-channel recordings of full-length and caspase-cleaved pannexin concatemers with defined numbers (0–6) of intact and truncated C termini revealed that Panx1 activation was increased in a sequential manner by stepwise removal of the autoinhibitory C termini. This also resulted in a graded increase in current and ATP/dye permeation.71 On the other hand, the reversible G-protein-coupled receptor-mediated mechanism is independent of caspase-mediated pannexin cleavage.74 Comparison of α1-adrenoceptor-activated with cleavage-activated Panx1 channels indicated that α1-adrenoceptor-activated Panx1 channels had a shorter mean open time, but progressively increasing conductance, suggesting that despite differences in gating kinetics, activation of Panx1 channels by both signaling mechanisms involves cumulative changes in open-channel properties.71
Pannexin signaling via ATP release
Panx1 channels can release ATP under physiological conditions and play critical roles in many pathological processes. ATP is a prominent extracellular signaling molecule in both physiological and pathological conditions. For example, ATP release is important for muscle differentiation and function,75–78 and ATP-receptor activation plays a role in regulation of cell proliferation, DNA synthesis, cell differentiation, and cell survival during the course of CNS development.79,80 At the same time, ATP may also serve as a major danger signal for cells,50 despite it having a very short half-life due to rapid degradation by surface ecto-ATPases.81 ATP is released from apoptotic, injured, and viable cells that are challenged by assorted cytokines, as well as mechanical or ischemic stress in the presence of elevated K+.82
ATP-mediated activation of Panx1, the ATP-release channel, typically ramps up in a vicious cycle only to a certain level, due to a retrograde feedback mechanism regulating activity of Panx1 HCs via a low-affinity ATP binding site.83 Therefore, the permeant (ATP) can inhibit the permeating channel when high extracellular ATP concentration is reached. Importantly, however, this inhibition is abrogated by an increased extracellular concentration of potassium ions (K+),82 suggesting a mechanism of toxicity of extracellular ATP in Panx1-expressing cells. In agreement with this mechanism are the findings that massive activation by Casp3/7 cleavage or expression of constitutively active Panx1 HCs results in cell death.65,84 Therefore, the balance between physiological and pathological activities of Panx1 depends on the open-state probability of the channel, which in turn is influenced by the increase in intracellular Ca2+ and extracellular ATP and K+. An additional level of Panx1-channel regulation is achieved via interactions with purinergic P2 (eg, P2X and P2Y) receptors, which are activated by binding extracellular ATP at the plasma membrane.85 Several salient aspects of Panx–P2 interactions, including the mechanisms and significance of such interactions, as well as their sensitivity and specificity, are detailed in the following sections.
Functional interactions of Panx HCs with purinergic P2X and P2Y receptors
There are two major families of purinergic P2 receptors: ionotropic P2X and metabotropic P2Y receptors. Reciprocal interactions, whereby P2 receptors directly activate Panx1 channels,86,87 suggest that these proteins can form a signaling complex at the cell surface34,88 that mediates both paracrine and autocrine purinergic communication.
The P2X-receptor family contains 7 isoforms (P2X1–7), and P2X receptors are classified as ligand-gated channels whose activation regulates cellular membrane potential and intracellular Ca2+ levels.89,90 More precisely, though, P2X family members are ATP-gated cation channels, selective for Na+, K+, and Ca2+ ions.91 In the nervous system, P2X receptors are pivotal transducers of ATP-mediated paracrine signals and have been implicated in physiological functions, such as chemotactic cell migration, intercellular calcium-wave propagation, as well as in nervous system dysfunction, leading to neuropathic pain or cell death.92 Five isoforms – P2X1, P2X2, P2X3, P2X4, and P2X7 – have been shown to interact with Panx1,93,94 among which P2X4 and P2X7 are the most common interaction partners in different cell types.51,95,96 Both P2X4 and P2X7 isoforms are calcium channels known to dilate into larger pores upon activation.97,98 P2X7-receptor activation results in the appearance of HC-like currents, reflecting the channel permeability for molecules up to 1 kDa and identified as Panx1 HC.52 Under pathological conditions, overactivation of the Panx1–P2X7 signalosome complex has been implicated in inflammation, cell death, and neuropathic pain.34,96,99,100
The P2Y-receptor family contains eight isoforms (P2Y1,2,4,6,11–14). P2Y receptors are metabotropic G-protein-coupled receptors that couple to Gq, Gs, or Gi in an isoform-specific manner, and their activation modulates intracellular inositol triphosphate, Ca2+, and cAMP levels.101,102 Different isoforms of purinergic P2Y receptors are activated by ATP and its degradation products ADP and UTP, and couple to distinct G proteins to induce cAMP production, activation of phospholipase C, or intracellular Ca2+ via inositol triphosphate second-messenger systems.103,104 Recent publications have implicated P2Y receptors and Panx1 signaling in the regulation of endothelial cell activation in vascular inflammation105 and cell-volume regulation.106 ATP release via Panx1 channels activates P2Y2 receptors to amplify signaling in sensing chemotactic gradients in neutrophils.107,108 Polarization of surface expression by translocation of Panx1, P2Y2, adenosine A3 receptors, and ENTPD1 ectonucleotidase to the leading cell edge allows neutrophils to polarize within the gradients.
Sensitivity and specificity of Panx1–P2X signaling during normal tissue function and chronic inflammation
The activity and downstream consequences of several P2X- and P2Y-receptor isoforms are dependent on ATP binding, with varying sensitivity. P2X7, expressed by microglia, astrocytes, and neurons, is the most studied isoform, and is implicated in cyto- and neurotoxicity via direct interaction with Panx1 channels.34,87,88 Due to its relatively low affinity to ATP (EC50 936 µM), it can only be activated by nonphysiological (≥1 mM) increases in extracellular ATP, which is locally achievable only at sites of injury or proximal to activated Panx1/Cx43 HCs. Conversely, the activation at low to medium (<900 µM) extracellular ATP levels can be mediated via an interaction between the high-affinity P2X4 receptor (P2X4; EC50 2.3 µM)109 and Panx149,110 (Figure 3A and B). This synergistic interaction was shown to coactivate P2X7, resulting in massive local efflux of ATP via the Panx1 channel, a forward-feeding autocrine amplification loop (Figure 3C).111,112 Functional synergy between P2X4 and cytotoxic P2X7110,113–115 is known to be pivotal for Panx1-dependent extracellular ATP-induced cell death,116 which can be suppressed by a blockade of either component in the Panx1–P2X4/7 complex or by extracellular ATP removal with apyrase.51,96 Similarly, in the retina, genetic ablation or pharmacological inhibition of Panx1,117 P2X7, or P2X4118–120 has protected retinal ganglion cells (RGCs) in both acute and chronic ocular hypertension (OHT)-injury models. In other studies, a similar blockade was shown to protect neurons and other cell types from death via a rise in ionized Ca2+ and the induction of the inflammasome in various injury paradigms.34,121,122
Figure 3.
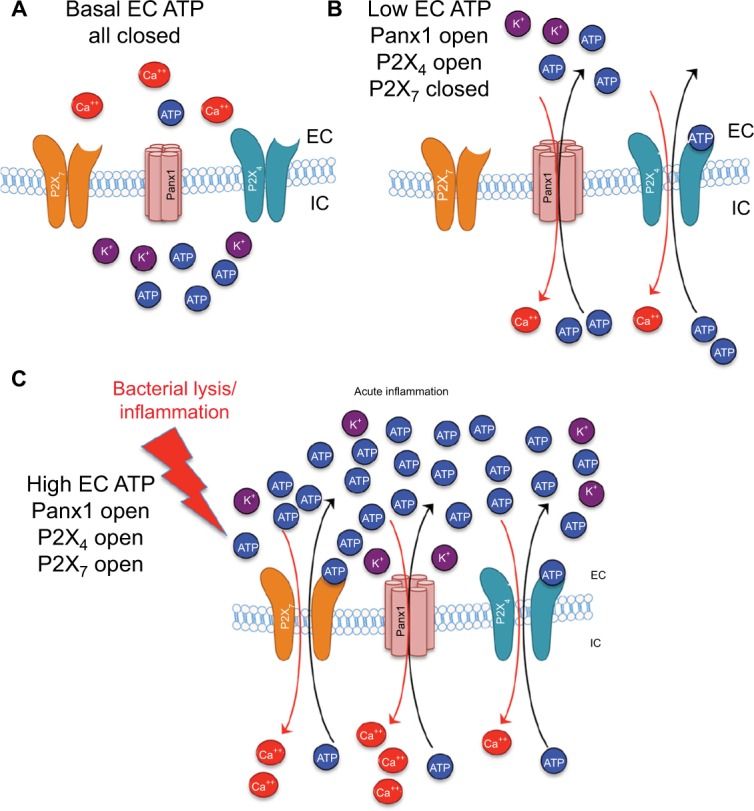
Differential ATP and ion movement depending on Panx1, P2X4, and P2X7 activity.
Notes: (A) Basal levels of EC ATP and normal concentration gradients of ATP and ions. (B) Panx1 opening with low levels of EC ATP results in P2X4 activation, but no P2X7 activity. (C) Higher levels of EC ATP, such as those resulting from bacterial lysis or chronic inflammation, result in opening of P2X4 and P2X7 channels and substantial movement of ATP and ions along their concentration gradients.
Abbreviations: EC, extracellular; IC, intracellular.
In contrast to P2X7, interactions between P2X4 receptor and Panx1 and their link to RGC loss and inflammasome in the retina still require exploration. However, strong evidence of a key cellular role of P2X4 in response to sublethal levels of ATP has been suggested in experiments on channel blockade with the 5-BDBD antagonist in macrophages,116 as well as on inflammasome activation in various tissue types.51,123 In contrast to P2X7, P2X4 blockade has been shown selectively to suppress IL1β but not IL18 cytokine levels,110 which was reported as potentially neuroprotective.124–126
More recently, an interesting phenomenon in acute-wound healing following the use of intracellular ATP delivery was described. In this study, ATP application was accompanied by a massive increase in macrophage trafficking, in situ proliferation, and direct collagen production within the wound.127 Although the signaling mechanism of this phenomenon has not been determined, other research128 has demonstrated that the recognition and clearance of dying cells and debris from focal points of inflammation is critical in both the induction and resolution of inflammation.28 Moreover, Panx1-mediated vesicular nucleotide transporters (responsible for ATP accumulation in secretory vesicles)-mediated ATP release have been shown to recruit neutrophils/macrophages to injury sites.129–131 It is quite possible that in some types of acute injury, an increase in or acceleration of postinjury inflammation may lead to more rapid resolution of inflammation through ADP or other signaling mechanisms.
Other mediators of Panx signaling and inflammation
In addition to Panx HCs, significant amounts of ATP can be released by bacteria, which trigger Panx1/P2X activity (Figure 3C).132 Bacterial ATP may affect different types of cells and lead to the production of proinflammatory cytokines and growth factors. A recent study showed that commensal bacteria-derived ATP activates CD70highCD11clow cells in the intestinal lamina propria, induce IL6 and IL23 production, as well as TGF-β pathway activation. This then led to local differentiation of IL17-producing CD4+ T lymphocytes (T-helper TH-17, cells involved in host defense and several immune disorders).133 Moreover, systemic or rectal administration of ATP into germ-free mice resulted in a marked increase in the number of lamina propria TH17 cells. The specific effect of ATP on TH17 differentiation was mediated by P2X and P2Y receptors, and ATP-induced TH17 differentiation was inhibited by P2X- and P2Y-receptor blockade. Interestingly, this mechanism commonly operates during the differentiation of both “naturally occurring” and “pathogenic” TH17 cells.127,133
Although ATP-gated unselective cation P2X channels are induced mainly by ATP, some studies report that they also may be activated by other molecules.134 β-Toxin produced by Clostridium perfringens is a key virulence factor in fatal hemorrhagic enterocolitis and enterotoxemia. This toxin belongs to a family of β-pore-forming toxins. The results of a recent study suggested that Panx1 opening is achieved through the interaction of β-toxin with the P2X7 receptor. Then, ATP released by Panx1-channel opening promotes oligomer formation of the toxin, leading to cell death.134 These studies suggest that Panx1 HC is an important contributor to P2X7-receptor signaling and provides a mechanistic link among bacterial stimuli, P2X7–Panx1 signaling, and inflammation.
Pannexins, mechanical signaling, and the cytoskeleton
A mechanosensitive role for connexin HCs in the propagation of intracellular calcium, initiated by the extracellular binding of ATP, was first noted in 1990.135 Numerous reports since then have demonstrated the sensitivity of connexin HCs to extracellular Ca2+, which are believed to keep connexin HCs in a closed state at physiological Ca2+ levels.136–138 In contrast, Panx HCs are not gated by external Ca2+,139 and the mechanical sensitivity of pannexin HCs was not noted until 2004, when single-channel currents were elicited by changes in pressure imposed pneumatically upon membrane patches of Xenopus oocytes expressing Panx1.140 Since then, mechanosensitive purinergic signaling pathways, including pannexin-mediated ATP release, have been demonstrated in many cell types in response to mechanical stimuli. For example, inhibition of pannexin function suppressed hypertonic stress-induced ATP release and reduced downstream transcriptional activation induced by hypertonicity,95,141 and inhibition of Panx1 and several P2X receptors reduced downstream transcriptional activation induced by hypertonicity. Similarly, pannexin- and/or P2-receptor-dependent ATP release has been observed in RGCs,69 lens epithelial cells,142 fibrosarcoma cells,143 urothelial cells,144 and astrocytes130 that were subjected to hypoosmotic conditions. In addition to altered tonicity, shear stress has been shown to activate mechanosensitive pannexin channels. Indeed, bone cells and red blood cells have demonstrated robust pannexin-mediated ATP release in response to oscillatory fluid shear stress.145,146 Consistent with this function, it was recently suggested that pannexin activity induced by transient fluid shear during media changes and manipulation of tissue-culture containers could confound the interpretation of cell-culture experiments.147
In contrast to the bulk of the literature, only one study in HEK293 cells subjected to hypotonic media has suggested that pannexin HCs are not directly mechanosensitive.148 It is likely that this controversial observation may have reflected a unique feature of the examined cell type and/or methodological differences. The authors’ choice to use ethidium bromide internalization as an indicator of pannexin HC activity may have led to different outcomes compared with more conventional and commonly used indicators of pannexin activity, such as dye uptake or ATP release.
While mechanical activation of pannexin HCs has been studied primarily in a general context, there is increasing evidence that mechanical signaling can facilitate pathological states, such as edema that stretches the plasma membrane. Mechanical strain was recently reported to trigger a robust inflammatory response, transcriptional priming of NLRP3 inflammasome formation, and IL1β production via activation of Panx1–P2X7 signaling.54
The eye has emerged as an important model in understanding pathological mechanotransductive roles for Panx1. When the retina is exposed to mechanical stress, ATP is released physiologically by glia and neurons via Panx1 channels.29,69,130 In OHT-injured retina, synergistic effects of mechanical stress induced by elevated pressure and massive ATP release facilitate sustained extracellular ATP elevation and prolonged activation of the Panx1–P2X pathway, a combination that is particularly toxic to RGCs, which are highly enriched in Panx1.119,130,149 Experimental data generated in the murine eye indicate that RGC loss and axonal damage strongly correlate with mechanical deformation150 and repetitive intraocular pressure spikes.29,130,151 An increase in extracellular ATP has been reported in eyes exposed to acute or chronic OHT in animal models, as well as in human primary open-angle glaucoma.69,111,149,151,152 Conversely, strain-activated, pannexin-regulated release of cytokines may also serve a protective role, as demonstrated by increased IL3 and IL6 expression in RGCs subjected to a 4% chronic strain in vitro or increased intraocular pressure in vivo.153 Furthermore, the release of IL18 via inflammasome activation has also been reported to be neuroprotective.125,154
Concurrent with Panx1-mediated ATP release, Cx43 HCs have been demonstrated as another key pathway of ATP release. Although the contribution of Cx43 vs Panx1 to ATP release has been heavily debated recently,155–157 the current consensus indicates that Panx1 channels initiate and Cx43 HCs facilitate the bulk of ATP release from macroglia, especially in the presence of TNFα and IL1β.158–160 Consistent with this view, upon their exposure to TNFα and IL1β cytokines, glial cells become activated and release ATP via Cx43 HCs.161
Given its structural role, the cytoskeleton is a leading candidate to participate in mechanical signaling. Actin dynamics, particularly those mediated by ARP2/3, have been reported to be regulated by pannexin activity,17 and pannexin interacts physically with actin (through the C terminus of Panx1), but not tubulin/microtubules.24,162 In a recent publication, it was reported that the Panx1–P2X7 autocrine loop induced by ATP increased the migration speed of dendritic cells by promoting reorganization of the actin cytoskeleton.163 In addition, pannexin influences a number of cellular changes that require cytoskeletal plasticity, including migration, differentiation, and proliferation.162,163 A role for actin is emphasized by evidence that Rho-kinase pathways, which have long been implicated in regulating actin dynamics, are dependent on pannexin-channel activity.164 The importance of pannexins in regulating cytoskeletal dynamics has also been suggested by the localization of pannexin to the actin-rich filopodia of directionally migrating or path-finding cells.17 We have shown that loss or inhibition of Panx1 increases neurite extension and branching in sensory neurons ex vivo.165 This finding suggests that pannexins may play a suppressive role in neuronal growth. Finally, pannexin channels may also somehow play a role in the mechanical sensitivity of other mechanosensitive channels, such as TRPV4 and TRPV1.166–168 It is not yet clear how the activity of these channels is coupled to pannexin activity. Such regulation may also be mediated by the cytoskeleton, the rigidity of which likely influences mechanosensitive-channel response and the stability of which may be regulated by pannexin–P2-mediated pathways, such as those already noted. Therefore, mechanosignaling through Panx HCs may play a role in normal cell migration, growth, and differentiation. However, persistent exposure to mechanical stress may facilitate sustained activation of a Panx1–P2X-signaling loop, contributing to chronic inflammation (Figure 4A and B).
Figure 4.
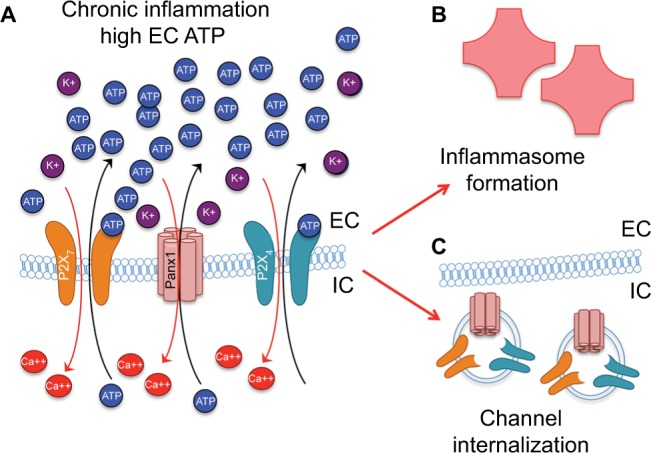
Persistent Panx1 and P2X activation (A) leads to inflammasome formation (B) and channel/receptor plasticity, including channel internalization through endocytosis (C).
Abbreviations: EC, extracellular; IC, intracellular.
Pannexin and receptor plasticity
The magnitude and persistence of an activating signal have been noted to influence the functional plasticity of pannexin and pannexin-associated purinergic receptors (Figure 4A and B). In addition to the low-affinity binding already mentioned, another negative-feedback mechanism response to increased extracellular ATP involves rapid internalization of Panx1 into endosomes (Figure 4C) in as little as 15 minutes, possibly through signaling initiated by P2X receptors.169 Positive feedback mechanisms include coregulation of pannexins with purinergic signaling proteins, detected in experiments with hypertonic saline treatment, which triggered both pannexin-channel activity and expression levels of P2X receptors in Jurkat T cells.95 Similar coregulation was observed in chronic mechanical strain in astrocytes that resulted in an increased expression of Panx1, -2, and -3 both in vitro and in vivo.130
Therapeutic implications of Panx1 inhibition
Pannexin has been implicated in regulating normal and pathological cellular function in a wide range of tissue types. In normal physiology, pannexin has been shown to modulate vascular tone,170 brain development,9 memory, sleep,166,171 skeletal muscle homeostasis,172,173 red blood-cell biomechanics,146 retinal signaling, response to ischemia,117,174 and leukocyte emigration.105 In pathology, pannexin-mediated signaling has been implicated in brain ischemia,64,175 ischemic stroke,176 pain,177,178 cardiomyocyte fibrosis,179 microbial infection,59,180 cancer,181 brain inflammation (autoimmune encephalomyelitis/multiple sclerosis),182–185 and immunogenic cell-death-inducing antineoplastic agents.96,186 On one hand, this potential breadth of functions renders pannexin a powerful and widely applicable therapeutic target. On other hand, this same breadth suggests potentially significant side effects of anti-pannexin therapy, manifested within the same cell type, within the same tissue, or systemically. In addition, as demonstrated by varying results of different commonly used pharmacological inhibitors of pannexin or P2 receptors, including probenecid, Panx1-blocking peptide (10Panx),105 carbenoxolone, P2-receptor-inhibiting peptides, and the extracellular ATP scavenger apyrase, the specificity of inhibition can appreciably impact a phenotypic response. In the following sections, we briefly describe possible outcomes related to the authors’ expertise, in which the diverse functions of Panx1 must be considered within translational therapeutic strategies.
Panx1 inhibition in the eye
Panx1 forms an ATP-, K+-, and Ca2+-permeable membrane channel that is highly expressed in the retina, making this easily accessible neural tissue a good model system for delineation of Panx1 function. In the retina, Panx1 has been shown to be activated by mechanical stress,29,69,111,130 intracellular Ca2+,117 extracellular K+,187 interactions with transient-receptor-potential channels,142,188 N-methyl-D-aspartate receptors,189 activation of C2+-dependent caspases 1/11 and NLRP1/3 inflammasomes,29,117,190,191 and purinergic receptors upon binding extracellular ATP.88 Several of these stressors and agonists are activated in the retina challenged by ischemia, OHT stress, or glaucoma, which can synergize to sustain prolonged Panx1 opening. Consistently with this observation, therapeutic Panx1 blockade protects RGCs and other neurons against mechanical stress and ischemia.29,174 However, due to the physiological significance of Panx1, only transient blockade and partial suppression represent therapeutically feasible options, as they are sufficient to block inflammasome and ionized Ca2+ influx without affecting global retina functionality.
Panx1 inhibition increases lacrimal-gland repair
Recent studies have proposed distinct roles for both Panx1 and P2X7 receptors in the control of inflammasome activation, leading to the release of mature IL1α and IL1β. These data support the model in which Panx1–P2X7 signaling is the key regulator of inflammatory response.52,192 Probenecid, a well-studied inhibitor of Panx1 and P2X7 receptors193,194 and organic anion transporters, has been traditionally used to treat an inflammatory gout disease.195 Treatment with probenecid has been found to affect ATP release195 and suppress neuronal death in ischemic stroke176,196 and cerebral edema.197 This suggests that modulation of Panx1 signaling may prevent inflammatory damage of brain tissue. Another study reports that in vivo administration of the P2X7R antagonist A438079 in the mouse model of salivary gland exocrinopathy could ameliorate salivary gland inflammation and enhance saliva secretion.198
Panx1 and P2 receptors are strongly upregulated during acute and chronic inflammation of the lacrimal gland,18 the primary contributor to the aqueous layer of tear film in humans. Moreover, lacrimal-gland injury due to inflammation leads to aqueous tear-deficiency dry eye. Most current therapies to treat lacrimal-gland disorders suggest topical treatments, including usage of artificial tears and autologous serum eye-drops, but they do not treat the cause of the disease and lead to limited success. Cell-based regenerative therapies may provide better and longer relief to dry-eye patients; however, survival of transplanted cells strictly depends on the degree of inflammation.199,200 We recently have shown that the best cell engraftment is observed when Panx1 has been blocked with specific Panx1 inhibitors, including 10Panx and self-deliverable RNAi (sdRNAi) specific to Panx1.200 Moreover, lacrimal-gland treatment with Panx1 sdRNAi resulted in significant reduction in IL1β and Nlrp3 expression in TSP1−/− mice, a mouse model of aqueous tear-deficiency dry eye.200 These findings have implications for therapeutic strategies targeting Panx1-signaling pathways for suppression of inflammation and/or increasing donor lacrimal-gland progenitor-cell engraftment.
The role of Panx1 in peripheral nerve disease and repair
Pannexins have been implicated in a number of pain-sensitization pathways, in both the peripheral nervous system and the CNS, through activity within and communication between neurons and their supporting cells.99,100,201,202 Zhang et al provided initial evidence that cell bodies of sensory neurons release ATP in response to electrical stimulation. Further, this release stimulated activation of P2X7 receptors and subsequent release of the inflammatory cytokine TNFα in neighboring glial cells.203 Pannexin interactions with various isoforms of P2 receptors may be localization-specific, as P2X4 receptors have also been implicated in purinergic pain pathways: in this case, through activity at sensory endings in the skin.204 Interestingly, P2X3 receptors have also been noted to play a prominent role in pannexin-mediated signaling in DRG neurons.203,205,206 Consistently with a role for pannexin-P2X3 activity in pain pathways, the neurotoxin BomoTx, from the Brazilian lancehead pit viper, activates ATP release through pannexin HCs and downstream P2X3-receptor activation, resulting in inflammatory pain, thermal hyperalgesia, and mechanical allodynia. Further, nerve injury results in increased Panx1 gene and protein expression due to epigenetic mechanisms.178 In contrast, a recent study revealed that Panx1 inhibition, genetically or through pharmacological reagents, reduced hypersensitivity induced by nerve injury.207
Although studied less comprehensively, pannexins also likely play an important role in peripheral nerve development and regeneration. For example, a role in myelination has been suggested for both P2X7 receptors and pannexin,208 likely due to communication between stimulated neurons and their flanking Schwann cells.209 In addition, a recent study by our research team indicated that pannexin negatively regulated developmental and regenerative growth of peripheral neurons, as suggested by the increased caliber of axons in the sciatic nerves of Panx1−/− mice and increased regenerative outgrowth and branching of cultured DRG explants harvested from Panx1−/− mice, as well as in wild-type DRG explants treated with inhibitors of the Panx1-signaling pathway, including apyrase, probenecid, and 10Panx.210 Based on its physiological properties and roles in inflammasome activation, it is feasible to suggest that pannexin activity can modulate the neuroregenerative environment, which is enriched in inflammatory cytokines, as well as immune and activated glial cells that respond to inflammatory cues. As such, pannexin reduction may be an effective strategy to reduce pain and promote regeneration after nerve injury.
Conclusion
The switch between normal (minor) and pathological (massive) ATP release from Panx HCs and downstream P2 receptors and other channels can determine whether an outcome will be “good” or “bad”. Low levels of extracellular ATP and K+ produced by physiological pannexin activity is required for homeostatic cell function (Figure 5A). Conversely, high levels of extracellular ATP and K+ and overload of the intracellular compartment with Ca2+ synergistically lead to sustained activation of Panx1–P2X7 signaling and inflammasome pathways, inducing Casp1/11-dependent pyroptotic cell death (Figure 5B). Therefore, therapeutic modulation of Panx1 channels represents a feasible new strategy to reduce inflammation and promote regeneration (Figure 5C).
Figure 5.
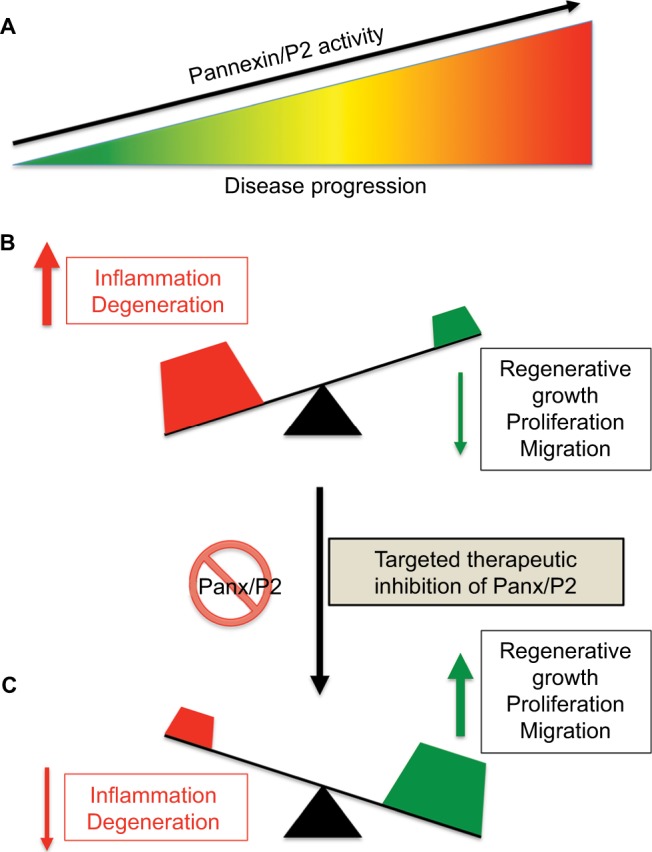
Effect of therapeutic suppression of pannexin function.
Notes: (A) Disease progression often correlates with increased Panx/P2-receptor activity and chronic inflammation (B). We hypothesize that therapeutic suppression of pannexin function may tilt the balance of a progressing disease from that of inflammation and degeneration to one of enhanced cellular growth, proliferation, and/or migration (C).
Acknowledgments
This work was supported by National Institutes of Health National Eye Institute grants 1R01EY026202 (to HPM) and R01EY021517 (to VIS), Russian Science Foundation grant N17-15-01433 (to VIS), and the Department of Veterans Affairs (Merit Award 1 I01 RX001471 to SBS).
Footnotes
Disclosure
The authors report no conflicts of interest in this work.
References
- 1.Phelan P, Bacon JP, Davies JA, et al. Innexins: a family of invertebrate gap-junction proteins. Trends Genet. 1998;14(9):348–349. doi: 10.1016/s0168-9525(98)01547-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Phelan P, Starich TA. Innexins get into the gap. Bioessays. 2001;23(5):388–396. doi: 10.1002/bies.1057. [DOI] [PubMed] [Google Scholar]
- 3.Herve JC, Phelan P, Bruzzone R, White TW. Connexins, innexins and pannexins: bridging the communication gap. Biochim Biophys Acta. 2005;1719(1–2):3–5. doi: 10.1016/j.bbamem.2005.11.013. [DOI] [PubMed] [Google Scholar]
- 4.Makowski L, Caspar DL, Phillips WC, Baker TS, Goodenough DA. Gap junction structures – VI: variation and conservation in connexon conformation and packing. Biophys J. 1984;45(1):208–218. doi: 10.1016/S0006-3495(84)84149-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Sáez JC, Retamal MA, Basilio D, Bukauskas FF, Bennett MV. Connexin-based gap junction hemichannels: gating mechanisms. Biochim Biophys Acta. 2005;1711(2):215–224. doi: 10.1016/j.bbamem.2005.01.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Söhl G, Willecke K. Gap junctions and the connexin protein family. Cardiovasc Res. 2004;62(2):228–232. doi: 10.1016/j.cardiores.2003.11.013. [DOI] [PubMed] [Google Scholar]
- 7.d’Hondt C, Ponsaerts R, de Smedt H, Bultynck G, Himpens B. Pannexins, distant relatives of the connexin family with specific cellular functions? Bioessays. 2009;31(9):953–974. doi: 10.1002/bies.200800236. [DOI] [PubMed] [Google Scholar]
- 8.Barbe MT, Monyer H, Bruzzone R. Cell-cell communication beyond connexins: the pannexin channels. Physiology (Bethesda) 2006;21:103–114. doi: 10.1152/physiol.00048.2005. [DOI] [PubMed] [Google Scholar]
- 9.Bruzzone R, Hormuzdi SG, Barbe MT, Herb A, Monyer H. Pannexins, a family of gap junction proteins expressed in brain. Proc Natl Acad Sci U S A. 2003;100(23):13644–13649. doi: 10.1073/pnas.2233464100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Shestopalov VI, Panchin Y. Pannexins and gap junction protein diversity. Cell Mol Life Sci. 2008;65(3):376–394. doi: 10.1007/s00018-007-7200-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Giaume C, Leybaert L, Naus CC, Sáez JC. Connexin and pannexin hemichannels in brain glial cells: properties, pharmacology, and roles. Front Pharmacol. 2013;4:88. doi: 10.3389/fphar.2013.00088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Rodriguez-Sinovas A, Sanchez JA, Fernandez-Sanz C, Ruiz-Meana M, Garcia-Dorado D. Connexin and pannexin as modulators of myocardial injury. Biochim Biophys Acta. 2012;1818(8):1962–1970. doi: 10.1016/j.bbamem.2011.07.041. [DOI] [PubMed] [Google Scholar]
- 13.Penuela S, Bhalla R, Gong XQ, et al. Pannexin 1 and pannexin 3 are glycoproteins that exhibit many distinct characteristics from the connexin family of gap junction proteins. J Cell Sci. 2007;120(Pt 21):3772–3783. doi: 10.1242/jcs.009514. [DOI] [PubMed] [Google Scholar]
- 14.Baranova A, Ivanov D, Petrash N, et al. The mammalian pannexin family is homologous to the invertebrate innexin gap junction proteins. Genomics. 2004;83(4):706–716. doi: 10.1016/j.ygeno.2003.09.025. [DOI] [PubMed] [Google Scholar]
- 15.Bond SR, Lau A, Penuela S, et al. Pannexin 3 is a novel target for Runx2, expressed by osteoblasts and mature growth plate chondrocytes. J Bone Miner Res. 2011;26(12):2911–2922. doi: 10.1002/jbmr.509. [DOI] [PubMed] [Google Scholar]
- 16.Penuela S, Kelly JJ, Churko JM, Barr KJ, Berger AC, Laird DW. Panx1 regulates cellular properties of keratinocytes and dermal fibroblasts in skin development and wound healing. J Invest Dermatol. 2014;134(7):2026–2035. doi: 10.1038/jid.2014.86. [DOI] [PubMed] [Google Scholar]
- 17.Wicki-Stordeur LE, Swayne LA. Panx1 regulates neural stem and progenitor cell behaviours associated with cytoskeletal dynamics and interacts with multiple cytoskeletal elements. Cell Commun Signal. 2013;11:62. doi: 10.1186/1478-811X-11-62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Basova LV, Tang X, Umasume T, et al. Manipulation of Panx1 Activity increases the engraftment of transplanted lacrimal gland epithelial progenitor cells. Invest Ophthalmol Vis Sci. 2017;58(13):5654–5665. doi: 10.1167/iovs.17-22071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kurtenbach S, Whyte-Fagundes P, Gelis L, et al. Investigation of olfactory function in a Panx1 knock out mouse model. Front Cell Neurosci. 2014;8:266. doi: 10.3389/fncel.2014.00266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lohman AW, Billaud M, Straub AC, et al. Expression of pannexin isoforms in the systemic murine arterial network. J Vasc Res. 2012;49(5):405–416. doi: 10.1159/000338758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.le Vasseur M, Lelowski J, Bechberger JF, Sin WC, Naus CC. Pannexin 2 protein expression is not restricted to the CNS. Front Cell Neurosci. 2014;8:392. doi: 10.3389/fncel.2014.00392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Swayne LA, Sorbara CD, Bennett SA. Pannexin 2 is expressed by postnatal hippocampal neural progenitors and modulates neuronal commitment. J Biol Chem. 2010;285(32):24977–24986. doi: 10.1074/jbc.M110.130054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zhang H, Chen Y, Zhang C. Patterns of heterogeneous expression of pannexin 1 and pannexin 2 transcripts in the olfactory epithelium and olfactory bulb. J Mol Histol. 2012;43(6):651–660. doi: 10.1007/s10735-012-9443-x. [DOI] [PubMed] [Google Scholar]
- 24.Bhalla-Gehi R, Penuela S, Churko JM, Shao Q, Laird DW. Pannexin1 and pannexin3 delivery, cell surface dynamics, and cytoskeletal interactions. J Biol Chem. 2010;285(12):9147–9160. doi: 10.1074/jbc.M109.082008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Langlois S, Xiang X, Young K, Cowan BJ, Penuela S, Cowan KN. Pannexin 1 and pannexin 3 channels regulate skeletal muscle myoblast proliferation and differentiation. J Biol Chem. 2014;289(44):30717–30731. doi: 10.1074/jbc.M114.572131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Penuela S, Bhalla R, Nag K, Laird DW. Glycosylation regulates pannexin intermixing and cellular localization. Mol Biol Cell. 2009;20(20):4313–4323. doi: 10.1091/mbc.E09-01-0067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Abeele FV, Bidaux G, Gordienko D, et al. Functional implications of calcium permeability of the channel formed by pannexin 1. J Cell Biol. 2006;174(4):535–546. doi: 10.1083/jcb.200601115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Makarenkova HP, Shestopalov VI. The role of pannexin hemichannels in inflammation and regeneration. Front Physiol. 2014;5:63. doi: 10.3389/fphys.2014.00063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Križaj D, Ryskamp DA, Tian N, et al. From mechanosensitivity to inflammatory responses: new players in the pathology of glaucoma. Curr Eye Res. 2014;39(2):105–119. doi: 10.3109/02713683.2013.836541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Retamal MA, Riquelme MA, Stehberg J, Alcayaga J. Connexin43 hemichannels in satellite glial cells, can they influence sensory neuron activity? Front Mol Neurosci. 2017;10:374. doi: 10.3389/fnmol.2017.00374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lapato AS, Tiwari-Woodruff SK. Connexins and pannexins: at the junction of neuro-glial homeostasis and disease. J Neurosci Res. 2018;96(1):31–44. doi: 10.1002/jnr.24088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sarrouilhe D, Dejean C, Mesnil M. Connexin43- and pannexin-based channels in neuroinflammation and cerebral neuropathies. Front Mol Neurosci. 2017;10:320. doi: 10.3389/fnmol.2017.00320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lohman AW, Isakson BE. Differentiating connexin hemichannels and pannexin channels in cellular ATP release. FEBS Lett. 2014;588(8):1379–1388. doi: 10.1016/j.febslet.2014.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Gulbransen BD, Bashashati M, Hirota SA, et al. Activation of neuronal P2X7 receptor-pannexin-1 mediates death of enteric neurons during colitis. Nat Med. 2012;18(4):600–604. doi: 10.1038/nm.2679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Pelegrin P. Targeting interleukin-1 signaling in chronic inflammation: focus on P2X7 receptor and pannexin-1. Drug News Perspect. 2008;21(8):424–433. doi: 10.1358/dnp.2008.21.8.1265800. [DOI] [PubMed] [Google Scholar]
- 36.Kanneganti TD, Lamkanfi M, Kim YG, et al. Pannexin-1-mediated recognition of bacterial molecules activates the cryopyrin inflammasome independent of Toll-like receptor signaling. Immunity. 2007;26(4):433–443. doi: 10.1016/j.immuni.2007.03.008. [DOI] [PubMed] [Google Scholar]
- 37.Martinon F, Tschopp J. Inflammatory caspases and inflammasomes: master switches of inflammation. Cell Death Differ. 2007;14(1):10–22. doi: 10.1038/sj.cdd.4402038. [DOI] [PubMed] [Google Scholar]
- 38.Martinon F, Burns K, Tschopp J. The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-β. Mol Cell. 2002;10(2):417–426. doi: 10.1016/s1097-2765(02)00599-3. [DOI] [PubMed] [Google Scholar]
- 39.Vaccari JP, Lotocki G, Marcillo AE, Dietrich WD, Keane RW. A molecular platform in neurons regulates inflammation after spinal cord injury. J Neurosci. 2008;28(13):3404–3414. doi: 10.1523/JNEUROSCI.0157-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kesavardhana S, Kanneganti TD. Mechanisms governing inflammasome activation, assembly and pyroptosis induction. Int Immunol. 2017;29(5):201–210. doi: 10.1093/intimm/dxx018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Malik A, Kanneganti TD. Inflammasome activation and assembly at a glance. J Cell Sci. 2017;130(23):3955–3963. doi: 10.1242/jcs.207365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Lei X, Zhang Z, Xiao X, Qi J, He B, Wang J. Enterovirus 71 inhibits pyroptosis through cleavage of gasdermin D. J Virol. 2017;91(18):e01069–17. doi: 10.1128/JVI.01069-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Chen X, He WT, Hu L, et al. Pyroptosis is driven by non-selective gasdermin-D pore and its morphology is different from MLKL channel-mediated necroptosis. Cell Res. 2016;26(9):1007–1020. doi: 10.1038/cr.2016.100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.He WT, Wan H, Hu L, et al. Gasdermin D is an executor of pyroptosis and required for interleukin-1β secretion. Cell Res. 2015;25(12):1285–1298. doi: 10.1038/cr.2015.139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kayagaki N, Stowe IB, Lee BL, et al. Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature. 2015;526(7575):666–671. doi: 10.1038/nature15541. [DOI] [PubMed] [Google Scholar]
- 46.Man SM, Kanneganti TD. Gasdermin D: the long-awaited executioner of pyroptosis. Cell Res. 2015;25(11):1183–1184. doi: 10.1038/cr.2015.124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Yue N, Huang H, Zhu X, et al. Activation of P2X7 receptor and NLRP3 inflammasome assembly in hippocampal glial cells mediates chronic stress-induced depressive-like behaviors. J Neuroinflammation. 2017;14(1):102. doi: 10.1186/s12974-017-0865-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Vaccari JP, Dietrich WD, Keane RW. Activation and regulation of cellular inflammasomes: gaps in our knowledge for central nervous system injury. J Cereb Blood Flow Metab. 2014;34(3):369–375. doi: 10.1038/jcbfm.2013.227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Silverman WR, Vaccari JP, Locovei S, et al. The pannexin 1 channel activates the inflammasome in neurons and astrocytes. J Biol Chem. 2009;284(27):18143–18151. doi: 10.1074/jbc.M109.004804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Riteau N, Gasse P, Fauconnier L, et al. Extracellular ATP is a danger signal activating P2X7 receptor in lung inflammation and fibrosis. Am J Respir Crit Care Med. 2010;182(6):774–783. doi: 10.1164/rccm.201003-0359OC. [DOI] [PubMed] [Google Scholar]
- 51.Hung SC, Choi CH, Said-Sadier N, et al. P2X4 assembles with P2X7 and pannexin-1 in gingival epithelial cells and modulates ATP-induced reactive oxygen species production and inflammasome activation. PLoS One. 2013;8(7):e70210. doi: 10.1371/journal.pone.0070210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Pelegrin P, Surprenant A. Pannexin-1 couples to maitotoxin- and nigericin-induced interleukin-1β release through a dye uptake-independent pathway. J Biol Chem. 2007;282(4):2386–2394. doi: 10.1074/jbc.M610351200. [DOI] [PubMed] [Google Scholar]
- 53.Pelegrin P, Surprenant A. Pannexin-1 mediates large pore formation and interleukin-1β release by the ATP-gated P2X7 receptor. EMBO J. 2006;25(21):5071–5082. doi: 10.1038/sj.emboj.7601378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Albalawi F, Lu W, Beckel JM, Lim JC, McCaughey SA, Mitchell CH. The P2X7 receptor primes IL-1β and the NLRP3 Inflammasome in astrocytes exposed to mechanical strain. Front Cell Neurosci. 2017;11:227. doi: 10.3389/fncel.2017.00227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Qu Y, Misaghi S, Newton K, et al. Pannexin-1 is required for ATP release during apoptosis but not for inflammasome activation. J Immunol. 2011;186(11):6553–6561. doi: 10.4049/jimmunol.1100478. [DOI] [PubMed] [Google Scholar]
- 56.Takahashi M. NLRP3 in myocardial ischaemia-reperfusion injury: inflammasome-dependent or -independent role in different cell types. Cardiovasc Res. 2013;99(1):4–5. doi: 10.1093/cvr/cvt142. [DOI] [PubMed] [Google Scholar]
- 57.Yanguas SC, Willebrords J, Johnstone SR, et al. Pannexin1 as mediator of inflammation and cell death. Biochim Biophys Acta. 2017;1864(1):51–61. doi: 10.1016/j.bbamcr.2016.10.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Brough D, Pelegrin P, Rothwell NJ. Pannexin-1-dependent caspase-1 activation and secretion of IL-1β is regulated by zinc. Eur J Immunol. 2009;39(2):352–358. doi: 10.1002/eji.200838843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.McKuen MJ, Dahl G, Fields KA. Assessing a potential role of host pannexin 1 during Chlamydia trachomatis infection. PLoS One. 2013;8(5):e63732. doi: 10.1371/journal.pone.0063732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Liu M, Wu Q, Wang M, Fu Y, Wang J. Lactobacillus rhamnosus GR-1 limits Escherichia coli-induced inflammatory responses via attenuating MyD88-dependent and MyD88-independent pathway activation in bovine endometrial epithelial cells. Inflammation. 2016;39(4):1483–1494. doi: 10.1007/s10753-016-0382-7. [DOI] [PubMed] [Google Scholar]
- 61.Correa-Costa M, Braga TT, Semedo P, et al. Pivotal role of Toll-like receptors 2 and 4, its adaptor molecule MyD88, and inflammasome complex in experimental tubule-interstitial nephritis. PLoS One. 2011;6(12):e29004. doi: 10.1371/journal.pone.0029004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Wang H, Xing Y, Mao L, Luo Y, Kang L, Meng G. Pannexin-1 influences peritoneal cavity cell population but is not involved in NLRP3 inflammasome activation. Protein Cell. 2013;4(4):259–265. doi: 10.1007/s13238-013-2114-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Parzych K, Zetterqvist AV, Wright WR, Kirkby NS, Mitchell JA, Paul-Clark MJ. Differential role of pannexin-1/ATP/P2X7 axis in IL-1β release by human monocytes. FASEB J. 2017;31(6):2439–2445. doi: 10.1096/fj.201600256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Weilinger NL, Lohman AW, Rakai BD, et al. Metabotropic NMDA receptor signaling couples Src family kinases to pannexin-1 during excitotoxicity. Nat Neurosci. 2016;19(3):432–442. doi: 10.1038/nn.4236. [DOI] [PubMed] [Google Scholar]
- 65.Chekeni FB, Elliott MR, Sandilos JK, et al. Pannexin 1 channels mediate ‘find-me’ signal release and membrane permeability during apoptosis. Nature. 2010;467(7317):863–867. doi: 10.1038/nature09413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Sridharan M, Adderley SP, Bowles EA, et al. Pannexin 1 is the conduit for low oxygen tension-induced ATP release from human erythrocytes. Am J Physiol Heart Circ Physiol. 2010;299(4):H1146–H1152. doi: 10.1152/ajpheart.00301.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Scemes E, Spray DC. Extracellular K+ and astrocyte signaling via connexin and pannexin channels. Neurochem Res. 2012;37(11):2310–2316. doi: 10.1007/s11064-012-0759-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Sandilos JK, Bayliss DA. Physiological mechanisms for the modulation of pannexin 1 channel activity. J Physiol. 2012;590(Pt 24):6257–6266. doi: 10.1113/jphysiol.2012.240911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Xia J, Lim JC, Lu W, et al. Neurons respond directly to mechanical deformation with pannexin-mediated ATP release and autostimulation of P2X7 receptors. J Physiol. 2012;590(10):2285–2304. doi: 10.1113/jphysiol.2012.227983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Penuela S, Simek J, Thompson RJ. Regulation of pannexin channels by post-translational modifications. FEBS Lett. 2014;588(8):1411–1415. doi: 10.1016/j.febslet.2014.01.028. [DOI] [PubMed] [Google Scholar]
- 71.Chiu YH, Jin X, Medina CB, et al. A quantized mechanism for activation of pannexin channels. Nat Commun. 2017;8:14324. doi: 10.1038/ncomms14324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Sandilos JK, Chiu YH, Chekeni FB, et al. Pannexin 1, an ATP release channel, is activated by caspase cleavage of its pore-associated C-terminal autoinhibitory region. J Biol Chem. 2012;287(14):11303–11311. doi: 10.1074/jbc.M111.323378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Yang D, He Y, Munoz-Planillo R, Liu Q, Nunez G. Caspase-11 requires the pannexin-1 channel and the purinergic P2X7 pore to mediate pyroptosis and endotoxic shock. Immunity. 2015;43(5):923–932. doi: 10.1016/j.immuni.2015.10.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Billaud M, Chiu YH, Lohman AW, et al. A molecular signature in the pannexin1 intracellular loop confers channel activation by the α1 adrenoreceptor in smooth muscle cells. Sci Signal. 2015;8(364):ra17. doi: 10.1126/scisignal.2005824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Martinello T, Baldoin MC, Morbiato L, et al. Extracellular ATP signaling during differentiation of C2C12 skeletal muscle cells: role in proliferation. Mol Cell Biochem. 2011;351(1–2):183–196. doi: 10.1007/s11010-011-0726-4. [DOI] [PubMed] [Google Scholar]
- 76.Meyer MP, Gröschel-Stewart U, Robson T, Burnstock G. Expression of two ATP-gated ion channels, P2X5 and P2X6, in developing chick skeletal muscle. Dev Dyn. 1999;216(4–5):442–449. doi: 10.1002/(SICI)1097-0177(199912)216:4/5<442::AID-DVDY12>3.0.CO;2-Z. [DOI] [PubMed] [Google Scholar]
- 77.Riquelme MA, Cea LA, Vega JL, et al. The ATP required for potentiation of skeletal muscle contraction is released via pannexin hemichannels. Neuropharmacology. 2013;75:594–603. doi: 10.1016/j.neuropharm.2013.03.022. [DOI] [PubMed] [Google Scholar]
- 78.Buvinic S, Almarza G, Bustamante M, et al. ATP released by electrical stimuli elicits calcium transients and gene expression in skeletal muscle. J Biol Chem. 2009;284(50):34490–34505. doi: 10.1074/jbc.M109.057315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Messemer N, Kunert C, Grohmann M, et al. P2X7 receptors at adult neural progenitor cells of the mouse subventricular zone. Neuropharmacology. 2013;73:122–137. doi: 10.1016/j.neuropharm.2013.05.017. [DOI] [PubMed] [Google Scholar]
- 80.Jacques-Silva MC, Rodnight R, Lenz G, et al. P2X7 receptors stimulate Akt phosphorylation in astrocytes. Br J Pharmacol. 2004;141(7):1106–1117. doi: 10.1038/sj.bjp.0705685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Plesner L. Ecto-ATPases: identities and functions. Int Rev Cytol. 1995;158:141–214. doi: 10.1016/s0074-7696(08)62487-0. [DOI] [PubMed] [Google Scholar]
- 82.Jackson DG, Wang J, Keane RW, Scemes E, Dahl G. ATP and potassium ions: a deadly combination for astrocytes. Sci Rep. 2014;4:4576. doi: 10.1038/srep04576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Qiu F, Dahl G. A permeant regulating its permeation pore: inhibition of pannexin 1 channels by ATP. Am J Physiol Cell Physiol. 2009;296(2):C250–C255. doi: 10.1152/ajpcell.00433.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.McIlwain DR, Berger T, Mak TW. Caspase functions in cell death and disease. Cold Spring Harb Perspect Biol. 2013;5(4):a008656. doi: 10.1101/cshperspect.a008656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Wilkaniec A, Gassowska M, Czapski GA, Cieslik M, Sulkowski G, Adamczyk A. P2X7 receptor-pannexin 1 interaction mediates extracellular alpha-synuclein-induced ATP release in neuroblastoma SH-SY5Y cells. Purinergic Signal. 2017;13(3):347–361. doi: 10.1007/s11302-017-9567-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Locovei S, Wang J, Dahl G. Activation of pannexin 1 channels by ATP through P2Y receptors and by cytoplasmic calcium. FEBS Lett. 2006;580(1):239–244. doi: 10.1016/j.febslet.2005.12.004. [DOI] [PubMed] [Google Scholar]
- 87.Iglesias R, Locovei S, Roque A, et al. P2X7 receptor-pannexin1 complex: pharmacology and signaling. Am J Physiol Cell Physiol. 2008;295(3):C752–C760. doi: 10.1152/ajpcell.00228.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Locovei S, Scemes E, Qiu F, Spray DC, Dahl G. Pannexin1 is part of the pore forming unit of the P2X7 receptor death complex. FEBS Lett. 2007;581(3):483–488. doi: 10.1016/j.febslet.2006.12.056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Roberts JA, Vial C, Digby HR, et al. Molecular properties of P2X receptors. Pflugers Arch. 2006;452(5):486–500. doi: 10.1007/s00424-006-0073-6. [DOI] [PubMed] [Google Scholar]
- 90.Kaczmarek-Hajek K, Lorinczi E, Hausmann R, Nicke A. Molecular and functional properties of P2X receptors: recent progress and persisting challenges. Purinergic Signal. 2012;8(3):375–417. doi: 10.1007/s11302-012-9314-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.North RA. Molecular physiology of P2X receptors. Physiol Rev. 2002;82(4):1013–1067. doi: 10.1152/physrev.00015.2002. [DOI] [PubMed] [Google Scholar]
- 92.Chen L, Liu YW, Yue K, et al. Differential expression of ATP-gated P2X receptors in DRG between chronic neuropathic pain and visceralgia rat models. Purinergic Signal. 2016;12(1):79–87. doi: 10.1007/s11302-015-9481-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Li S, Tomić M, Stojilkovic SS. Characterization of novel pannexin 1 isoforms from rat pituitary cells and their association with ATP-gated P2X channels. Gen Comp Endocrinol. 2011;174(2):202–210. doi: 10.1016/j.ygcen.2011.08.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Woehrle T, Yip L, Elkhal A, et al. Pannexin-1 hemichannel-mediated ATP release together with P2X1 and P2X4 receptors regulate T-cell activation at the immune synapse. Blood. 2010;116(18):3475–3484. doi: 10.1182/blood-2010-04-277707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Woehrle T, Yip L, Manohar M, et al. Hypertonic stress regulates T cell function via pannexin-1 hemichannels and P2X receptors. J Leukoc Biol. 2010;88(6):1181–1189. doi: 10.1189/jlb.0410211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Draganov D, Gopalakrishna-Pillai S, Chen YR, et al. Modulation of P2X4/P2X7/pannexin-1 sensitivity to extracellular ATP via Ivermectin induces a non-apoptotic and inflammatory form of cancer cell death. Sci Rep. 2015;5:16222. doi: 10.1038/srep16222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Khakh BS, Proctor WR, Dunwiddie TV, Labarca C, Lester HA. Allosteric control of gating and kinetics at P2X4 receptor channels. J Neurosci. 1999;19(17):7289–7299. doi: 10.1523/JNEUROSCI.19-17-07289.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Khakh BS, Bao XR, Labarca C, Lester HA. Neuronal P2X transmitter-gated cation channels change their ion selectivity in seconds. Nat Neurosci. 1999;2(4):322–330. doi: 10.1038/7233. [DOI] [PubMed] [Google Scholar]
- 99.Bravo D, Maturana CJ, Pelissier T, Hernandez A, Constandil L. Interactions of pannexin 1 with NMDA and P2X7 receptors in central nervous system pathologies: possible role on chronic pain. Pharmacol Res. 2015;101:86–93. doi: 10.1016/j.phrs.2015.07.016. [DOI] [PubMed] [Google Scholar]
- 100.Bravo D, Ibarra P, Retamal J, et al. Pannexin 1: a novel participant in neuropathic pain signaling in the rat spinal cord. Pain. 2014;155(10):2108–2115. doi: 10.1016/j.pain.2014.07.024. [DOI] [PubMed] [Google Scholar]
- 101.Lommen J, Stahr A, Ingenwerth M, Ali AA, von Gall C. Time-of-day-dependent expression of purinergic receptors in mouse suprachiasmatic nucleus. Cell Tissue Res. 2017;369(3):579–590. doi: 10.1007/s00441-017-2634-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Kashfi S, Peymani M, Ghaedi K, Baharvand H, Nasr-Esfahani MH, Javan M. Purinergic receptor expression and potential association with human embryonic stem cell-derived oligodendrocyte progenitor cell development. Cell J. 2017;19(3):386–402. doi: 10.22074/cellj.2017.3906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Jacobson KA, Ivanov AA, de Castro S, Harden TK, Ko H. Development of selective agonists and antagonists of P2Y receptors. Purinergic Signal. 2009;5(1):75–89. doi: 10.1007/s11302-008-9106-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Burnstock G. Purine and pyrimidine receptors. Cell Mol Life Sci. 2007;64(12):1471–1483. doi: 10.1007/s00018-007-6497-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Lohman AW, Leskov IL, Butcher JT, et al. Pannexin 1 channels regulate leukocyte emigration through the venous endothelium during acute inflammation. Nat Commun. 2015;6:7965. doi: 10.1038/ncomms8965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Sanderson J, Dartt DA, Trinkaus-Randall V, et al. Purines in the eye: recent evidence for the physiological and pathological role of purines in the RPE, retinal neurons, astrocytes, Müller cells, lens, trabecular meshwork, cornea and lacrimal gland. Exp Eye Res. 2014;127:270–279. doi: 10.1016/j.exer.2014.08.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Bao Y, Chen Y, Ledderose C, Li L, Junger WG. Pannexin 1 channels link chemoattractant receptor signaling to local excitation and global inhibition responses at the front and back of polarized neutrophils. J Biol Chem. 2013;288(31):22650–22657. doi: 10.1074/jbc.M113.476283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Chen Y, Corriden R, Inoue Y, et al. ATP release guides neutrophil chemotaxis via P2Y2 and A3 receptors. Science. 2006;314(5806):1792–1795. doi: 10.1126/science.1132559. [DOI] [PubMed] [Google Scholar]
- 109.Young MT, Pelegrin P, Surprenant A. Amino acid residues in the P2X7 receptor that mediate differential sensitivity to ATP and BzATP. Mol Pharmacol. 2007;71(1):92–100. doi: 10.1124/mol.106.030163. [DOI] [PubMed] [Google Scholar]
- 110.Vaccari JP, Bastien D, Yurcisin G, et al. P2X4 receptors influence inflammasome activation after spinal cord injury. J Neurosci. 2012;32(9):3058–3066. doi: 10.1523/JNEUROSCI.4930-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Reigada D, Lu W, Zhang M, Mitchell CH. Elevated pressure triggers a physiological release of ATP from the retina: possible role for pannexin hemichannels. Neuroscience. 2008;157(2):396–404. doi: 10.1016/j.neuroscience.2008.08.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Xia J, Lim JC, Lu W, et al. Neurons respond directly to mechanical deformation with pannexin-mediated ATP release and autostimulation of P2X7 receptors. J Physiol. 2012;590(Pt 10):2285–2304. doi: 10.1113/jphysiol.2012.227983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Boumechache M, Masin M, Edwardson JM, Gorecki DC, Murrell-Lagnado R. Analysis of assembly and trafficking of native P2X4 and P2X7 receptor complexes in rodent immune cells. J Biol Chem. 2009;284(20):13446–13454. doi: 10.1074/jbc.M901255200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Craigie E, Birch RE, Unwin RJ, Wildman SS. The relationship between P2X4 and P2X7: a physiologically important interaction? Front Physiol. 2013;4:216. doi: 10.3389/fphys.2013.00216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Casas-Pruneda G, Reyes JP, Perez-Flores G, Perez-Cornejo P, Arreola J. Functional interactions between P2X4 and P2X7 receptors from mouse salivary epithelia. J Physiol. 2009;587(Pt 12):2887–2901. doi: 10.1113/jphysiol.2008.167395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Kawano A, Tsukimoto M, Noguchi T, et al. Involvement of P2X4 receptor in P2X7 receptor-dependent cell death of mouse macrophages. Biochem Biophys Res Commun. 2012;419(2):374–380. doi: 10.1016/j.bbrc.2012.01.156. [DOI] [PubMed] [Google Scholar]
- 117.Dvoriantchikova G, Ivanov D, Barakat D, et al. Genetic ablation of pannexin1 protects retinal neurons from ischemic injury. PLoS One. 2012;7(2):e31991. doi: 10.1371/journal.pone.0031991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Mitchell CH, Lu W, Hu H, Zhang X, Reigada D, Zhang M. The P2X7 receptor in retinal ganglion cells: a neuronal model of pressure-induced damage and protection by a shifting purinergic balance. Purinergic Signal. 2009;5(2):241–249. doi: 10.1007/s11302-009-9142-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Zhang X, Zhang M, Laties AM, Mitchell CH. Stimulation of P2X7 receptors elevates Ca2+ and kills retinal ganglion cells. Invest Ophthalmol Vis Sci. 2005;46(6):2183–2191. doi: 10.1167/iovs.05-0052. [DOI] [PubMed] [Google Scholar]
- 120.Kakurai K, Sugiyama T, Kurimoto T, Oku H, Ikeda T. Involvement of P2X7 receptors in retinal ganglion cell death after optic nerve crush injury in rats. Neurosci Lett. 2013;534:237–241. doi: 10.1016/j.neulet.2012.11.060. [DOI] [PubMed] [Google Scholar]
- 121.Domercq M, Perez-Samartin A, Aparicio D, Alberdi E, Pampliega O, Matute C. P2X7 receptors mediate ischemic damage to oligodendrocytes. Glia. 2010;(6):730–740. doi: 10.1002/glia.20958. [DOI] [PubMed] [Google Scholar]
- 122.Shoji KF, Saez PJ, Harcha PA, Aguila HL, Sáez JC. Pannexin1 channels act downstream of P2X 7 receptors in ATP-induced murine T-cell death. Channels (Austin) 2014;8(2):142–156. doi: 10.4161/chan.28122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Chen K, Zhang J, Zhang W, et al. ATP-P2X4 signaling mediates NLRP3 inflammasome activation: a novel pathway of diabetic nephropathy. Int J Biochem Cell Biol. 2013;45(5):932–943. doi: 10.1016/j.biocel.2013.02.009. [DOI] [PubMed] [Google Scholar]
- 124.Shen J, Choy DF, Yoshida T, et al. Interleukin-18 has antipermeability and antiangiogenic activities in the eye: reciprocal suppression with VEGF. J Cell Physiol. 2014;229(8):974–983. doi: 10.1002/jcp.24575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Doyle SL, Lopez FJ, Celkova L, et al. IL-18 immunotherapy for neovascular AMD: tolerability and efficacy in nonhuman primates. Invest Ophthalmol Vis Sci. 2015;56(9):5424–5430. doi: 10.1167/iovs.15-17264. [DOI] [PubMed] [Google Scholar]
- 126.Murphy AJ, Kraakman MJ, Kammoun HL, et al. IL-18 production from the NLRP1 inflammasome prevents obesity and metabolic syndrome. Cell Metab. 2016;23(1):155–164. doi: 10.1016/j.cmet.2015.09.024. [DOI] [PubMed] [Google Scholar]
- 127.Sarojini H, Billeter AT, Eichenberger S, et al. Rapid tissue regeneration induced by intracellular ATP delivery: a preliminary mechanistic study. PLoS One. 2017;12(4):e0174899. doi: 10.1371/journal.pone.0174899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Lee SR, Guo SZ, Scannevin RH, et al. Induction of matrix metalloproteinase, cytokines and chemokines in rat cortical astrocytes exposed to plasminogen activators. Neurosci Lett. 2007;417(1):1–5. doi: 10.1016/j.neulet.2007.01.017. [DOI] [PubMed] [Google Scholar]
- 129.Adamson SE, Leitinger N. The role of pannexin1 in the induction and resolution of inflammation. FEBS Lett. 2014;588(8):1416–1422. doi: 10.1016/j.febslet.2014.03.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Beckel JM, Argall AJ, Lim JC, et al. Mechanosensitive release of adenosine 5′-triphosphate through pannexin channels and mechanosensitive upregulation of pannexin channels in optic nerve head astrocytes: a mechanism for purinergic involvement in chronic strain. Glia. 2014;62(9):1486–1501. doi: 10.1002/glia.22695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Sakaki H, Tsukimoto M, Harada H, Moriyama Y, Kojima S. Autocrine regulation of macrophage activation via exocytosis of ATP and activation of P2Y11 receptor. PLoS One. 2013;8(4):e59778. doi: 10.1371/journal.pone.0059778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Yaginuma H, Kawai S, Tabata KV, et al. Diversity in ATP concentrations in a single bacterial cell population revealed by quantitative single-cell imaging. Sci Rep. 2014;4:6522. doi: 10.1038/srep06522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Atarashi K, Nishimura J, Shima T, et al. ATP drives lamina propria TH17 cell differentiation. Nature. 2008;455(7214):808–812. doi: 10.1038/nature07240. [DOI] [PubMed] [Google Scholar]
- 134.Seike S, Takehara M, Kobayashi K, Nagahama M. Role of pannexin 1 in Clostridium perfringens beta-toxin-caused cell death. Biochim Biophys Acta. 2016;1858(12):3150–3156. doi: 10.1016/j.bbamem.2016.10.003. [DOI] [PubMed] [Google Scholar]
- 135.Sáez JC, Nairn AC, Czernik AJ, et al. Phosphorylation of connexin 32, a hepatocyte gap-junction protein, by cAMP-dependent protein kinase, protein kinase C and Ca2+/calmodulin-dependent protein kinase II. Eur J Biochem. 1990;192(2):263–273. doi: 10.1111/j.1432-1033.1990.tb19223.x. [DOI] [PubMed] [Google Scholar]
- 136.Carrer A, Leparulo A, Crispino G, et al. Cx32 hemichannel opening by cytosolic Ca2+ is inhibited by the R220X mutation that causes Charcot-Marie-Tooth disease. Hum Mol Genet. 2018;27(1):80–94. doi: 10.1093/hmg/ddx386. [DOI] [PubMed] [Google Scholar]
- 137.Dhein S, Duerrschmidt N, Scholl A, et al. A new role for extracellular Ca2+ in gap-junction remodeling: studies in humans and rats. Naunyn Schmiedebergs Arch Pharmacol. 2008;377(2):125–138. doi: 10.1007/s00210-008-0265-z. [DOI] [PubMed] [Google Scholar]
- 138.Ebihara L. New roles for connexons. News Physiol Sci. 2003;18:100–103. doi: 10.1152/nips.01431.2002. [DOI] [PubMed] [Google Scholar]
- 139.Bruzzone R, Barbe MT, Jakob NJ, Monyer H. Pharmacological properties of homomeric and heteromeric pannexin hemichannels expressed in Xenopus oocytes. J Neurochem. 2005;92(5):1033–1043. doi: 10.1111/j.1471-4159.2004.02947.x. [DOI] [PubMed] [Google Scholar]
- 140.Bao L, Locovei S, Dahl G. Pannexin membrane channels are mechanosensitive conduits for ATP. FEBS Lett. 2004;572(1–3):65–68. doi: 10.1016/j.febslet.2004.07.009. [DOI] [PubMed] [Google Scholar]
- 141.Yip L, Cheung CW, Corriden R, Chen Y, Insel PA, Junger WG. Hypertonic stress regulates T-cell function by the opposing actions of extracellular adenosine triphosphate and adenosine. Shock. 2007;27(3):242–250. doi: 10.1097/01.shk.0000245014.96419.3a. [DOI] [PubMed] [Google Scholar]
- 142.Shahidullah M, Mandal A, Beimgraben C, Delamere NA. Hyposmotic stress causes ATP release and stimulates Na, K-ATPase activity in porcine lens. J Cell Physiol. 2012;227(4):1428–1437. doi: 10.1002/jcp.22858. [DOI] [PubMed] [Google Scholar]
- 143.Islam MR, Uramoto H, Okada T, Sabirov RZ, Okada Y. Maxi-anion channel and pannexin 1 hemichannel constitute separate pathways for swelling-induced ATP release in murine L929 fibrosarcoma cells. Am J Physiol Cell Physiol. 2012;303(9):C924–C935. doi: 10.1152/ajpcell.00459.2011. [DOI] [PubMed] [Google Scholar]
- 144.Negoro H, Urban-Maldonado M, Liou LS, Spray DC, Thi MM, Suadicani SO. Pannexin 1 channels play essential roles in urothelial mechanotransduction and intercellular signaling. PLoS One. 2014;9(8):e106269. doi: 10.1371/journal.pone.0106269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Seref-Ferlengez Z, Maung S, Schaffler MB, Spray DC, Suadicani SO, Thi MM. P2X7R-Panx1 complex impairs bone mechanosignaling under high glucose levels associated with type-1 diabetes. PLoS One. 2016;11(5):e0155107. doi: 10.1371/journal.pone.0155107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Forsyth AM, Wan J, Owrutsky PD, Abkarian M, Stone HA. Multiscale approach to link red blood cell dynamics, shear viscosity, and ATP release. Proc Natl Acad Sci U S A. 2011;108(27):10986–10991. doi: 10.1073/pnas.1101315108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Burnstock G, Knight GE. Cell culture: complications due to mechanical release of ATP and activation of purinoceptors. Cell Tissue Res. 2017;370(1):1–11. doi: 10.1007/s00441-017-2618-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Reyes JP, Hernandez-Carballo CY, Perez-Flores G, Perez-Cornejo P, Arreola J. Lack of coupling between membrane stretching and pannexin-1 hemichannels. Biochem Biophys Res Commun. 2009;380(1):50–53. doi: 10.1016/j.bbrc.2009.01.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Li A, Zhang X, Zheng D, Ge J, Laties AM, Mitchell CH. Sustained elevation of extracellular ATP in aqueous humor from humans with primary chronic angle-closure glaucoma. Exp Eye Res. 2011;93(4):528–533. doi: 10.1016/j.exer.2011.06.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Cone FE, Gelman SE, Son JL, Pease ME, Quigley HA. Differential susceptibility to experimental glaucoma among 3 mouse strains using bead and viscoelastic injection. Exp Eye Res. 2010;91(3):415–424. doi: 10.1016/j.exer.2010.06.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Resta V, Novelli E, Vozzi G, et al. Acute retinal ganglion cell injury caused by intraocular pressure spikes is mediated by endogenous extracellular ATP. Eur J Neurosci. 2007;25(9):2741–2754. doi: 10.1111/j.1460-9568.2007.05528.x. [DOI] [PubMed] [Google Scholar]
- 152.Lu W, Hu H, Sevigny J, et al. Rat, mouse, and primate models of chronic glaucoma show sustained elevation of extracellular ATP and altered purinergic signaling in the posterior eye. Invest Ophthalmol Vis Sci. 2015;56(5):3075–3083. doi: 10.1167/iovs.14-15891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Lim JC, Lu W, Beckel JM, Mitchell CH. Neuronal release of cytokine IL-3 triggered by mechanosensitive autostimulation of the P2X7 receptor is neuroprotective. Front Cell Neurosci. 2016;10:270. doi: 10.3389/fncel.2016.00270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Campbell M, Doyle SL, Ozaki E, et al. An overview of the involvement of interleukin-18 in degenerative retinopathies. Adv Exp Med Biol. 2014;801:409–415. doi: 10.1007/978-1-4614-3209-8_52. [DOI] [PubMed] [Google Scholar]
- 155.Iglesias R, Dahl G, Qiu F, Spray DC, Scemes E. Pannexin 1: the molecular substrate of astrocyte “hemichannels”. J Neurosci. 2009;29(21):7092–7097. doi: 10.1523/JNEUROSCI.6062-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Kang J, Kang N, Lovatt D, et al. Connexin 43 hemichannels are permeable to ATP. J Neurosci. 2008;28(18):4702–4711. doi: 10.1523/JNEUROSCI.5048-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Suadicani SO, Brosnan CF, Scemes E. P2X7 receptors mediate ATP release and amplification of astrocytic intercellular Ca2+ signaling. J Neurosci. 2006;26(5):1378–1385. doi: 10.1523/JNEUROSCI.3902-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Bennett MV, Garré JM, Orellana JA, Bukauskas FF, Nedergaard M, Sáez JC. Connexin and pannexin hemichannels in inflammatory responses of glia and neurons. Brain Res. 2012;1487:3–15. doi: 10.1016/j.brainres.2012.08.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Orellana JA, Saez PJ, Shoji KF, et al. Modulation of brain hemichannels and gap junction channels by pro-inflammatory agents and their possible role in neurodegeneration. Antioxid Redox Signal. 2009;11(2):369–399. doi: 10.1089/ars.2008.2130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Garre JM, Retamal MA, Cassina P, et al. FGF-1 induces ATP release from spinal astrocytes in culture and opens pannexin and connexin hemichannels. Proc Natl Acad Sci U S A. 2010;107(52):22659–22664. doi: 10.1073/pnas.1013793107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Froger N, Orellana JA, Calvo CF, et al. Inhibition of cytokine-induced connexin43 hemichannel activity in astrocytes is neuroprotective. Mol Cell Neurosci. 2010;45(1):37–46. doi: 10.1016/j.mcn.2010.05.007. [DOI] [PubMed] [Google Scholar]
- 162.Boyce AK, Wicki-Stordeur LE, Swayne LA. Powerful partnership: crosstalk between pannexin 1 and the cytoskeleton. Front Physiol. 2014;5:27. doi: 10.3389/fphys.2014.00027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Sáez PJ, Vargas P, Shoji KF, Harcha PA, Lennon-Duménil AM, Sáez JC. ATP promotes the fast migration of dendritic cells through the activity of pannexin 1 channels and P2X7 receptors. Sci Signal. 2017;10(506):eaah7107. doi: 10.1126/scisignal.aah7107. [DOI] [PubMed] [Google Scholar]
- 164.Stankova V, Tsikolia N, Viebahn C. Rho kinase activity controls directional cell movements during primitive streak formation in the rabbit embryo. Development. 2015;142(1):92–98. doi: 10.1242/dev.111583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Horton SM, Lopez CL, Blevins E, et al. Pannexin 1 modulates axonal growth in mouse peripheral nerves. Front Cell Neurosci. 2017;11:365. doi: 10.3389/fncel.2017.00365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Kovalzon VM, Moiseenko LS, Ambaryan AV, Kurtenbach S, Shestopalov VI, Panchin YV. Sleep-wakefulness cycle and behavior in pannexin1 knockout mice. Behav Brain Res. 2017;318:24–27. doi: 10.1016/j.bbr.2016.10.015. [DOI] [PubMed] [Google Scholar]
- 167.Sappington RM, Calkins DJ. Contribution of TRPV1 to microglia-derived IL-6 and NFκB translocation with elevated hydrostatic pressure. Invest Ophthalmol Vis Sci. 2008;49(7):3004–3017. doi: 10.1167/iovs.07-1355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Sappington RM, Sidorova T, Long DJ, Calkins DJ. TRPV1: contribution to retinal ganglion cell apoptosis and increased intracellular Ca2+ with exposure to hydrostatic pressure. Invest Ophthalmol Vis Sci. 2009;50(2):717–728. doi: 10.1167/iovs.08-2321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Boyce AK, Kim MS, Wicki-Stordeur LE, Swayne LA. ATP stimulates pannexin 1 internalization to endosomal compartments. Biochem J. 2015;470(3):319–330. doi: 10.1042/BJ20141551. [DOI] [PubMed] [Google Scholar]
- 170.Billaud M, Sandilos JK, Isakson BE. Pannexin 1 in the regulation of vascular tone. Trends Cardiovasc Med. 2012;22(3):68–72. doi: 10.1016/j.tcm.2012.06.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Prochnow N, Abdulazim A, Kurtenbach S, et al. Pannexin1 stabilizes synaptic plasticity and is needed for learning. PLoS One. 2012;7(12):e51767. doi: 10.1371/journal.pone.0051767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Cea LA, Riquelme MA, Cisterna BA, et al. Connexin- and pannexin-based channels in normal skeletal muscles and their possible role in muscle atrophy. J Membr Biol. 2012;245(8):423–436. doi: 10.1007/s00232-012-9485-8. [DOI] [PubMed] [Google Scholar]
- 173.Cea LA, Riquelme MA, Vargas AA, Urrutia C, Sáez JC. Pannexin 1 channels in skeletal muscles. Front Physiol. 2014;5:139. doi: 10.3389/fphys.2014.00139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Dvoriantchikova G, Ivanov D, Panchin Y, Shestopalov VI. Expression of pannexin family of proteins in the retina. FEBS Lett. 2006;580(9):2178–2182. doi: 10.1016/j.febslet.2006.03.026. [DOI] [PubMed] [Google Scholar]
- 175.Weilinger NL, Maslieieva V, Bialecki J, Sridharan SS, Tang PL, Thompson RJ. Ionotropic receptors and ion channels in ischemic neuronal death and dysfunction. Acta Pharmacol Sin. 2013;34(1):39–48. doi: 10.1038/aps.2012.95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Wicki-Stordeur LE, Sanchez-Arias JC, Dhaliwal J, et al. Pannexin 1 differentially affects neural precursor cell maintenance in the ventricular zone and peri-infarct cortex. J Neurosci. 2016;36(4):1203–1210. doi: 10.1523/JNEUROSCI.0436-15.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Mannelli LC, Marcoli M, Micheli L, et al. Oxaliplatin evokes P2X7-dependent glutamate release in the cerebral cortex: a pain mechanism mediated by pannexin 1. Neuropharmacology. 2015;97:133–141. doi: 10.1016/j.neuropharm.2015.05.037. [DOI] [PubMed] [Google Scholar]
- 178.Zhang Y, Laumet G, Chen SR, Hittelman WN, Pan HL. Pannexin-1 up-regulation in the dorsal root ganglion contributes to neuropathic pain development. J Biol Chem. 2015;290(23):14647–14655. doi: 10.1074/jbc.M115.650218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Dolmatova E, Spagnol G, Boassa D, et al. Cardiomyocyte ATP release through pannexin 1 aids in early fibroblast activation. Am J Physiol Heart Circ Physiol. 2012;303(10):H1208–H1218. doi: 10.1152/ajpheart.00251.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Wonnenberg B, Tschernig T, Voss M, et al. Probenecid reduces infection and inflammation in acute Pseudomonas aeruginosa pneumonia. Int J Med Microbiol. 2014;304(5–6):725–729. doi: 10.1016/j.ijmm.2014.05.002. [DOI] [PubMed] [Google Scholar]
- 181.Lai CP, Bechberger JF, Thompson RJ, MacVicar BA, Bruzzone R, Naus CC. Tumor-suppressive effects of pannexin 1 in C6 glioma cells. Cancer Res. 2007;67(4):1545–1554. doi: 10.1158/0008-5472.CAN-06-1396. [DOI] [PubMed] [Google Scholar]
- 182.Lutz SE, Gonzalez-Fernandez E, Ventura JC, et al. Contribution of pannexin1 to experimental autoimmune encephalomyelitis. PLoS One. 2013;8(6):e66657. doi: 10.1371/journal.pone.0066657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Hainz N, Wolf S, Tschernig T, Meier C. Probenecid application prevents clinical symptoms and inflammation in experimental autoimmune encephalomyelitis. Inflammation. 2016;39(1):123–128. doi: 10.1007/s10753-015-0230-1. [DOI] [PubMed] [Google Scholar]
- 184.Velasquez S, Malik S, Lutz SE, Scemes E, Eugenin EA. Pannexin1 channels are required for chemokine-mediated migration of CD4+ T lymphocytes: role in inflammation and experimental autoimmune encephalomyelitis. J Immunol. 2016;196(10):4338–4347. doi: 10.4049/jimmunol.1502440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Jin Z, Ding Y, Xue R, et al. Involvement of interstitial cells of Cajal in bladder dysfunction in mice with experimental autoimmune encephalomyelitis. Int Urol Nephrol. 2017;49(8):1353–1359. doi: 10.1007/s11255-017-1597-8. [DOI] [PubMed] [Google Scholar]
- 186.Martins I, Wang Y, Michaud M, et al. Molecular mechanisms of ATP secretion during immunogenic cell death. Cell Death Differ. 2014;21(1):79–91. doi: 10.1038/cdd.2013.75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Bunse S, Locovei S, Schmidt M, et al. The potassium channel subunit Kvβ3 interacts with pannexin 1 and attenuates its sensitivity to changes in redox potentials. FEBS J. 2009;276(21):6258–6270. doi: 10.1111/j.1742-4658.2009.07334.x. [DOI] [PubMed] [Google Scholar]
- 188.Shibukawa Y, Sato M, Kimura M, et al. Odontoblasts as sensory receptors: transient receptor potential channels, pannexin-1, and ionotropic ATP receptors mediate intercellular odontoblast-neuron signal transduction. Pflugers Arch. 2015;467(4):843–863. doi: 10.1007/s00424-014-1551-x. [DOI] [PubMed] [Google Scholar]
- 189.Weilinger NL, Tang PL, Thompson RJ. Anoxia-induced NMDA receptor activation opens pannexin channels via Src family kinases. J Neurosci. 2012;32(36):12579–12588. doi: 10.1523/JNEUROSCI.1267-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Vaccari JP, Lotocki G, Alonso OF, Bramlett HM, Dietrich WD, Keane RW. Therapeutic neutralization of the NLRP1 inflammasome reduces the innate immune response and improves histopathology after traumatic brain injury. J Cereb Blood Flow Metab. 2009;29(7):1251–1261. doi: 10.1038/jcbfm.2009.46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Meng XF, Wang XL, Tian XJ, et al. Nod-like receptor protein 1 inflammasome mediates neuron injury under high glucose. Mol Neurobiol. 2014;49(2):673–684. doi: 10.1007/s12035-013-8551-2. [DOI] [PubMed] [Google Scholar]
- 192.Pelegrin P, Barroso-Gutierrez C, Surprenant A. P2X7 receptor differentially couples to distinct release pathways for IL-1β in mouse macrophage. J Immunol. 2008;180(11):7147–7157. doi: 10.4049/jimmunol.180.11.7147. [DOI] [PubMed] [Google Scholar]
- 193.Silverman W, Locovei S, Dahl G. Probenecid, a gout remedy, inhibits pannexin 1 channels. Am J Physiol Cell Physiol. 2008;295(3):C761–C767. doi: 10.1152/ajpcell.00227.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Bhaskaracharya A, Dao-Ung P, Jalilian I, et al. Probenecid blocks human P2X7 receptor-induced dye uptake via a pannexin-1 independent mechanism. PLoS One. 2014;9(3):e93058. doi: 10.1371/journal.pone.0093058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Xiong XX, Gu LJ, Shen J, et al. Probenecid protects against transient focal cerebral ischemic injury by inhibiting HMGB1 release and attenuating AQP4 expression in mice. Neurochem Res. 2014;39(1):216–224. doi: 10.1007/s11064-013-1212-z. [DOI] [PubMed] [Google Scholar]
- 196.Dahl G, Keane RW. Pannexin: from discovery to bedside in 11±4 years? Brain Res. 2012;1487:150–159. doi: 10.1016/j.brainres.2012.04.058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Orellana JA, Avendano BC, Montero TD. Role of connexins and pannexins in ischemic stroke. Curr Med Chem. 2014;21(19):2165–2182. doi: 10.2174/0929867321666131228191714. [DOI] [PubMed] [Google Scholar]
- 198.Khalafalla MG, Woods LT, Camden JM, et al. P2X7 receptor antagonism prevents IL-1beta release from salivary epithelial cells and reduces inflammation in a mouse model of autoimmune exocrinopathy. J Biol Chem. 2017;292:16626–16637. doi: 10.1074/jbc.M117.790741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Umazume T, Thomas WM, Campbell S, et al. Lacrimal gland inflammation deregulates extracellular matrix remodeling and alters molecular signature of epithelial stem/progenitor cells. Invest Ophthalmol Vis Sci. 2015;56(13):8392–8402. doi: 10.1167/iovs.15-17477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Gromova A, Voronov DA, Yoshida M, et al. Lacrimal gland repair using progenitor cells. Stem Cells Transl Med. 2017;6(1):88–98. doi: 10.5966/sctm.2016-0191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Spray DC, Hanani M. Gap junctions, pannexins and pain. Neurosci Lett. 2017 Jun 22; doi: 10.1016/j.neulet.2017.06.035. Epub. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Huang LY, Gu Y, Chen Y. Communication between neuronal somata and satellite glial cells in sensory ganglia. Glia. 2013;61(10):1571–1581. doi: 10.1002/glia.22541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Zhang X, Chen Y, Wang C, Huang LY. Neuronal somatic ATP release triggers neuron-satellite glial cell communication in dorsal root ganglia. Proc Natl Acad Sci U S A. 2007;104(23):9864–9869. doi: 10.1073/pnas.0611048104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Barr TP, Hrnjic A, Khodorova A, Sprague JM, Strichartz GR. Sensitization of cutaneous neuronal purinergic receptors contributes to endothelin-1-induced mechanical hypersensitivity. Pain. 2014;155(6):1091–1101. doi: 10.1016/j.pain.2014.02.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Fabbretti E. ATP-gated P2X3 receptors are specialised sensors of the extracellular environment. Adv Exp Med Biol. 2017;1051:7–16. doi: 10.1007/5584_2017_56. [DOI] [PubMed] [Google Scholar]
- 206.Fabbretti E. ATP P2X3 receptors and neuronal sensitization. Front Cell Neurosci. 2013;7:236. doi: 10.3389/fncel.2013.00236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Weaver JL, Arandjelovic S, Brown G, et al. Hematopoietic pannexin 1 function is critical for neuropathic pain. Sci Rep. 2017;7:42550. doi: 10.1038/srep42550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Faroni A, Smith RJ, Procacci P, et al. Purinergic signaling mediated by P2X7 receptors controls myelination in sciatic nerves. J Neurosci Res. 2014;92(10):1259–1269. doi: 10.1002/jnr.23417. [DOI] [PubMed] [Google Scholar]
- 209.Ino D, Sagara H, Suzuki J, Kanemaru K, Okubo Y, Iino M. Neuronal regulation of Schwann cell mitochondrial Ca2+ signaling during myelination. Cell Rep. 2015;12(12):1951–1959. doi: 10.1016/j.celrep.2015.08.039. [DOI] [PubMed] [Google Scholar]
- 210.Horton SM, Luna Lopez C, Blevins E, et al. Pannexin 1 modulates axonal growth in mouse peripheral nerves. Front Cell Neurosci. 2017;11:365. doi: 10.3389/fncel.2017.00365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Penuela S, Gehi R, Laird DW. The biochemistry and function of pannexin channels. Biochim Biophys Acta. 2013;1828(1):15–22. doi: 10.1016/j.bbamem.2012.01.017. [DOI] [PubMed] [Google Scholar]
- 212.Kwon TJ, Kim DB, Bae JW, et al. Molecular cloning, characterization, and expression of pannexin genes in chicken. Poult Sci. 2014;93(9):2253–2261. doi: 10.3382/ps.2013-03867. [DOI] [PubMed] [Google Scholar]
- 213.Fagerberg L, Hallström BM, Oksvold P, et al. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Mol Cell Proteomics. 2014;13(2):397–406. doi: 10.1074/mcp.M113.035600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Iwamoto T, Nakamura T, Doyle A, et al. Pannexin 3 regulates intracellular ATP/cAMP levels and promotes chondrocyte differentiation. J Biol Chem. 2010;285(24):18948–18958. doi: 10.1074/jbc.M110.127027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Ray A, Zoidl G, Weickert S, Wahle P, Dermietzel R. Site-specific and developmental expression of pannexin1 in the mouse nervous system. Eur J Neurosci. 2005;21(12):3277–3290. doi: 10.1111/j.1460-9568.2005.04139.x. [DOI] [PubMed] [Google Scholar]
- 216.Diezmos EF, Sandow SL, Markus I, et al. Expression and localization of pannexin-1 hemichannels in human colon in health and disease. Neurogastroenterol Motil. 2013;25(6):e395–e405. doi: 10.1111/nmo.12130. [DOI] [PubMed] [Google Scholar]
- 217.Zoidl G, Petrasch-Parwez E, Ray A, et al. Localization of the pannexin1 protein at postsynaptic sites in the cerebral cortex and hippocampus. Neuroscience. 2007;146(1):9–16. doi: 10.1016/j.neuroscience.2007.01.061. [DOI] [PubMed] [Google Scholar]
- 218.Penuela S, Celetti SJ, Bhalla R, Shao Q, Laird DW. Diverse subcellular distribution profiles of pannexin 1 and pannexin 3. Cell Commun Adhes. 2008;15(1):133–142. doi: 10.1080/15419060802014115. [DOI] [PubMed] [Google Scholar]
- 219.Ishikawa M, Iwamoto T, Nakamura T, Doyle A, Fukumoto S, Yamada Y. Pannexin 3 functions as an ER Ca2+ channel, hemichannel, and gap junction to promote osteoblast differentiation. J Cell Biol. 2011;193(7):1257–1274. doi: 10.1083/jcb.201101050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Cowan KN, Langlois S, Penuela S, Cowan BJ, Laird DW. Pannexin1 and pannexin3 exhibit distinct localization patterns in human skin appendages and are regulated during keratinocyte differentiation and carcinogenesis. Cell Commun Adhes. 2012;19(3–4):45–53. doi: 10.3109/15419061.2012.712575. [DOI] [PubMed] [Google Scholar]
- 221.Ambrosi C, Gassmann O, Pranskevich JN, et al. Pannexin1 and Pannexin2 channels show quaternary similarities to connexons and different oligomerization numbers from each other. J Biol Chem. 2010;285(32):24420–24431. doi: 10.1074/jbc.M110.115444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Ding J, Wang K, Liu W, et al. Pore-forming activity and structural autoinhibition of the gasdermin family. Nature. 2016;535(7610):111–116. doi: 10.1038/nature18590. [DOI] [PubMed] [Google Scholar]