

Abstract
Cancer stem cells (CSCs), which have a close connection with tumor microenvironment, play a pivotal role in tumorigenesis, tumor progression, and metastasis. The inflammatory microenvironment is an essential component of tumor microenvironment. In the recent years, many studies have demonstrated that the inflammatory microenvironment induces the initiation of tumors, and contributes to the process of the progression of tumors, as well as metastasis. In this review, we summarize the relationship between CSCs and inflammatory components, such as inflammatory cytokines (IFNs, TNF, IL-6, IL-17) and inflammatory cells (myeloid-derived suppressor cells, tumor-associated macrophages). To illuminate the key factors that exert important actions in the tumor process would be important to improve the clinical outcome of the treatment for different types of cancer.
Keywords: tumor microenvironment, inflammatory factors, cancer stem cells, interferons, tumor necrosis factor, myeloid-derived suppressor cells, tumor-associated macrophages
1. Introduction
Cancer stem cells (CSCs), which was first reported by Dick and his colleagues in acute myeloid leukemia (AML) (1), have been identified in many solid tumors. It is extensively accepted the importance of CSCs in tumorigenesis, tumor progression, metastasis and relapse. CSCs are capable of preserving cancer heterogeneity through reserving self-renewal and differentiation abilities. In addition, CSCs activate the resistance mechanisms, including downregulation of replication, expression of drug export systems, epithelial-to-mesenchymal transition (EMT) and enhanced resistance to hypoxia with induction of angiogenesis and immune escape by reducing tumor specific antigens, while increasing anti-inflammatory cytokines and growth factors (2). The positioning of stem cells in normal organs has been identified, and it has been suggested that the interaction of microenvironment with stem cells is pivotal for the maintenance of stemness (3). CSCs may localize at specific niche like stem cells in normal organs (4). Due to the significance of tumor microenvironment, it is a reasonable strategy to regulate the stem cell niche to inhibit CSC survival. Among the multiple causes of malignant tumor, infection has been considered as a major navigator of inflammation-induced tumorigenesis (5). It has been accepted that the inflammatory microenvironment is an essential component of CSCs niche. Cancer can be promoted by inflammation, via enhancing proliferative and survival signaling, induction of invasion and metastasis. In this review, we summarize the major molecular and cellular mechanisms that participate in the reciprocal action between CSCs and inflammatory mediators (Figs. 1 and 2; Table I). The emerging mechanisms of inflammatory factors-induced stemness of CSCs are outlined. A better understanding of the role of the crosstalk between CSCs and inflammatory microenvironment might prospectively lead to more efficient therapies against cancer initiation and progression.
Figure 1.

IFN-γ, by binding to the IFNAR on CSCs, lead to the activation of NFκB, p38/MAPK and STAT pathways. After integrated with the TNFR, TNF-α increases the stemness property and enhance the proliferation of CSCs. IL-6 acts on the IL-6 receptor expressed by CSCs and activates JAK/STAT, PI3K/Akt pathways, then to facilitate the self-renewal of stem-like cells and enhance their tumorigenic potential. The JAK/STAT, NFκB and p38/MAPK pathways also could be activated by IL-17 and IL-17R. CSCs, cancer stem cells; IFN, interferon; JAK, Janus kinase; STAT, signal transducer and activator of transcription; p38, tumor protein 38; MAPK, mitogen-activated protein kinase; Akt, protein kinase B; TNF, tumor necrosis factor; TNFR, TNF receptor; NFκB; nuclear factor κB; JNK, c-Jun N-terminal kinases; PI3K, phosphatidylinositol 3-kinase; IL, interleukin.
Figure 2.
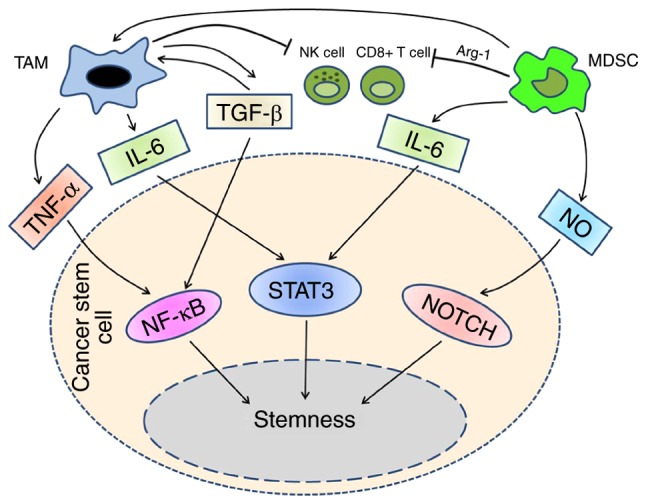
MDSC population could differentiate into TAMs, in tumor microenvironment, they suppress NK cells, CD8+ cells and other immune cells synergistically. The cytokines exist in tumor microenvironment mediate the interaction between TAMs and MDSCs with CSCs. TAMs activates NFκB and STAT3 pathways through TNF-α, IL-6 and TGF-β. In addition to IL-6, MDSCs utilize NO to activate STAT3 and NOTCH pathways. By means of these signal pathways, TAMs and MDSCs drive CSCs development, enhance the invasion and migration. CSCs, cancer stem cells; STAT, signal transducer and activator of transcription; TNF, tumor necrosis factor; NFκB; nuclear factor κB; IL, interleukin; MDSCs, myeloid-derived suppressor cells; TAMs, tumor-associated macrophages; TGF, transforming growth factor.
Table I.
A list of some CSC-related inflammatory cytokines, ligands, origins and their functions.
| Item | Origin | Receptors | Signal pathway | Functions (in tumor) | (Refs.) |
|---|---|---|---|---|---|
| IFN | Host cells | IFNGR1,2 | JAK/STAT | Induce EMT | 1,2,7–21 |
| (virus-infected cells) | IFNAR1,2 | p38/MAPK | Downregulate tumor-antigens | ||
| Fibroblasts | IFNLR1 | PI3K/Akt | Facilitate angiogenesis | ||
| Macrophages | C3G/Rap1 | Promote CSCs proliferation | |||
| T helper cells | |||||
| TNF | Activated-macrophages | TNFR1 | NFκB/HIF1a | Induce EMT | 7-9,22–27,31–37 |
| CD4+ lymphocytes | TNFR2 | JNK/MAPK | Increase stemness property | ||
| NK cells | Support CSCs survival | ||||
| Neutrophils | Enhance proliferation | ||||
| Mast cells | |||||
| IL-6 | Macrophages | CD126/IL6RA | JAK/STAT | Downregulate E-cadherin | 39-42,43,44,47–50 |
| Th2 cells | CD130/IR6RB | PI3K/Akt | Moderate CSCs' differentiation | ||
| B cells | Protect CSCs from apoptosis | ||||
| Astrocytes | Alter DNA methylation | ||||
| Endothelium | |||||
| IL-17 | Th17 | IL-17RA, B, | p38/MAPK | Drive CSCs' development | 57-59,61–66 |
| C, D and E | AP-1 | Promote CSCs' self-renewal | |||
| JAK/STAT3 | Enhance stemness, invasion, | ||||
| NFκB | and migration |
CSC, cancer stem cells; IFN, interferon; IFNAR, IFN-α/β receptor; JAK, Janus kinase; STAT, signal transducer and activator of transcription; p38, tumor protein 38; MAPK, mitogen-activated protein kinase; Akt, RAC-α serine/threonine protein kinase; C3G, Rap1, Ras-related protein 1; TNF, tumor necrosis factor; TNFR, TNF receptor; NFκB; nuclear factor κB; HIF1a, hypoxic inducible factor 1α; JNK, c-Jun N-terminal kinases; PI3K, phosphatidylinositol 3-kinase; AP-1, activator protein 1; EMT, epithelial-to-mesenchymal transition; Th, T helper cells; NK cells, Natural killer cells; CD, cluster of differentiation; IL, interleukin; IL6RA, IL 6 receptor.
2. Inflammatory cytokines
Interferons (IFNs)
IFNs are a group of pleiotropic cytokines that participate in multiple biological activities, such as antivirus infection, regulation of cell proliferation and immunological response (6). They are classically divided among three species: Type I IFNs; Type II IFNs; and Type III IFNs, and the two main categories in mammals are Type I and II. Type I contains two primary members, IFN-α and IFN-β, and the IFN-γ is the lonely member of the type II (7). Indeed, IFNs have been verified in the therapies of several types of cancer with success, such as chronic myeloid leukemia, T and B cell lymphomas, melanoma, renal carcinoma and so on. However, under certain conditions, IFNs was found to augment the numbers of T regulatory cells (Tregs) and Th17 cells. The myeloid-derived suppressor cells (MDSCs) that induced by IFNs, can express as protumorigenesis factors and promote the tumor cells escape from immunosurveillance (8). Therefore, IFNs participate in tumor immunology as a ‘double-edged sword’.
Mesenchymal stem cells (MSCs) are multipotential stromal cells that can differentiate into kinds of cell types, such as osteoblasts, chondrocytes, myocytes and adipocytes (9), and with self-renewal and pluripotent differentiation abilities, possessing the potential to displace damaged and diseased tissues (10). There is a closely interaction between the MSCs and the host immune system, the ability to inhibit proinflammatory cytokines of MSCs sets up the foundation for clinical applications in treating autoimmune diseases (11). In recent years, the MSCs have been proved to accelerate cancer progression in several types of cancer, and the IFNs have been shown to play critical roles in it. Wang et al demonstrated that IFN-γ and TNF-α, as two important representative inflammatory factors, play a crucial role in synergistically inducing MSCs deficiency and eventual MSCs tumorigenesis via the NFκB pathway in OVX-induced osteoporotic mice (12). Their research indicated that IFN-γ and TNF-α contributes in the MSCs-based tissue regeneration (13). It is also reported that IFN-γ and TNF-α in the MSCs enhanced tumor cells malignancy, induced EMT of breast cancer cells, and papillary thyroid cancer cells (14,15).
Numerous studies have demonstrated that IFNs are closely related to the CSCs, in the tumor cells proliferation, therapy resistance, and metastasis. Schürch, et al gived evidences of that IFN-γ induce proliferation and differentiation of chronic myeloid leukemia stem cells (16). In pancreatic carcinoma cells, IFN-α up-regulates the expression of CSC markers, promotes the metastasis formation (17). Yamashina et al revealed that the cancer stem-like cells from chemo-resistant tumors are able to produce IFN-regulated transcription factors, which promote macrophage colony-stimulating factor (M-CSF) production and generate tumorigenic myeloid cells, then facilitate the tumorigenic and stem cell activities of bulk tumors (18). In glioma stem cells, it is also demonstrated that the IFN-regulated factors promote tumorigenicity, angiogenesis, microglia recruitment and maintain glioma stem cells properties through induction of interleukin 6, C-X-C motif chemokine 1 and C-C motif chemokine 2 (19). Ojha R and his colleagues reported that in bladder cancer cells, JAK-mediated autophagy regulates stemness and cell survival via IFN-γ (20). In hepatocellular carcinoma, the researchers demonstrated that the IFN-γ treatment enriched the CD133+ liver CSCs population in vitro and in vivo (21).
In addition to the above, the IFNs could promote tumor progression via downregulating tumor antigens, facilitating angiogenesis, and maintaining an immunosuppressive tumor microenvironment (22,23). The roles of IFNs in malignancies maybe determined by tumor microenvironment, tumor types, and tumor stage and so on, for the two faces of IFNs in cancer, further studies are in great request to provide a promising prospect for IFNs-based treatment.
Tumor necrosis factor (TNF)
TNF superfamily refers to a group of cytokines that can cause cell death, the two main members of the family are TNF-α and TNF-β. Given that TNF-α accounts for 70~95% of TNF biological activities, it can represent the TNF superfamily in general (24). By virtue of the ability to cause cytolysis of certain tumor cell lines, TNF-α has been utilized as a potential anticancer agent for many years (25), but with the development of research, emerging evidences suggest that TNF-α is significant in promoting tumor progression, in particular, with CSCs.
TNF-α can induce EMT and increase stemness properties, that is demonstrated in renal cell carcinoma, hypopharyngeal cancer and cholangiocarcinoma cells (26,27). Synergized with IFN-γ, TNF-α stimulates MSCs to enhance malignancy of cancer cells, tumorigenesis (12–14), and resistance to chemotherapy (28). Yu et al validated that TNF-α-activated MSCs promote breast cancer metastasis via recruiting CXCR2+ neutrophils (29), a similar result is reported by Katanov C and his colleague (30).
In recent studies, it is revealed that TNF-α enhances CSCs phenotype of oral squamous cell carcinoma (OSCC) cells, such as an increase in tumor sphere-formation ability, stem cell associated genes expression, chemo-radioresistance, and tumorigenesis (31). Besides that, TNF-α upregulates SLUG (a mediator of EMT process) with a dependency on canonical NFκB/HIF1α signaling, then imparts breast cancer cells with stem cell-like features (32). In melanoma, it is evidenced that after treatment of TNF-α, the self-renewing capacity of stem-like cells is upregulated (33). The transcription factor Atonal homolog 1 (Atoh1) protein, stabilized by TNF-α, could enrich colon CSCs, and induce high malignant potential (34). In myeloid leukemia, TNF-α secreted by the CSCs could promote NFκB pathway/p65 pathway and support stem cells survival (35,36). Similarly, TNF-α induces NFκB pathway activation to protect colorectal CSCs from death, and induce tumor regression (37).
For the importance of TNF-α in promoting tumorigenesis and progression, it represents a novel target in tumor prevention and therapy (28). However, more studies are needed before ideal clinical outcomes can be achieved (38).
IL-6
IL-6 is one member of the ILs, which acts as both a pro-inflammatory cytokine and an anti-inflammatory myokine (39). IL-6, also referred to as B-cell stimulatory factor-2 (BSF-2) and IFN β-2, is a cytokine that regulates the immune response but also plays a role in modulating cell growth, differentiation, and survival. It is mainly secreted by T cells, macrophages and fibroblasts. IL-6 belongs to the ‘IL-6 type cytokine’ family that also includes leukemia inhibitory factor (LIF), IL-11, ciliary neurotrophic factor (CNTF), cardiotrophin-1 (CT-1) and oncostatin M (OSM) (40). IL-6 mediates its activity by a cell-surface typeIcytokine receptor complex, which includes the ligand-binding IL-6Rα chain (IL-6R, CD126), and the signal-transducing component gp130 (CD130) (41). After interaction with its receptor, IL-6 triggers the IL-6R and gp130 proteins to form a complex, which stimulates distinct pathways to perform its oncogenic consequence, these pathways consist of Janus kinases (JAK)/signal transducer and activator of transcription (STAT), mitogen-activated protein kinase (MAPK) and phosphatidylinositol 3 kinase (PI3K)/protein kinase B (Akt) (42).
The varying biological functions of IL-6 make it critical in the enhancement or suppression of tumor growth and progress, and it is involved in the proliferation and differentiation of malignant cells and found to be high in serum and tumor tissues of a majority of cancers, including prostate cancer (43), multiple myeloma (44), renal cell carcinoma (45) and breast cancer (46). Emerging evidences suggest that the IL-6 levels in the serum are implicated in aggressive tumor growth and response to therapies in many types of cancer, and also could be a prognostic marker for assessing disease stage and disease progression (43).
To be valid in the protumorigenesis, IL-6 frequently interacts with CSCs, such as moderating CSCs differentiation and maintaining the stemness of cancer cells. In lung cancer cells, IL-6 promotes the self-renewal of CD133+ CSC-like cells, and the IL-6 expressing CD133+ cells have significantly higher expression of EMT-related molecules (lower E-cadherin, higher N-cadherin, vimentin, and TWIST), as well as higher expression of metastasis-related molecules [MMP9, transforming growth factor (TGF)-β1] than IL-6 knocked down CD133+ cells (47). In subsequent study, it is reported that IL-6 signaling induces DNA repair while preventing CD133+ CSC-like cells from apoptotic death after radiation for lung cancer (44). Likewise, in cisplatin-resistant lung cancer cells, IL-6 plays an important role in triggering enhanced stemness during cisplatin resistance development, through the upregulation of hypoxia-inducible factors (HIFs) (48). Liu et al (49) reported the IL-6/JAK2/STAT3 pathway upregulates DNA methyltransferase 1 (DNMT1) and promotes cancer initiation and lung CSCs proliferation by downregulation of p53 and p21 resulting from DNA hypermethylation. By the activation of IL-6/JAK2/STAT3 pathway, MSCs can enhance the capability of tumor initiation in lung cancer cells (50). Correspondingly, TME and/or tumor cells can impact the function of IL-6. Lu et al (51) revealed that the adipose-derived stem cells (ADSCs) enhance the malignant characteristics of Lewis lung carcinoma cells, including cell growth ability and especially CSC property. Glioma stem cells (GSCs) initiate microglial IL-6 secretion via TLR4 signaling and that IL-6 induces glioma growth by supporting GSCs, and glioma-associated microglia/brain macrophages are the primary source of IL-6 in tumor (52). By secreting high levels of IL-6, ovarian MSCs (OvMSCs) enhance the proliferation, sphere and colony formation and tumorigenesis of SKOV3 cells (53). In colorectal cancer (CRC), it is proposed that CD90+ fibroblasts/myofibroblasts may be the major source of IL-6 in T2-T3 CRC tumors, then supports the stemness of tumor cells and mediates the immune adaptive inflammatory response favoring tumor growth (54). The role of epigenetic control in cancer onset and development has been extensively accepted in recent years, e.g., the IL-6/JAK2/STAT3 pathway upregulates DNA methyltransferase 1 (54), the long non-coding RNAs (lncRNAs) in liver CSCs (LCSCs) mediate the crosstalk between TNF-α/NFκB signaling and autocrine IL-6/STAT3 cascade, then suppressed LCSCs expansion by inhibiting IL-6 transcription and STAT3 activation (55). In monoclonal gammopathy of undetermined significance (MGUS) and multiple myeloma, the IL-6 production in the bone marrow MSC (BMMSC) is driven by the citrullination of histone H3 (56).
Overall, these evidences suggest that IL-6 is involved in mechanisms of CSCs stemness, and the significance of IL-6 in the crosstalk between tumor and its microenvironment. Therefore, the inhibition of IL-6 may be a useful adjunct to cancer therapy, with a need of targeting this protein to combat cancer.
IL-17
IL-17 is a potent proinflammatory cytokine mainly produced by activated T-helper 17 cells (Th17) and activated by IL-23. The IL-17, also called IL-17A, is the fundamental member of a group of cytokines called IL-17 family. Besides of IL-17A, there are IL-17B, IL-17C, IL-17D, IL-17E (also called IL-25), and IL-17F constitute this family. The IL-17 receptor group includes five receptors, IL-17RA, B, C, D and E (57). IL-17 binds IL-17R, and then forms a complex to mediate a variety of signaling pathway, such as JNK, Erk1/2, p38, AP-1, JAK/STAT3 and NFκB (58). By virtue of these pathways, IL-17 induces the production of many other cytokines (such as IL-6, G-CSF, GM-CSF, IL-1β, TGF-β, and TNF-α), chemokines (IL-8, GRO-α, and MCP-1), and prostaglandins (e.g., PGE2) from many cell types, then to play an important part in the initiation and progression of many diseases. As a pivotal factor of tumor microenvironment, IL-17 could facilitate cancer progression, from tumorigenesis, migration, invasion, and metastasis, to adapting the tumor in its ability to confer upon itself both immune evasion, and chemotherapy resistance (59).
In view of the significance of CSCs in the cancer progression, the interplay between IL-17 and CSCs has aroused great interests of the scientists and researchers in recent years. In CRC, the sphere cells cocultured with Foxp3+IL-17+ cells could express more CRC cell markers (CD133, CD44s, CD166, EpCAM, and ALDH1) than the control sphere cells, and when neutralizing anti-IL-17 antibody was added to the culture, the expression of CD133 and the rest of the CRC cell markers was abolished. These data suggest that Foxp3+IL-17+ cells are capable of driving cancer-initiating cells (CICs) development (60). In this study, they also demonstrated that hypoxia induces Foxp3+Tregs to express IL-17. Consistently, it is revealed that Th17 stemness may be partially controlled by signaling pathways such as hypoxia inducible factor HIF1α, Notch and Bcl (61). Lotti et al (62) reported that IL-17A can contribute to CICs maintenance through IL-17A receptor in CRC, and then promote protumorigenic CICs behavior, as well as contribute to CIC therapeutic resistance. It is also demonstrated that one role of IL-17 in ovarian CD133+ cancer stem-like cells (CSLCs) is to promote the self-renewal (63). By IL-17 overexpression, the growth and sphere formation capacities of ovarian CD133+ CSLCs were dramaticlly enhanced. The incentive function of IL-17 might be mediated by the NFκB and p38/MAPK signaling pathway. In human hepatocellular carcinoma (HCC), IL-17E binding to IL-17RB motivates NFκB and JAK/STAT3 pathways to facilitate propagation and self-renewal ability of CSCs, and these beneficial effects could be prevented by specific inhibitors of JAK and NFκB signals (64). Similarly, by NFκB and STAT3 pathways, IL-17-IL-17R interaction in glioma stem cells (GSCs) induces an autocrine and/or paracrine cytokine feedback loop, that may provide an important signaling pathway for the stemness of GSCs (65). In addition to that, the upregulation of IL-17 in gastric cancer results in the transformation of quiescent gastric CSCs into invasive gastric CSCs, from invasion, migration to tumor formation ability (66).
For the significance of the interaction between IL-17 and CSCs, targeting IL-17 may emerge as a feasible novel therapeutic strategy. That clearly demonstrates a need for more research in this area, and the potential application of anti-IL-17 in the overall management will be hopeful.
3. Inflammatory cells
MDSCs
MDSCs are one of the major cell populations in charge of mediating immune responses, and are a heterogeneous population of cells comprised of macrophages, dendritic cells (DCs) and granulocytes, which expand during tumor progression, autoimmune disease, infection and other pathological conditions, and can efficiently suppress T cell function (67). The immune cells of MDSCs are from the myeloid lineage (a family of cells that originate from bone marrow stem cells) and are consist of myeloid-cell progenitors and precursors of myeloid cells. Under normal condition, the immature myeloid cells (IMCs) quickly differentiate into mature granulocytes, macrophages or DCs, but in abnormal conditions such as cancer, infectious diseases, trauma or some autoimmune disorders, IMCs differentiate into MDSCs (68). A number of immune suppressive factors are associated with the immunosuppressive function of MDSCs, including arginase (encoded by ARG1), inducible nitric oxide synthase (iNOS, or NOS2), nitric oxide (NO) and reactive oxygen species (ROS). The signaling pathways activated in MDSCs induced biological process are mainly STAT6, STAT1, and NFκB.
In addition to the immunological functions, non-immunological functions of MDSC also should been illustrated, such as the promotion of tumorigenesis, tumor invasion and metastasis (69). Given that CSCs are responsible for cancer initiating, progression, metastasis and recurrence, it is reasonable to hypothesize that there are mutual effects between CSCs and MDSCs, and then mounting evidences verify the existence of the relationship. In an early study, the CD44+ cancer stem-like cells are shown to not only more strongly inhibit T-cell proliferation, but also more efficiently induce regulatory T cells (Treg cells) and MDSCs as compared with CD44− cells (70). In glioblastoma, it is shown that MDSCs recruited to the tumor microenvironment by CSCs, can promote the survival of CSCs, and also be responsible for the immune-evasive properties. Moreover, the CSCs conditioned media inhibits MDSCs apoptosis and increases arginase 1 (Arg1) production of murine bone marrow derived MDSCs (71). Based on the known evidence and their experimental findings, Gao et al (72) hypothesize that T cell immunoglobulin mucin-3 (Tim-3), which specifically expresses on Leukemic stem cells (LSCs), is beneficial for LSCs survival and AML progression by promoting expansion of MDSCs and differentiating into tumor-associated macrophages (TAMs) at the leukemia site. In a further study, it is demonstrated that the macrophage migration inhibitory factor (MIF), which was produced at high levels by glioblastoma stem cells, could increase production of the immune-suppressive enzyme Arg1 in MDSCs as a CXCR2-dependent manner (73). In return, MDSCs could enhance the stemness of CSCs in certain ways. In ovarian carcinoma, MDSCs trigger the expression of miRNA101 in cancer cells, which subsequently repressed the corepressor gene C-terminal binding protein-2 (CtBP2), and CtBP2 directly targets stem cell core genes leading to increased cancer cell stem cells properties and increases metastatic and tumorigenic potential (74). Panni et al (75) revealed that, depending on the activation of the STAT3 pathway, the pancreatic cancer TME transforms monocytes to monocytic MDSCs (Mo-MDSCs), and these cells enhance the stemness and mesenchymal properties CSCs. In a recent study, it is revealed that MDSCs promote tumor initiation by endowing breast cancer cell stemness as well as repressing T-cell activation, and these effects are depended on the crosstalk between STAT3 and NOTCH pathways in cancer cells (76).
As a critical member of the tumor microenvironment, MDSCs lead to not only the tumor-associated immunosuppression, but also the CSCs promotion, and could be a prospective target in anti-tumor treatment, but there are still a lot of efforts needed to put in, such as how to identify MDSCs, the mechanism of MDSCs accumulation in TME, and by which means can suppress MDSCs effectively.
Tumor-associated macrophages (TAMs)
TAMs are the macrophages infiltrating in tumor tissue, are the most immune cells in the tumor microenvironment. At first, it is believed that TAMs are able to recognize and eliminate tumor cells by a variety of cytokines, but as the development of research, it is now generally accepted that TAMs play a key role in tumor growth, invasion and migration (77). TAMs derive from monocytic precursors circulating in the vessels, when there is a tumor exists, monocytes could be recruited into TME, via the chemokines and cytokines secreted by tumor cells, tumor stromal cells, and immune cells, including chemokine (C-C motif) ligand-2 (CCL2), CCL5, Chemokine (C-X-C motif) ligand-1 (CXCL1), vascular endothelial growth factor (VEGF), platelet derived growth factor (PDGF), TGF and macrophage colony stimulating factor (M-CSF). Affected by the cells and cytokines, mononuclear phagocytes polarize into different types of TAM. Typically activated M1 macrophages are induced by IFN-γ alone or in cooperation with microbial stimuli (i.e., LPS) or cytokines (i.e., TNF and GM-CSF). On the other hand, IL-4 and IL-13 induce M2 macrophages alternatively (78). M1 macrophages often show a capability of anti-tumor, such as releasing toxic intermediates and proinflammatory cytokines, inducing specific immune response, and enhancing the antigen presenting. In contrast, M2 macrophages represent as a pro-tumor factor, through suppressing the inflammatory responses, promoting angiogenesis, inducing the invasion and metastasis of tumor. It is accepted that TAMs have functions and phenotype more similar to M2 macrophages, so TAMs are often considered as promoter of tumor progression (79).
A lot of evidences have clarified that TAMs and CSCs have close contact with each other, and the interaction between them play a critical role in tumorigenesis, progression, invasion, drug resistance and so on (80). Jinushi et al (81) found that TAMs produced amount of Milk-fat globule EGF-8 (MFG-E8) in stimulation with lung CSCs. The MFG-E8 enhanced tumorigenicity and drug resistance in CSCs, by means of the coordinated activation of STAT3 and Sonic Hedgehog signals. Furthermore, MFG-E8 and IL-6 from TAMs could synergistically mediate tumorigenesis and drug resistance within CSC. In breast cancer, it is reported that after the cocultivation of M2 macrophages and breast cancer cell lines MCF-7, hybrids can be isolated from the fusion, and they represent a more invasive phenotype, including increased migration, aggression and tumorigenicity, which implicated the role of TAMs in promoting CSCs (82). Additionally, by releasing high levels of TGF-β1, TAMs promote the invasion of glioma stem-like cells (GSLCs) and increase the production of MMP-9, then induce EMT (83). The role of TGF-β1/EMT is also revealed in hepatocellular carcinoma (84). Likewise, the importance of EGFR/Stat3/Sox-2 and IL-6/STAT3 signaling pathway in regulating CSCs also shows up in breast cancer and hepatocellular carcinoma (85–87).
Through the way of interaction, CSCs could recruit TAMs to promote tumor growth. It is demonstrated that the CD34− melanoma tumor-initiating cells (TICs) interact specifically with M2 macrophages, then support their resistance to chemotherapeutic drugs and accelerate cancer progression by TGF-β and arginase pathway (88). Zhou et al (89) and Shi et al (90) found that in Glioblastoma, the periostin (POSTN) preferentially secreted by GSCs could correlates with TAM density in primary GBMs. Silencing POSTN in GSCs can reduce TAM recruitment, inhibit tumor growth, and extend survival of mice bearing GSC-derived xenografts. Furthermore, they revealed that POSTN recruits TAMs via integrin αvβ3 signaling.
For the significance of the relationship between TAMs and CSCs, the coexpression of them could be a meaningful marker to the tumor diagnosis and prognostic, as well as a promising therapy target (91,92). We propose that the elimination or re-differentiation of macrophages within the tumor microenvironment could be a significant prong of combination therapies designed to treat malignancies.
4. Conclusion
Currently, cancer is still one of the biggest threats to human life. Despite of all the efforts have been put in the studies of tumor treatment in recent decades, the survival rate has not apparently been improved. Inflammatory TME mainly show up the promotion of CSCs, but sometimes it also represent as an inhibitory factor to the tumor. The mechanism of the role transformation is still not clear, so furthermore experimental and clinical researches are needed to shed light on the fundamental mechanisms. Given that Inflammatory TME and CSCs are two major barriers to valid cancer therapy, targeting them may provide a promising approach to developing novel treatments.
Acknowledgements
This study was supported by Research Grants (15411950300) from Science and Technology Commission of Shanghai Municipality, Science and Technology Commission Foundation of Shanghai (no. 10DZ1951300) and Shanghai Summit & Plateau Disciplines (to CPZ), the National Natural Science Foundation of China (no. 81771046) and Shanghai Summit & Plateau Disciplines (to LW), Natural Science Foundation of China (no. 81602367 to XY).
References
- 1.Bonnet D, Dick JE. Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat Med. 1997;3:730–737. doi: 10.1038/nm0797-730. [DOI] [PubMed] [Google Scholar]
- 2.Albini A, Bruno A, Gallo C, Pajardi G, Noonan DM, Dallaglio K. Cancer stem cells and the tumor microenvironment: Interplay in tumor heterogeneity. Connect Tissue Res. 2015;56:414–425. doi: 10.3109/03008207.2015.1066780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Voog J, Jones DL. Stem cells and the niche: A dynamic duo. Cell Stem Cell. 2010;6:103–115. doi: 10.1016/j.stem.2010.01.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kise K, Kinugasa-Katayama Y, Takakura N. Tumor microenvironment for cancer stem cells. Adv Drug Deliv Rev. 2016;99:197–205. doi: 10.1016/j.addr.2015.08.005. [DOI] [PubMed] [Google Scholar]
- 5.Elinav E, Nowarski R, Thaiss CA, Hu B, Jin C, Flavell RA. Inflammation-induced cancer: Crosstalk between tumours, immune cells and microorganisms. Nat Rev Cancer. 2013;13:759–771. doi: 10.1038/nrc3611. [DOI] [PubMed] [Google Scholar]
- 6.Corrales L, Gajewski TF. Molecular pathways: Targeting the stimulator of interferon genes (STING) in the immunotherapy of cancer. Clin Cancer Res. 2015;21:4774–4779. doi: 10.1158/1078-0432.CCR-15-1362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Trinchieri G. Type I interferon: Friend or foe? J Exp Med. 2010;207:2053–2063. doi: 10.1084/jem.20101664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zaidi MR, Merlino G. The two faces of interferon-γ in cancer. Clin Cancer Res. 2011;17:6118–6124. doi: 10.1158/1078-0432.CCR-11-0482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Nardi Beyer N, da Silva Meirelles L. Mesenchymal stem cells: Isolation, in vitro expansion and characterization. Handb Exp Pharmacol. 2006:249–282. doi: 10.1007/3-540-31265-X_11. [DOI] [PubMed] [Google Scholar]
- 10.Shi S, Gronthos S. Perivascular niche of postnatal mesenchymal stem cells in human bone marrow and dental pulp. J Bone Miner Res. 2003;18:696–704. doi: 10.1359/jbmr.2003.18.4.696. [DOI] [PubMed] [Google Scholar]
- 11.Sharma RR, Pollock K, Hubel A, Mckenna D. Mesenchymal stem or stromal cells: A review of clinical applications and manufacturing practices. Transfusion. 2014;54:1418–1437. doi: 10.1111/trf.12421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wang L, Zhao Y, Liu Y, Akiyama K, Chen C, Qu C, Jin Y, Shi S. IFN-γ and TNF-α synergistically induce mesenchymal stem cell impairment and tumorigenesis via NFκB signaling. Stem Cells. 2013;31:1383–1395. doi: 10.1002/stem.1388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Liu Y, Wang L, Kikuiri T, Akiyama K, Chen C, Xu X, Yang R, Chen W, Wang S, Shi S. Mesenchymal stem cell-based tissue regeneration is governed by recipient T lymphocytes via IFN-γ and TNF-α. Nat Med. 2011;17:1594–1601. doi: 10.1038/nm.2542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Trivanović D, Jauković A, Krstić J, Nikolić S, Okić Djordjević I, Kukolj T, Obradović H, Mojsilović S, Ilić V, Santibanez JF, Bugarski D. Inflammatory cytokines prime adipose tissue mesenchymal stem cells to enhance malignancy of MCF-7 breast cancer cells via transforming growth factor-β1. IUBMB Life. 2016;68:190–200. doi: 10.1002/iub.1473. [DOI] [PubMed] [Google Scholar]
- 15.Lv N, Gao Y, Guan H, Wu D, Ding S, Teng W, Shan Z. Inflammatory mediators, tumor necrosis factor-α and interferon-γ, induce EMT in human PTC cell lines. Oncol Lett. 2015;10:2591–2597. doi: 10.3892/ol.2015.3518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Schürch C, Riether C, Amrein MA, Ochsenbein AF. Cytotoxic T cells induce proliferation of chronic myeloid leukemia stem cells by secreting interferon-γ. J Exp Med. 2013;210:605–621. doi: 10.1084/jem.20121229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Zhu Y, Karakhanova S, Huang X, Deng SP, Werner J, Bazhin AV. Influence of interferon-α on the expression of the cancer stem cell markers in pancreatic carcinoma cells. Exp Cell Res. 2014;324:146–156. doi: 10.1016/j.yexcr.2014.03.020. [DOI] [PubMed] [Google Scholar]
- 18.Yamashina T, Baghdadi M, Yoneda A, Kinoshita I, Suzu S, Dosakaakita H, Jinushi M. Cancer stem-like cells derived from chemoresistant tumors have a unique capacity to prime tumorigenic myeloid cells. Cancer Res. 2014;74:2698–2709. doi: 10.1158/0008-5472.CAN-13-2169. [DOI] [PubMed] [Google Scholar]
- 19.Jin X, Kim SH, Jeon HM, Beck S, Sohn YW, Yin J, Kim JK, Lim YC, Lee JH, Kim SH, et al. Interferon regulatory factor 7 regulates glioma stem cells via interleukin-6 and Notch signalling. Brain. 2012;135:1055–1069. doi: 10.1093/brain/aws028. [DOI] [PubMed] [Google Scholar]
- 20.Ojha R, Singh SK, Bhattacharyya S. JAK-mediated autophagy regulates stemness and cell survival in cisplatin resistant bladder cancer cells. Biochim Biophys Acta. 2016;1860:2484–2497. doi: 10.1016/j.bbagen.2016.07.021. [DOI] [PubMed] [Google Scholar]
- 21.Li J, Chen JN, Zeng TT, He F, Chen SP, Ma S, Bi J, Zhu XF, Guan XY. CD133+ liver cancer stem cells resist interferon-gamma-induced autophagy. BMC Cancer. 2016;16:15. doi: 10.1186/s12885-016-2050-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Furuta J, Inozume T, Harada K, Shimada S. CD271 on melanoma cell is an IFN-γ-inducible immunosuppressive factor that mediates downregulation of melanoma antigens. J Invest Dermatol. 2014;134:1369–1377. doi: 10.1038/jid.2013.490. [DOI] [PubMed] [Google Scholar]
- 23.Kharma B, Baba T, Matsumura N, Kang HS, Hamanishi J, Murakami R, McConechy MM, Leung S, Yamaguchi K, Hosoe Y, et al. STAT1 drives tumor progression in serous papillary endometrial cancer. Cancer Res. 2014;74:6519–6530. doi: 10.1158/1538-7445.AM2014-LB-123. [DOI] [PubMed] [Google Scholar]
- 24.Wang AM, Creasey AA, Ladner MB, Lin LS, Strickler J, Van Arsdell JN, Yamamoto R, Mark DF. Molecular cloning of the complementary DNA for human tumor necrosis factor. Science. 1985;228:149–154. doi: 10.1126/science.3856324. [DOI] [PubMed] [Google Scholar]
- 25.Roberts NJ, Zhou S, Diaz LA, Matthias H. Systemic use of tumor necrosis factor alpha as an anticancer agent. Oncotarget. 2011;2:739–751. doi: 10.18632/oncotarget.344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Zhang L, Jiao M, Wu K, Li L, Zhu G, Wang X, He D, Wu D. TNF-α induced epithelial mesenchymal transition increases stemness properties in renal cell carcinoma cells. Int J Clin Exp Med. 2014;7:4951–4958. [PMC free article] [PubMed] [Google Scholar]
- 27.Techasen A, Namwat N, Loilome W, Bungkanjana P, Khuntikeo N, Puapairoj A, Jearanaikoon P, Saya H, Yongvanit P. Tumor necrosis factor-α (TNF-α) stimulates the epithelial-mesenchymal transition regulator Snail in cholangiocarcinoma. Med Oncol. 2012;29:3083–3091. doi: 10.1007/s12032-012-0305-x. [DOI] [PubMed] [Google Scholar]
- 28.Valizadeh A, Ahmadzadeh A, Saki G, Khodadadi A, Teimoori A. Role of tumor necrosis factor-producing mesenchymal stem cells on apoptosis of chronic B-lymphocytic tumor cells resistant to fludarabine-based chemotherapy. Asian Pac J Cancer Prev. 2015;16:8533–8539. doi: 10.7314/APJCP.2015.16.18.8533. [DOI] [PubMed] [Google Scholar]
- 29.Yu PF, Huang Y, Han YY, Lin LY, Sun WH, Rabson AB, Wang Y, Shi YF. TNFα-activated mesenchymal stromal cells promote breast cancer metastasis by recruiting CXCR2+ neutrophils. Oncogene. 2017;36:482–490. doi: 10.1038/onc.2016.252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Katanov C, Lerrer S, Liubomirski Y, Leider-Trejo L, Meshel T, Bar J, Feniger-Barish R, Kamer I, Soria-Artzi G, Kahani H, et al. Regulation of the inflammatory profile of stromal cells in human breast cancer: Prominent roles for TNF-α and the NF-κB pathway. Stem Cell Res Ther. 2015;6:87. doi: 10.1186/s13287-015-0080-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lee SH, Hong HS, Liu ZX, Kim RH, Kang MK, Park NH, Shin KH. TNFα enhances cancer stem cell-like phenotype via Notch-Hes1 activation in oral squamous cell carcinoma cells. Biochem Biophys Res Commun. 2012;424:58–64. doi: 10.1016/j.bbrc.2012.06.065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Storci G, Sansone P, Mari S, D'Uva G, Tavolari S, Guarnieri T, Taffurelli M, Ceccarelli C, Santini D, Chieco P, et al. TNFalpha up-regulates SLUG via the NF-kappaB/HIF1alpha axis, which imparts breast cancer cells with a stem cell-like phenotype. J Cell Physiol. 2010;225:682–691. doi: 10.1002/jcp.22264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ostyn P, El Machhour R, Begard S, Kotecki N, Vandomme J, Flamenco P, Segard P, Masselot B, Formstecher P, Touil Y, Polakowska R. Transient TNF regulates the self-renewing capacity of stem-like label-retaining cells in sphere and skin equivalent models of melanoma. Cell Commun Signal. 2014;12:52. doi: 10.1186/s12964-014-0052-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Fukushima K, Tsuchiya K, Kano Y, Horita N, Hibiya S, Hayashi R, Kitagaki K, Negi M, Itoh E, Akashi T, et al. Atonal homolog 1 protein stabilized by tumor necrosis factor α induces high malignant potential in colon cancer cell line. Cancer Sci. 2015;106:1000–1007. doi: 10.1111/cas.12703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Gallipoli P, Pellicano F, Morrison H, Laidlaw K, Allan EK, Bhatia R, Copland M, Jørgensen HG, Holyoake TL. Autocrine TNF-α production supports CML stem and progenitor cell survival and enhances their proliferation. Blood. 2013;122:3335–3339. doi: 10.1182/blood-2013-02-485607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Zhou X, Zhou S, Li B, Li Q, Gao L, Li D, Gong Q, Zhu L, Wang J, Wang N, et al. Transmembrane TNF-α preferentially expressed by leukemia stem cells and blasts is a potent target for antibody therapy. Blood. 2015;126:1433–1442. doi: 10.1182/blood-2015-01-624833. [DOI] [PubMed] [Google Scholar]
- 37.Sheng YH, He Y, Hasnain SZ, Wang R, Tong H, Clarke DT, Lourie R, Oancea I, Wong KY, Lumley JW, et al. MUC13 protects colorectal cancer cells from death by activating the NF-κB pathway and is a potential therapeutic target. Oncogene. 2017;36:700–713. doi: 10.1038/onc.2016.241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Nenu I, Tudor D, Filip AG, Baldea I. Current position of TNF-α in melanomagenesis. Tumor Biol. 2015;36:6589–6602. doi: 10.1007/s13277-015-3639-0. [DOI] [PubMed] [Google Scholar]
- 39.Bromberg J, Wang TC. Inflammation and cancer: IL-6 and STAT3 complete the link. Cancer Cell. 2009;15:79–80. doi: 10.1016/j.ccr.2009.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Smith PC, Hobisch A, Lin DL, Culig Z, Keller ET. Interleukin-6 and prostate cancer progression. Cytokine Growth Factor Rev. 2001;12:33–40. doi: 10.1016/S1359-6101(00)00021-6. [DOI] [PubMed] [Google Scholar]
- 41.Yao X, Huang J, Zhong H, Shen N, Faggioni R, Fung M, Yao Y. Targeting interleukin-6 in inflammatory autoimmune diseases and cancers. Pharmacol Ther. 2014;141:125–139. doi: 10.1016/j.pharmthera.2013.09.004. [DOI] [PubMed] [Google Scholar]
- 42.Hideshima T, Nakamura N, Chauhan D, Anderson KC. Biologic sequelae of interleukin-6 induced PI3-K/Akt signaling in multiple myeloma. Oncogene. 2001;20:5991–6000. doi: 10.1038/sj.onc.1204833. [DOI] [PubMed] [Google Scholar]
- 43.Kumari N, Dwarakanath BS, Das A, Bhatt AN. Role of interleukin-6 in cancer progression and therapeutic resistance. Tumor Biol. 2016;37:11553–11572. doi: 10.1007/s13277-016-5098-7. [DOI] [PubMed] [Google Scholar]
- 44.Chen Y, Zhang F, Tsai Y, Yang X, Yang L, Duan S, Wang X, Keng P, Lee SO. IL-6 signaling promotes DNA repair and prevents apoptosis in CD133+ stem-like cells of lung cancer after radiation. Radiat Oncol. 2015;10:227. doi: 10.1186/s13014-015-0534-1. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 45.Altundag O, Altundag K, Gunduz E. Interleukin-6 and C-reactive protein in metastatic renal cell carcinoma. J Clin Oncol. 2005;23:1044–1045. doi: 10.1200/JCO.2005.05.155. [DOI] [PubMed] [Google Scholar]
- 46.Knüpfer H, Preiß R. Significance of interleukin-6 (IL-6) in breast cancer (review) Breast Cancer Res Treat. 2007;102:129–135. doi: 10.1007/s10549-006-9328-3. [DOI] [PubMed] [Google Scholar]
- 47.Ok LS, Yang X, Duan S, Ying T, Strojny LR, Peter K, et al. IL-6 promotes growth and epithelial-mesenchymal transition of CD133+ cells of non-small cell lung cancer. Oncotarget. 2016;7:6626–6638. doi: 10.18632/oncotarget.6570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Zhang F, Duan S, Ying T, Keng PC, Chen Y, Lee SO, Chen Y. Cisplatin treatment increases stemness through upregulation of hypoxia-inducible factors by interleukin-6 in non-small cell lung cancer. Cancer Sci. 2016;107:746–754. doi: 10.1111/cas.12937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Liu CC, Lin JH, Hsu TW, Su K, Li AF, Hsu HS, Hung SC. IL-6 enriched lung cancer stem-like cell population by inhibition of cell cycle regulators via DNMT1 upregulation. Int J Cancer. 2015;136:547–559. doi: 10.1002/ijc.29033. [DOI] [PubMed] [Google Scholar]
- 50.Hsu HS, Lin JH, Hsu TW, Su K, Wang CW, Yang KY, Chiou SH, Hung SC. Mesenchymal stem cells enhance lung cancer initiation through activation of IL-6/JAK2/STAT3 pathway. Lung Cancer. 2012;75:167–177. doi: 10.1016/j.lungcan.2011.07.001. [DOI] [PubMed] [Google Scholar]
- 51.Lu JH, Wei HJ, Peng BY, Chou HH, Chen WH, Liu HY, Deng WP. Adipose-derived stem cells enhance cancer stem cell property and tumor formation capacity in lewis lung carcinoma cells through an interleukin-6 paracrine circuit. Stem Cells Dev. 2016;25:1833–1842. doi: 10.1089/scd.2016.0163. [DOI] [PubMed] [Google Scholar]
- 52.a Dzaye OD, Hu F, Derkow K, Haage V, Euskirchen P, Harms C, Lehnardt S, Synowitz M, Wolf SA, Kettenmann H. Glioma stem cells but not bulk glioma cells upregulate IL-6 secretion in microglia/brain macrophages via toll-like receptor 4 signaling. J Neuropathol Exp Neurol. 2016;75:429–440. doi: 10.1093/jnen/nlw016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Ding DC, Liu HW, Chu TY. Interleukin-6 from ovarian mesenchymal stem cells promotes proliferation, sphere and colony formation and tumorigenesis of an ovarian cancer cell line SKOV3. J Cancer. 2016;7:1815–1823. doi: 10.7150/jca.16116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Huynh PT, Beswick EJ, Yun AC, Johnson P, O'Connell MR, Watts T, Singh P, Qiu S, Morris K, Powell DW, Pinchuk IV. CD90(+) stromal cells are the major source of IL-6, which supports cancer stem-like cells and inflammation in colorectal cancer. Int J Cancer. 2016;138:1971–1981. doi: 10.1002/ijc.29939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Wang X, Sun W, Shen W, Xia M, Chen C, Xiang D, Ning B, Cui X, Li H, Li X, et al. Long non-coding RNA DILC regulates liver cancer stem cells via IL-6/STAT3 axis. J Hepatol. 2016;64:1283–1294. doi: 10.1016/j.jhep.2016.01.019. [DOI] [PubMed] [Google Scholar]
- 56.Mcnee G, Eales KL, Wei W, Williams DS, Barkhuizen A, Bartlett DB, Essex S, Anandram S, Filer A, Moss PA, et al. Citrullination of histone H3 drives IL-6 production by bone marrow mesenchymal stem cells in MGUS and multiple myeloma. Leukemia. 2017;31:373–381. doi: 10.1038/leu.2016.187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Kolls JK, Lindén A. Interleukin-17 family members and inflammation. Immunity. 2004;21:467–476. doi: 10.1016/j.immuni.2004.08.018. [DOI] [PubMed] [Google Scholar]
- 58.Guéry L, Hugues S. Th17 cell plasticity and functions in cancer immunity. Biomed Res Int. 2015;2015:314620. doi: 10.1155/2015/314620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Yang B, Fung A, Zhao H, Wang T, Ma D. The Role of Interleukin 17 in Tumour Proliferation, Angiogenesis and Metastasis. Mediators of Inflammation. 2014;2014:390–392. doi: 10.1155/2014/623759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Yang S, Wang B, Guan C, Wu B, Cai C, Wang M, Zhang B, Liu T, Yang P. Foxp3+IL-17+ T cells promote development of cancer-initiating cells in colorectal cancer. J Leukoc Biol. 2011;89:85–91. doi: 10.1189/jlb.0910506. [DOI] [PubMed] [Google Scholar]
- 61.Wei S, Zhao E, Kryczek I, Zou W. Th17 cells have stem cell-like features and promote long-term immunity. Oncoimmunology. 2012;1:516–519. doi: 10.4161/onci.19440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Lotti F, Jarrar AM, Pai RK, Hitomi M, Lathia J, Mace A, Gantt GA, Jr, Sukhdeo K, DeVecchio J, Vasanji A, et al. Chemotherapy activates cancer-associated fibroblasts to maintain colorectal cancer-initiating cells by IL-17A. J Exp Med. 2013;210:2851–2872. doi: 10.1084/jem.20131195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Xiang T, Long H, He L, Han X, Lin K, Liang Z, Zhuo W, Xie R, Zhu B. Interleukin-17 produced by tumor microenvironment promotes self-renewal of CD133+ cancer stem-like cells in ovarian cancer. Oncogene. 2015;34:165–176. doi: 10.1038/onc.2013.537. [DOI] [PubMed] [Google Scholar]
- 64.Luo Y, Yang Z, Su L, Shan J, Xu H, Xu Y, Liu L, Zhu W, Chen X, Liu C, et al. Non-CSCs nourish CSCs through interleukin-17E-mediated activation of NF-κB and JAK/STAT3 signaling in human hepatocellular carcinoma. Cancer Lett. 2016;375:390–399. doi: 10.1016/j.canlet.2016.03.012. [DOI] [PubMed] [Google Scholar]
- 65.Parajuli P, Anand R, Mandalaparty C, Suryadevara R, Sriranga PU, Michelhaugh SK, Cazacu S, Finniss S, Thakur A, Lum LG, et al. Preferential expression of functional IL-17R in glioma stem cells: Potential role in self-renewal. Oncotarget. 2016;7:6121–6135. doi: 10.18632/oncotarget.6847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Jiang YX, Yang SW, Li PA, Luo X, Li ZY, Hao YX, Yu PW. The promotion of the transformation of quiescent gastric cancer stem cells by IL-17 and the underlying mechanisms. Oncogene. 2017;36:1256–1264. doi: 10.1038/onc.2016.291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Gabrilovich DI, Nagaraj S. Myeloid-derived suppressor cells as regulators of the immune system. Nat Rev Immunol. 2009;9:162–174. doi: 10.1038/nri2506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Condamine T, Gabrilovich DI. Molecular mechanisms regulating myeloid-derived suppressor cell differentiation and function. Trends Immunol. 2011;32:19–25. doi: 10.1016/j.it.2010.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Medina-Echeverz J, Aranda F, Berraondo P. Myeloid-derived cells are key targets of tumor immunotherapy. Oncoimmunology. 2014;3:e28398. doi: 10.4161/onci.28398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Chikamatsu K, Okamoto A, Sakakura K, Hatsushika K, Takahashi G, Masuyama K. P3.14. Immunoregulatory properties of CD44+ cancer stem-like cells in squamous cell carcinoma of the head and neck. Oral Oncol Suppl. 2009;3:205–206. doi: 10.1016/j.oos.2009.06.540. [DOI] [Google Scholar]
- 71.Otvos B, Finke J, Vogelbaum M, Lathia JD. Interrogating the interactions between myeloid derived suppressor cells and cancer stem cells in glioblastoma. J Immuno Ther Cancer. 2013;1:268. doi: 10.1186/2051-1426-1-S1-P268. [DOI] [Google Scholar]
- 72.Gao L, Yu S, Zhang X. Hypothesis: Tim-3/galectin-9, a new pathway for leukemia stem cells survival by promoting expansion of myeloid-derived suppressor cells and differentiating into tumor-associated macrophages. Cell Biochem Biophys. 2014;70:273–277. doi: 10.1007/s12013-014-9900-0. [DOI] [PubMed] [Google Scholar]
- 73.Otvos B, Silver DJ, Mulkearns-Hubert EE, Alvarado AG, Turaga SM, Sorensen MD, Rayman P, Flavahan WA, Hale JS, Stoltz K, et al. Cancer stem cell-secreted macrophage migration inhibitory factor stimulates myeloid derived suppressor cell function and facilitates glioblastoma immune evasion. Stem Cells. 2016;34:2026–2039. doi: 10.1002/stem.2393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Cui TX, Kryczek I, Zhao L, Zhao E, Kuick R, Roh MH, Vatan L, Szeliga W, Mao Y, Thomas DG, et al. Myeloid-derived suppressor cells enhance stemness of cancer cells by inducing microRNA101 and suppressing the corepressor CtBP2. Immunity. 2013;39:611–621. doi: 10.1016/j.immuni.2013.08.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Panni RZ, Sanford DE, Belt BA, Mitchem JB, Worley LA, Goetz BD, Mukherjee P, Wang-Gillam A, Link DC, Denardo DG, et al. Tumor-induced STAT3 activation in monocytic myeloid-derived suppressor cells enhances stemness and mesenchymal properties in human pancreatic cancer. Cancer Immunol Immunother. 2014;63:513–528. doi: 10.1007/s00262-014-1527-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Peng D, Tanikawa T, Li W, Zhao L, Vatan L, Szeliga W, Wan S, Wei S, Wang Y, Liu Y, et al. Myeloid-derived suppressor cells endow stem-like qualities to breast cancer cells through IL6/STAT3 and NO/NOTCH cross-talk signaling. Cancer Res. 2016;76:3156–3165. doi: 10.1158/0008-5472.CAN-15-2528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Shih JY, Yuan A, Chen JW, Yang PC. Tumor-associated macrophage: Its role in cancer invasion and metastasis. J Cancer Mol. 2006;2:101–106. [Google Scholar]
- 78.Allavena P, Sica A, Solinas G, Porta C, Mantovani A. The inflammatory micro-environment in tumor progression: The role of tumor-associated macrophages. Crit Rev Oncol Hematol. 2008;66:1–9. doi: 10.1016/j.critrevonc.2007.07.004. [DOI] [PubMed] [Google Scholar]
- 79.Franklin RA, Liao W, Sarkar A, Kim MV, Bivona MR, Liu K, Pamer EG, Li MO. The cellular and molecular origin of tumor-associated macrophages. Science. 2014;344:921–925. doi: 10.1126/science.1252510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Raggi C, Mousa HS, Correnti M, Sica A, Invernizzi P. Cancer stem cells and tumor-associated macrophages: A roadmap for multitargeting strategies. Oncogene. 2016;35:671–682. doi: 10.1038/onc.2015.132. [DOI] [PubMed] [Google Scholar]
- 81.Jinushi M, Chiba S, Yoshiyama H, Masutomi K, Kinoshita I, Dosaka-Akita H, Yagita H, Takaoka A, Tahara H. Tumor-associated macrophages regulate tumorigenicity and anticancer drug responses of cancer stem/initiating cells. Proc Natl Acad Sci USA. 2011;108:12425–12430. doi: 10.1073/pnas.1106645108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Ding J, Jin W, Chen C, Shao Z, Wu J. Tumor associated macrophage × cancer cell hybrids may acquire cancer stem cell properties in breast cancer. PLoS One. 2012;7:e41942. doi: 10.1371/journal.pone.0041942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Ye XZ, Xu SL, Xin YH, Yu SC, Ping YF, Chen L, Xiao HL, Wang B, Yi L, Wang QL, et al. Tumor-associated microglia/macrophages enhance the invasion of glioma stem-like cells via TGF-β1 signaling pathway. J Immunol. 2012;189:444–453. doi: 10.4049/jimmunol.1103248. [DOI] [PubMed] [Google Scholar]
- 84.Fan QM, Jing YY, Yu GF, Kou XR, Ye F, Gao L, Li R, Zhao QD, Yang Y, Lu ZH, Wei LX. Tumor-associated macrophages promote cancer stem cell-like properties via transforming growth factor-beta1-induced epithelial-mesenchymal transition in hepatocellular carcinoma. Cancer Lett. 2014;352:160–168. doi: 10.1016/j.canlet.2014.05.008. [DOI] [PubMed] [Google Scholar]
- 85.Yang J, Liao D, Chen C, Liu Y, Chuang TH, Xiang R, Markowitz D, Reisfeld RA, Luo Y. Tumor-associated macrophages regulate murine breast cancer stem cells through a novel paracrine EGFR/Stat3/Sox-2 signaling pathway. Stem Cells. 2013;31:248–258. doi: 10.1002/stem.1281. [DOI] [PubMed] [Google Scholar]
- 86.Zhou N, Zhang Y, Zhang X, Lei Z, Hu R, Li H, Mao Y, Wang X, Irwin DM, Niu G, Tan H. Exposure of tumor-associated macrophages to apoptotic MCF-7 cells promotes breast cancer growth and metastasis. Int J Mol Sci. 2015;16:11966–11982. doi: 10.3390/ijms160611966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Wan S, Zhao E, Kryczek I, Vatan L, Sadovskaya A, Ludema G, Simeone DM, Zou W, Welling TH. Tumor-associated macrophages produce interleukin 6 and signal via STAT3 to promote expansion of human hepatocellular carcinoma stem cells. Gastroenterology. 2014;147:1393–1404. doi: 10.1053/j.gastro.2014.08.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Tham M, Tan KW, Keeble J, Wang X, Hubert S, Barron L, Tan NS, Kato M, Prevost-Blondel A, Angeli V, Abastado JP. Melanoma-initiating cells exploit M2 macrophage TGFβ and arginase pathway for survival and proliferation. Oncotarget. 2014;5:12027–12042. doi: 10.18632/oncotarget.2482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Zhou W, Ke SQ, Huang Z, Flavahan W, Fang X, Paul J, Wu L, Sloan AE, McLendon RE, Li X, et al. Periostin secreted by glioblastoma stem cells recruits M2 tumour-associated macrophages and promotes malignant growth. Nat Cell Biol. 2015;17:170–182. doi: 10.1038/ncb3090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Shi Y, Ping Y, Zhang X, Bian XW. Hostile takeover: Glioma stem cells recruit TAMs to support tumor progression. Cell Stem Cell. 2015;16:219–220. doi: 10.1016/j.stem.2015.02.008. [DOI] [PubMed] [Google Scholar]
- 91.He KF, Zhang L, Huang CF, Ma SR, Wang YF, Wang WM, Zhao ZL, Liu B, Zhao YF, Zhang WF, Sun ZJ. CD163+ tumor-associated macrophages correlated with poor prognosis and cancer stem cells in oral squamous cell carcinoma. Biomed Res Int. 2014;2014:838632. doi: 10.1155/2014/838632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Hou YC, Chao YJ, Tung HL, Wang HC, Shan YS. Coexpression of CD44-positive/CD133-positive cancer stem cells and CD204-positive tumor-associated macrophages is a predictor of survival in pancreatic ductal adenocarcinoma. Cancer. 2014;120:2766–2777. doi: 10.1002/cncr.28774. [DOI] [PMC free article] [PubMed] [Google Scholar]