

Abstract
The ability of athletes to train day after day depends in large part on adequate restoration of muscle glycogen stores, a process that requires the consumption of sufficient dietary carbohydrates and ample time. Providing effective guidance to athletes and others wishing to enhance training adaptations and improve performance requires an understanding of the normal variations in muscle glycogen content in response to training and diet; the time required for adequate restoration of glycogen stores; the influence of the amount, type, and timing of carbohydrate intake on glycogen resynthesis; and the impact of other nutrients on glycogenesis. This review highlights the practical implications of the latest research related to glycogen metabolism in physically active individuals to help sports dietitians, coaches, personal trainers, and other sports health professionals gain a fundamental understanding of glycogen metabolism, as well as related practical applications for enhancing training adaptations and preparing for competition.
Keywords: dietary carbohydrates, exercise recovery, muscle glycogen, sports nutrition
INTRODUCTION
During intense, intermittent exercise and throughout prolonged physical activity, muscle glycogen particles are broken down, freeing glucose molecules that muscle cells then oxidize through anaerobic and aerobic processes to produce the adenosine triphosphate (ATP) molecules required for muscle contraction.1 The rate at which muscle glycogen is degraded depends primarily upon the intensity of physical activity; the greater the exercise intensity, the greater the rate at which muscle glycogen is degraded. As a result, high-intensity activity, such as repeated sprinting, can quickly lower glycogen stores in active muscle cells, even though the total time of activity might be relatively brief (eg, 10 × 30-s sprints with short recovery intervals). In comparison, an endurance athlete who trains for hours at a time will also experience a marked decline in muscle glycogen, although at a slower rate of degradation than the sprinter.
The US Institute of Medicine’s recommended daily allowance for carbohydrate consumption in sedentary adult men and women is 130 g,2 with additional carbohydrates required to match that oxidized during physical activity, a value that varies with the duration and intensity of exercise. For instance, on days that involve only light physical activity of relatively short duration, considerably less carbohydrate is required to restore muscle and liver glycogen than on heavier training days. For that reason, current recommendations for carbohydrate intake in athletes vary to reflect the daily training load.3 However, athletes often do not consume enough dietary carbohydrates to meet current recommendations for the daily carbohydrate intake (8–12 g carbohydrate/kg of body weight [BW])3,4 considered necessary to fully replenish muscle glycogen stores.5–7 Less-than-optimal daily carbohydrate intake is likely a result of demanding training schedules, busy lives, confusion regarding the benefits of dietary carbohydrates, and inadequate understanding of basic sports nutrition concepts.8,9
Glycogen is a ubiquitous fuel source stored in the cytosol of cells, occupying 2% of the volume of cardiac cells,10 1%–2% of the volume of skeletal muscle cells, and 5%–6% of the volume of liver cells.11 The glycogen particles in liver cells can be 10 times larger than those in skeletal muscle cells, with each particle containing > 50 000 glucose molecules.12,13 Each gram of glycogen is stored with at least 3 g of water,14,15 making weight gain a noticeable response to glycogen super-compensation in many athletes. Conversely, rapid weight loss accompanies diets inadequate in carbohydrate content; as muscle glycogen is broken down, the associated water molecules are excreted in urine. After a few days, this weight loss subsides, and weight gain may occur as glycogen stores are replenished and additional water molecules are retained. Interestingly, neither short-term fasting nor prolonged sedentary behavior affect muscle glycogen stores, although cardiac muscle glycogen may be increased during fasting as gluconeogenic substrates such as amino acids and glycerol are converted to glucose and stored as glycogen to ensure the heart has adequate energy stores.10
For the body to be prepared for subsequent training and competition, it is essential that glycogen stores in muscle and liver be replenished as rapidly as possible. The intent of this review paper is to summarize how the science related to glycogen metabolism supports current recommendations for diet, training, and recovery in athletes and other engaged in regular physical activity.
RELIANCE ON CARBOHYDRATE OXIDATION DURING EXERCISE
To sustain the contraction of skeletal muscles during intermittent and continuous exercise of varying intensities and durations, active muscle cells require a constant supply energy in the form of ATP. That ATP is produced by the oxidation of fatty acids from the bloodstream and from intramuscular triglyceride stores, along with glucose supplied by the bloodstream and intramuscular glycogen stores. During training and competition, ATP production in muscle is prodigious. Even at rest, each muscle cell contains roughly 1 billion ATP molecules, all of which will be used and replaced every 2 minutes; during intense exercise, muscle ATP production can increase 1000-fold to meet the demands of intense muscle contraction.16
During exercise at intensities greater than approximately 60% maximal oxygen consumption (VO2max), blood glucose and muscle glycogen are the primary fuels oxidized to produce the ATP required to sustain exercise, in large part because more fast-twitch motor units are recruited as exercise intensity increases, increasing the reliance on carbohydrate as the predominant fuel source.1,17
GLYCOGEN AS A FUEL SOURCE
In addition to human muscle and liver cells, glycogen is stored in small amounts in brain cells, heart cells, smooth muscle cells, kidney cells, red and white blood cells, and even adipose cells.18 Glucose is a critical energy source for neurons in the brain and throughout the body,19 and under normal circumstances, glucose is the only fuel the brain uses to produce ATP; at rest, approximately 60% of the glucose found in blood is metabolized by the brain.20 Because the brain is an obligatory user of blood glucose, it is critical to maintain euglycemia (normal blood glucose concentration) during rest and exercise. The brain’s constant requirement for glucose is the primary factor underlying the current recommended daily allowance (RDA) for carbohydrates at 130 g/day.2
The > 80 g of glucose stored as liver glycogen is used to constantly replenish the 4 g of glucose circulating in the blood.20 To ensure the brain has an ample supply of glucose, the liver releases glucose into the bloodstream at a rate similar to the uptake of glucose from the blood into tissues, thereby stabilizing blood glucose concentration between 4.0 and 5.5 mmol/L (70–100 mg/dL). When liver glycogen stores fall to low levels, the liver can increase its reliance on gluconeogenic metabolism to produce glucose from amino acids and glycerol, although the rate of this production is limited and cannot keep pace with glucose removal from the blood during exercise. The use of muscle glycogen during exercise reduces glucose uptake from the blood, thereby helping to maintain blood glucose in the absence of exogenous carbohydrate intake. Sufficient carbohydrate ingestion during exercise helps maintain liver glycogen stores21,22 and has been reported to spare glycogen in type II (fast-twitch) muscle cells.23
In addition to the glycogen stored in muscle and liver, a small amount of glycogen is stored in brain cells (100× less than the glycogen content of muscle cells), specifically in astrocytes, nonneuronal glial cells that play an important role in stabilizing, insulating, and nourishing neurons.19,24,25 Astrocyte glycogen is metabolized to lactate, which then diffuses to nearby neurons to support their energy needs, an example of a very localized lactate shuttle.26 Lactate transporters (monocarboxylate transporter [MCT]) have been found in the neuronal membrane.24,27 Neurons also contain a small amount of glycogen.28
CARBOHYDRATES AND PERFORMANCE
In the 1920s, it became apparent that carbohydrate was an important fuel source for exercising muscles29; that blood glucose concentration seemed to be linked to fatigue during marathon running30; and that increasing dietary carbohydrate intake before a marathon, as well as consuming hard candies during the race, prevented weakness and fatigue.31 Despite those observations and the much earlier discovery of glycogen in 1858,13 the link among the carbohydrate content of the diet, muscle glycogen, and exercise capacity was not confirmed until the 1960s when Scandinavian researchers used the muscle biopsy technique to establish that muscle glycogen content had a major impact on endurance performance.32–35
It is now widely accepted that consuming a diet sufficient in carbohydrates, along with ingesting carbohydrates during and following exercise, can improve performance and speed recovery.1,3,36 In fact, the ability of carbohydrates to improve exercise performance is not limited to ingesting carbohydrates; simply rinsing the mouth with carbohydrate solutions without swallowing has been shown to improve aspects of exercise performance such as endurance capacity and bench-press repetitions to failure.37–39 It is also well established that beginning exercise with ample muscle glycogen stores is an important contributor to improved exercise performance; further, restoration of glycogen stores is essential for complete recovery and the maintenance of subsequent exercise capacity.3,4
WHOLE-BODY GLYCOGEN STORES
Whole-body glycogen content is approximately 600 g, a figure that varies widely based on body mass, diet, fitness, and recent exercise. Not surprisingly, skeletal muscle cells represent the largest depot of glycogen (see Table 1).40 During intense and prolonged exercise, the glycogen content of active muscle cells can be substantially reduced but does not fall further than approximately 10% of initial values.4 The content of glycogen in liver cells varies throughout each day depending upon the carbohydrate content of the diet, the time between meals, and the intensity and duration of recent physical activity. Although muscle and liver glycogen represent just 4% of the body’s total fuel stores, muscle glycogen is the primary fuel during exercise of moderate or greater intensity.37,41
Table 1.
The glycogen content of liver and muscle
| Tissue | Average (g) | Normal range (g) |
|---|---|---|
| Muscle | 500 | 300–700 |
| Liver | 80 | 0–160 |
Adapted from Hargreaves (2012).40
SHAPE OF THE GLYCOGEN PARTICLE
Because glycogen is comprised of individual glucose molecules, there is no such thing as a glycogen molecule; it is more accurate to refer to a glycogen particle, knowing that the particles can vary widely in size due to differences in the total number of glucose molecules within each particle. All glycogen particles are initiated by glycogenin, an enzyme that forms a complex with glycogen synthase to link uridine diphosphate–glucose molecules to create the beginnings of a glycogen particle.42 Glycogen synthase and branching enzyme then work in concert to enlarge the glycogen particle. As shown in Figure 1, glycogen synthase creates α-1,4-glycosidic linkages to create a strand of glucose molecules, and the branching enzyme establishes α-1,6 bonds between glucose molecules to create branches every 8–12 glucose molecules; the branches increase the density, solubility, and surface area of the glycogen particle.13,42
Figure 1.
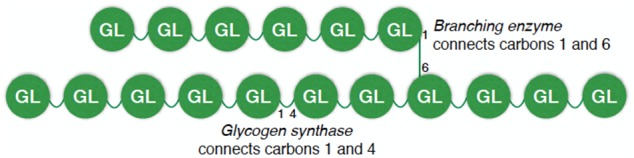
Depiction of glycogen, a large spherical particle formed by linking glucose molecules into strands and branches.
The regulation of glycogenin formation is not well understood, but the cellular content of glycogenin influences the rate and extent of glycogen storage.43,44 Glycogen particles have been categorized into 2 forms based upon their size: 1) proglycogen and 2) macroglycogen.43–47 Proglycogen particles comprise roughly 15% of total glycogen content, are sensitive to dietary carbohydrates, and are first to add glucose units after glycogen depletion; additional glucose units are then more slowly added to create the larger macroglycogen particles. This observation may explain the biphasic nature of glycogen repletion: rapid for the first few hours and more slowly thereafter.
LOCATION OF GLYCOGEN STORAGE
Glycogen particles are distributed within the muscle cell to support the local energy needs of the cell during exercise (see Figure 2).48 Intermyofibrillar glycogen particles constitute roughly 75% of total muscle glycogen and are conveniently located adjacent to the sarcoplasmic reticulum and mitochondria.48 Intramyofibrillar glycogen particles are dispersed among the contractile filaments and represent 5%–15% of total glycogen; subsarcolemmal glycogen particles—another 5%–15% of total glycogen—are found between the sarcolemma and the contractile filaments.48,49
Figure 2.

The intracellular locations of skeletal muscle glycogen. Image © Human Kinetics. Used with permission. Values for glycogen distribution are from Schweitzer et al (2017).48
Glycogen from all 3 cellular “compartments” is used during exercise, but it appears that the intramyofibrillar glycogen use is greater in both type I (slow-twitch) and type II (fast-twitch) fibers.50 Nielsen et al.51 used transmission electron microscopy to show that intramyofibrillar glycogen was preferentially oxidized in both types I and II muscle fibers during exhaustive cross-country ski racing. Intramyofibrillar glycogen is used by the sarcoplasmic reticulum to allow for calcium release and muscle contraction, so its depletion likely contributes to fatigue.51,52
ROLES OF GLYCOGEN
Muscle glycogen is not only a fuel source, but it is also a fuel sensor, a regulator of signaling pathways involved in training adaptation, and an influencer of intracellular osmolality.4,13 Additional details on muscle glycogen metabolism can be found in excellent reviews by Burke et al,4 Ivy,41 Jensen and Richter,17 Knuiman et al.,53 Nielsen and Ortenblad,49 Philp et al.,13 and Schweitzer et al.,48 as well as in other reviews published over the years.
MEASUREMENT OF GLYCOGEN
Much of our understanding of how muscle glycogen stores decline during physical activity and are restored during subsequent rest comes from studies that used the muscle biopsy technique. A muscle biopsy requires a small incision through the skin and underlying tissue so that a biopsy needle can be inserted into a muscle and a small piece of tissue can be extracted. The tissue is quickly frozen and eventually prepared for microscopic analysis, sometimes using special dyes to help identify various cellular structures and components.
The vastus lateralis muscle of the quadriceps muscle group on the front of the thigh is a common site for muscle biopsies because that particular muscle is active during running and cycling, exercise modalities often used in laboratory studies. A biopsy taken prior to exercise establishes the baseline muscle glycogen concentration, and subsequent biopsies taken during and after exercise are used to chart the changes in glycogen levels over time. Liver glycogen stores in humans are indirectly estimated by nuclear magnetic spectroscopy or other noninvasive techniques. Muscle glycogen concentration is often expressed as millimoles of glycogen per kilogram of tissue, either as wet weight or dry weight (tissue water removed.) Dry-weight values are 4.325 times greater than wet-weight values.
FACTORS INFLUENCING GLYCOGEN STORES
Glycogen stores in liver and muscle decrease during physical activity; the longer and more intense the activity, the greater the rate and overall reduction of glycogen stores. Consuming a diet that supplies ample carbohydrates and energy (calories) to match or exceed daily expenditures results in a gradual supercompensation of muscle glycogen stores over days and weeks, a response that can be further enhanced by dietary interventions (see Table 2).33,54–68 Improved physical fitness is an additional stimulus for enhanced muscle glycogen stores, helping ensure that ample carbohydrate energy is available to fuel intense and prolonged training and competition.
Table 2.
Dietary and exercise interventions that influence muscle glycogen synthesis
| Intervention | Response | Reference |
|---|---|---|
| Regular training + diet high in CHO content (>8 g/kg BW/d) | Supercompensated muscle glycogen stores compared with the start of training and greater than if a low-CHO diet is consumed | Ahlborg and Brohult (1967)54 |
| Classic glycogen loading: 3 d of hard training on a low-CHO diet (<5 g/kg BW/d) followed by 3 d of tapered training on a high-CHO diet | Supercompensated muscle glycogen stores compared with before the intervention but training is very difficult both physically and psychologically during the low-CHO phase | Bergstrom et al. (1967)33 and (1972)55 |
| Modified glycogen loading: 3-d taper on a high-CHO diet with 24-h rest prior to competitions | Supercompensated muscle glycogen stores similar to the classic loading regimen | Sherman et al. (1981)56; Bussau et al. (2002)57 |
| Train low, compete high: purposefully reduce daily CHO intake or train after an overnight fast or withhold CHO intake during and for 2 h following a hard training session to promote adaptations that result in glycogen supercompensation | Training low reduces the capacity to train hard and makes training psychologically challenging. No clear evidence of additional benefits to glycogen stores or performance | Bartlett et al. (2015)58; Burke (2010)59 |
| Train high, sleep low: train with high CHO availability in the evening, no CHO replacement prior to sleep, train with low CHO availability in the morning | Evidence of improved performance compared with consuming a consistently high-CHO diet during training. Performance benefits could be due to higher muscle glycogen | Hawley (2014)60; Marquet et al. (2016)61,62; Yeo et al. (2008)63 |
| Protein supplementation | When dietary CHO intake is not adequate, consuming 0.3–0.4 g protein/kg BW has been shown to augment glycogen synthesis | Betts and Williams (2010)64; Ivy et al. (2002)65 |
| Creatine loading | Some studies found enhanced muscle glycogen storage with creatine loading, whereas other studies found no effect | Roberts et al. (2016)66; Sewell et al. (2008)67 |
| Fat loading, train low | >2 weeks on a high-fat, low-CHO diet increases muscle fat oxidation and spares muscle glycogen during moderate-intensity exercise (eg, 60% VO2max). However, fat loading can impair muscle CHO oxidation and performance at higher intensities | Bartlett et al. (2015)58’ Burke and Kiens (2006)68 |
Abbreviations: BW, body weight; CHO, carbohydrate; VO2max, maximal oxygen consumption.
It is clear from the scientific evidence that consuming a diet high in carbohydrate content (eg, often > 60% of total calories or ideally the amount of carbohydrate sufficient to replace the carbohydrate oxidized during rest and training), combined with proper training and adequate rest, results in supercompensation of muscle glycogen stores. However, it appears that many athletes may not be consuming enough carbohydrates on a daily basis to fully restore muscle glycogen. For example, Mullins et al.7 reported that a group of 19 female heptathletes consumed an average of 5.2 g of carbohydrate/kg BW/ day (339 g/d), compared with the current carbohydrate-intake recommendation of 6.0–12.0 g/kg of BW/day.3,4 Devlin et al.6 assessed the diets of 66 elite and subelite soccer and Australian Rules football players using multiple-pass 24-hour dietary recalls and found that daily carbohydrate intake ranged from 2.9 to 4.6 g/kg BW. The results were similar for professional English soccer players who ingested an average of 4.2–6.4 g/kg BW/day during match and training days, respectively.5 This apparent gap between actual carbohydrate intake and current recommendations may not be as worrisome as it may seem because, as discussed later in this review, even a carbohydrate intake of < 5–6 g/kg BW/day may be sufficient to maintain muscle glycogen at levels that can sustain the demands of normal training.
MUSCLE GLYCOGEN STORAGE
The synthesis of muscle glycogen depends upon uptake of glucose molecules from the blood into muscle cells. When carbohydrates are ingested at rest—and in the recovery period after exercise—the entry of glucose into muscle cells is facilitated by the hormone insulin. When glucose enters the muscle cell at rest or during exercise, it is immediately phosphorylated to glucose-6-phosphate by the enzyme hexokinase. Glucose-6-phosphate can then be oxidized through glycolysis and the Krebs cycle to produce ATP for immediate use by the cell, or it can be stored as glycogen. In fact, glucose-6-phosphate allosterically activates glycogen synthase, stimulating the addition of glucose molecules to the glycogen particle. The activity of the glycogen synthase enzyme is controlled by a cascade of events that rely on phosphorylation and dephosphorylation reactions that decrease and increase the activity of the enzyme in concert with similar phosphorylation-dephosphorylation reactions that control muscle glycogenolysis via the glycogen phosphorylation enzyme described below (see Figure 3).
Figure 3.

A simplified overview of glycogen metabolism at rest and during exercise. The sarcolemma separates the muscle cell interior from the interstitial fluid that surrounds the cell. At rest (left side), the consumption of carbohydrate stimulates the release of insulin from the pancreas. Insulin molecules bind to insulin receptors embedded in the sarcolemma. That binding sparks a cascade of intracellular responses that result in the movement of GLUT4 glucose transporters from the interior of the muscle cell into the sarcolemma, allowing for glucose to move into the cell. Once inside the muscle cell, glucose molecules are readied for inclusion into glycogen. Glycogenin is an enzyme that forms the center of glycogen particles, allowing for the initial formation of glycogen strands. During exercise (right side), GLUT4 transporters move into the sarcolemma without the assistance of insulin, aiding in glucose uptake into the cell. Simultaneously, glycogen degradation increases in response to changes in the concentration of metabolites inside the cell. The glucose molecules from the blood and those released from glycogen are oxidized to produce the adenosine triphosphate (ATP) molecules required to sustain muscle contraction.
The activity of glycogen synthase is also influenced by the glycogen content of the muscle cell; high glycogen synthase activity is associated with low glycogen levels.17,37 After exercise of sufficient intensity and duration to substantially lower muscle glycogen stores, the increase in the permeability of the sarcolemma to glucose—thanks to increased GLUT4 translocation to the membrane during exercise and the presence of GLUT1 transporters at rest12—aids glucose uptake and glycogen synthesis. If postexercise carbohydrate supplementation is not maintained, GLUT4 transporters are removed from the membrane after 30–60 minutes.41 In short, the reduction in muscle glycogen stores that occurs during exercise is a major driving factor for subsequent glycogenesis.69
After exercise, the restoration of muscle glycogen occurs in a biphasic manner. During the first phase, glycogen synthesis is rapid (12–30 mmol/g wet weight/h), does not require insulin, and lasts 30–40 minutes if glycogen depletion is substantial. The second phase depends on insulin and occurs at a slower rate with euglycemia (2–3 mmol/g wet weight/h), a rate that can be increased to 8–12 mmol/g wet weight/hour with additional carbohydrate intake. Periodic carbohydrate supplementation can result in supercompensation of glycogen stores, an advantage after tasks requiring hours of sustained physical effort. In fact, the second-phase effect can be sustained for several days when carbohydrate intake is maintained.37 Liver glycogen is rapidly restored during postexercise feeding,22 helping ensure the maintenance of normal blood glucose. Even when carbohydrates are not ingested after exercise, glycogen repletion can occur at slow rates (1–2 mmol/kg wet weight/h) from gluconeogenesis70 and the conversion of lactate to glucose.71
During vigorous exercise, insulin release is blunted, and epinephrine (adrenalin) is released from the adrenal glands. Epinephrine causes phosphorylation of intramyofibrillar glycogen synthase, ensuring that glycogen synthesis is slowed as glycogen degradation rapidly increases.41 The rate of glycogen degradation (glycogenolysis) depends upon exercise intensity; during all-out exercise, glycogen can release glucose molecules at a rate of 40 mmol glucose/kg wet weight/minute. During low-intensity exercise, glycogen breakdown may occur at only 1–2 mmol glucose/kg wet weight/minute.40 In addition to the stimulating effect of epinephrine binding to β-adrenergic receptors on the sarcolemma, muscle contraction produces a variety of metabolites, such as adenosine diphosphate (ADP), adenosine monophosphate (AMP), and inosine monophosphate (IMP), along with increased levels of calcium and inorganic phosphate, which also enhance the activity of glycogen phosphorylase.40,72
As exercise progresses, the activity of glycogen phosphorylase falls as glycogen stores are reduced and as plasma free fatty acids become more available as substrates. Endurance training increases muscle glycogen stores and reduces the reliance on glycogen as a result of the increased use of free fatty acids by active muscle cells,40 a metabolic adaptation that allows for improved performance. Conversely, the depletion of muscle glycogen causes fatigue.32–35 When muscle glycogen stores are low, muscle cells cannot produce ATP rapidly enough to maintain exercise intensity,73 the very definition of fatigue. For example, Krustrup et al.74 reported that after a high-level soccer game, muscle glycogen depletion was associated with altered calcium cycling in the sarcoplasmic reticulum, reduced force of maximal voluntary contraction, and increased muscle soreness. If daily carbohydrate intake is insufficient to fully replace the glycogen metabolized during hard labor or training, muscle glycogen concentration in active muscles will fall progressively over a period of days, a circumstance that is well established in the scientific literature.75–77
GLYCOGEN AND ADAPTATION TO TRAINING
As noted in Table 2, training and/or sleeping with low muscle glycogen levels enhances intracellular signaling and consequent adaptations that upregulate the oxidative capacity of muscle cells and possibly improve endurance performance.58–63,78,79 Athletes can follow train-low regimens periodically during their training season, but it is likely that many athletes train with low muscle glycogen on occasions when their dietary carbohydrate intake is inadequate. As a result, they accidentally benefit from the enhanced metabolic signaling associated with low muscle glycogen.
There is even less certainty regarding how muscle glycogen stores influence the adaptations associated with resistance training because there are far fewer studies compared to the number of studies that have focused on the influence of glycogen levels on the adaptations to endurance and interval training. Muscle glycogen is an important substrate for resistance training because repeated contractions of near-maximal loads stimulate glycogenolysis, resulting in a reduction in glycogen stores of 25%–40%.80,81 However, based upon the available literature, it appears as though low muscle glycogen levels do not impede muscle protein synthesis or the overall anabolic response to resistance training.53,82,83
NORMAL VARIATION IN GLYCOGEN CONCENTRATION
In trained and well-fed endurance athletes, muscle glycogen concentration is roughly 150 mmol/kg wet weight following at least 8–12 hours of rest but can reach supercompensated levels of 200 mmol/kg wet weight in highly fit, rested athletes after a few days on high-carbohydrate diets56; after prolonged intense exercise, muscle glycogen might fall to < 50 mmol/kg wet weight.4,84 When muscle glycogen falls to < 70 mmol/kg wet weight, calcium release from the sarcoplasmic reticulum is impaired, as is peak power output,48,53 implying that there is a threshold level of muscle glycogen below which muscle function and performance are impaired. It may be that the average value for muscle glycogen concentration does not accurately reflect the intramyofibrillar glycogen stores, which appear to have the greatest impact on muscle function.
As an example, an athlete who begins a hard 2-hour training session with a muscle glycogen concentration of 150 mmol/kg wet weight may experience a 50% decline in muscle glycogen to 75 mmol/kg wet weight, close to the theoretical threshold at which muscle function is impaired, making subsequent training or performance difficult.85 Consuming sufficient carbohydrates to raise muscle glycogen concentration above 100 mmol/kg wet weight would be enough to allow for additional exercise > 4 hours later, assuming a resynthesis rate of 5–6 mmol/kg/hour. Figure 4 depicts how muscle glycogen levels might vary during 4 days of hard training followed by 2 days of light training.85 Because muscle glycogen resynthesis is a relatively slow process (see below), athletes typically train with varied muscle glycogen stores that are well below supercompensated levels.
Figure 4.

Muscle glycogen levels can vary widely during training, only reaching supercompensated levels after a few days of rest and light training. In this example, muscle glycogen levels decline during training sessions and are partially restored during subsequent rest and after adequate carbohydrate intake. During hard 2-a-day training sessions (day 3), glycogen concentration can be lowered to the point at which contractile dysfunction (fatigue) occurs. Athletes typically train with muscle glycogen stores that are adequate to meet the demands of training (eg, between 75 and 150 mmol/kg wet weight) even though those stores might be considered suboptimal. Illustration based on data from Sherman and Wimer (1991).85
TIME COURSE OF GLYCOGEN REPLENISHMENT
Whenever muscle glycogen stores are reduced as a result of physical activity, consumption of an adequate amount of carbohydrate is required to restore glycogen to normal levels or above (supercompensation).86 An average glycogen repletion rate of 5–6 mmol/kg wet weight/hour is usually required for complete glycogen restoration within 24 hours.87 For example, Starling et al.88 reported that 24 hours on a high-carbohydrate diet (9.8 g/kg BW/d) restored 93% of the muscle glycogen oxidized during a prior 2-hour cycling bout at 65% VO2max; a low-carbohydrate diet (1.9 g/kg BW/d) restored only 13%. Other researchers89 demonstrated that when individuals consumed a high-glycemic carbohydrate diet (∼10 g carbohydrate/kg BW/d; including corn flakes, bread, potatoes), the muscle glycogen storage rate was 106 mmol/kg wet weight/day (an hourly average of 4.4 mmol/kg wet weight).
GLYCOGEN STORES AND TRANING CAPACITY
Athletes who train hard most days of the week, at times completing multiple training sessions each day, likely do so with muscle glycogen stores that are rarely fully replenished. For example, Sherman et al.77 found that participants who were fed moderate- or high-carbohydrate diets (5 vs 10 g/kg BW/d) over 7 days of training were able to maintain their glycogen stores from day to day on the high-carbohydrate diet but experienced a 30%–36% decline in muscle glycogen on the moderate-carbohydrate diet. However, the lower muscle glycogen levels did not impair training capacity or exercise performance (run or cycle to exhaustion at 80% VO2 peak.) In their review of the literature, Sherman and Wimer85 came to the conclusion that high-carbohydrate diets can prevent a fall in muscle glycogen stores over weeks of intense training; in contrast, moderate-carbohydrate diets maintain muscle glycogen stores at levels that are lower but still sufficient to meet the demands of hard training. In an exhaustive review of the literature on dietary carbohydrate intake among athletes, Burke et al.90 hypothesized that athletes can adapt to lower muscle glycogen stores in ways that protect training capacity and performance. Although training capacity and performance may not be adversely affected by the consumption of moderate-carbohydrate diets, performance is impaired on low-carbohydrate diets. Burke et al.91 reported that race walkers who trained for 3 weeks on a low-carbohydrate diet (2.1 g/kg BW/d) did not improve their 10-km race walk performance compared with those who consumed a high-carbohydrate diet (8.6 g/kg BW/d). The high-carbohydrate group improved their performance by 6.6%, whereas performance in the low-carbohydrate group did not change (−1.6%) from baseline measures. In practical terms, athletes should be educated and encouraged to consume enough carbohydrates to replenish at least a sizable portion of their muscle glycogen stores so that training intensity can be maintained from day to day. Fortunately, athletes in training tend to gravitate to high-carbohydrate diets,1 helping ensure that glycogen stores do not drop so low that training is impaired. In addition, the unavoidable daily fluctuations in muscle glycogen stores are an important intracellular signal to stimulate the adaptations required for improved performance,92 augmented intracellular responses that also follow purposeful reductions in muscle glycogen as part of “periodized nutrition” strategies.93
GLYCOGEN RESYNTHESIS AFTER EXERCISE
Immediately after physical activity, muscle cells that sustained a substantial decrease in glycogen content are metabolically prepared for rapid glycogenesis. In brief, glycogen use during exercise turns on glycogen synthesis during recovery. When carbohydrates are ingested soon after exercise, insulin release from the pancreas, insulin sensitivity in muscle cells, glucose uptake by muscle cells, and glycogen synthase activity within muscle cells all increase,94 responses that can remain elevated for 48 hours.4,95 Repeated intake of 1.0–1.2 g carbohydrate/kg BW/hour after exercise takes advantage of these metabolic circumstances to stimulate high rates of glycogen synthesis (10–11 mmol/kg wet weight/h).4,96 When 24 hours of rest occur between exercise sessions, ingesting a total of 10 g/kg BW, along with enough energy (calories) to maintain energy balance, will maximize glycogen restoration.4,65,97
As noted above, timing of carbohydrate intake following physical activity is very important during training and competition requiring multiple efforts during a single day. If a 2-hour bout of exercise reduces muscle glycogen content by 75 mmol/kg wet weight and an athlete has 6 hours of rest prior to the next exercise session, ingesting 1.0–1.2 g carbohydrate/kg BW/hour in meals or snacks could theoretically restore 60 mmol/kg wet weight, or 80% of the oxidized glycogen.
Consuming > 10 g carbohydrate/kg BW/day appears to be of no additional benefit to glycogen restoration.4 A major factor that influences the rate and extent of muscle glycogen repletion is total energy intake; even if adequate amounts of carbohydrates are ingested after exercise, glycogen repletion will not be complete unless sufficient energy (calorie) intake occurs.98 This can be an important consideration for athletes involved in weight-class sports and those attempting to lose weight for other reasons.
To be clear, consumption of 10 g carbohydrate/kg BW/day is necessary only when glycogen deficits are large or on those occasions when glycogen loading is desired prior to competition or exhaustive physical activity. Consumption of 10 g carbohydrate/kg BW/day represents a sizable amount of carbohydrate; for a 50-kg (110-lb) athlete, 10 g carbohydrate/kg BW/day is the equivalent of 500 g of carbohydrate (2000 kcal). Daily carbohydrate intake should reflect the extent of carbohydrate oxidation during training: low on light training days, substantially higher on days of intense or prolonged training. Table 3 contains related practical recommendations.3,4
Table 3.
Recommendations for daily carbohydrate intake for athletes involved in repeated days of strenuous, prolonged physical activity and training
| Exercise intensity | Description | Dietary carbohydrate | Comments |
|---|---|---|---|
| Low | Easy activity such as yoga, tai chi, walking, or any exercise done at a light effort (can easily talk or sing during the activity) | 3–5 g/kg BW/d | Normal dietary intake is usually sufficient to restore muscle glycogen content |
| Moderate | One hour or more of activity such as walking, jogging, swimming, bicycling at a modest effort (can carry on a conversation without problem, but cannot sing) | 5–7 g/kg BW/d | A diet in which at least 50% of the energy (calories) comes from carbohydrate food is usually sufficient to restore muscle glycogen content |
| High | One hour or more hard exercise such as interval training, running, swimming, bicycling at a modest effort (can carry on only very brief conversations) | 6–10 g/kg BW/d | Postexercise carbohydrate/protein intake, with high-carbohydrate meals and snacks, is needed to fully restore muscle glycogen within 24–36 h |
| Very-high | Very hard exercise for an hour or more or very prolonged exercise such as interval training, ice hockey, soccer, basketball, running, swimming, bicycling at an intense effort (cannot speak during the effort) | 8–12 g/kg BW/d | Postexercise carbohydrate/protein intake, with high-carbohydrate meals and snacks, is needed to fully restore muscle glycogen within 24–36 h |
TIME COURSE OF GLYCOGEN RESYNTHESIS
If glycogen depletion is only 40 mmol/kg wet wt and a sufficient amount of high-glycemic carbohydrates are ingested immediately after exercise and again at 30-minute intervals, complete glycogen restoration might occur after 4 or 5 hours. On the other hand, if glycogen depletion is 150 mmol/kg wet wt, full repletion might require close to 24 hours because the maximal rate of glycogen synthesis (10 mmol/kg wet wt/h) can be maintained for only approximately 4 hours before the rate slowly declines to roughly 50% of maximum (4–6 mmol/kg wet wt/h).4,41
Supercompensation of muscle glycogen stores can occur following training if sufficient amounts of carbohydrates are consumed (eg, 8–10 g/kg BW/d),33,34,55–57,99 requiring 24–72 hours of rest or very light exercise following the last training session.4 Muscle damage can retard the rate of muscle glycogen synthesis,99–103 a response thought to be caused by a reduction in glucose uptake by muscle cells due to the disruption of the sarcolemmal membrane, a finding that underscores the importance of adequate rest following hard training to allow for adequate glycogen restoration.
AMOUNT AND TYPE OF DIETARY CARBOHYDRATE
In their review of the literature, Burke et al.4 concluded that long-term glycogen recovery (eg, ≥24 h) is not affected by timing or carbohydrate type but is most influenced by the total amount of carbohydrate ingested. It is true that fructose better stimulates liver glycogen restoration and glucose does the same for muscle glycogen,104 but most physically active people normally ingest enough fructose and glucose in foods and beverages to restore liver glycogen. Consequently, there is no need to be concerned with the adequacy of dietary fructose intake. It should be noted that combinations of glucose, fructose, and sucrose consumed in sports drinks during exercise have been shown to enhance the rate of fluid absorption from the proximal small intestine105 and improve the rate of carbohydrate oxidation during exercise,106,107 2 important factors in sustaining exercise performance.
Solid and liquid forms of carbohydrates are associated with similar rates of glycogen synthesis,108–109 so athletes can meet their daily carbohydrate needs by consuming the carbohydrate-rich foods and beverages they most enjoy. Interestingly, Cramer et al.110 showed that fast foods (in 2 meals, including 1 meal with french fries) were similar to sports supplements of similar carbohydrate and energy content in promoting muscle glycogen resynthesis in the 4 hours following 90 minutes of glycogen-depleting cycling exercise.
HIGH-GLYCEMIC CARBOHYDRATES
In the hours soon after exercise, consuming high–glycemic index (GI) foods can speed muscle glycogen restoration. Low-GI foods are digested and absorbed more slowly than high-GI foods, differences that result in a slower rise in blood glucose and insulin levels,111 an effect that can last for hours after eating.112 Lower blood insulin levels are associated with greater oxidation of fatty acids during exercise113 and reduced degradation of muscle glycogen.114
The effect of high- versus low-GI meals/foods on exercise performance during continuous or intermittent exercise is not clear.115 Little et al.116 studied the effect of consuming a low-GI meal (lentils; GI = 26) or a comparatively high-GI meal (mashed potatoes, bread, and egg whites; GI = 76) prior to 90-minutes of intermittent treadmill exercise. The meals were consumed 2 hours prior to exercise. Compared with a placebo treatment (no meal), both the low- and high-GI meals improved the total run distance during sprints conducted in the last 15 minutes of the 90-minute session. In contrast with no pre-exercise meal, muscle glycogen levels prior to the final 15-minute segment of exercise were similarly higher with both low- and high-GI meals. The authors attributed improved run performance to higher muscle (and possibly liver) glycogen levels prior to the final sprints.
Consuming high-GI carbohydrates is effective in increasing muscle glycogen stores after exercise. Burke et al.89 fed participants either high- or low-GI meals during 24 hours of recovery after completing 2 hours of cycling exercise at 75% VO2max and four 30-second, all-out sprints. The high-GI diet resulted in greater glycemic and insulinemic responses, along with greater restoration of muscle glycogen (106 mmol/kg wet weight vs 72 mmol/kg wet weight for the low-GI diet.) Other researchers have reported enhanced pre-exercise muscle glycogen storage and greater muscle glycogen use during exercise 3 hours following a high-GI (GI = 80) meal compared with a low-GI (GI = 36) meal.115,116 Rauch et al.117 used potato starch to supplement the diets of participants for 3 days prior to 3 hours of cycling exercise, including a 60-minute performance ride. Increasing the carbohydrate content of the diet to 10.5 g/kg BW/day (vs 6.2 g/kg BW/day) resulted in 47% greater pre-exercise muscle glycogen stores, better cycling performance, and enhanced reliance on muscle glycogen as fuel.
Consuming high-GI foods is important on occasions when rapid resynthesis of muscle glycogen is critical, as is the case during 2-a-day training and competitions requiring multiple games/matches during a single day. However, Brown et al.118 reported that consuming high-glycemic foods (only 2 g/kg BW) during a 3-hour period following glycogen-depleting exercise did not improve a 5-km cycling time trial compared with the consumption of an isoenergetic, low-glycemic meal, even though the high-GI meal promoted a greater insulinemic response. As is often the case in science, additional research is needed to further clarify the conditions in which consuming high-GI foods benefits glycogen restoration and performance.
WAXY STARCH VERSUS RESISTANT STARCH
Waxy starches from varietals of potatoes, corn (maize), and barley are high in amylopectin and low in amylose; amylopectin is less resistant to digestion because its glucose chains are more highly branched compared with amylose. For that reason, waxy starches have been studied to assess how their ingestion influences glycogen metabolism and exercise performance.119–121 In 1996, Jozsi et al.121 published the results of a study in which 8 male participants completed 4 exercise trials in which they cycled for 60 minutes at 75% VO2max, followed by six 1-minutes sprints, a protocol that lowered muscle glycogen content. During the subsequent 12 hours in each trial, the participants consumed 3000 kcal, 65% of which was carbohydrate as 1) glucose, 2) maltodextrin (often referred to as glucose polymers, relatively short chains of glucose molecules), 3) waxy corn starch (100% amylopectin), and 4) resistant starch (100% amylose). Postexercise muscle glycogen concentrations were similar among treatments, but 24 hours later, less glycogen had been replenished with resistant starch compared with the other treatments. Total glycogen repletion with glucose was greater than that with waxy starch was greater than that with maltodextrin was greater than that with resistant starch. A companion study by Goodpastor et al.119 reported that cycling performance in 10 male participants was improved by the pre-exercise consumption of glucose and waxy starch, but not with resistant starch, findings that further emphasize that the digestibility of carbohydrates is an important factor in performance improvement and glycogen replenishment. Additional research on the metabolic and performance responses to the ingestion of waxy starches is warranted simply because of the dearth of research in this area.
In terms of overall health, high-quality carbohydrates from unprocessed or minimally processed whole grains, vegetables, beans, dairy foods, and fruits also provide numerous vitamins, minerals, fiber, and many important phytonutrients. For individuals who are physically active on a daily basis, energy needs can easily exceed 3000 kcal/day, resulting in increased dietary intake of carbohydrates, proteins, and a wide range of micronutrients. Increased consumption of high-quality carbohydrate foods, such as potatoes and grains, can help ensure adequate consumption of nutrients vital to health, recovery, repair, adaptation, growth, and performance.
EXTENDED FASTING AND KETOSIS
Aside from the purposeful manipulation of muscle glycogen concentration by diet and training (nutrition periodization), periods of extended fasting during Ramadan or in attempts to lose body weight result in metabolic responses that are usually contrary to maintaining high muscle glycogen concentrations, especially if training continues during the fasted state.122 The fasting that occurs between meals, during sleep, or even during more extended periods of fasting has minimal effect on muscle glycogen concentration in resting individuals because muscle glycogen is not a major fuel substrate at rest. Prolonged fasting and very low–carbohydrate diets result in ketosis (ketoacidosis), sparing liver and muscle glycogen. As a result, ketotic diets and the ingestion of ketone bodies have been suggested as possible ergogenic aids, particularly for endurance and ultra-endurance athletes.123 However, in a 2017 review of the literature, Scott and Deuster124 concluded that the existing evidence for ketosis to benefit performance or glycogen replenishment is not persuasive. Related to that conclusion, Vandoorne et al.125 reported that ingesting ketone esters during a 5-hour recovery period following glycogen-depleting exercise had no effect on the replenishment of muscle glycogen but did activate the intracellular signaling pathways of adenosine monophosphate kinase (AMPK) and mechanistic target of rapamycin complex 1 (mTORC1) in ways that suggest that muscle protein synthesis might be stimulated. In short, more research is needed to further clarify the metabolic and performance responses to ketosis—whether induced by fasting, prolonged low-carbohydrate diets, or by the ingestion of ketone bodies—across performance parameters, with special reference to the mental and physical responses during ultra-endurance events when fat oxidation normally predominates.
OTHER NUTRIENT INTERVENTIONS
In practical terms, 2 hours or more of even moderate physical activity (eg, 65% VO2max) is sufficient to markedly lower muscle glycogen stores. At least 24 hours of rest and consumption of a high-carbohydrate diet (10 g/kg BW/d) are required to fully restore muscle glycogen concentration. This relatively slow time course makes it impossible for those engaged in multiple bouts of intense exercise during a single day to fully restore muscle glycogen between training sessions or competitive efforts. However, it is possible to maximize the rate of short-term muscle glycogen repletion so that athletes can replenish more muscle glycogen than might otherwise be possible.
Consuming proteins with carbohydrates may be beneficial in stimulating rapid glycogenesis in the hours immediately following exercise,65 a finding that has implications for speeding recovery between demanding bouts of exercise within the same day. A greater glycogen storage rate may be due to increased muscle glucose uptake and enhanced signaling pathways made possible by the influx of amino acids. Protein consumption also induces a rise in blood insulin concentration that augments the insulinemic response to carbohydrate ingestion, increasing the rate of glycogen repletion.96 However, when ample carbohydrates (>1.0 g/kg BW/h) are ingested, the addition of proteins does not further enhance glycogenesis.64,126,127 From a practical standpoint, protein is an important nutrient during recovery from exercise because it stimulates the muscle protein synthesis required for muscle recovery, repair, and growth. Consumption of 0.3–0.4 g protein/kg BW soon after exercise has been shown to optimize muscle protein synthesis.64,128 There are no negative implications associated with consuming a mixture of carbohydrates and proteins; many snacks and sports foods contain a sufficient amount of carbohydrates to stimulate muscle glycogen synthesis. It is clear that adequate consumption of proteins stimulates muscle protein synthesis during rest,129 although consuming proteins during exercise does not appear to benefit performance or immune function or reduce muscle damage.130,131
Multiday supplementation with creatine monohydrate along with an adequate amount of carbohydrates has been reported to increase muscle glycogen synthesis compared with carbohydrate ingestion alone.66,67,132 Other interventions, including consumption of large doses of caffeine133,134 and postexercise heat and cold therapy, have produced equivocal results in stimulating glycogenesis.135–137
EFFECTS OF SEX AND AGE
Males and females appear to restore muscle glycogen at similar rates following exercise, as long as sufficient carbohydrates and energy are consumed.98 In older adults, regular exercise training increases the GLUT4 and glycogen content of skeletal muscle, responses similar to those seen in younger adults; however, resting muscle glycogen does not seem to increase to levels seen in younger adults.138,139
Among masters athletes (those aged >55 y), Doering et al.140 reported that, although exercise-induced muscle damage is similar to that seen in younger athletes, the rate of muscle repair is slower in older adults,141 suggesting that the restoration of muscle glycogen may also be slower. As a result, older athletes are advised to consume 35–40 g of high-quality proteins in addition to sufficient carbohydrate to maximally stimulate muscle protein synthesis after exercise. The additional protein intake might also help facilitate glycogen synthesis, especially when carbohydrate intake is low.65
HIGH-QUALITY CARBOHYDRATE FOODS
Although there is no universally accepted definition of a high-quality carbohydrate food, for the purpose of advising athletes, unprocessed or minimally processed vegetables, grains, and fruits are examples of foods that provide ample amounts of carbohydrates per serving (eg, >20 g/serving), along with important micronutrients and “gap nutrients,” those nutrients that are commonly underconsumed in a typical American diet. The Dietary Guidelines for Americans142 identifies gap nutrients as dietary fiber, choline, potassium, magnesium, calcium, and vitamins A, D, E, and C. For example, the recommended intake for dietary fiber is 25 g/day for women and 38 g/day for men, yet the average US intake is 14–19 g/day.143 Iron is often underconsumed by adolescent girls and adult women, ages 19–50 years.144 For athletes, nutrients of concern reflect those of the general public: iron, vitamin D, calcium, and antioxidant nutrients, including vitamins C and E.144Table 4 identifies examples of high-quality carbohydrate foods.
Table 4.
High-quality carbohydrate foods
| Food | Portion | Energy | CHO (g) | PRO (g) | Fat (g) | Fiber (g) | Calcium (mg) | Vitamin C (mg) | Iron (mg) |
|---|---|---|---|---|---|---|---|---|---|
| Baked potato with skin | Medium, 2¼–3¼″ diameter | 161 | 36 | 4.3 | 0.2 | 3.8 | 26 | 16 | 1.9 |
| Black beans, canned, drained | 1 cup | 218 | 40 | 14.7 | 0.7 | 16.6 | 84 | 6.5 | 4.5 |
| Enriched penne pasta, boiled | 1 cup | 169 | 33 | 6.2 | 1.0 | 1.9 | 7 | 0 | 1.4 |
| Enriched wheat bran cereal with raisins | 1 cup | 191 | 46 | 4.5 | 0.9 | 8.1 | 26 | 0.5 | 10.8 |
| Whole wheat bread | 1 slice | 81 | 14 | 4 | 1.1 | 1.9 | 52 | 0 | 0.8 |
Data are from the US Department of Agriculture.144
Abbreviations: CHO, carbohydrate; PRO, protein.
HELPING ATHLETES WITH FOOD SELECTION
Nutrient-rich foods that are high in carbohydrates include grains (cereal, rice, pasta, breads, etc), most fruits, some vegetables (especially starch vegetables such as potatoes, beans, and peas), and dairy foods. Fruit and dairy foods contain simple sugars yet are rich in key nutrients. Fruit, especially whole fruit, is a good source of dietary fiber, vitamins, minerals, and water. Dairy foods, such as milk, are a good source of calcium, vitamin D, and potassium. The nutrition facts panel on packaged food can steer athletes to high-quality carbohydrate foods. The nutrition facts panel contains 2 pieces of information that athletes can readily use to identify carbohydrate content: the serving size and the grams of carbohydrates per serving. For example, 1.0–1.2 g carbohydrate/kg BW/hour after exercise stimulates the highest rate of glycogen synthesis and is an important strategy for athletes involved in competition requiring many trials or bouts in a single day. A 70-kg athlete should aim for 70–84 g carbohydrate/hour to rapidly restore glycogen. Using the nutrition facts panel, a serving of pita chips is about 7 chips and contains 19 g of carbohydrates. A serving of dried cherries is ¼ cup and containing 32 g of carbohydrates. Therefore, 2 servings of pita chips and 1 serving of dried cherries provides 70 g of carbohydrates and would be an appropriate snack for an athlete between bouts of training or competition. Currently, the nutrition facts panel does not distinguish naturally occurring sugar from added sugar, but proposed changes to the nutrition label will eventually separate the 2 sugars on product labels. This is important because the current nutrition label identifies milk as containing 12 grams of sugar per cup, but this sugar is the naturally occurring milk sugar, lactose, not sugar added in processing.
HELPING ATHLETES WITH PORTION SIZE
Identifying portion sizes can be a challenge for many athletes. Frequently, portion sizes are identified in ounces (or grams) or common household measures, such as cups, but many athletes are unfamiliar with the translation of those units to what constitutes a single serving. Using common household objects as a benchmark can help athletes understand serving sizes. A useful guide can be found at Web MD,145 and athletes can use these tools to help them easily visualize a serving.
For example, 1 cup of raw or cooked vegetables, 1 cup of popcorn, or a medium piece of fruit is similar to the size of a baseball; 3 ounces of meat, fish, or poultry is the size of a deck of cards; ¼ cup of dried fruit or nuts is the size of a large egg; 2 tablespoons of peanut butter (1 ounce) has the proportions of a golf ball; and a 3-ounce muffin or biscuit is of similar dimensions to a hockey puck.
Achieving a high-carbohydrate diet
As identified in Table 3, athletes who exercise very hard every day or perform very prolonged exercise have a high requirement for dietary carbohydrates. It can be daunting to consume the amount of food needed to achieve 8–12 g carbohydrate/kg BW/day, but identification of how much carbohydrate is in an athlete’s usual diet and how much carbohydrate is in an athlete’s favorite foods can help athletes understand what foods and beverages can contribute to meeting carbohydrate goals. There are also many high-carbohydrate drinks, gels, bars, and shakes that can be consumed to boost carbohydrate intake. Table 5 shows an athlete’s usual dietary intake, along with suggestions to increase carbohydrate consumption.
Table 5.
Example of eating program for a 70-kilogram athlete training for an ultra-endurance event, requiring 8–12 grams of carbohydrate per kilogram of body weight per day
| Usual intake | Total grams of carbohydrate | Additional foods to boost carbohydrate | Total grams of carbohydrate |
|---|---|---|---|
| Breakfast | |||
| Fast-food egg and cheese biscuit (32 g) and coffee (0 g) | 32 |
|
124 |
| Morning snack | |||
|
40 |
|
145 |
| Lunch | |||
|
10 |
|
112 |
| During workouts | |||
|
15 | 16-oz carbohydrate energy drink (30 g) | 45 g |
| Post workout | |||
|
38 |
|
100 g |
| Dinner | |||
|
76 |
|
198 |
| Bedtime snack | |||
|
20 |
|
84 |
| Total | 231 g or 3.3 g/kg BW | Total | 808 g or 11.5 g/kg BW |
Data from National Nutrient Database for Standard Reference (Release September 25, 2015).144
Abbreviations: BW, body weight.
PRACTICAL TIPS TO OPTIMIZE RESTORATION OF GLYCOGEN STORES
Athletes should pack snacks to consume between training sessions and not rely on coaches or the venue to provide quality snacks. Several sports foods (carbohydrate chews, bars, gels) are available in a variety of flavors and can be convenient for athletes. However, athletes are advised to try sports foods during training to know which foods they tolerate and enjoy.
Examples of carbohydrate foods include energy chews and gels that typically contain 25–30 g of carbohydrates per serving. Energy/sports bars often contain 25–45 g of carbohydrates per bar. Trail mix (nuts and dried fruit) provides 30–40 g of carbohydrates per ½ cup serving. Two fig-bar cookies contain 24 g of carbohydrates, and a peanut-butter-and-jelly sandwich typically contains 35–40 g of carbohydrates.
CONCLUSION
A high-carbohydrate diet remains the evidence-based recommendation for athletes who engage in hours of physical activity on a daily basis. To maintain muscle glycogen stores, athletes are advised to consume a high-carbohydrate diet that contains adequate energy (calories), along with proteins to stimulate muscle repair and growth and fluids to ensure normal hydration. Glycogen supercompensation results from a combination of ample rest, reduced training volume, and the consumption of a high-carbohydrate diet.
Consumption of a variety of carbohydrate foods ensures adequate muscle and liver glycogen restoration between bouts of physical activity. High-quality, nutrient-rich carbohydrates, such as potatoes, pastas, breads, vegetables, and fruits, provide concentrated amounts of simple (mono- and disaccharides, maltodextrins) and complex (starches and fibers) carbohydrates along with an array of micronutrients, such as vitamins, minerals, and other nutrient compounds.
To allow for sufficient muscle glycogen restoration between training sessions and overnight, athletes should consume enough carbohydrates to replace all or at least a substantial amount of the glucose oxidized during the day. Glucose oxidation can vary widely from day to day, so carbohydrate and energy intake should also vary—higher on days of hard training, lower on days of easier training. Depending upon training intensity and duration, carbohydrate intake might vary from 3 to 10 g/kg BW/day (1.4–4.5 g/lb BW/d).
After hard exercise, nutritious, carbohydrate-rich foods such as potatoes, pastas, grains, vegetables, and fruits are important sources of carbohydrates that can be quickly digested, absorbed, and transported in the blood and taken up by muscles and liver for the restoration of glycogen stores. The consumption of high-glycemic carbohydrates soon after exercise can maximize and sustain the rate of glycogen synthesis to help speed glycogen restoration on days that require > 1 session of hard exercise. When rapid glycogen resynthesis is required, consuming 0.5–0.6 g/kg BW of high-glycemic carbohydrates every 30 minutes (0.23–0.28 g/lb BW/30 min) for 2–4 hours (or until the next full meal) will sustain high rates of muscle glycogen synthesis. When 24 hours or more are available for glycogen restoration, the frequency of carbohydrate intake is less important than the total amounts of carbohydrates and energy consumed.
Techniques such as training with high muscle glycogen stores but sleeping and then training the next morning with low muscle glycogen stores have been shown in some studies to enhance glycogen storage and performance. However, more research is needed to confirm the consistency and magnitude of these responses.
For anyone engaged in regular physical activity (eg, laborers and soldiers) or sports training, adequate restoration of muscle and liver glycogen stores from day to day is required to sustain the capacity for continued exercise and high-intensity activity. If glycogen stores in muscle reach a critically low level, exercise intensity markedly decreases and performance is impaired. If liver glycogen stores are compromised, blood glucose declines, creating a hypoglycemic state that impairs both physical and mental function. Consumption of a variety of foods containing carbohydrates is needed to replenish all or a substantial portion of the glycogen that is oxidized during physical activity. Athletes, laborers, and soldiers are well advised to consume diets containing a variety of foods rich in carbohydrates and other nutrients and to begin consuming such foods as soon as possible following glycogen-depleting exercise or activity.
Acknowledgments
Author contributions.
B.M. conducted the literature review and authored content related to the scientific aspects of glycogen metabolism. C.R. contributed text and table content to this paper related to the practical aspects of nutritional guidance.
Funding.
B.M. received a stipend from the Alliance for Potato Research and Education to conduct a literature review on glycogen metabolism.
Declaration of interest.
B.M. served as a scientific consultant to the Alliance for Potato Research and Education. Both authors are members of The Clif Bar Company Nutrition Advisory Council.
References
- 1. Hawley JA, Leckey JJ.. Carbohydrate dependence during prolonged, intense endurance exercise. Sports Med. 2015;45(suppl 1):S5–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Institute of Medicine. Dietary carbohydrates, sugars and starches. In: Dietary Reference Intakes for Energy, Carbohydrates, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids. Washington, DC: National Academies Press; 2005:265–338. [Google Scholar]
- 3. Thomas TD, Erdman KA, Burke LM.. Nutrition and athletic performance. Med Sci Sports Exerc. 2016;48:543–568.http://dx.doi.org/10.1249/MSS.0000000000000852 [DOI] [PubMed] [Google Scholar]
- 4. Burke LM, van Loon LJ, Hawley JA.. Post-exercise muscle glycogen resynthesis in humans. J Appl Physiol. 2017;122:1055–1067.http://dx.doi.org/10.1152/japplphysiol.00860.2016 [DOI] [PubMed] [Google Scholar]
- 5. Anderson L, Orme P, Naughton RJ et al. , Energy intake and expenditure of professional soccer players of the English Premier League: evidence of carbohydrate periodization. Int J Sports Nutr Exerc Metab. 2017;27:228–238.http://dx.doi.org/10.1123/ijsnem.2016-0259 [DOI] [PubMed] [Google Scholar]
- 6. Devlin BL, Leveritt MD, Kingsley M et al. , Dietary intake, body composition and nutrition knowledge of Australian football and soccer players: implications for sports nutrition professionals in practice. Int J Sports Nutr Exerc Metab. 2017;27:130–138.http://dx.doi.org/10.1123/ijsnem.2016-0191 [DOI] [PubMed] [Google Scholar]
- 7. Mullins VA, Houtkooper LB, Howell WH et al. , Nutritional status of US elite female heptathletes during training. Int J Sport Nutr Exer Metab. 2001;11:299–314.http://dx.doi.org/10.1123/ijsnem.11.3.299 [DOI] [PubMed] [Google Scholar]
- 8. Birkenhead KL, Slater G.. A review of factors influencing athletes’ food choices. Sports Med. 2015;45:1511–1522. [DOI] [PubMed] [Google Scholar]
- 9. Trakman GL, Forsyth A, Devlin BL et al. , A systematic review of athletes’ and coaches’ nutrition knowledge and reflections on the quality of current nutrition knowledge measures. Nutrients. 2016;8:1–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Depre C, Vanoverschelde JJ, Taegtmeyer H.. Glucose for the heart. Circulation. 1999;99:578–588.http://dx.doi.org/10.1161/01.CIR.99.4.578 [DOI] [PubMed] [Google Scholar]
- 11. Guyton AC, Hall JE.. Guyton and Hall Textbook of Medical Physiology. New York: Saunders/Elsevier; 2011. [Google Scholar]
- 12. Adeva-Andany MM, Gonzalez-Lucan M, Donapetry-Garcia C et al. , Glycogen metabolism in humans. BBA Clin. 2016;5:85–100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Philp A, Hargreaves M, Baar K.. More than a store: regulatory roles for glycogen in skeletal muscle adaptation to exercise. Am J Physiol Endocrin Metab. 2012;302:E1343–E1351. [DOI] [PubMed] [Google Scholar]
- 14. Fernandez-Elias VE, Ortega JF, Nelson RK et al. , Relationship between muscle water and glycogen recovery after prolonged exercise in the heat in humans. Eur J Appl Physiol. 2015;115:1919–1926.http://dx.doi.org/10.1007/s00421-015-3175-z [DOI] [PubMed] [Google Scholar]
- 15. Kreitzman SN, Coxon AY, Szaz KF.. Glycogen storage: illusions of easy weight loss, excessive weight regain, and distortions in estimates of body composition. Am J Clin Nutr. 1992;56(1 suppl):292S–293S. [DOI] [PubMed] [Google Scholar]
- 16. Meyer RA, Wiseman RW.. The metabolic systems: control of ATP synthesis in skeletal muscle. In: Farrell PA, Joyner MJ, Caiozzo VJ, ed. ACSM’s Advanced Exercise Physiology. 2nd ed.Philadelphia, PA: Wolters Kluwer; 2012:363–378. [Google Scholar]
- 17. Jensen TE, Richter EA.. Regulation of glucose and glycogen metabolism during and after exercise. J Physiol (London). 2012;590:1069–1076.http://dx.doi.org/10.1113/jphysiol.2011.224972 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Ceperuelo-Mallafre V, Ejarque M, Serena C et al. , Adipose tissue glycogen accumulation is associated with obesity-linked inflammation in humans. Molec Metab. 2016;5:5–18.http://dx.doi.org/10.1016/j.molmet.2015.10.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Brown AM, Ransom BR.. Astrocyte glycogen and brain energy metabolism. Glia. 2007;55:1263–1271.http://dx.doi.org/10.1002/glia.20557 [DOI] [PubMed] [Google Scholar]
- 20. Wasserman DH. Four grams of glucose. Am J Physiol Endocrinol Metab. 2009;296:E11–E21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Bosch AN, Weltan SM, Dennis SC et al. , Fuel substrate turnover and oxidation and glycogen sparing with carbohydrate ingestion in non-carbohydrate-loaded cyclists. Pflügers Arch. 1996;432:1003–1010. [DOI] [PubMed] [Google Scholar]
- 22. Gonzalez JT, Fuchs CJ, Betts JA et al. , Liver glycogen metabolism during and after prolonged endurance-type exercise. Am J Physiol Endo Metab. 2016;311:E543–E553. [DOI] [PubMed] [Google Scholar]
- 23. De Bock K, Derave W, Ramaekers M et al. , Fiber type-specific muscle glycogen sparing due to carbohyrate intake befor and during exercise. J Appl Physiol. 2007;102:183–188.http://dx.doi.org/10.1152/japplphysiol.00799.2006 [DOI] [PubMed] [Google Scholar]
- 24. Fryer KL, Brown AM.. Pluralistic roles for glycogen in the central and peripheral nervous systems. Metab Brain Dis. 2015;30:299–306.http://dx.doi.org/10.1007/s11011-014-9516-5 [DOI] [PubMed] [Google Scholar]
- 25. Obel LF, Muller MS, Walls AB et al. , Brain glycogen-new perspectives on its metabolic function and regulation at the subcellular level. Front Neuroenerg. 2012;4:1–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Chambers TW, Daly TP, Hockley A et al. , Contribution of glycogen in supporting axon conduction in the peripheral and central nervous systems: the role of lactate. Front Neurosci. 2014;8:1–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Bergersen LH. Is lactate food for neurons? Comparison of monocarboxylate transporter subtypes in brain and muscle. Neuroscience. 2007;145:11–19.http://dx.doi.org/10.1016/j.neuroscience.2006.11.062 [DOI] [PubMed] [Google Scholar]
- 28. Duran J, Guinovart JJ.. Brain glycogen in health and disease. Molec Aspects Med. 2015;46:70–77.http://dx.doi.org/10.1016/j.mam.2015.08.007 [DOI] [PubMed] [Google Scholar]
- 29. Krogh A, Lindhard J.. The relative value of fat and carbohydrate as sources of muscular energy: with appendices on the correlation between standard metabolism and the respiratory quotient during rest and work. Biochem J. 1920;14:290–363.http://dx.doi.org/10.1042/bj0140290 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Levine SA, Gordon B, Derick CL.. Some changes in the chemical constituents of the blood following a marathon—with especial reference to the development of hypoglycemia. JAMA. 1924;82:1778–1779. [Google Scholar]
- 31. Gordon B, Kohn LA, Levine SA et al. , Sugar content of the blood in runners following a marathon race—especial reference to the prevention of hypoglycemia. JAMA. 1925;83:508–509. [Google Scholar]
- 32. Bergstrom J, Hultman E.. Muscle glycogen synthesis after exercise: an enhancing factor localized to the muscle cells in man. Nature. 1966;210:309–310.http://dx.doi.org/10.1038/210309a0 [DOI] [PubMed] [Google Scholar]
- 33. Bergstrom J, Hermansen L, Hultman E et al. , Diet, muscle glycogen and physical performance. Acta Physiol Scand. 1967;71:140–150.http://dx.doi.org/10.1111/j.1748-1716.1967.tb03720.x [DOI] [PubMed] [Google Scholar]
- 34. Hermansen L, Hultman E, Saltin B.. Muscle glycogen during prolonged severe exercise. Acta Physiol Scand. 1967;71:129–139.http://dx.doi.org/10.1111/j.1748-1716.1967.tb03719.x [DOI] [PubMed] [Google Scholar]
- 35. Hultman E, Bergstrom J.. Muscle glycogen synthesis in relation to diet studied in normal subjects. Acta Med Scand. 1967;182:109–117. [DOI] [PubMed] [Google Scholar]
- 36. Murray R, Sugar, sports drinks, and performance. In: Rippe JM, ed. Fructose, High Fructose Corn Syrup, Sucrose and Health. New York: Humana Press; 2014:293–305. [Google Scholar]
- 37. Burke LM, Maughan RJ.. The governor has a sweet tooth—mouth sensing of nutrients to enhance sports performance. Eur J Sports Sci. 2015;15:29–40. [DOI] [PubMed] [Google Scholar]
- 38. Carter J, Jeukendrup A, Jones D.. The effect of carbohydrate mouth rinse on 1-h cycle time trial performance. Med Sci Sports Exerc. 2004;36:2107–2111. [DOI] [PubMed] [Google Scholar]
- 39. Clarke ND, Hammond S, Kornilios E et al. , Carbohydrate mouth rinse improves morning high-intensity exercise performance. Eur J Sports Sci. 2017;17:955–963.http://dx.doi.org/10.1080/17461391.2017.1333159 [DOI] [PubMed] [Google Scholar]
- 40. Hargreaves M. The metabolic systems: carbohydrate metabolism. In: Farrell P, ed. ACSM's Advanced Exercise Physiology. 2nd ed.Philadelphia: Lippincott Williams & Wilkins; 2012:379–391. [Google Scholar]
- 41. Ivy J. The regulation and synthesis of muscle glycogen by means of nutrient intervention. In: Maughan R, ed. Sports Nutrition. Vol. 19 West Sussex, UK: Wiley Blackwell; 2014:113–125. [Google Scholar]
- 42. Smythe C, Cohen P.. The discovery of glycogenin and the priming mechanism for glycogen biogenesis. Eur J Biochem. 1991;200:625–631.http://dx.doi.org/10.1111/j.1432-1033.1991.tb16225.x [DOI] [PubMed] [Google Scholar]
- 43. Shearer J, Wilson RJ, Battram DS et al. , Increases in glycogenin and glycogenin mRNA accompany glycogen resynthesis in human skeletal muscle. Am J Physiol Endocrin Metab. 2005;289:E508–E514. [DOI] [PubMed] [Google Scholar]
- 44. Shearer J, Graham TE, Battram DS et al. , Glycogenin activity and mRNA expression in response to volitional exhaustion in human skeletal muscle. J Appl Physiol. 2005;99:957–962.http://dx.doi.org/10.1152/japplphysiol.00275.2005 [DOI] [PubMed] [Google Scholar]
- 45. Lomako J, Lomako WM, Whelan WJ.. The biogenesis of glycogen: nature of the carbohydrate in the protein primer. Biochem Int. 1990;21:251–260. [PubMed] [Google Scholar]
- 46. Lomako J, Lomako WM, Whelan WJ.. The nature of the primer for glycogen synthesis in muscle. FEBS Lett. 1990;268:8–12.http://dx.doi.org/10.1016/0014-5793(90)80959-M [DOI] [PubMed] [Google Scholar]
- 47. Lomako J, Lomako WM, Whelan WJ.. Proglycogen: a low-molecular-weight form of muscle glycogen. FEBS Lett. 1991;279:223–228.http://dx.doi.org/10.1016/0014-5793(91)80154-U [DOI] [PubMed] [Google Scholar]
- 48. Schweitzer GG, Kearney ML, Mittendorfer B.. Muscle glycogen: where did you come from, where did you go? J Physiol. 2017;595:2771–2772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Nielsen J, Ortenblad N.. Physiological aspects of the subcellular localization of glycogen in skeletal muscle. Appl Physiol Nutr Metab. 2013;38:91–99.http://dx.doi.org/10.1139/apnm-2012-0184 [DOI] [PubMed] [Google Scholar]
- 50. Gejl KD, Ortenblad N, Andersson E et al. , Local depletion of glycogen with supramaximal exercise in human skeletal muscle fibres. J Physiol (London). 2017;595:2809–2821.http://dx.doi.org/10.1113/JP273109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Nielsen J, Holmberg HC, Schroder HD et al. , Human skeletal muscle glycogen utilization in exhaustive exercise: role of subcellular localization and fibre type. J Physiol. 2011;589(pt 11):2871–2885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Ortenblad N, Nielsen J, Saltin B, Holmberg HC.. Role of glycogen availability in sarcoplasmic reticulum Ca2+ kinetics in human skeletal muscle. J Physiol. 2011;589(pt 3):711–725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Knuiman P, Hopman MT, Mensink M.. Glycogen availability and skeletal muscle adaptations with endurance and resistance exercise. Nutr Metab (London). 2015;12:59.http://dx.doi.org/10.1186/s12986-015-0055-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Ahlborg B, Brohult J.. Immediate and delayed metabolic reactions in well-trained subjects after prolonged physical exercise. Acta Med Scand. 1967;182:41–54. [DOI] [PubMed] [Google Scholar]
- 55. Bergstrom J, Hultman E, Roch-Norlund AE.. Muscle glycogen synthetase in normal subjects. Basal values, effect of glycogen depletion by exercise and of a carbohydrate-rich diet following exercise. Scand J Clin Lab Invest. 1972;29:231–236. [DOI] [PubMed] [Google Scholar]
- 56. Sherman WM, Costill DL, Fink WJ et al. , Effect of exercise-diet manipulation on muscle glycogen and its subsequent utilization during performance. Int J Sports Med. 1981;2:114–118.http://dx.doi.org/10.1055/s-2008-1034594 [DOI] [PubMed] [Google Scholar]
- 57. Bussau VA, Fairchild TJ, Rao A et al. , Carbohydrate loading in human muscle: an improved 1 day protocol. Eur J Appl Physiol. 2002;87:290–295.http://dx.doi.org/10.1007/s00421-002-0621-5 [DOI] [PubMed] [Google Scholar]
- 58. Bartlett JD, Hawley JA, Morton JP.. Carbohydrate availability and exercise training adaptation: too much of a good thing?. Eur J Sports Sci. 2015;15:3–12.http://dx.doi.org/10.1080/17461391.2014.920926 [DOI] [PubMed] [Google Scholar]
- 59. Burke LM. Fueling strategies to optimize performance: training high or training low? Scand J Med Sci Sports. 2010;20(suppl 2):48–58. [DOI] [PubMed] [Google Scholar]
- 60. Hawley J. Manipulating carbohydrate availability to promote training adaptation. Sports Sci Exchange. 2014;27:1–7. [Google Scholar]
- 61. Marquet LA, Brisswalter J, Louis J et al. , Enhanced endurance performance by periodization of carbohydrate intake: “sleep low” strategy. Med Sci Sports Exerc. 2016;48:663–672. [DOI] [PubMed] [Google Scholar]
- 62. Marquet LA, Hausswirth C, Molle O et al. , Periodization of carbohydrate intake: short-term effect on performance. Nutrients. 2016;8:2–13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Yeo WK, Paton CD, Garnham AP et al. , Skeletal muscle adaptation and performance responses to once a day versus twice every second day endurance training regimens. J Appl Physiol. 2008;105:1462–1470.http://dx.doi.org/10.1152/japplphysiol.90882.2008 [DOI] [PubMed] [Google Scholar]
- 64. Betts JA, Williams C.. Short-term recovery from prolonged exercise: exploring the potential for protein ingestion to accentuate the benefits of carbohydrate supplements. Sports Med. 2010;40:941–959.http://dx.doi.org/10.2165/11536900-000000000-00000 [DOI] [PubMed] [Google Scholar]
- 65. Ivy JL, Goforth HW Jr, Damon BM et al. , Early postexercise muscle glycogen recovery is enhanced with a carbohydrate-protein supplement. J Appl Physiol. 2002;93:1337–1344.http://dx.doi.org/10.1152/japplphysiol.00394.2002 [DOI] [PubMed] [Google Scholar]
- 66. Roberts PA, Fox J, Peirce N et al. , Creatine ingestion augments dietary carbohydrate mediated muscle glycogen supercompensation during the initial 24 h of recovery following prolonged exhaustive exercise in humans. Amino Acids. 2016;48:1831–1842.http://dx.doi.org/10.1007/s00726-016-2252-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Sewell DA, Robinson TM, Greenhaff PL.. Creatine supplementation does not affect human skeletal muscle glycogen content in the absence of prior exercise. J Appl Physiol. 2008;104:508–512.http://dx.doi.org/10.1152/japplphysiol.00787.2007 [DOI] [PubMed] [Google Scholar]
- 68. Burke LM, Kiens B.. “Fat adaptation” for athletic performance: the nail in the coffin? J Appl Physiol. 2006;100:7–8. [DOI] [PubMed] [Google Scholar]
- 69. Zachwieja JJ, Costill DL, Pascoe DD et al. , Influence of muscle glycogen depletion on the rate of resynthesis. Med Sci Sports Exerc. 1991;23:44–48. [PubMed] [Google Scholar]
- 70. McInerney P, Lessard SJ, Burke LM et al. , Failure to repeatedly supercompensate muscle glycogen stores in highly trained men. Med Sci Sports Exerc. 2005;37:404–411.http://dx.doi.org/10.1249/01.MSS.0000155699.51360.2F [DOI] [PubMed] [Google Scholar]
- 71. Hermansen L, Vaage O.. Lactate disappearance and glycogen synthesis in human muscle after maximal exercise. Am J Physiol. 1977;233:E422–E429. [DOI] [PubMed] [Google Scholar]
- 72. Mul JD, Stanford KI, Hirshman MF et al. , Exercise and regulation of carbohydrate metabolism. Prog Mol Biol Transl Sci. 2015;135:17–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Shulman RG, Rothman DL.. The “glycogen shunt” in exercising muscle: a role for glycogen in muscle energetics and fatigue. Proc Natl Acad Sci U S A. 2001;98:457–461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Krustrup P, Ortenblad N, Nielsen J et al. , Maximal voluntary contraction force, SR function and glycogen resynthesis during the first 72 h after a high-level competitive soccer game. Eur J Appl Physiol. 2011;111:2987–2995.http://dx.doi.org/10.1007/s00421-011-1919-y [DOI] [PubMed] [Google Scholar]
- 75. Costill DL, Sparks K, Gregor R et al. , Muscle glycogen utilization during exhaustive running. J Appl Physiol. 1971;31:353–356.http://dx.doi.org/10.1152/jappl.1971.31.3.353 [DOI] [PubMed] [Google Scholar]
- 76. Costill DL, Bowers R, Branam G et al. , Muscle glycogen utilization during prolonged exercise on successive days. J Appl Physiol. 1971;31:834–838.http://dx.doi.org/10.1152/jappl.1971.31.6.834 [DOI] [PubMed] [Google Scholar]
- 77. Sherman WM, Doyle JA, Lamb DR et al. , Dietary carbohydrate, muscle glycogen, and exercise performance during 7 d of training. Am J Clin Nutr. 1993;57:27–31.http://dx.doi.org/10.1093/ajcn/57.1.27 [DOI] [PubMed] [Google Scholar]
- 78. Cochran AJ, Myslik F, MacInnis MJ et al. , Manipulating carbohydrate availability between twice-daily sessions of high-intensity interval training over 2 weeks improves time-trial performance. Int J Sports Nutr Exerc Metab. 2015;25:463–470.http://dx.doi.org/10.1123/ijsnem.2014-0263 [DOI] [PubMed] [Google Scholar]
- 79. Hansen AK, Fischer CP, Plomgaard P et al. , Skeletal muscle adaptation: training twice every second day vs. training once daily. J Appl Physiol. 2005;98:93–99. [DOI] [PubMed] [Google Scholar]
- 80. Pascoe DD, Costill DL, Fink WJ et al. , Glycogen resynthesis in skeletal muscle following resistive exercise. Med Sci Sports Exerc. 1993;25:349–354. [PubMed] [Google Scholar]
- 81. Tesch PA, Colliander EB, Kaiser P.. Muscle metabolism during intense, heavy-resistance exercise. Eur J Appl Physiol Occup Physiol. 1986;55:362–366.http://dx.doi.org/10.1007/BF00422734 [DOI] [PubMed] [Google Scholar]
- 82. Camera DM, Hawley JA, Coffey VG.. Resistance exercise with low glycogen increases p53 phosphorylation and PGC-1alpha mRNA in skeletal muscle. Eur J Appl Physiol. 2015;115:1185–1194.http://dx.doi.org/10.1007/s00421-015-3116-x [DOI] [PubMed] [Google Scholar]
- 83. Escobar KA, VanDusseldorp TA, Kerksick CM.. Carbohydrate intake and resistance-based exercise: are current recommendations reflective of actual need? Br J Nutr. 2016;116:2053–2065. [DOI] [PubMed] [Google Scholar]
- 84. Coyle EF, Coggan AR, Hemmert MK et al. , Muscle glycogen utilization during prolonged strenuous exercise when fed carbohydrate. J Appl Physiol. 1986;61:165–172.http://dx.doi.org/10.1152/jappl.1986.61.1.165 [DOI] [PubMed] [Google Scholar]
- 85. Sherman WM, Wimer GS.. Insufficient dietary carbohydrate during training: does it impair athletic performance? Int J Sports Nutr. 1991;1:28–44. [DOI] [PubMed] [Google Scholar]
- 86. Helge JW. A high carbohydrate diet remains the evidence based choice for elite athletes to optimise performance. J Physiol (London). 2017;595:2775.http://dx.doi.org/10.1113/JP273830 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Coyle EF. Timing and method of increased carbohydrate intake to cope with heavy training, competition and recovery. J Sports Sci. 1991;9:29–51; discussion 51–22. [DOI] [PubMed] [Google Scholar]
- 88. Starling RD, Trappe TA, Parcell AC et al. , Effects of diet on muscle triglyceride and endurance performance. J Appl Physiol. 1997;82:1185–1189.http://dx.doi.org/10.1152/jappl.1997.82.4.1185 [DOI] [PubMed] [Google Scholar]
- 89. Burke LM, Collier GR, Hargreaves M.. Muscle glycogen storage after prolonged exercise: effect of the glycemic index of carbohydrate feedings. J Appl Physiol. 1993;75:1019–1023.http://dx.doi.org/10.1152/jappl.1993.75.2.1019 [DOI] [PubMed] [Google Scholar]
- 90. Burke LM, Cox GR, Cummings NK et al. , Guidelines for daily carbohydrate intake. Sports Med. 2001;31:267–299.http://dx.doi.org/10.2165/00007256-200131040-00003 [DOI] [PubMed] [Google Scholar]
- 91. Burke LM, Ross ML, Garvican-Lewis LA et al. , Low carbohydrate, high fat diet impairs exercise economy and negates the performance benefit from intensified training in elite race walkers. J Physiol. 2017;595:2785–2807.http://dx.doi.org/10.1113/JP273230 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Hawley JA, Hargreaves M, Zierath JR.. Signalling mechanisms in skeletal muscle: role in substrate selection and muscle adaptation. Essays Biochem. 2006;42:1–12.http://dx.doi.org/10.1042/bse0420001 [DOI] [PubMed] [Google Scholar]
- 93. Jeukendrup AE. Periodized nutrition for athletes. Sports Med. 2017;47(Suppl 1):51–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94. Richter EA, Derave W, Wojtaszewski JF.. Glucose, exercise and insulin: emerging concepts. J Physiol (London). 2001;535(pt 2):313–322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95. Mikines KJ, Farrell PA, Sonne B et al. , Postexercise dose-response relationship between plasma glucose and insulin secretion. J Appl Physiol. 1988;64:988–999.http://dx.doi.org/10.1152/jappl.1988.64.3.988 [DOI] [PubMed] [Google Scholar]
- 96. van Loon LJ, Saris WH, Kruijshoop M et al. , Maximizing postexercise muscle glycogen synthesis: carbohydrate supplementation and the application of amino acid or protein hydrolysate mixtures. Am J Clin Nutr. 2000;72:106–111. [DOI] [PubMed] [Google Scholar]
- 97. Burke LM, Hawley JA, Wong SH et al. , Carbohydrates for training and competition. J Sports Sci. 2011;29(suppl 1):S17–S27. [DOI] [PubMed] [Google Scholar]
- 98. Tarnopolsky MA, Zawada C, Richmond LB et al. , Gender differences in carbohydrate loading are related to energy intake. J Appl Physiol. 2001;91:225–230.http://dx.doi.org/10.1152/jappl.2001.91.1.225 [DOI] [PubMed] [Google Scholar]
- 99. Asp S, Daugaard JR, Richter EA.. Eccentric exercise decreases glucose transporter GLUT4 protein in human skeletal muscle. J Physiol. 1995;482(pt 3):705–712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100. Asp S, Daugaard JR, Kristiansen S et al. , Eccentric exercise decreases maximal insulin action in humans: muscle and systemic effects. J Physiol. 1996;494(pt 3):891–898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101. Blom PC, Costill DL, Vollestad NK.. Exhaustive running: inappropriate as a stimulus of muscle glycogen super-compensation. Med Sci Sports Exerc. 1987;19:398-403. [PubMed] [Google Scholar]
- 102. Costill DL, Pascoe DD, Fink WJ et al. , Impaired muscle glycogen resynthesis after eccentric exercise. J Appl Physiol. 1990;69:46–50.http://dx.doi.org/10.1152/jappl.1990.69.1.46 [DOI] [PubMed] [Google Scholar]
- 103. Widrick JJ, Costill DL, McConell GK et al. , Time course of glycogen accumulation after eccentric exercise. J Appl Physiol. 1992;72:1999–2004.http://dx.doi.org/10.1152/jappl.1992.72.5.1999 [DOI] [PubMed] [Google Scholar]
- 104. Blom PC, Hostmark AT, Vaage O et al. , Effect of different post-exercise sugar diets on the rate of muscle glycogen synthesis. Med Sci Sports Exerc. 1987;19:491–496. [PubMed] [Google Scholar]
- 105. Shi X, Passe DH.. Water and solute absorption from carbohydrate-electrolyte solutions in the human proximal small intestine: a review and statistical analysis. Int J Sport Nutr Exer Metab. 2010;20:427–442.http://dx.doi.org/10.1123/ijsnem.20.5.427 [DOI] [PubMed] [Google Scholar]
- 106. Baker LB, Jeukendrup AE.. Optimal composition of fluid-replacement beverages. Compr Physiol. 2014;4:575–620. [DOI] [PubMed] [Google Scholar]
- 107. Jentjens R, Achten J, Jeukendrup A.. High oxidation rates from combined carbohydrates ingested during exercise. Med Sci Sports Exerc. 2004;36:1551–1558.http://dx.doi.org/10.1249/01.MSS.0000139796.07843.1D [DOI] [PubMed] [Google Scholar]
- 108. Keizer HA, Kuipers H, van Kranenburg G et al. , Influence of liquid and solid meals on muscle glycogen resynthesis, plasma fuel hormone response, and maximal physical working capacity. Int J Sports Med. 1987;8:99–104.http://dx.doi.org/10.1055/s-2008-1025649 [DOI] [PubMed] [Google Scholar]
- 109. Reed MJ, Brozinick JT Jr, Lee MC et al. , Muscle glycogen storage postexercise: effect of mode of carbohydrate administration. J Appl Physiol. 1989;66:720–726.http://dx.doi.org/10.1152/jappl.1989.66.2.720 [DOI] [PubMed] [Google Scholar]
- 110. Cramer MJ, Dumke CL, Hailes WS et al. , Postexercise glycogen recovery and exerciseperformance is not significantly different between fast food and sport supplements. Int J Sport Nutr Exerc Metab. 2015;25:448–455.http://dx.doi.org/10.1123/ijsnem.2014-0230 [DOI] [PubMed] [Google Scholar]
- 111. Jenkins DJ, Wolever TM, Taylor RH et al. , Glycemic index of foods: a physiological basis for carbohydrate exchange. Am J Clin Nutr. 1981;34:362–366.http://dx.doi.org/10.1093/ajcn/34.3.362 [DOI] [PubMed] [Google Scholar]
- 112. Montain SJ, Hopper MK, Coggan AR et al. , Exercise metabolism at different time intervals after a meal. J Appl Physiol. 1991;70:882–888.http://dx.doi.org/10.1152/jappl.1991.70.2.882 [DOI] [PubMed] [Google Scholar]
- 113. Wu CL, Williams C.. A low glycemic index meal before exercise improves endurance running capacity in men. Int J Sports Nutr Exerc Metab. 2006;16:510–527.http://dx.doi.org/10.1123/ijsnem.16.5.510 [DOI] [PubMed] [Google Scholar]
- 114. Wee SL, Williams C, Tsintzas K et al. , Ingestion of a high-glycemic index meal increases muscle glycogen storage at rest but augments its utilization during subsequent exercise. J Appl Physiol. 2005;99:707–714.http://dx.doi.org/10.1152/japplphysiol.01261.2004 [DOI] [PubMed] [Google Scholar]
- 115. Little JP, Chilibeck PD, Ciona D et al. , The effects of low- and high-glycemic index foods on high-intensity intermittent exercise. Int J Sports Physiol Perf. 2009;4:367–380.http://dx.doi.org/10.1123/ijspp.4.3.367 [DOI] [PubMed] [Google Scholar]
- 116. Little JP, Chilibeck PD, Ciona D et al. , Effect of low- and high-glycemic-index meals on metabolism and performance during high-intensity, intermittent exercise. Int J Sports Nutr Exerc Metab. 2010;20:447–456.http://dx.doi.org/10.1123/ijsnem.20.6.447 [DOI] [PubMed] [Google Scholar]
- 117. Rauch LH, Rodger I, Wilson GR et al. , The effects of carbohydrate loading on muscle glycogen content and cycling performance. Int J Sports Nutr. 1995;5:25–36.http://dx.doi.org/10.1123/ijsn.5.1.25 [DOI] [PubMed] [Google Scholar]
- 118. Brown LJ, Midgley AW, Vince RV et al. , High versus low glycemic index 3-h recovery diets following glycogen-depleting exercise has no effect on subsequent 5-km cycling time trial performance. J Sci Med Sport. 2013;16:450–454.http://dx.doi.org/10.1016/j.jsams.2012.10.006 [DOI] [PubMed] [Google Scholar]
- 119. Goodpaster BH, Costill DL, Fink WJ et al. , The effects of pre-exercise starch ingestion on endurance performance. Int J Sports Med. 1996;17:366–372.http://dx.doi.org/10.1055/s-2007-972862 [DOI] [PubMed] [Google Scholar]
- 120. Gray BJ, Page R, Turner D et al. , Improved end-stage high-intensity performance but similar glycemic responses after waxy barley starch ingestion compared to dextrose in type 1 diabetes. J Sports Med Phys Fit. 2016;56:1392–1400. [PubMed] [Google Scholar]
- 121. Jozsi AC, Trappe TA, Starling RD et al. , The influence of starch structure on glycogen resynthesis and subsequent cycling performance. Int J Sports Med. 1996;17:373–378.http://dx.doi.org/10.1055/s-2007-972863 [DOI] [PubMed] [Google Scholar]
- 122. Vicente-Salar N, Urdampilleta Otegui A, Roche Collado E.. Endurance training in fasting conditions: biological adaptations and body weight management. Nutr Hosp. 2015;32:2409–2420. [DOI] [PubMed] [Google Scholar]
- 123. Cox PJ, Clarke K.. Acute nutritional ketosis: implications for exercise performance and metabolism. Extreme Physiol Med. 2014;3:1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Scott JM, Deuster PA.. Ketones and human performance. J Spec Oper Med. 2017;17:112–116. [DOI] [PubMed] [Google Scholar]
- 125. Vandoorne T, De Smet S, Ramaekers M et al. , Intake of a ketone ester drink during recovery from exercise promotes mTORC1 signaling but not glycogen resynthesis in human muscle. Front Physiol. 2017;8:1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126. Jentjens RL, van Loon LJ, Mann CH et al. , Addition of protein and amino acids to carbohydrates does not enhance postexercise muscle glycogen synthesis. J Appl Physiol. 2001;91:839–846.http://dx.doi.org/10.1152/jappl.2001.91.2.839 [DOI] [PubMed] [Google Scholar]
- 127. van Hall G, Shirreffs SM, Calbet JAL.. Muscle glycogen resynthesis during recovery from cycle exercise: no effect of additional protein ingestion. J Appl Physiol. 2000;88:1631–1636.http://dx.doi.org/10.1152/jappl.2000.88.5.1631 [DOI] [PubMed] [Google Scholar]
- 128. Morton RW, McGlory C, Phillips SM.. Nutritional interventions to augment resistance training-induced skeletal muscle hypertrophy. Front Physiol. 2015;6:245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129. Phillips SM, Chevalier S, Leidy HJ.. Protein “requirements” beyond the RDA: implications for optimizing health. Appl Physiol Nutr Metab. 2016;41:565–572. [DOI] [PubMed] [Google Scholar]
- 130. Breen L, Tipton KD, Jeukendrup AE.. No effect of carbohydrate-protein on cycling performance and indices of recovery. Med Sci Sports Exerc. 2010;42:1140–1148. [DOI] [PubMed] [Google Scholar]
- 131. Hansen M, Bangsbo J, Jensen J et al. , Protein intake during training sessions has no effect on performance and recovery during a strenuous training camp for elite cyclists. J Int Soc Sports Nutr. 2016;13:1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132. Robinson TM, Sewell DA, Hultman E et al. , Role of submaximal exercise in promoting creatine and glycogen accumulation in human skeletal muscle. J Appl Physiol. 1999;87:598–604.http://dx.doi.org/10.1152/jappl.1999.87.2.598 [DOI] [PubMed] [Google Scholar]
- 133. Beelen M, Burke LM, Gibala MJ et al. , Nutritional strategies to promote postexercise recovery. Int J Sports Nutr Exerc Metab. 2010;20:515–532.http://dx.doi.org/10.1123/ijsnem.20.6.515 [DOI] [PubMed] [Google Scholar]
- 134. Pedersen D, Lessard S, Coffey V et al. , High rates of muscle glycogen resynthesis after exhaustive exercise when carbohydrate is coingested with caffeine. J Appl Physiol. 2008;105:7–13.http://dx.doi.org/10.1152/japplphysiol.01121.2007 [DOI] [PubMed] [Google Scholar]
- 135. Gregson W, Allan R, Holden S et al. , Postexercise cold-water immersion does not attenuate muscle glycogen resynthesis. Med Sci Sports Exerc. 2013;45:1174–1181.http://dx.doi.org/10.1249/MSS.0b013e3182814462 [DOI] [PubMed] [Google Scholar]
- 136. Slivka D, Tucker T, Cuddy J et al. , Local heat application enhances glycogenesis. Appl Physiol Nutr Metab. 2012;37:247–251.http://dx.doi.org/10.1139/h11-157 [DOI] [PubMed] [Google Scholar]
- 137. Tucker TJ, Slivka DR, Cuddy JS et al. , Effect of local cold application on glycogen recovery. J Sports Med Phys Fitness. 2012;52:158–164. [PubMed] [Google Scholar]
- 138. Cartee GD. Influence of age on skeletal muscle glucose transport and glycogen metabolism. Med Sci Sports Exerc. 1994;26:577–585. [PubMed] [Google Scholar]
- 139. Cartee GD. Aging skeletal muscle: response to exercise. Exerc Sport Sci Rev. 1994;22:91–120. [PubMed] [Google Scholar]
- 140. Doering TM, Reaburn PR, Phillips SM et al. , Postexercise dietary protein strategies to maximize skeletal muscle repair and remodeling in masters endurance athletes: a review. Int J Sports Nutr Exerc Metab. 2016;26:168–178.http://dx.doi.org/10.1123/ijsnem.2015-0102 [DOI] [PubMed] [Google Scholar]
- 141. Brisswalter J, Nosaka K.. Neuromuscular factors associated with decline in long-distance running performance in master athletes. Sports Med. 2013;43:51–63.http://dx.doi.org/10.1007/s40279-012-0006-9 [DOI] [PubMed] [Google Scholar]
- 142. US Department of Health and Human Services; US Department of Agriculture. 2015–2020 Dietary Guidelines for Americans. 8th ed.Washington, DC: US Dept of Health and Human Services; 2015. http://www.health.gov/DietaryGuidelines. Accessed February 20, 2017. [Google Scholar]
- 143. Quagliani D, Felt-Gunderson P.. Closing America’s fiber intake gap: communication strategies from a food and fiber summit. Am J Lifestyle Med. 2017;11:80–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144. US Department of Agriculture. National Nutrition Database. https://ndb.nal.usda.gov/ndb/. Accessed May 28, 2017. [Google Scholar]
- 145. Zelman K. Web MD Portion Size Guide. http://img.webmd.com/dtmcms/live/webmd/consumer_assets/site_images/media/pdf/diet/portion-control-guide.pdf. Accessed February 20, 2017. [Google Scholar]