

Abstract
Although the last 50 years of clinical and preclinical research have demonstrated that addiction is a brain disease, we still have no neural circuit–based treatments for substance dependence or cue reactivity at large. Now, for the first time, it appears that a noninvasive brain stimulation technique known as transcranial magnetic stimulation (TMS), which is Food and Drug Administration approved to treat depression, may be the first tool available to fill this critical void in addiction treatment development. The goals of this review are to 1) introduce TMS as a tool to induce causal change in behavior, cortical excitability, and frontal–striatal activity; 2) describe repetitive TMS (rTMS) as an interventional tool; 3) provide an overview of the studies that have evaluated rTMS as a therapeutic tool for alcohol and drug use disorders; and 4) outline a conceptual framework for target selection when designing future rTMS clinical trials in substance use disorders. The manuscript concludes with some suggestions for methodological innovation, specifically with regard to combining rTMS with pharmacotherapy as well as cognitive behavioral training paradigms. We have attempted to create a comprehensive manuscript that provides the reader with a basic set of knowledge and an introduction to the primary experimental questions that will likely drive the field of TMS treatment development forward for the next several years.
I. Introduction
Substance dependence is an intransigent, worldwide health problem that affects over 1 billion individuals. Approximately 1.1 billion people smoke cigarettes on a regular basis; 6 million deaths every year are attributable to smoking (World Health Organization, 2012); and at least 76 million individuals meet criteria for alcohol use disorder, which is associated with 1.8 million deaths per year (World Health Organization, 2004). Although addiction affects populations that vary in age from adolescents to senior citizens, from low to high socioeconomic status, and includes substances that are naturally grown and synthetically processed, all of these substance-dependent populations are united by two common features—elevated reactivity to environmental cues and disrupted activity in frontal–striatal circuits.
The basic recovery process for any substance use disorder includes management of acute withdrawal (detoxification), abstinence initiation, and relapse prevention. Whereas the majority of medical attention is spent on management of acute withdrawal and short-term detoxification programs, there is very little evidence to suggest these short-term interventions improve long-term abstinence rates.
The earliest long-term treatments for addiction sought to address the underlying maladaptive behaviors. In 1935, Bill Wilson and Robert Smith developed the Alcoholics Anonymous group, which has been expanded into the 12-step facilitation program for other drugs of abuse. The 12-step facilitation program is based on the application of spiritual and moral principles. Other behavioral therapies include cognitive behavioral therapy, motivational interviewing, and contingency management. These therapies all seek to increase the patient’s cognitive engagement in treatment and develop behavioral strategies to reduce the likelihood of relapse. Although these behavioral therapies play a very important role in addiction treatment and relapse prevention, the efficacy of behavioral therapy alone varies from low to moderate based on the primary substance of abuse (Dutra et al., 2008).
As knowledge grew about the biologic mechanisms for addiction, pharmacotherapies were explored based on the implicated receptor systems. Although the biologic target of these interventions differs by class of substance use disorder, the majority of these drugs rely upon full or partial agonist or antagonist effects on a specific neurotransmitter system. There are several Food and Drug Administration (FDA)–approved medications for alcohol use disorder (e.g., naltrexone, acamprosate), nicotine use disorder (e.g., varenicline, nicotine replacement), as well as opiate use disorder (e.g., naloxone, buprenorphine, methadone). To date, there are no FDA-approved pharmacologic treatments for either cocaine or methamphetamine. With technological advances in drug development, it is now possible to design pharmacotherapeutics for substance use disorders that can specifically target receptor subtypes believed to be involved in various portions of the addiction process (i.e., drug-cued reinstatement) and precisely manipulate the pharmacokinetic properties of these compounds (i.e., long-acting formulations). Although these neurochemically targeted pharmacotherapeutics are often very elegant and effective at modulating a specific neurotransmitter system, it is still difficult to limit their site of action to a specific neural region or circuit in the brain. This is particularly true for pharmacotherapeutic agents that rely on glutamatergic and GABAergic mechanisms, given the broad distribution of these neurotransmitter systems throughout the brain.
Recently, there has been an emerging emphasis in designing neural circuit–specific treatment options for patients with substance use disorders. This neural circuit–based emphasis in the substance use research community has been hastened by advances in neuroimaging technology and preclinical studies in the 1990s and 2000s. Preclinical studies have revealed that there is a causal relationship between activity in the frontal–striatal circuits and drug use behavior, whereas clinical studies have demonstrated that activity in these frontal–striatal circuits are also biomarkers for predicting vulnerability to relapse across multiple substance use disorders.
During this same period of time, the depression research community was in the process of developing a noninvasive neural circuit–based treatment of major depression disorder. This treatment, known as repetitive transcranial magnetic stimulation (rTMS), was approved as a therapy for major depressive disorder by the FDA in 2005 and is now widely covered by Medicare. Repetitive transcranial magnetic stimulation (TMS) (section Transcranial Magnetic Stimulation: An Interventional Tool) was an extension of single-pulse TMS (section Single Pulse Transcranial Magnetic Stimulation: A Biomarker and Probe), which had been invented by Dr. Anthony Barker in 1984 in Sheffield, England. A group of investigators at the National Institute of Neurologic Disease and Stroke was the first to demonstrate that repeated pulses of TMS (rTMS) at 1 Hz over the motor cortex decreased cortical excitability. These decreases in cortical excitability associated with 1 Hz rTMS are referred to as long-term depression (LTD)–like effects of rTMS—a reference to the preclinical concept of LTD. Likewise, rTMS at 10 Hz (and 20 Hz) increases cortical excitability. These are referred to as long-term potentiation (LTP)–like effects of rTMS—a reference to the preclinical concept of LTP. These foundational studies over the motor cortex were based on parameters that had been evaluated in preclinical electrophysiology models of neural plasticity. These studies paved the way for psychiatry researchers at the National Institute of Mental Health (and several other universities) who were interested in evaluating the therapeutic potential that rTMS might have for their patients—many of whom had elevated or depressed activity in the prefrontal cortex. Twenty years after Dr. Barker’s initial manuscript was published (1985), TMS has become an FDA-approved treatment tool for depression (2005). As of 2018 there are four FDA-approved devices to deliver rTMS as a treatment tool for depression, and there are many other clinical indications for which rTMS has a lot of potential promise—one of which is substance use disorders.
In the next sections, we will introduce TMS as a novel neuromodulation tool being applied to various substance-dependent populations and describe its utility as a probe for cortical excitability (section Single-Pulse Transcranial Magnetic Stimulation: A Biomarker and Probe), its effects on pharmacology (section Single-Pulse Transcranial Magnetic Stimulation: A Biomarker and Probe), as an intervention (section Repetitive Transcranial Magnetic Stimulation: An Interventional Tool), and how it is currently being evaluated as a novel treatment tool for alcohol, nicotine, and cocaine use disorders (section Repetitive Transcranial Magnetic Stimulation: An Interventional Tool).
II. Single-Pulse Transcranial Magnetic Stimulation: A Biomarker and Probe
A. Fundamentals
1. What Is Single-Pulse Transcranial Magnetic Stimulation?
Although a comprehensive review of the basic mechanisms of TMS is beyond the scope of this manuscript, prior behavioral, electrophysiological, and neuroimaging work in this area is well described and summarized in several review articles (see: Fitzgerald et al., 2006; Thickbroom, 2007; Hoogendam et al., 2010; Ziemann, 2011). In its most basic form, TMS is administered through a handheld circular coil that contains 100–200 windings of metal wire contained in a magnetically translucent casing (i.e., plastic). When electricity is passed through the wire, a magnetic field is created perpendicular to the plane of the wound coil. By applying pulses of electricity through the coil, the magnetic field will also have a pulsatile nature, and, consistent with Maxwell’s third law, will induce an electrical field perpendicular to the magnetic field.
When this field is induced in the vicinity of an electrically sensitive biologic system (such as muscle or neurons), an electrical current will be induced in the biologic medium. In practice, this is easily demonstrated by holding a TMS coil parallel to someone’s forearm. When pulses of electricity are allowed to run through the coil, a pulsatile magnetic field will be created, and in turn the muscles of the forearm that are in the spatial distribution of the induced electrical field will contract. The strength of the muscle contraction is proportional to the strength of the induced magnetic field, which is related to the current (Maxwell, 1861).
For transcranial magnetic stimulation, these same principles are applied to cortical neurons (rather than forearm muscles). In the first demonstration of TMS, Dr. Barker was able to show that when a circular TMS coil was positioned over the area of the left primary motor cortex somatotopically related to the right hand, he could induce a motor-evoked potential (MEP) in the right hand that was proportional to the strength of the TMS field. Since Dr. Barker’s discovery in 1984, there have been significant advancements in TMS coil designs, including figure-of-eight coils (which can induce a more focused field change) and H-coil designs (which allow a deeper penetration of the effects of TMS, enabling broader TMS target engagement).
2. The Spatial Topography of a Single Transcranial Magnetic Stimulation Pulse
One of the most common questions posed by clinical and preclinical researchers is the following: “How precise is a TMS pulse?” With a growing number of TMS coil designs, this is an increasingly complex question to answer. The focality of TMS is related to the shape of the coil. There is a substantial body of literature devoted to computational modeling of electric field distributions associated with different coil shapes. In one of the most comprehensive papers, Deng et al. (2013) investigated the focality and penetration depth of 50 existing TMS coils (Deng et al., 2013). Their computational models revealed that typical figure-of-eight coil affected approximately 10 cm2 of cortical surface, circular coils affected approximately 50 cm2, and H-coil designs affected approximately 100 cm2. Most flat coil designs had penetration depths from 1 to 2 cm2, whereas the H-coil designs had consistently higher depths of 2 to 3 cm. A single TMS pulse from a standard figure-of-eight coil stimulates a 12.5-cm2 area, which is approximately 1/125 (0.8%) of the cortical surface area. By comparison, deep brain stimulation is an order of magnitude more precise, with volumes ranging from 1 to 20 cm (Wei and Grill, 2005), whereas electroconvulsive therapy and magnetic seizure therapy affect approximately 94% and 21% of the brain, respectively (Lee et al., 2016).
With TMS it is also possible to modulate monosynaptic (and possibly polysynaptic) targets of these cortical areas. The indirect effects of cortical TMS on monosynaptic afferent targets can be demonstrated through a behavioral assessment of the recruitment curve. When TMS is applied to the hand area of the primary motor cortex, there is a dose-dependent change in the amplitude of the MEP on the hand contralateral to the TMS coil. This pathway from the motor cortex to the hand requires at least two neurons—the upper motor neuron, which originates in the motor cortex and terminates in the spinal cord, and the lower motor neuron, which originates in the spinal cord and terminates in the muscles that will contract to produce the MEP. The majority of upper motor neurons, however, terminate on interneurons, which then facilitate lower motor neuron activity—suggesting that TMS can have polysynaptic effects. This ability to stimulate monosynaptic and possibly polysynaptic neurons is important for substance abuse treatment development research because many areas of interest for stimulation in substance use treatment are located in subcortical areas. Whereas these areas (i.e., striatum, amygdala) are not directly reachable with most TMS coils, neuroimaging data and electrophysiology data have demonstrated that they can be modulated via the cortical source of their afferent input.
3. Single-Pulse Transcranial Magnetic Stimulation as a Measurement Tool
Single and paired pulses of TMS are often used as a method to probe cortical excitatory and inhibitory tone (review Chen, 2000). The majority of our knowledge regarding the basic electrophysiological effects of TMS on the brain is from studies in the motor system. As mentioned above, a single pulse of TMS over the motor cortex results in a MEP in the limb contralateral to the stimulation site. A summary of the TMS probes commonly used and their sensitivity as markers of neurochemistry is displayed in Table 1. The most commonly reported dependent measure in TMS literature is the resting motor threshold (MT). This parameter is defined as the minimum TMS intensity required to generate a detectable MEP on 50% of the observations. Nearly all clinical treatment studies of rTMS use the MT as their dosing metric. In the FDA-approved protocol for TMS in depression, for example, patients are given 10 Hz stimulation over the dorsolateral prefrontal cortex at 120% of their resting MT. This enables rTMS to be titrated to each individual patient—an important step toward precision medicine. Although MT is consistent within an individual, there is a significant between-individual variability— nearly 70% of which is accounted for by variability in scalp-to-cortex distance (Herbsman et al., 2009).
TABLE 1.
A brief summary of TMS measures of cortical excitability and their relationship to neurotransmitters
| TMS Measure | MEP (right anterior pollicis brevis response, following TMS to the left motor cortex) | Neurochemistry Involved | Select References |
|---|---|---|---|
| Resting motor threshold |  |
Ion channels (voltage-gated Na+, AMPA glutamate) | Ziemann and Rothwell (2000), Di Lazzaro et al. (2008) |
| Paired pulse facilitation (long interval facilitation, 15 ms) | 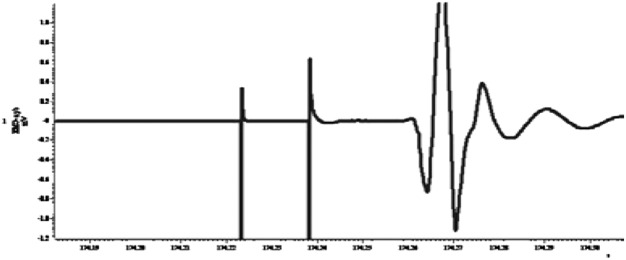 |
Glutamate (AMPA, NMDA) | Reis et al. (2008) |
| Paired pulse inhibition (short interval inhibition, 3 ms) | 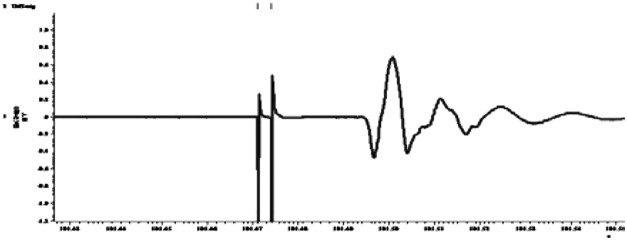 |
GABAa | Ziemann et al. (1996), Di Lazzaro et al. (2000a), Korchounov et al. (2005) |
| Cortical silent period | 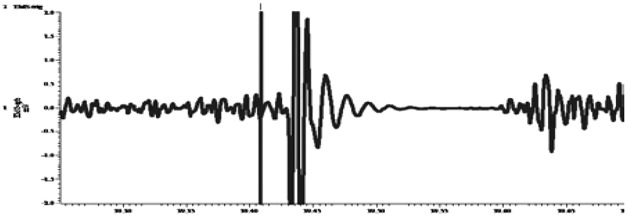 |
GABAb, dopamine | Ziemann et al. (1993) |
| Recruitment curve (dose–response curve) | 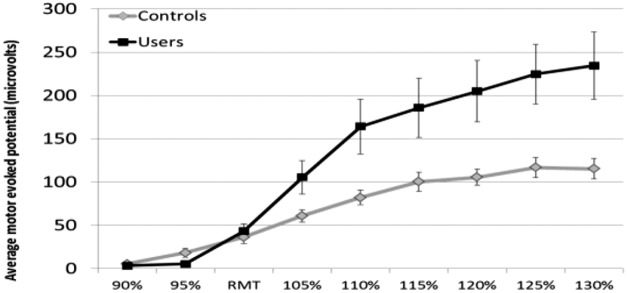 |
Causal relationship between TMS input and MEP output | Hanlon et al. (2015a) |
Beyond calculating the resting MT, single pulses of TMS at variable intensities (80%–120% of MT) can also be used to calculate a recruitment curve. This is expressed as a dose-response relationship wherein the amplitude of the TMS intensity is plotted against the amplitude of the average evoked MEP at that intensity. The shape of the recruitment curve is influenced by the balance of local inhibition and facilitation. Paired pulses of TMS can be used to investigate intracortical inhibition (ICI) and intracortical facilitation (ICF). In these paired-pulse procedures, a subthreshold conditioning stimulus (typically 80% motor threshold) is followed by suprathreshold test stimulus (typically 120% motor threshold) (Ziemann et al., 1996; Kujirai et al., 2006). Short interstimulus intervals (1–4 milliseconds) lead to inhibition, whereas longer interstimulus intervals (8–15 milliseconds) lead to facilitation.
B. Relationship between Pharmacology and Transcranial Magnetic Stimulation Measures
Beyond simply evaluating cortical facilitation and inhibition, however, these TMS measures can be used as a tool to evaluate the effects of pharmaceutical agents on cortical excitability in clinical populations. In fact, TMS and electroencephalography (EEG) are currently the only noninvasive electrophysiology tools available to assess the effects of a pharmacologic treatment on cortical excitability. The interaction between TMS and pharmacology has been an active area of research since the 1990s. Many of these studies have been elegantly summarized in several reviews (Ziemann, 2004; Ziemann et al., 2015). As stated above, TMS can be used as both a probe (single and paired pulses) and an interventional tool (rTMS). Consequently, studies that combine both pharmacology with TMS electrophysiology can be divided into two categories: 1) studies that investigated the effects of pharmaceutical agents on TMS measures of cortical excitability, and 2) studies that investigated the effects of various rTMS protocols on endogenous neurotransmitters (discussed in section Repetitive Transcranial Magnetic Stimulation: An Interventional Tool). As of 2015 there were 34 studies that took the first approach—evaluating the effect of various drugs on TMS electrophysiology parameters. These parameters include resting motor threshold, cortical silent period (CSP), paired pulse inhibition, and paired pulse facilitation. In a review of these studies, Ziemann et al. (2015) classified each study by the predominant neurotransmitter system that the drug targeted (voltage gated ion channels, antiglutamatergic, GABAergic, dopaminergic, noradrenergic) (Fig. 1). Of the drugs that were investigated, vigabatrin (GABA-ergic agonist) and lorazepam (GABA antagonist) had the largest effects on cortical silent period—a measure of cortical inhibitory tone, wherein vigabatrin caused a significant increase in inhibition (Pierantozzi et al., 2004), whereas lorazapam caused a significant decrease in inhibition (Kimiskidis et al., 2006). Of the dopaminergic and noradrenergic drugs, levodopa had a significant inhibitory effect (lengthening the cortical silent period) (Priori et al., 1994), whereas methylphenidate appears to have dose-dependent effect—increasing cortical facilitation at 12 mg (Moll et al., 2003) but increasing inhibition at 30 mg (Gilbert et al., 2006) and 40 mg (Ilic et al., 2003).
Fig. 1.

A summary of studies that have investigated the effects of various FDA-approved medications on cortical excitability using TMS. In a previous review, the extant literature in this field (n = 34 studies; Ziemann et al., 2015) grouped each drug into a class (based on its stated mechanism of action in that study) (top left). The average effect size (Cohen’s d) of the drugs in that class was then compiled for several TMS electrophysiology tests (e.g., cortical silent period). The data are reported as follows: average effect size (number of published studies with significant results, number of drugs investigated in that class). Effect sizes with an absolute value greater than 0.8 are strong, greater than 0.5 are moderate, and less than 0.5 are weak.
C. Using Transcranial Magnetic Stimulation To Study Cortical Excitability in Addiction
1. Cocaine
Single-pulse TMS protocols (such as paired pulse inhibition, facilitation, and cortical silent period) have been used to investigate disruptions in cortical excitability by multiple groups (Boutros et al., 2001, 2005; Sundaresan et al., 2007; Gjini et al., 2012; Bunse et al., 2014; Hanlon et al., 2015a). Our group, for example, demonstrated that the slope of the TMS dose-response curve was significantly steeper in current, chronic cocaine users than healthy controls. Additionally, the baseline motor threshold in cocaine users is higher than controls (Hanlon et al., 2015a). This difference means that it takes more energy to generate an initial MEP in cocaine users, but, once a threshold had been reached in cocaine users, the slope of the dose-response curve was very steep. These data suggest that the ratio of local excitatory to inhibitory processes in the motor cortex (e.g., likely glutamate or GABA mediated) is higher in cocaine users.
Several studies have demonstrated that abstinent cocaine users have lower motor cortex excitability than nondrug-using controls (Boutros et al., 2001, 2005; Sundaresan et al., 2007; Gjini et al., 2012). These lower MEPs in abstinent cocaine users are likely due to lower cortical (rather than peripheral nervous system) excitability (Ziemann et al., 1996; Di Lazzaro et al., 2008).
Furthermore, we have demonstrated that cocaine users had a lower ventral striatal blood-oxygen-level dependent (BOLD) response to single pulses of TMS stimulation to the medial prefrontal cortex (MPFC) than healthy controls (Hanlon et al., 2016). In this study, transcranial magnetic stimulation was applied to the left frontal pole (MPFC area of Brodmann area 10) and the left dorsolateral prefrontal cortex (DLPFC; lateral Brodmann 9) of 36 individuals—18 cocaine-dependent individuals with a history of failed quit attempts and 18 age-matched controls. Cocaine users had a lower ventral striatal BOLD response to MPFC stimulation. There was no between-group difference, however, in dorsal striatal response to DLPFC stimulation. Consistent with the competing neurobehavioral decision system (CNDS) theory, DLPFC stimulation led to a reciprocal decrease in MPFC activity (BA 10) in the controls. This reciprocal pattern did not exist, however, in the cocaine users.
2. Alcohol
In healthy controls, acute ethanol exposure leads to an increase in the duration of the cortical silent period (Ziemann et al., 1995; Conte et al., 2008). Despite this acute change, CSP is not altered in individuals who have consumed alcohol chronically (Conte et al., 2008; Nardone et al., 2010b; Naim-Feil et al., 2016), even during withdrawal states (Nardone et al., 2010b). Similarly, inhibition can be enhanced and facilitation reduced by acute alcohol consumption in healthy controls (Ziemann et al., 1995), but no difference in magnitude could be found among detoxified individuals with a history of chronic alcohol use (Nardone et al., 2010a,b; Naim-Feil et al., 2016), despite facilitation being increased in withdrawal states (Nardone et al., 2010b).
Some groups have used TMS to induce either LTD- or LTP-like plasticity. In healthy controls, acute alcohol exposure enhances LTD-like plasticity (Fuhl et al., 2015) but blocks LTP-like effects (Lucke et al., 2014; Loheswaran et al., 2016). In multiple studies, neither acute alcohol administration nor a history of chronic alcohol use had an effect on resting or active motor threshold (Ziemann et al., 1995; Conte et al., 2008; Nardone et al., 2010a,b; Kaarre et al., 2018), although one group did find a small difference between healthy controls and those with an alcohol use disorder when controlling for depression and intellectual function (Naim-Feil et al., 2016).
The primary findings, that of increased inhibition, reflect GABAergic properties. Similar intracortical inhibition can be seen after administration of a benzodiazepine such as lorazepam, a GABA-A agonist (Di Lazzaro et al., 2005a), and CSP elongation is observed following inhibition of GABA reuptake inhibition with tiagabine (Werhahn et al., 1999) or reduced GABA degradation with vigabatrin (Pierantozzi et al., 2004). Further work will be needed to explore the compensation methods that differentiate acute from chronic changes.
3. Nicotine
Chronic nicotine usage is related with a prolonged CSP duration, compared with healthy controls (Lang et al., 2008). Neither resting motor threshold (rMT) nor active motor threshold measured at the periphery appears to be sensitive to acute (Orth et al., 2005; Grundey et al., 2012a, 2013) or chronic (Orth et al., 2005; Lang et al., 2008; Grundey et al., 2012a; Strube et al., 2015) nicotine usage. rMT measured at the tongue, evaluating corticobulbar rather than corticospinal excitability, did find that chronic smokers had lower motor thresholds (Vicario et al., 2014). Evidence for changes in the recruitment curve is mixed, with chronic smokers showing greater MEPs at 150% of rMT (Grundey et al., 2013), but not 130% (Lang et al., 2008).
No effect of past nicotine usage was found for ICI (Lang et al., 2008; Grundey et al., 2013), although chronic smokers did show reduced ICF (Lang et al., 2008; Grundey et al., 2013). In deprived smokers, this loss of facilitation was restored to healthy control levels following the administration of nicotine (Grundey et al., 2013); however, acute nicotine did not alter ICF in healthy controls, but instead enhanced ICI (Grundey et al., 2013). Deprived chronic smokers showed enhanced short-interval afferent inhibition (SAI) at 20 milliseconds compared with controls, and this was not altered by a nicotine patch (Lang et al., 2008; Grundey et al., 2013). In contrast to this, SAI was enhanced in nonsmokers after a nicotine patch (Grundey et al., 2013).
Depending on the route of administration, nicotine was effective at blocking LTD-like plasticity in healthy controls (Thirugnanasambandam et al., 2011) as well as LTP-like effects (Grundey et al., 2012a). In deprived smokers, nicotine administration restored LTP-like plasticity with no effect on LTD-like plasticity (Grundey et al., 2012b). Further work is needed to determine what role these acute changes play in the addiction process and whether they can be used as an index of recovery or intervention effectiveness.
Together, these results highlight the complex interaction of the cholinergic system with both GABAergic and glutamatergic function. SAI is particularly relevant in studies of nicotine, as it is sensitive to cholinergic function and can be blocked by scopolamine, a muscarinic antagonist (Di Lazzaro et al., 2000b). This is a promising method for future work, as it appears to be sensitive to both acute and chronic nicotine effects.
D. Evaluating Cortical–Striatal Connectivity
In addition to investigating cortical excitability, it is also possible to evaluate cortical–striatal connectivity by combining single pulses of TMS with either functional magnetic resonance imaging (MRI) or EEG. As stated previously, the direct effects of TMS are spatially restricted to cortical areas directly exposed to the TMS-induced field (typically 2–4 cm from the center of the coil). When this depolarizing current is strong enough, however, it is possible to induce activity in monosynaptic targets of the neurons directly affected by the field. In this manner, cortical pulses of TMS can be used to investigate frontal–striatal connectivity. This has been documented using interleaved TMS/BOLD imaging wherein a single pulse of TMS induces an elevation in the BOLD signal in the vicinity of the TMS coil and in monosynaptic target regions (Bohning et al., 1998). For example, when TMS is applied to the motor cortex, the TMS-evoked BOLD signal in the motor system is similar to a task-evoked BOLD signal in the motor system (i.e., instructing the participant to squeeze his or her hand) (Denslow et al., 2005). In one of the first sham-controlled interleaved TMS/MRI studies, Dowdle et al. (2018) demonstrated that, relative to sham TMS, single pulses of real TMS to the DLPFC induce a significant increase in the BOLD signal in the left caudate and the anterior cingulate cortex—regions that both receive axonal projections from the left DLPFC (Fig. 2) (Dowdle et al., 2018). With interleaved TMS/MRI, it is also possible to differentially activate the frontal–striatal circuits involved in limbic versus executive control—neural circuits that are both involved in the substance use and recovery process. Specifically, TMS pulses to the left DLPFC induce an increase in dorsal striatal activity, whereas TMS pulses to the left ventral MPFC (frontal pole) induce an increase in ventral striatal activity (Hanlon et al., 2013b). This technique was originally used to investigate frontal–striatal integrity in healthy controls, and has now been applied to individuals with nicotine, cocaine, and alcohol use disorders. Although this interleaved TMS/MRI imaging technique can be a technological challenge due to physical constraints of having a TMS coil inside a MRI scanner, it provides researchers with a very unique opportunity to evaluate the functional connectivity of specific neural circuits in a causal manner.
Fig. 2.

TMS to the DLPFC induces a direct increase in brain activity (BOLD signal) in the caudate and anterior cingulate cortex. Healthy young adults received 40 single pulses to the left DLPFC at TMS intensities (Multiband T2*, TR = 1.0, TMS pulse fired in 100-ms gap, 10 pulses per dose, randomized order). Sham TMS was delivered by increasing the distance from the scalp via reticulated open-cell foam. Data were analyzed with an event-related design using the general linear model with weighted values for the dose covariate and a canonical hemodynamic response function (SPM 8, image threshold at P < 0.001). Relative to the sham/control condition, TMS to the left DLPFC induced a significant increase in activity in the caudate and the anterior cingulate cortex, but not in the putamen, right DLPFC, or auditory cortex (positive control region) (P < 0.05, corrected) (as described in Dowdle et al., 2018).
III. Repetitive Transcranial Magnetic Stimulation: An Interventional Tool
Whereas the period from 1999 to 2006 was a time of tremendous growth regarding the basic effects of various TMS protocols on motor cortex excitability, from 2006 to the present day, the momentum in rTMS research has shifted toward the clinical utility of rTMS as a tool to treat psychiatric disease. Section Single-Pulse Transcranial Magnetic Stimulation: A Biomarker and Probe focused on the use of TMS as a tool to probe cortical excitability—potentially a biomarker for future drug discovery trials. The remainder of this review will focus on TMS as an interventional tool. In rTMS, single TMS pulses are delivered at various frequencies (typically 1–20 Hz) in either a fixed or bursting pattern, for anywhere from 600 to 4000 pulses per session. These parameters can induce LTP of cortical excitability or LTD of cortical excitability depending on the frequency chosen (reviewed in Fitzgerald et al., 2006; Thickbroom, 2007). The FDA-approved protocol for depression is 10 Hz stimulation to the left DLPFC at 120% of the patient’s resting MT. In standard clinical practice, each session is typically 2000–4000 pulses and occurs 3–5 days per week for 6 weeks. There have been several multisite clinical trials evaluating the safety, efficacy, and durability of rTMS as a treatment of depression. Although the treatment outcomes and durability data are mixed, the effects of 4–6 weeks of treatment appear to last for at least 6 months and may be facilitated by intermittent shorter treatments of TMS. There are now TMS clinics on six of the seven continents—including all 50 states in the United States; nearly all countries in the European Union; established professional networks in Australia and South America; large high volume centers in China, South Korea, and Japan; and emerging clinics in Africa. Although the majority of the research in TMS is focused on optimizing treatment protocols for depression, there has been an exponential growth in the application of TMS to investigate and modulate these networks in populations with substance use disorders.
A. Fundamentals
There are a number of important factors that influence the efficacy of a given rTMS protocol (e.g., frequency, amplitude, location, coil shape, number of pulses, number of sessions). There is general agreement that low-frequency stimulation (e.g., 1 Hz) causes a decrease in cortical excitability, whereas higher-frequency stimulation (e.g., 10, 20 Hz) causes an increase in cortical excitability (review: Fitzgerald et al., 2006).One of the first studies in this field was by Pascual-Leone et al. (1994), who discovered that 20 pulses at 10 and 20 Hz stimulation over the motor cortex produced an increase in the amplitude of the motor-evoked potential, suggesting there are LTP-like effects on cortical excitability. Chen et al. (1997) then tested the hypothesis that 15 minutes of 0.9 Hz TMS stimulation (810 pulses) to the motor cortex would induce a LTD-like effect on motor cortex excitability. This hypothesis was built on preclinical studies that demonstrated that 15 minutes of 1 Hz stimulation induces LTD of activity in slice preparations of the motor cortex, visual cortex, and hippocampus. In a sample of 14 individuals, they demonstrated that 1 Hz TMS to the motor cortex for 15 minutes decreased the motor-evoked potential by 20% for at least 15 minutes after stimulation. Whereas these were some of the first studies, from 1995 to 2006 there were at least 46 other original research reports published evaluating the effects of various frequencies, amplitudes, and pulses on motor cortex excitability (review: Fitzgerald et al., 2006).
These excitatory and inhibitory effects for rTMS can also be achieved through θ burst stimulation (TBS). Built from a preclinical foundation (Bear and Malenka, 1994; Malenka and Bear, 2004), human TMS protocols apply rTMS in a continuous or intermittent bursting pattern (Di Lazzaro et al., 2005b; Huang et al., 2005). With continuous TBS (cTBS), three pulse bursts at 50 Hz are applied at a frequency of 5 Hz. In most protocols, this cycle continues until 600 pulses have been delivered (20 seconds). For intermittent TBS (iTBS), bursts are applied at the same rate (five groups of three pulse bursts per second) for 2 seconds, followed by an 8-second pause. In most protocols this 10-second cycle occurs until 600 pulses have been delivered (190 seconds). When performed over the primary motor cortex, 600 pulses of cTBS inhibit cortical excitability, whereas 600 pulses of iTBS amplify cortical excitability (Huang et al., 2005). The advantage of TBS protocols is that effect sizes comparable to fixed frequency protocols can be achieved significantly faster (1–2 minutes versus 20–30 minutes). These similar effect sizes have also been observed in clinical studies for depression, wherein there was no significant difference in response rates to TMS when the treatment was delivered at 10 Hz (30 minutes/session for 20–30 sessions) versus iTBS (6 minutes/session for 20–30 sessions).
B. Pharmacological Effects of Repetitive Transcranial Magnetic Stimulation
Although there have been many studies that have evaluated the effects of various rTMS protocols on behavior and cortical excitability (via EEG and functional MRI), very little is known about the effects of rTMS on neuropharmacology.
1. Dopamine
The most cited studies in this domain have been done using positron emission tomography (PET) imaging, wherein the radioligand [11C]raclopride, which is sensitive to D2 dopamine receptors, is used to detect changes in levels of extracellular dopamine after rTMS. In a series of unique and important studies in this field, Strafella et al. have demonstrated the following: 1) 10 Hz rTMS to the left primary motor cortex (which increases cortical excitability) increases dopamine release in the left putamen (Strafella et al., 2003); 2) 10 Hz rTMS to the left DLPFC increases dopamine release in the caudate (Strafella et al., 2001); and 3) cTBS to the left DLPFC (which decreases cortical excitability) decreases dopamine release in the left and right caudate and disrupted performance on a card-sorting task (Ko et al., 2008). Notably, cTBS to the right DLPFC did not change dopamine release nor affect task performance, suggesting a hemispheric asymmetry in rTMS effects. Additionally, when using the radioligand [11C]FLB457, which is a high-affinity D2 ligand sensitive to extrastriatal dopamine, 10 Hz rTMS to the left DLPFC increased dopamine release in the anterior cingulate and medial prefrontal cortex (Cho and Strafella, 2009). Again, 10 Hz TMS to the right DLPFC did not change dopamine release. This hemispheric asymmetry is consistent with clinical studies of rTMS for depression, which typically observe that whereas 10 Hz TMS to the left DLPFC (which increases cortical excitability) can be effective, 1 Hz TMS is typically used when targeting treatment to the right DLPFC (which decreases cortical excitability).
2. Glutamate and GABA
The rTMS-induced dopamine release is most likely due to direct effects of rTMS on glutamatergic pyramidal cells that originate at the site of stimulation and project to the striatum. One of the first studies to investigate the effects of single and multiple sessions of rTMS on neurochemistry was performed by Michael et al. in 2003 using proton magnetic resonance spectroscopy. A small group of healthy individuals was assigned to receive real (n = 7) or sham (n = 5) 20 Hz rTMS to the DLPFC. They demonstrated that both one and five sessions of rTMS increased glutamate/glutamine levels relative to sham rTMS. They also observed a rate-dependent effect of rTMS on glutamate/glutamine changes, wherein individuals with low baseline concentrations had highest levels of real TMS-associated increases in glutamate/glutamine (Michael et al., 2003). In addition to glutamate changes, several studies have also demonstrated that rTMS has direct effects on cortical GABA (Iwabuchi et al., 2017; Stagg et al., 2009; Vidal-Pineiro et al., 2015). Stagg et al. (2009), for example, demonstrated that the attenuating effects of cTBS on cortical excitability are related to an increase in GABA at the area of stimulation rather than a change in glutamate (Stagg et al., 2009). Iwabuchi et al. (2017) demonstrated that iTBS to the DLPFC (which typically increases cortical excitability) decreases the GABA/glutamate ratio in both the DLPFC and the insula (Iwabuchi et al., 2017).
3. Serotonin
Although the primary clinical indication for rTMS is major depressive disorder, a disease that is typically treated with selective serotonin reuptake inhibitors, very little is known about the effects of rTMS on serotonin. One of the few clinical studies done in this area used 123I-5-I-R91150 single-photon emission-computed tomography to evaluate the effect of 10 sessions of 10 Hz left DLPFC rTMS on postsynaptic 5-HT(2A) receptor binding (Baeken and De Raedt, 2011). Relative to controls (n = 21), at baseline the depressed patients had lower 5-HT(2A) receptor binding in the DLPFC bilaterally and higher binding in the hippocampus. Following 10 sessions of 10 Hz rTMS to the left DLPFC, there was a positive correlation between the improvement in depressive symptoms (measured by the Hamilton depression rating scale) and the change in 5-HT(2A) receptor binding in the DLPFC. This relationship between baseline 5-HT(2A) receptor binding and clinical response to rTMS is aligned with evidence that 5-HT(2A) receptor binding is also related to treatment response with selective serotonin uptake inhibitors (Zanardi et al., 2001).
IV. Developing Repetitive Transcranial Magnetic Stimulation as a Treatment Tool for Addiction
In the last 5 years, there has been an exponential growth in the number of published manuscripts and review articles evaluating rTMS as a tool to decrease craving (Feil and Zangen, 2010; Wing et al., 2013; Bellamoli et al., 2014; Gorelick et al., 2014; Hanlon et al., 2015b). Of these studies, the majority have applied high-frequency stimulation (10, 20 Hz, iTBS) to the left DLPFC, the same target used in depression. Although this may be valuable, as both diseases share common deficits in executive control (which is largely modulated by the DLPFC), scientifically, it is not obvious that the standard treatments for depression will be the optimal target sites for drug and alcohol use. In the following sections, we will review the body of literature that exists to date regarding the effects of TMS as a tool to change behavior in populations with alcohol use disorder, tobacco use disorder, and cocaine use disorder. We will then propose a conceptual model (section Moving Forward— Important Methodological Considerations) for TMS treatment development moving forward, and propose some novel biologic targets for future treatment development.
Many compounds have been evaluated as potential pharmacotherapies for substance use disorders. As of early 2018, however, there is only a handful of FDA-approved medications. Most pharmacotherapeutic strategies fall within three main categories: full agonists, partial agonists, and antagonists. Full agonists, such as methadone, directly stimulate receptor sites and are used as a replacement for the abused drug of choice. Full agonists are taken daily to avoid craving and withdrawal that can lead to relapse. Partial agonists, such as buprenorphine, are often used for detoxification and as a maintenance medication for those at risk of relapse into physical dependence. Antagonists, such as naltrexone, block some of the rewarding aspects of alcohol, making it easier to reduce consumption by blocking opioid receptors.
A. State of the Science in Alcohol
1. Approved Pharmacotherapy
The pharmacotherapy of alcohol dependence is broken into therapies for acute alcohol withdrawal syndrome and therapies for relapse prevention and harm reduction. For acute alcohol withdrawal syndrome, benzodiazepines are safe, effective, and the preferred treatment. Benzodiazepines are thought to increase brain GABA levels via GABA A binding that are systemically decreased with chronic alcohol exposure (Sachdeva et al., 2015). All subclasses of benzodiazepines have shown to be effective in treating alcohol withdrawal syndrome, with the most commonly used being the longer-acting chlordiazepoxide and diazepam or the shorter- to intermediate-acting lorazepam and oxazepam. There are three pharmacotherapies FDA approved for the treatment of alcohol dependence. Disulfram has been used for relapse prevention for over 50 years and works by inhibiting the enzyme acetaldehyde dehydrogenase, resulting in unpleasant effects (headache, nausea, vomiting, chest pain, etc.) when combined with even a small amount of alcohol. This extinguishing effect of drinking behavior is effective, although patients are often not compliant. The FDA-approved opiate antagonist, naltrexone, has shown promise in relapse prevention, stemming the endorphin-mediated reinforcing effects of drinking alcohol. There is strong evidence that naltrexone significantly reduces alcohol relapse to heavy drinking, the frequency and quantity of alcohol in those who do drink, and alcohol craving. Naltrexone also comes in long-acting, one-time-per-month injectable form to help with compliance. Acamprosate is the third medication to receive FDA approval for post withdrawal maintenance of alcohol abstinence. It is thought to reduce craving for alcohol by modulating GABA and glutamate systems that are disrupted by chronic alcohol use.
2. Brain Stimulation Research
At the time of writing, there have been nine studies published on the use of rTMS for alcohol addiction (Table 2). Six of these studies targeted the DLPFC, with five delivering stimulation to the right side and one targeting the left. Two recent studies chose the MPFC as an alternative target. High-frequency stimulation (10 or 20 Hz) was used for all studies that targeted the DLPFC, and for one of the studies targeting the MPFC. The remaining MPFC study used cTBS. Most of these studies (n = 6) evaluated outcomes after 10 or more sessions. The optimal parameters, such as the number of sessions or target, are currently unknown. One difficulty is the large number of factors that may modulate TMS efficacy. For example, a study in 2016 used 20 Hz right DLPFC stimulation and found that the change in cue reactivity depended on the cue reactivity as measured before TMS was delivered (Herremans et al., 2016). These rate-dependent effects may represent a biomarker that can guide treatment targets or inform the interpretation of results. Another such biomarker can be derived from functional imaging of cue induction, which consistently highlights the role of the MPFC in cue reactivity (Schacht et al., 2013). By targeting this region with a single session of cTBS, Hanlon et al. (2017) found reductions in evoked responses in orbitofrontal and insular cortex relative to sham stimulation. (More studies will be needed to determine how these factors contribute to the effectiveness of rTMS.)
TABLE 2.
Studies that have used repetitive TMS as a tool to change alcohol craving
Multiple sessions were given in a single day. This study performed 10 Hz TMS on the superior frontal gyrus and found an increase in craving but did not find a decrease in craving with 1 Hz to the superior frontal gyrus (L, left; R, right).
| Author | Sample Size | Site of TMS | Frequency | Sessions | Behavioral Effect? | Active Sham Control |
|---|---|---|---|---|---|---|
| De Ridder et al. (2011) | 1 | ACC | 1–35 Hz | 3 mo | Yes | No |
| Herremans et al. (2012) | 15 | R DLPFC | 20 Hz | 1 | No | Between groups |
| Herremans et al. (2013) | 29 | R DLPFC | 20 Hz | 2 | No | Within subject |
| Hoppner et al. (2011) | 10 | L DLPFC | 20 Hz | 10 | No | Between groups |
| Mishra et al. (2010) | 30 | R DLPFC | 10 Hz | 1 | Yes | Between groups |
| Ceccanti et al. (2015) | 18 | dMPFCa | 20 Hz | 10 | Yes | Between groups |
| Herremans et al. (2015) | 26 | R DLPFC | 20 Hz | 15 (over 4 days) | No | No |
| Herremans et al. (2016) | 19 | R DLPFC | 14 (over 3 days) | Rate-dependent change (ACC) | ||
| Hanlon et al. (2017) | 25 | L MPFC | cTBS | 1 (3600 pulses)z | No | Yes, between |
Studies used H-coil TMS devices (Brainsway, Jerusalem, Israel). This deep TMS coil geometry has a very different field distribution than the typical figure of eight coils.
B. State of the Science in Cocaine
Despite many years of research into the development of an effective medication for cocaine dependence, there still is no medication that has emerged as consistently effective. Research has explored a variety of neurobiological targets, including dopamine, serotonin, GABA, norepinephrine, and glutamate.
1. Brain Stimulation Research
To date, there have been seven published reports on the effects of rTMS for cocaine use disorder (Table 3). The majority of these studies have targeted the left DLPFC (n = 4), with one targeting the prefrontal cortex bilaterally. Two have chosen the MPFC as a target. Early studies evaluated the effects of just one session, whereas more recent work has explored outcomes after 8–12 sessions. All studies that stimulated the DLPFC, whether right, left, or both, have used high-frequency TMS (10 or 15 Hz). For studies that chose the MPFC as a target, cTBS was used. Although the exact parameters have yet to be determined, there are promising findings. For example, Terraneo et al. (2016) reported that left DLPFC stimulation alone led to a significantly greater number of cocaine-free urine tests compared with a control group that used pharmacotherapies. As in alcohol research, the MPFC is being explored as an alternative to the conventional DLPFC target. Hanlon et al. (2015b) reported that a single session of cTBS led to reduction in the stimulus evoke BOLD signal in the orbitofrontal cortex and striatum. Together these findings suggest that rTMS may be an alternative treatment or adjunct to a yet-to-be-discovered pharmacotherapy.
TABLE 3.
Studies that have used repetitive TMS as a tool to cocaine or methamphetamine craving
This study performed 10 Hz TMS on the superior frontal gyrus and found an increase in craving but did not find a decrease in craving with 1 Hz to the superior frontal gyrus (L, left; R, right).
| Author | Drug of Abuse | Sample Size | Site of TMS | Frequency | Sessions | Behavioral Effect? | Active Sham Control |
|---|---|---|---|---|---|---|---|
| Camprodon et al. (2007) | Cocaine | 6 | L/R DLPFC | 10 Hz | 1 | Yes | Within subject |
| Hanlon et al. (2015b) | Cocaine | 11 | L vMPFC | cTBS | 6a | Yes | Within subject |
| Hanlon et al. (2017) | Cocaine | 25 | L MPFC | cTBS | 6a | Yes | Within subject |
| Politi et al. (2008) | Cocaine | 36 | L DLPFC | 15 Hz | 10 | Yes | No |
| Rapinesi et al. (2016) | Cocaine | 7 | L DLPFCb | 20 Hz | 12 | Yes | Between groups |
| Bolloni et al. (2016) | Cocaine | 10 | Bilat PFC/Insb | 10 Hz | 12 | No | Between groups |
| Terraneo et al. (2016) | Cocaine | 32 | L DLPFC | 15 Hz | >8 | Yes | No |
| Li et al. (2013b) | Meth. | 10 | L DLPFC | 1 Hz | 1 day | No | Within subject |
PFC, prefrontal cortex.
Multiple sessions were given in a single day.
Studies used H-coil TMS devices (Brainsway, Jerusalem, Israel). This deep TMS coil geometry has a very different field distribution than the typical figure of eight coils.
C. State of the Science in Nicotine
1. Approved Pharmacotherapy
There are three FDA-approved medications used for smoking cessation. Nicotine replacement therapies such as transdermal patches, gum, nasal spray, inhaler, and lozenges offer the patient a safer and therapeutically manageable form of nicotine to alleviate the symptoms of withdrawal and craving. One drawback with nicotine replacement therapies is the potential to become dependent on the replacement if the patient does not wean as directed. The antidepressant, bupropion, acts as a neuronal reuptake inhibitor of dopamine and noradrenalin and has been shown to be efficacious as a first-line treatment of nicotine dependence (Hurt et al., 1997). Varenicline is a partial agonist that has high and selective activity at α4-β2 receptor. It has been FDA approved due to its efficacy in reducing craving and withdrawal symptoms during abstinence and blocking the reinforcing effects of nicotine (Coe et al., 2005; Gonzales et al., 2006; Jorenby et al., 2006).
2. Brain Stimulation Research
Since 2003, there have been 10 studies on rTMS and cigarette smoking (Table 4). As with other drugs of abuse, the DLPFC is the primary target (six left, one right, one both). The insula and superior frontal gyrus have been targeted in one study each. Each left DLPFC study chose to use high-frequency stimulation (10–20 Hz). Half of these studies used more than one session. In the largest randomized controlled trial to date, Dinur-Klein et al. (2014) delivered 13 sessions of either high- or low-frequency rTMS bilaterally to the lateral prefrontal cortex and insulae, with and without cue exposure immediately before stimulation. High- but not low-frequency TMS was effective in reducing the number of cigarettes smoked. Individuals randomized to the high-frequency and cue-exposed condition showed the largest reduction in cigarettes smoked. These context-dependent effects may have large consequences on the efficacy of rTMS and should be considered as an additional parameter for optimization. Although cue induction is common in the addiction field, and can lead to reliable limbic system activation, there are a number of alternatives. One example is executive control circuitry, as engaging this before or during treatment may also be effective. Toward this end, Sheffer et al. (2018) delivered eight sessions of self-help therapy combined with active rTMS or sham TMS. Despite the use of therapy in both groups, active rTMS showed significantly greater abstinence groups relative to sham. These findings add to the number of parameters that must be explored when designing a TMS trial; however, they both suggest that there is an opportunity to improve TMS treatment outcomes.
TABLE 4.
Studies that have used repetitive TMS as a tool to change nicotine craving
Multiple sessions were given in a single day. This study performed 10 Hz TMS on the superior frontal gyrus (SFG) and found an increase in craving but did not find a decrease in craving with 1 Hz to the SFG (L, left; R, right).
| Author | Sample Size | Site of TMS | Frequency | Sessions | Behavioral Effect? | Active Sham Control |
|---|---|---|---|---|---|---|
| Amiaz et al. (2009) | 22 | L DLPFC | 10 Hz | 10 days | Yes | Between groups |
| Dinur-Klein et al. (2014) | 115 | Bilat PFC/Insa | 10 Hz | 13 | Yes | Between groups |
| Dinur-Klein et al. (2014) | 115 | Bilat PFC/Insa | 1 Hz | 13 | No | Between groups |
| Eichhammer et al. (2003) | 14 | L DLPFC | 20 Hz | 4 days | No | Within subject |
| Li et al. (2013a) | 16 | L DLPFC | 10 Hz | 2 days | Yes | Within subject |
| Pripfl et al. (2014) | 14 | L DLPFC | 10 Hz | 1 | Yes | Within subject |
| Rose et al. (2011) | 15 | SFG | 10 Hz | 3 days | Yes | Within subject |
| Rose et al. (2011) | 15 | SFG | 1 Hz | 3 days | No | Within subject |
| Trojak et al. (2015) | 37 | R DLPFC | 1 Hz | 10 | Yes | Within subject |
| Sheffer et al. (2018) | 29 | L DLPFC | 20 Hz | 8 | Yes | Within subject |
| Li et al. (2017) | 10 | L DLPFC | 10 Hz | 1 | Yes | Within subject |
PFC, prefrontal cortex.
Studies used H-coil TMS devices (Brainsway, Jerusalem, Israel). This deep TMS coil geometry has a very different field distribution than the typical figure of eight coils.
D. State of the Science in Opiates
1. Approved Pharmacotherapy
There are two general pharmacotherapy paths for opioid dependence, opioid maintenance treatment, or detoxification. Agonist and partial agonist medications are used for both maintenance and detoxification: α-2-adrenergic agonists used to enhance detoxification outcomes; antagonist medications used to accelerate the detoxification process. Methadone is a μ-opioid receptor agonist that is typically used as a replacement therapy for opioid dependence. Its long half-life (24–36 hours) allows it to be used as either a detoxification agent or for maintenance therapy (Ward et al., 1996). Buprenorphine is a partial agonist of μ-opioid receptors that has a slow onset and long duration, allowing for alternate day dosing for opioid maintenance treatment. As a partial agonist, it has a more reduced risk of unintentional overdose than methadone, but this may also reduce buprenorphine’s potential efficacy. A number of studies have shown the efficacy of buprenorphine over placebo for opioid maintenance. Mattick et al. (2004) showed that both methadone and buprenorphine are effective at decreased illicit opiate use and increased retention rates when higher doses of methadone (>60 mg) and buprenorphine (>8 mg) are used. The third FDA-approved pharmacotherapy for opioid dependence is the opioid antagonist naltrexone used for opioid maintenance. Naltrexone is safe, well tolerated, and effective at blocking the reinforcing properties of opiates. Unfortunately, in an analysis of 10 studies, Minozzi et al. (2011) showed weak support for the efficacy of naltrexone.
2. Brain Stimulation Research
In contrast to the variety of pharmacological treatments available for opiate dependence, there is a paucity of published reports on the effectiveness of rTMS. To the best of our knowledge, the only report of rTMS in opiate dependence is a short correspondence on 20 male heroin users. In this, Shen et al. (2016) reported a reduction in cue-induced craving after a single, 10-minute session of high-frequency stimulation to the left DLPFC, with further effects after an additional four treatments. Although the literature is currently limited, it is likely that this will soon change as a number of clinical trials are currently ongoing. Although present findings specific to opiate addiction are limited, there is a large body of evidence that rTMS is effective for reducing pain (for a review, see Galhardoni et al., 2015). Although many of these studies have focused on using the motor cortex as a target for stimulation, there is evidence that the DLPFC is also an effective target for pain reduction (Borckardt et al., 2007, 2008; Brighina et al., 2011; Taylor et al., 2013). This overlap represents a clinically meaningful and treatable target for both pain disorders and substance abuse. This is particularly important, considering opiate use disorders, which can be initiated or maintained by the presence of pain (Passik et al., 2006; Back et al., 2011; Barth et al., 2013; Lusted et al., 2013).
V. Moving Forward—Important Methodological Considerations
A. Conceptual Model
Despite the variety of TMS parameters used across different substance use disorders, a few clear trends have emerged. The DLPFC, whether left or right, is the most common target, although early results support alternative targets such as the MPFC or insula. Additionally, there is a growing interest in the context of TMS delivery, which includes current craving levels, cue induction, or executive control engagement. To make significant progress in TMS treatment development is critical to start from a robust conceptual model, which addresses the role of both the limbic system and the executive control system in maintaining successful abstinence in treatment-seeking individuals. Previous studies have demonstrated that high rates of relapse may be due to dysfunction in at least two neural circuits: 1) elevated functional activity within limbic neural circuitry, in the presence of a salient cue (Moeller et al., 2001; Ersche et al., 2011) (including the MPFC and ventral striatum) or 2) depressed activity in executive control circuitry (Goldstein et al., 2004; Kubler et al., 2005; Moeller et al., 2010), which can counteract limbic drive (including the DLPFC and dorsal striatum) (Fig. 3). One model that addresses both of these aspects is the CNDS theory. The CNDS theory poses that, in a pathologic drug use state, there is elevated activity in frontal–striatal circuits involved in limbic reward and impulsive action, as well as relatively lower activity in executive control circuitry. Intervention efforts could therefore be directed at either decreasing impulsive reward circuit activity to cues or increasing the executive control circuit activity to cues. This model logically extends to at least two potential strategies for improving treatment outcomes in patients, as follows: strategy 1, increase executive control network, or strategy 2, decrease craving/cue-reactivity networks through ventromedial prefrontal cortex (vMPFC) or orbital frontal cortex (OFC) TMS. Whereas most of the studies to date have evaluated strategy 1, the importance of strategy 2 is underscored by the transdiagnostic relevance of cue reactivity as a predictive indicator of relapse.
Fig. 3.

Frontal–striatal circuits that contribute to vulnerability to cues and brain stimulation strategies to modulate these circuits.
Many studies investigating cue reactivity in either cocaine-, nicotine-, or alcohol-dependent populations have independently demonstrated that drug cues evoke elevated activity in the medial prefrontal cortex, anterior cingulate, and insula cortex. Most of these studies focus on one substance-using class, however, and therefore it is difficult to distinguish which aspects of cue reactivity are transdiagnostic biomarkers of the addiction process versus those that are specific to alcohol, cocaine, or nicotine dependence. Several retrospective meta-analyses have demonstrated that the medial prefrontal cortex and cingulate cortex are reliably activated to drug cues (Schacht et al., 2013). Other meta-analyses have demonstrated that activity in these brain regions may predict relapse across multiple substances (Killen and Fortmann, 1997; O’Connell et al., 2011; Courtney et al., 2016).
B. Choosing a Target
The DLPFC and vMPFC are only two of many potential treatment targets for TMS treatment development in addiction. The prefrontal cortex is often divided into six discrete areas (known as Brodmann areas 9, 10, 11, 12, 46, and 47), which were defined in 1909 by German scientist Korbinian Brodmann. For example, Brodmann areas 9 and 46, often referred to as the DLPFC, contribute to executive processing tasks, including planning future events, decision making, and the coordination of purposeful behavior (Miller and Cohen, 2001). Brodmann areas 10, 11, 12, and 47 are each involved in reward signaling, assigning value, arousal, and emotional processing (Goldstein and Volkow, 2002). Together, these prefrontal regions all play key roles in addiction disorders.
1. Dorsolateral Prefrontal Cortex
The importance of the DLPFC in conscious decision making, reasoning, working memory, inhibition, as well as outcome prediction (see Krawczyk, 2002), and its established reputation as a successful treatment target for TMS in patients with depression, all suggest that the DLPFC is most likely a very good candidate for TMS treatment development in substance use disorder. The DLPFC can be directly stimulated with traditional figure-of-eight TMS coils and circular TMS coils.
The majority of studies in TMS to date have focused on this brain region. The biggest trial to date was a cohort study of 115 treatment-seeking smokers randomized to receive 13 sessions of 10, 1 Hz, or sham TMS after viewing smoking cues. An H-coil (which induces a wider, deeper electromagnetic field in the brain than a figure-of-eight coil) focused on the left insula and lateral prefrontal cortex was used for this experiment. Ten hertz (LTP-like) rTMS decreased cigarette consumption and improved 6-month abstinence rates (Dinur-Klein et al., 2014). These data complement other evidence that the insula can be modulated by TMS (Gratton et al., 2013), and that functional connectivity (Moran-Santa Maria et al., 2015) and anatomic integrity (Naqvi et al., 2007) of the insula are critical for craving. A greater understanding of the neurobiological effects of these brain stimulation tools and protocols may allow us to more effectively translate preclinical knowledge into efficient, evidence-based treatments for patients.
2. Ventral Medial Prefrontal Cortex and Orbitofrontal Cortex
The MPFC extends throughout the medial section of the frontal cortex and into the OFC (BA 10). The ventral aspects of the MPFC typically are involved in mediating arousal, whereas the dorsal aspects of the MPFC play an important role in the cognitive decrements seen in individuals with substance use disorders (Bolla et al., 2003; Kaufman et al., 2003; Kubler et al., 2005). The OFC is located on the ventral section of the PFC and contributes to reinforcement of cocaine (e.g., with nucleus accumbens, thalamus, ventral striatum, and indirectly with amygdala, cingulate, hippocampus). Acute cocaine administration, for example, leads to elevated activity in the OFC, which may act as an information integrator, leading to reinforcement of drug taking (Volkow and Fowler, 2000). The OFC is involved in response inhibition and learning the association between a stimulus and outcome (Kringelbach and Rolls, 2004). Dysregulation of the OFC most likely perpetuates habitual drug use behavior as well as risky decision making, an inability to anticipate outcomes (see Krawczyk, 2002).
3. Relationship to Transcranial Magnetic Stimulation Treatment Development
The primary cortical inputs to the ventral striatum are the medial and orbital prefrontal cortices. Given that the ventral striatum is one of the primary brain regions involved in processing drug-associated reward, it seems that targeting the MPFC would be a more direct method to modulate ventral striatal activity than DLPFC stimulation. Specifically, a single pulse of TMS to the MPFC in healthy individuals leads to an increase in BOLD signal in the ventral striatum (Hanlon et al., 2013b). A recent study in cocaine users demonstrated that they have a hyperactive BOLD response in the dorsal and ventral striatum relative to controls (Hanlon et al., 2016). This elevated ventral striatal sensitivity following MPFC stimulation, a frontal–striatal circuit involved in the limbic aspects of craving, may be a prime circuit to attenuate to make these individuals less vulnerable to drug-related cues. Given that craving is associated with an increase in activity in the striatum (e.g., BOLD signal), LTD-like rTMS over the MPFC may attenuate activity in this neural circuit.
Our laboratory has spent the last 7 years systematically developing a TMS as an empirically derived treatment of cocaine use disorder and alcohol use disorder, wherein the ventral MPFC and OFC are the prime targets of interest. We have demonstrated that it is possible to differentially activate the vMPFC from the DLPFC using interleaved TMS/functional MRI (Hanlon et al., 2013b); shown that connectivity with the vMPFC was significantly different in the cocaine users relative to healthy controls and DLPFC connectivity was similar to controls (Hanlon et al., 2016); evaluated the feasibility of a vMPFC-focused treatment strategy for TMS (Hanlon et al., 2015b); and demonstrated that continuous TBS could dampen activity in the vMPFC, insula, and cingulate cortex (Hanlon et al., 2017). This attenuation was present in both the cocaine users (Fig. 4) and the alcohol users.
Fig. 4.

The effect of frontal pole cTBS on brain reactivity in cocaine users and alcohol users (adapted from Hanlon et al., 2015, 2017). Interleaved TMS/BOLD imaging was used to measure TMS-evoked BOLD signal immediately before and after cocaine users (B) and heavy alcohol users (C) were given a dose of cTBS to the left frontal pole (A). The TMS coil was placed over FP1 (EEG:10-20 system) for both the interleaved TMS/BOLD scan (left and right panel) and the cTBS session (center panel). The red area represents the region of interest to which the coil is targeted (AAL: left superior and middle orbital prefrontal cortex inferior to the anterior commissure). Real cTBS (LTD-like) led to a significant decrease in BOLD signal in the left orbital/medial prefrontal cortex and ventral striatum (SPM 8, P < 0.05 family wise error correction, negative Z-scores shown). The cTBS protocol was two trains of 1800 pulses, 110% rMT, 60-second intertrain interval, intensity ramped from 80% to 110% over first 30-second L = left hemisphere. These data reveal that 3600 pulses of real cTBS to the left frontal pole decrease stimulus-evoked activity in the MPFC, cingulate, and striatum of cocaine users and MPFC and insula of alcohol users significantly more than sham cTBS.
4. Anterior Cingulate Cortex
The anterior cingulate cortex (ACC) is part of the limbic system and is involved in attention, emotion regulation, error monitoring, inhibitory control, and motivation (Bush et al., 2000). Relative to nondrug-using controls, cocaine users have lower rates of glucose usage in the ACC and OFC as measured with fluorodeoxyglucose and PET (Volkow et al., 1991, 1992, 2005; Goldstein and Volkow, 2002; Goldstein et al., 2004). These depressed rates of functional activity have been reported to persist for up to 3 (Volkow et al., 1993) to 5 months (Hanlon et al., 2013a) of abstinence. The ACC has increased activity during cocaine craving following administration (Breiter et al., 1997; Garavan et al., 2000; Maas et al., 1998; Risinger et al., 2005), and its activity is negatively correlated with reports of cocaine high. There is, however, a positive correlation with rush (Breiter et al., 1997). Cocaine users have decreased activity relative to controls in the ACC when completing a task requiring cognitive inhibition (Kaufman et al., 2003; Hester and Garavan, 2004) and visual attention (Kubler et al., 2005). From a structural perspective, cocaine users have decreased gray matter density in the ACC compared with controls (Matochik et al., 2003), and this reduction has been found to be greater for those with a longer history of abuse (Franklin et al., 2002; Ersche et al., 2011).
Additionally, activity in the ACC has been consistently linked to an enhanced attentional bias to smoking cues (Janes et al., 2009), alcohol cues (Vollstadt-Klein et al., 2012), and cocaine cues (Goldstein et al., 2009). Attentional bias is one of the most studied cognitive processes in addiction, as it is often linked to drug craving and relapse (Field and Cox, 2008). Several studies have, in fact, demonstrated that higher levels of ACC activity during an attentional bias task predict higher relapse probability in smokers (Janes et al., 2010) and cocaine users (Marhe et al., 2013).
5. Insula
The insula is a cortical structure located deep in the brain between the frontal and the temporal lobes, which has four subregions that are defined based on their cytoarchitecture: posterior/dorsal insula (granular), intermediate insula (dysgranular), anterior/ventral insula (agranular), and the fronto–insular cortex (agranular), which is only present in humans and great apes. Anatomical, visual, and functional imaging studies have shown that the fronto-insular and anterior subregions of the insula are extensively connected with the orbitofrontal cortex, anterior cingulate cortex, and the dorsal lateral prefrontal cortex (Bauernfeind et al., 2013). From a functional perspective, there appears to be laterality within the insula wherein the right anterior insula is elevated by negative emotional valence and sympathetic activation, whereas the left anterior insula is associated with positive affect and parasympathetic activation (Craig et al., 2000; Critchley et al., 2004).The insula has dense structural connectivity with the amygdala, basal ganglia, thalamus, OFC, and PFC, placing it as an important structural and functional hub involved in drug and alcohol use and relapse (see Singer et al., 2009). The insula has elevated activity, for example, in paradigms exploring cocaine craving as well as acute cocaine high (Kilts et al., 2001; Bonson et al., 2002; Risinger et al., 2005). The volume of the insula is also smaller and correlated with length of cocaine user among cocaine users compared with sibling controls (Ersche et al., 2011). Small insula volume is also associated with decrements in attentional control (Ersche et al., 2011; Hanlon et al., 2011). A large study by McHugh et al. (2013) demonstrated that cocaine users (n = 45) had lower connectivity between the insula and putamen than healthy controls (n = 22). They also showed increased impulsivity in cocaine-addicted individuals that was mediated by reduced insula–putamen connectivity.
6. Relationship to Transcranial Magnetic Stimulation Treatment Development—Anterior Cingulate Cortex and Insula
The ACC and the insula are nodes in the Salience network, a functional connectivity network in the brain which is involved in attentional bias and task switching. Given the importance of these processes to addiction, there is a lot of interest in directly modulating this network with rTMS. Whereas traditional figure-of-eight coils cannot directly reach the insular cortex or the ACC due to the magnetic field decay. It is possible to indirectly modulate the insula and ACC by stimulating the DLPFC (Nahas et al., 2001) or the frontal pole (Hanlon et al., 2017). This indirect modulation via both the dorsal lateral and ventral medial aspects of the cortex is due to the very high anatomic connectivity that exists between the anterior insula and these prefrontal regions. It is likely also possible to directly target the ACC and the anterior or posterior insula through novel TMS coil designs such as the H-coil design series. These coils have a unique geometry that enables their direct penetration depth to be as deep as 3–5 cm, which is most likely sufficient to engage the ACC and anterior insula (Deng et al., 2015). Hence, H-coils are actively being evaluated to addiction treatment development (Dinur-Klein et al., 2014). Targeting the insula via cTBS treatment of the MPFC may reduce the activation when experiencing cocaine craving and high shown by Bonson et al. (2002) and Kilts et al. (2001).
C. Combining Pharmacology with Brain Stimulation
In section Single Pulse Transcranial Magnetic Stimulation: A Biomarker and Probe, we discussed ways in which TMS has been used as a tool to evaluate the effect of various pharmacotherapies on the brain. In section Repetitive Transcranial Magnetic Stimulation: An Interventional Tool, we discussed ways in which pharmacologically sensitive tools (e.g., PET and spectroscopy) have been used to evaluate the effect of rTMS on the brain. A third, very interesting concept to consider is how these two therapeutic approaches—brain stimulation and pharmacotherapy—might be combined in a synergistic manner. Although there are presently no studies in the drug or alcohol addiction literature that have directly evaluated the combined effects of rTMS and pharmacotherapy, there are several studies in healthy individuals that suggest that combining these two approaches may have supraordinal effects on neuroplasticity (for a review, see Ziemann et al., 2015). In this work, we highlight the possibilities that these combinations represent, but also draw attention to the variety of new challenges that are created.
Ideally a protocol could combine a safe pharmacological treatment with brain stimulation to enhance or stabilize the desired effects. Most of these studies have been done using paired associative stimulation (PAS) and transcranial direct current stimulation (TDCS). PAS and TDCS are unique noninvasive brain stimulation protocols that, like rTMS, can induce increases or decreases in cortical excitability. With PAS, peripheral stimulation is delivered to the median nerve, whereas TMS is delivered to the cortex. With a 10-ms delay between peripheral and cortical stimulation, MEPs are reduced (PAS-LTD) (Wolters et al., 2003); at 25 milliseconds, the resulting MEPs are larger (PAS-LTP) (Stefan et al., 2000). With TDCS, a pair or an array of electrodes is placed on the scalp. The electrodes are typically 2–20 cm apart. One electrode acts as the anode, leading to increased cortical excitability in the underlying cortex, and the other as the cathode, causing inhibition (Nitsche and Paulus, 2000). Several studies using these approaches have demonstrated that D-cycloserine (Nitsche et al., 2004b), amphetamine (Nitsche et al., 2004a), and nicotine (Thirugnanasambandam et al., 2011) may increase the LTP-like effects of brain stimulation. There have not been any studies to date that have evaluated these pharmacotherapies as potential adjuvants to rTMS treatment.
Although these studies show promising synergistic effects with brain stimulation, the majority of pharmacological interventions evaluated to date appear to block the facilitation or inhibition associated with brain stimulation. This includes medications such as dextromethorphan (Liebetanz et al., 2002; Nitsche et al., 2003) diazepam (Heidegger et al., 2010), baclofen (McDonnell et al., 2007), and propranolol (Nitsche et al., 2004a). Although, ideally, we would like to discover a combination of pharmacotherapy and brain stimulation that has additive effects, knowledge about pharmacotherapeutic agents that blunt the effects of rTMS is also very valuable to the field. For example, a treatment-seeking alcohol user who is using diazepam (a benzodiazepine) to treat coexisting anxiety may not receive the same therapeutic effect from rTMS as one of his/her peers who is not taking diazepam. It is unclear, of course, whether these blunting effects apply to all benzodiazepines. Additionally, whereas D-cycloserine appears to increase the effects of TDCS (Nitsche et al., 2004b), another study demonstrated that it reverses the effects of intermittent TBS, causing iTBS to inhibit cortical excitability, rather than facilitate it as would have been predicted (Teo et al., 2007).
Presently there are not enough data combining pharmacology with rTMS to make any definitive conclusions. Further work is also required to determine whether these early results extend beyond the scope of motor cortex stimulation. In the near future, we are optimistic that significantly more attention will be given to multimodal approaches to neurologic and psychiatric treatment—harnessing both pharmacotherapy and brain stimulation interventions. By combining these approaches with magnetically sensitive nanoparticle technology, it may even be possible to move toward a drug delivery platform that uses targeted brain stimulation focus pharmaceutical delivery to a specific neural circuit. There are, of course, still many wide gaps in our knowledge that need to be filled before this type of precision-medicine approach can be reached.
D. Combining Cognitive Training/Therapy with Brain Stimulation
This review has focused on potential neural targets for rTMS development and the pharmacological effects of rTMS. Another critical aspect of TMS treatment development for substance use disorder is if, and how, to combine it with existing behavioral therapies for substance use disorder. At the end of the day, all treatment development approaches for substance use need to focus on changing behavior. Whereas changing drug-taking behavior (e.g., cigarette abstinence, reduction in alcohol drinking) is the dependent measure currently deemed most important for FDA approval, there are a number of other critical, disabling behavioral hallmarks of the addiction recovery process that clinical treatment researchers are investigating. These include impulsivity, craving, cognitive set shifting, and assigning value to future rewards (e.g., delayed discounting). These behavioral domains are often described as transdiagnostic phenotypes of addiction and have been evaluated in both clinical and preclinical substance use research (Garavan and Hester, 2007; Beveridge et al., 2008; Stalnaker et al., 2009). Moving forward it will be important to identify which of these behavioral phenotypes are most responsive to various forms of pharmacotherapy, behavioral retraining therapy, and ultimately brain stimulation therapy. Through the integration of these components, we will be much more likely to generate individually tailored therapies for individuals that are at various stages of the continuum from drug use vulnerability, to dependence, relapse, and ultimate freedom from powerful, short-term reward of the drug.
1. Task-Dependent Amplification of Transcranial Magnetic Stimulation Effects
One of the most interesting areas of recent methods development in neuromodulation involves choosing a task for the participants to perform (or a thought pattern to engage in) before or during the intervention. Translating established principles of task-dependent plasticity in animals to human clinical experiments, Dinur-Klein et al. (2014) were the first to demonstrate that it is possible to amplify the effects of TMS on smoking cessation by having individuals engage in a smoking cue-reactivity task immediately before the TMS session. In this large, double-blind, sham-controlled study of 115 cigarette smokers, individuals were randomized to receive 10 or 1 real or sham TMS for 13 sessions. Half of these participants were presented with visual cues of smoking for 20 minutes before the TMS session (Brainsway H-ADD coil; 120% resting motor threshold, left insula, and lateral prefrontal cortex). The group receiving 10 Hz TMS (but not 1 Hz) had significantly lower cigarette consumption and nicotine dependence than sham TMS. There was a significantly better outcome in the individuals that had been exposed to the smoking cues as measured by their change in the Fagerstrom test for nicotine dependence score and their abstinence rates (44% at the end of the trial; 33% 6 months later).
These results are comparable to treatment with varenicline, which has a 44% abstinence rate at the end of treatment (12 weeks) and 29.5% 6 months later. TMS treatment in this study appears to be better than treatment with bupropion hydrochloride (Wellbutrin Sustained Release), which has a 29.5% abstinence rate at the end of treatment (12 weeks) and 20.7% 6 months later (Gonzales et al., 2006).
Task-dependent amplification of TMS effects also applies to treating post-traumatic stress disorder (PTSD) (Isserles et al., 2013). In this study, 30 PTSD patients were randomized to one of three groups: sham rTMS, real rTMS following exposure to a 30-second patient-tailored trauma script, or real rTMS following exposure to a 30-second patient-tailored neutral script. The participants received 12 sessions of real or sham rTMS (three sessions/week, 4 weeks, Brainsway H-coil). The only group with a significant improvement in the clinician-administered PTSD scale was the group that received exposure to 30-second trauma scripts before their rTMS treatment. Although the mechanism through which cue exposure enhances the behavioral effects of rTMS is not clear, one possibility is that cue exposure reactivates latent memory traces (frequently referred to as the engram) (Josselyn et al., 2015), enabling them to be manipulated and reconsolidated (Dudai and Eisenberg, 2004; Dudai, 2006). To our knowledge, this has not been directly evaluated in patients with either substance dependence or PTSD.
Based on these empirical results from clinical studies and a strong preclinical foundation for task-dependent plasticity, all experiments in our brain stimulation laboratory now use a standardized behavioral prime, which is given before and during the brain stimulation intervention. For our present investigations using the vMPFC as a target, this behavioral prime includes a standardized drug-cue exposure paradigm based on standardized exposure therapy treatment questions (Concurrent Treatment of PTSD and Substance Use Disorder Using Prolonged Exposure Handbook). Specifically, participants are prompted to describe their most recent (index) drug use event (e.g., day of the week, time of the day, people they were with, and the sensory aspects of the experience). For some of our studies, individuals are then prompted: “During the stimulation, it is important for you to try to focus on (insert their drug of choice: cocaine, crack, alcohol, liquor, beer, wine, cigarettes, painkillers). This tablet will be displaying various pictures of (insert: cocaine/crack, alcohol drinking, cigarettes, painkillers). We would also like you to keep your eyes open, look at the screen, and think about your most recent experience with (insert: cocaine/crack, alcohol drinking, cigarettes, painkillers).” For other protocols, especially those in which we are using the DLPFC as our target (based on a conceptual framework of increasing cognitive control), they are prompted with statements that they are taught in cognitive behavioral therapy as they view the drug cues. Although the cues are routinely tailored to the patient group, and will change based on conceptual strategy for a specific TMS protocol, one thing that remains stable is the existence of a behavioral prime. Full details of the standard operating protocol and a description of the questions used to create these participant-tailored drug use scripts are included in prior work (Hanlon et al., 2017). In the future we anticipate that cutting-edge brain stimulation laboratories and clinics will capitalize on the increasingly widespread availability of virtual reality gaming environments that could be customized and adapted to treatment-seeking drug users and drug use treatment.
E. Summary
TMS can be used as both a probe of cortical excitability and an interventional tool to change neural excitability. When applied to addiction treatment development, TMS may be an innovative way to examine the effects of single or combinations of various pharmacotherapeutic agents on cortical excitability. Given that the chemical composition of the cortex is dynamic and heterogenous, it may be useful to use TMS as a tool to quantify the effects of these novel therapies on excitatory or inhibitory tone in the cortex—an opportunity that is particularly appealing when the effects are combined with various neuroimaging approaches such as interleaving TMS and MRI or interleaving TMS with magnetic resonance spectroscopy.
As an interventional tool, we are most likely only in the very first chapter of our education on its utility as a treatment tool in various substance use disorders. Building on data from the depression field (for which TMS is currently FDA approved), we are now beginning to build a foundation of knowledge regarding its utility as a tool to change smoking, drinking, and cocaine use behavior. Although there are several concepts of rTMS treatment that are robust and replicable (e.g., regional specificity, depth of the magnetic field, dose-dependent amplification of behavior, polysynaptic engagement, frequency-dependent effects), there are still many key components of TMS treatment development that have not been widely explored (e.g., optimal number of sessions per day, optimal number of sessions total, effects of TMS on drug use behavior when it is applied to areas other than the DLPFC, the use of behavioral primes to amplify TMS treatment effects, the effects of applying TMS before versus after behavioral therapy, the use of TMS to amplify pharmacotherapy treatment). As the field of addiction moves forward with pursuing rTMS as a new tool to modulate craving and the frontal-striatal circuits that contribute to chronic use and relapse, it will be important to consider all of the important factors including but not limited to: optimal anatomical site, frequency of stimulation, and target patient population.
The data presented in this review demonstrate that whereas most of the efforts for rTMS in addiction have been focused on increasing activity in the DLPFC, decreasing activity in the MPFC and ventral striatum may also be a feasible and fruitful target to consider. It seems plausible that either increasing neural firing in the executive control circuit (perhaps via 10 Hz TMS in the DLPFC) or decreasing firing in the limbic circuit in the presence of cues (perhaps via cTBS TMS in the MPFC) may be valuable strategies for decreasing vulnerability to drug- related cues among our patients. Before moving forward with slow and expensive clinical trials, however, it is important to have a comprehensive understanding of limbic and executive circuit functioning in a diverse cross section of patients with drug use disorders. With this knowledge, we will be able to develop circuit-specific treatment strategies for these populations. It will be critical to determine which patients are likely to maximally benefit from TMS and whether strategy 1 or strategy 2 is ideal for a given patient based upon his or her neurobiological and behavioral phenotype.
We have attempted to introduce TMS as an innovative new tool that can modulate brain activity in a circuit-specific, frequency-dependent manner as well as to review the current knowledge regarding the pharmacologic effects of TMS. Although the development of TMS as a new treatment tool for addiction is still in its infancy, we hope we have planted several seeds in the reader’s mind regarding the need to develop a neural circuit–based treatment tool that is available to our patients and the need to increase our knowledge regarding the synergy between pharmacotherapeutics and brain stimulation interventions. Based on a large body of knowledge that suggests frontal–striatal circuit activity is a significant biomarker for predicting treatment outcome, through noninvasive brain stimulation techniques such as rTMS, it may be possible to develop a neural circuit–based treatment of our patients. It is important to note, however, that brain stimulation alone is likely not sufficient for inducing lasting behavioral change. Just as the plasticity potential of a primed neuron is higher than an unprimed neuron, TMS likely has higher efficacy when an individual is engaged in the cognitive process they wish to amplify or attenuate. Hence, TMS is likely to be most effective when combined with a pharmacotherapeutic agent that lowers the threshold for cortical excitability or with behavioral interventions such as cognitive behavioral therapy, exposure therapy, or contingency management. Unfortunately, whereas the aforementioned statements are based on preclinical literature and human learning theory, they have not yet been rigorously evaluated.
As the field continues to grow, we hope to see more interactions between clinical and preclinical neuroscience researchers from both an electrophysiology background and a pharmacology background. Rigorous preclinical TMS-dosing studies in various addiction models are needed to comprehensively evaluate the full parameter space of dosing variables (e.g., optimal stimulation frequency, optimal number of pulses per day, optimal number of days required for a durable effect, optimal stimulation site). With any luck, through the continued refinement of TMS as a tool to modulate neural circuits, we will soon be rigorously evaluating noninvasive brain stimulation as a tool to enhance and refine pharmacotherapeutic development for substance use disorders. Just as Perseus required the aid of Cadmus and Bellerophon to ultimately slay Medusa, and Jason required the band of Argonauts to obtain the quest for the Golden Fleece, the quest for a sustainable addiction treatment solution will undoubtedly require a complementary approach to modifying the pharmacology, neural circuitry, and ultimately the behavioral phenotypes present in our patients.
Acknowledgments
We thank Sarah Hamilton and Dr. Audrey Padula for their assistance and contributions to the editing and formatting of this manuscript.
Abbreviations
- ACC
anterior cingulate cortex
- BOLD
blood-oxygen-level dependent
- CNDS
competing neurobehavioral decision system
- CSP
cortical silent period
- cTBS
continuous TBS
- DLPFC
dorsolateral prefrontal cortex
- EEG
electroencephalography
- FDA
Food and Drug Administration
- ICF
intracortical facilitation
- ICI
intracortical inhibition
- iTBS
intermittent TBS
- LTD
long-term depression
- LTP
long-term potentiation
- MEP
motor-evoked potential
- MPFC
medial prefrontal cortex
- MRI
magnetic resonance imaging
- MT
motor threshold
- OFC
orbital frontal cortex
- PAS
paired associative stimulation
- PET
positron emission tomography
- PTSD
post-traumatic stress disorder
- rMT
resting motor threshold
- rTMS
repetitive TMS
- SAI
short-interval afferent inhibition
- TBS
θ burst stimulation
- TDCS
transcranial direct current stimulation
- TMS
transcranial magnetic stimulation
- VMPFC
ventral medial prefrontal cortex
Authorship Contributions
Participated in research design: Hanlon.
Performed data analysis: Hanlon, Dowdle, Henderson.
Wrote or contributed to the writing of the manuscript: Hanlon, Dowdle, Henderson.
Footnotes
This work was supported by National Institutes of Health National Institute of Drug Abuse [Grants R01DA03661, T32DA007288, and F31DA043330], National Institute of General Medical Science [Grant P20 GM109040-04], and National Institutes of Health [Grants R01DA03661, R21DA041610, P50AA010761, P20GM109040, T32DA007288, F31DA043330, and P20 GM109040].
References
- Amiaz R, Levy D, Vainiger D, Grunhaus L, Zangen A. (2009) Repeated high-frequency transcranial magnetic stimulation over the dorsolateral prefrontal cortex reduces cigarette craving and consumption. Addiction 104:653–660. [DOI] [PubMed] [Google Scholar]
- Back SE, Lawson KM, Singleton LM, Brady KT. (2011) Characteristics and correlates of men and women with prescription opioid dependence. Addict Behav 36:829–834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baeken C, De Raedt R. (2011) Neurobiological mechanisms of repetitive transcranial magnetic stimulation on the underlying neurocircuitry in unipolar depression. Dialogues Clin Neurosci 13:139–145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barth KS, Maria MM, Lawson K, Shaftman S, Brady KT, Back SE. (2013) Pain and motives for use among non-treatment seeking individuals with prescription opioid dependence. Am J Addict 22:486–491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bauernfeind AL, de Sousa AA, Avasthi T, Dobson SD, Raghanti MA, AH Lewandowski, Zilles K, Semendeferi K, Allman JM, Craig AD, et al. (2013) A volumetric comparison of the insular cortex and its subregions in primates. J Hum Evol 64:263–279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bear MF, Malenka RC. (1994) Synaptic plasticity: LTP and LTD. Curr Opin Neurobiol 4:389–399. [DOI] [PubMed] [Google Scholar]
- Bellamoli E, Manganotti P, Schwartz RP, Rimondo C, Gomma M, Serpelloni G. (2014) rTMS in the treatment of drug addiction: an update about human studies. Behav Neurol 2014:815215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beveridge TJ, Gill KE, Hanlon CA, Porrino LJ. (2008) Review: parallel studies of cocaine-related neural and cognitive impairment in humans and monkeys. Philos Trans R Soc Lond B Biol Sci 363:3257–3266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bohning DE, Shastri A, Nahas Z, Lorberbaum JP, Andersen SW, Dannels WR, Haxthausen EU, Vincent DJ, George MS. (1998) Echoplanar BOLD fMRI of brain activation induced by concurrent transcranial magnetic stimulation. Invest Radiol 33:336–340. [DOI] [PubMed] [Google Scholar]
- Bolla KI, Eldreth DA, London ED, Kiehl KA, Mouratidis M, Contoreggi C, Matochik JA, Kurian V, Cadet JL, Kimes AS, et al. (2003) Orbitofrontal cortex dysfunction in abstinent cocaine abusers performing a decision-making task. Neuroimage 19:1085–1094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolloni C, Panella R, Pedetti M, Frascella AG, Gambelunghe C, Piccoli T, Maniaci G, Brancato A, Cannizzaro C, Diana M. (2016) Bilateral transcranial magnetic stimulation of the prefrontal cortex reduces cocaine intake: a pilot study. Front Psychiatry 7:133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonson KR, Grant SJ, Contoreggi CS, Links JM, Metcalfe J, Weyl HL, Kurian V, Ernst M, London ED. (2002) Neural systems and cue-induced cocaine craving. Neuropsychopharmacology 26:376–386. [DOI] [PubMed] [Google Scholar]
- Borckardt JJ, Reeves ST, Weinstein M, Smith AR, Shelley N, Kozel FA, Nahas Z, Byrne KT, Morgan K, George MS. (2008) Significant analgesic effects of one session of postoperative left prefrontal cortex repetitive transcranial magnetic stimulation: a replication study. Brain Stimul 1:122–127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borckardt JJ, Smith AR, Reeves ST, Weinstein M, Kozel FA, Nahas Z, Shelley N, Branham RK, Thomas KJ, George MS. (2007) Fifteen minutes of left prefrontal repetitive transcranial magnetic stimulation acutely increases thermal pain thresholds in healthy adults. Pain Res Manag 12:287–290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boutros NN, Lisanby SH, McClain-Furmanski D, Oliwa G, Gooding D, Kosten TR. (2005) Cortical excitability in cocaine-dependent patients: a replication and extension of TMS findings. J Psychiatr Res 39:295–302. [DOI] [PubMed] [Google Scholar]
- Boutros NN, Lisanby SH, Tokuno H, Torello MW, Campbell D, Berman R, Malison R, Krystal JH, Kosten T. (2001) Elevated motor threshold in drug-free, cocaine-dependent patients assessed with transcranial magnetic stimulation. Biol Psychiatry 49:369–373. [DOI] [PubMed] [Google Scholar]
- Breiter HC, Gollub RL, Weisskoff RM, Kennedy DN, Makris N, Berke JD, Goodman JM, Kantor HL, Gastfriend DR, Riorden JP, et al. (1997) Acute effects of cocaine on human brain activity and emotion. Neuron 19:591–611. [DOI] [PubMed] [Google Scholar]
- Brighina F, De Tommaso M, Giglia F, Scalia S, Cosentino G, Puma A, Panetta M, Giglia G, Fierro B. (2011) Modulation of pain perception by transcranial magnetic stimulation of left prefrontal cortex. J Headache Pain 12:185–191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bunse T, Wobrock T, Strube W, Padberg F, Palm U, Falkai P, Hasan A. (2014) Motor cortical excitability assessed by transcranial magnetic stimulation in psychiatric disorders: a systematic review. Brain Stimul 7:158–169. [DOI] [PubMed] [Google Scholar]
- Bush G, Luu P, Posner MI. (2000) Cognitive and emotional influences in anterior cingulate cortex. Trends Cogn Sci 4:215–222. [DOI] [PubMed] [Google Scholar]
- Camprodon JA, Martínez-Raga J, Alonso-Alonso M, Shih MC, Pascual-Leone A. (2007) One session of high frequency repetitive transcranial magnetic stimulation (rTMS) to the right prefrontal cortex transiently reduces cocaine craving. Drug Alcohol Depend 86:91–94. [DOI] [PubMed] [Google Scholar]
- Ceccanti M, Inghilleri M, Attilia ML, Raccah R, Fiore M, Zangen A, Ceccanti M. (2015) Deep TMS on alcoholics: effects on cortisolemia and dopamine pathway modulation: a pilot study. Can J Physiol Pharmacol 93:283–290. [DOI] [PubMed] [Google Scholar]
- Chen R. (2000) Studies of human motor physiology with transcranial magnetic stimulation. Muscle Nerve Suppl 9:S26–S32. [DOI] [PubMed] [Google Scholar]
- Chen R, Classen J, Gerloff C, Celnik P, Wassermann EM, Hallett M, Cohen LG. (1997) Depression of motor cortex excitability by low-frequency transcranial magnetic stimulation. Neurology 48:1398–1403. [DOI] [PubMed] [Google Scholar]
- Cho SS, Strafella AP. (2009) rTMS of the left dorsolateral prefrontal cortex modulates dopamine release in the ipsilateral anterior cingulate cortex and orbitofrontal cortex. PLoS One 4:e6725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coe JW, Brooks PR, Vetelino MG, Wirtz MC, Arnold EP, Huang J, Sands SB, Davis TI, Lebel LA, Fox CB, et al. (2005) Varenicline: an alpha4beta2 nicotinic receptor partial agonist for smoking cessation. J Med Chem 48:3474–3477. [DOI] [PubMed] [Google Scholar]
- Conte A, Attilia ML, Gilio F, Iacovelli E, Frasca V, Bettolo CM, Gabriele M, Giacomelli E, Prencipe M, Berardelli A, et al. (2008) Acute and chronic effects of ethanol on cortical excitability. Clin Neurophysiol 119:667–674. [DOI] [PubMed] [Google Scholar]
- Courtney KE, Schacht JP, Hutchison K, Roche DJ, Ray LA. (2016) Neural substrates of cue reactivity: association with treatment outcomes and relapse. Addict Biol 21:3–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Craig AD, Chen K, Bandy D, Reiman EM. (2000) Thermosensory activation of insular cortex. Nat Neurosci 3:184–190. [DOI] [PubMed] [Google Scholar]
- Critchley HD, Wiens S, Rotshtein P, Ohman A, Dolan RJ. (2004) Neural systems supporting interoceptive awareness. Nat Neurosci 7:189–195. [DOI] [PubMed] [Google Scholar]
- Deng ZD, Lisanby SH, Peterchev AV. (2013) Electric field depth-focality tradeoff in transcranial magnetic stimulation: simulation comparison of 50 coil designs. Brain Stimul 6:1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deng ZD, Lisanby SH, Peterchev AV. (2015) On the stimulation depth of transcranial magnetic stimulation coils. Clin Neurophysiol 126:843–844. [DOI] [PubMed] [Google Scholar]
- Denslow S, Lomarev M, George MS, Bohning DE. (2005) Cortical and subcortical brain effects of transcranial magnetic stimulation (TMS)-induced movement: an interleaved TMS/functional magnetic resonance imaging study. Biol Psychiatry 57:752–760. [DOI] [PubMed] [Google Scholar]
- De Ridder D, Vanneste S, Kovacs S, Sunaert S, Dom G. (2011) Transient alcohol craving suppression by rTMS of dorsal anterior cingulate: an fMRI and LORETA EEG study. Neurosci Lett 496:5–10. [DOI] [PubMed] [Google Scholar]
- Di Lazzaro V, Oliviero A, Meglio M, Cioni B, Tamburrini G, Tonali P, Rothwell JC. (2000a) Direct demonstration of the effect of lorazepam on the excitability of the human motor cortex. Clin Neurophysiol 111:794–799. [DOI] [PubMed] [Google Scholar]
- Di Lazzaro V, Oliviero A, Profice P, Pennisi MA, Di Giovanni S, Zito G, Tonali P, Rothwell JC. (2000b) Muscarinic receptor blockade has differential effects on the excitability of intracortical circuits in the human motor cortex. Exp Brain Res 135:455–461. [DOI] [PubMed] [Google Scholar]
- Di Lazzaro V, Oliviero A, Saturno E, Dileone M, Pilato F, Nardone R, Ranieri F, Musumeci G, Fiorilla T, Tonali P. (2005a) Effects of lorazepam on short latency afferent inhibition and short latency intracortical inhibition in humans. J Physiol 564:661–668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Lazzaro V, Pilato F, Saturno E, Oliviero A, Dileone M, Mazzone P, Insola A, Tonali PA, Ranieri F, Huang YZ, et al. (2005b) Theta-burst repetitive transcranial magnetic stimulation suppresses specific excitatory circuits in the human motor cortex. J Physiol 565:945–950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Lazzaro V, Ziemann U, Lemon RN. (2008) State of the art: physiology of transcranial motor cortex stimulation. Brain Stimul 1:345–362. [DOI] [PubMed] [Google Scholar]
- Dinur-Klein L, Dannon P, Hadar A, Rosenberg O, Roth Y, Kotler M, Zangen A. (2014) Smoking cessation induced by deep repetitive transcranial magnetic stimulation of the prefrontal and insular cortices: a prospective, randomized controlled trial. Biol Psychiatry 76:742–749. [DOI] [PubMed] [Google Scholar]
- Dowdle LT, Brown TR, George MS, Hanlon CA. (2018) Single pulse TMS to the DLPFC, compared to a matched sham control, induces a direct, causal increase in caudate, cingulate, and thalamic BOLD signal. Brain Stimul DOI: 10.1016/j.brs.2018.02.014. [DOI] [PMC free article] [PubMed]
- Dudai Y. (2006) Reconsolidation: the advantage of being refocused. Curr Opin Neurobiol 16:174–178. [DOI] [PubMed] [Google Scholar]
- Dudai Y, Eisenberg M. (2004) Rites of passage of the engram: reconsolidation and the lingering consolidation hypothesis. Neuron 44:93–100. [DOI] [PubMed] [Google Scholar]
- Dutra L, Stathopoulou G, Basden SL, Leyro TM, Powers MB, Otto MW. (2008) A meta-analytic review of psychosocial interventions for substance use disorders. Am J Psychiatry 165:179–187. [DOI] [PubMed] [Google Scholar]
- Eichhammer P, Johann M, Kharraz A, Binder H, Pittrow D, Wodarz N, Hajak G. (2003) High-frequency repetitive transcranial magnetic stimulation decreases cigarette smoking. J Clin Psychiatry 64:951–953. [DOI] [PubMed] [Google Scholar]
- Ersche KD, Barnes A, Jones PS, Morein-Zamir S, Robbins TW, Bullmore ET. (2011) Abnormal structure of frontostriatal brain systems is associated with aspects of impulsivity and compulsivity in cocaine dependence. Brain 134:2013–2024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feil J, Zangen A. (2010) Brain stimulation in the study and treatment of addiction. Neurosci Biobehav Rev 34:559–574. [DOI] [PubMed] [Google Scholar]
- Field M, Cox WM. (2008) Attentional bias in addictive behaviors: a review of its development, causes, and consequences. Drug Alcohol Depend 97:1–20. [DOI] [PubMed] [Google Scholar]
- Fitzgerald PB, Fountain S, Daskalakis ZJ. (2006) A comprehensive review of the effects of rTMS on motor cortical excitability and inhibition. Clin Neurophysiol 117:2584–2596. [DOI] [PubMed] [Google Scholar]
- Franklin TR, Acton PD, Maldjian JA, Gray JD, Croft JR, Dackis CA, O’Brien CP, Childress AR. (2002) Decreased gray matter concentration in the insular, orbitofrontal, cingulate, and temporal cortices of cocaine patients. Biol Psychiatry 51:134–142. [DOI] [PubMed] [Google Scholar]
- Fuhl A, Müller-Dahlhaus F, Lücke C, Toennes SW, Ziemann U. (2015) Low doses of ethanol enhance LTD-like plasticity in human motor cortex. Neuropsychopharmacology 40:2969–2980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galhardoni R, Correia GS, Araujo H, Yeng LT, Fernandes DT, Kaziyama HH, Marcolin MA, Bouhassira D, Teixeira MJ, de Andrade DC. (2015) Repetitive transcranial magnetic stimulation in chronic pain: a review of the literature. Arch Phys Med Rehabil 96 (Suppl):S156–S172. [DOI] [PubMed] [Google Scholar]
- Garavan H, Hester R. (2007) The role of cognitive control in cocaine dependence. Neuropsychol Rev 17:337–345. [DOI] [PubMed] [Google Scholar]
- Garavan H, Pankiewicz J, Bloom A, Cho JK, Sperry L, Ross TJ, Salmeron BJ, Risinger R, Kelley D, Stein EA. (2000) Cue-induced cocaine craving: neuroanatomical specificity for drug users and drug stimuli. Am J Psychiatry 157:1789–1798. [DOI] [PubMed] [Google Scholar]
- Gilbert DL, Ridel KR, Sallee FR, Zhang J, Lipps TD, Wassermann EM. (2006) Comparison of the inhibitory and excitatory effects of ADHD medications methylphenidate and atomoxetine on motor cortex. Neuropsychopharmacology 31:442–449. [DOI] [PubMed] [Google Scholar]
- Gjini K, Ziemann U, Napier TC, Boutros N. (2012) Dysbalance of cortical inhibition and excitation in abstinent cocaine-dependent patients. J Psychiatr Res 46:248–255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldstein RZ, Leskovjan AC, Hoff AL, Hitzemann R, Bashan F, Khalsa SS, Wang GJ, Fowler JS, Volkow ND. (2004) Severity of neuropsychological impairment in cocaine and alcohol addiction: association with metabolism in the prefrontal cortex. Neuropsychologia 42:1447–1458. [DOI] [PubMed] [Google Scholar]
- Goldstein RZ, Tomasi D, Alia-Klein N, Honorio Carrillo J, Maloney T, Woicik PA, Wang R, Telang F, Volkow ND. (2009) Dopaminergic response to drug words in cocaine addiction. J Neurosci 29:6001–6006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldstein RZ, Volkow ND. (2002) Drug addiction and its underlying neurobiological basis: neuroimaging evidence for the involvement of the frontal cortex. Am J Psychiatry 159:1642–1652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gonzales D, Rennard SI, Nides M, Oncken C, Azoulay S, Billing CB, Watsky EJ, Gong J, Williams KE, Reeves KR, Varenicline Phase 3 Study Group (2006) Varenicline, an alpha4beta2 nicotinic acetylcholine receptor partial agonist, vs sustained-release bupropion and placebo for smoking cessation: a randomized controlled trial. JAMA 296:47–55. [DOI] [PubMed] [Google Scholar]
- Gorelick DA, Zangen A, George MS. (2014) Transcranial magnetic stimulation in the treatment of substance addiction. Ann N Y Acad Sci 1327:79–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gratton C, Lee TG, Nomura EM, D’Esposito M. (2013) The effect of theta-burst TMS on cognitive control networks measured with resting state fMRI. Front Syst Neurosci 7:124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grundey J, Freznosa S, Klinker F, Lang N, Paulus W, Nitsche MA. (2013) Cortical excitability in smoking and not smoking individuals with and without nicotine. Psychopharmacology (Berl) 229:653–664. [DOI] [PubMed] [Google Scholar]
- Grundey J, Thirugnanasambandam N, Kaminsky K, Drees A, Skwirba AC, Lang N, Paulus W, Nitsche MA. (2012a) Rapid effect of nicotine intake on neuroplasticity in non-smoking humans. Front Pharmacol 3:186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grundey J, Thirugnanasambandam N, Kaminsky K, Drees A, Skwirba AC, Lang N, Paulus W, Nitsche MA. (2012b) Neuroplasticity in cigarette smokers is altered under withdrawal and partially restituted by nicotine exposition. J Neurosci 32:4156–4162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanlon CA, Beveridge TJ, Porrino LJ. (2013a) Recovering from cocaine: insights from clinical and preclinical investigations. Neurosci Biobehav Rev 37:2037–2046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanlon CA, Canterberry M, Taylor JJ, DeVries W, Li X, Brown TR, George MS. (2013b) Probing the frontostriatal loops involved in executive and limbic processing via interleaved TMS and functional MRI at two prefrontal locations: a pilot study. PLoS One 8:e67917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanlon CA, DeVries W, Dowdle LT, West JA, Siekman B, Li X, George MS. (2015a) A comprehensive study of sensorimotor cortex excitability in chronic cocaine users: integrating TMS and functional MRI data. Drug Alcohol Depend 157:28–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanlon CA, Dowdle LT, Austelle CW, DeVries W, Mithoefer O, Badran BW, George MS. (2015b) What goes up, can come down: novel brain stimulation paradigms may attenuate craving and craving-related neural circuitry in substance dependent individuals. Brain Res 1628:199–209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanlon CA, Dowdle LT, Correia B, Mithoefer O, Kearney-Ramos T, Lench D, Griffin M, Anton RF, George MS. (2017) Left frontal pole theta burst stimulation decreases orbitofrontal and insula activity in cocaine users and alcohol users. Drug Alcohol Depend 178:310–317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanlon CA, Dowdle LT, Moss H, Canterberry M, George MS. (2016) Mobilization of medial and lateral frontal-striatal circuits in cocaine users and controls: an interleaved TMS/BOLD functional connectivity study. Neuropsychopharmacology 41:3032–3041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanlon CA, Dufault DL, Wesley MJ, Porrino LJ. (2011) Elevated gray and white matter densities in cocaine abstainers compared to current users. Psychopharmacology (Berl) 218:681–692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heidegger T, Krakow K, Ziemann U. (2010) Effects of antiepileptic drugs on associative LTP-like plasticity in human motor cortex. Eur J Neurosci 32:1215–1222. [DOI] [PubMed] [Google Scholar]
- Herbsman T, Forster L, Molnar C, Dougherty R, Christie D, Koola J, Ramsey D, Morgan PS, Bohning DE, George MS, et al. (2009) Motor threshold in transcranial magnetic stimulation: the impact of white matter fiber orientation and skull-to-cortex distance. Hum Brain Mapp 30:2044–2055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herremans SC, Baeken C, Vanderbruggen N, Vanderhasselt MA, Zeeuws D, Santermans L, De Raedt R. (2012) No influence of one right-sided prefrontal HF-rTMS session on alcohol craving in recently detoxified alcohol-dependent patients: results of a naturalistic study. Drug Alcohol Depend 120:209–213. [DOI] [PubMed] [Google Scholar]
- Herremans SC, De Raedt R, Van Schuerbeek P, Marinazzo D, Matthys F, De Mey J, Baeken C. (2016) Accelerated HF-rTMS protocol has a rate-dependent effect on dACC activation in alcohol-dependent patients: an open-label feasibility study. Alcohol Clin Exp Res 40:196–205. [DOI] [PubMed] [Google Scholar]
- Herremans SC, Vanderhasselt MA, De Raedt R, Baeken C. (2013) Reduced intra-individual reaction time variability during a Go-NoGo task in detoxified alcohol-dependent patients after one right-sided dorsolateral prefrontal HF-rTMS session. Alcohol Alcohol 48:552–557. [DOI] [PubMed] [Google Scholar]
- Herremans SC, Van Schuerbeek P, De Raedt R, Matthys F, Buyl R, De Mey J, Baeken C. (2015) The impact of accelerated right prefrontal high-frequency repetitive transcranial magnetic stimulation (rTMS) on cue-reactivity: an fMRI study on craving in recently detoxified alcohol-dependent patients. PLoS One 10:e0136182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hester R, Garavan H. (2004) Executive dysfunction in cocaine addiction: evidence for discordant frontal, cingulate, and cerebellar activity. J Neurosci 24:11017–11022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoogendam JM, Ramakers GM, Di Lazzaro V. (2010) Physiology of repetitive transcranial magnetic stimulation of the human brain. Brain Stimul 3:95–118. [DOI] [PubMed] [Google Scholar]
- Höppner J, Broese T, Wendler L, Berger C, Thome J. (2011) Repetitive transcranial magnetic stimulation (rTMS) for treatment of alcohol dependence. World J Biol Psychiatry 12 (Suppl 1):57–62. [DOI] [PubMed] [Google Scholar]
- Huang YZ, Edwards MJ, Rounis E, Bhatia KP, Rothwell JC. (2005) Theta burst stimulation of the human motor cortex. Neuron 45:201–206. [DOI] [PubMed] [Google Scholar]
- Hurt RD, Sachs DP, Glover ED, Offord KP, Johnston JA, Dale LC, Khayrallah MA, Schroeder DR, Glover PN, Sullivan CR, et al. (1997) A comparison of sustained-release bupropion and placebo for smoking cessation. N Engl J Med 337:1195–1202. [DOI] [PubMed] [Google Scholar]
- Ilić TV, Korchounov A, Ziemann U. (2003) Methylphenidate facilitates and disinhibits the motor cortex in intact humans. Neuroreport 14:773–776. [DOI] [PubMed] [Google Scholar]
- Isserles M, Shalev AY, Roth Y, Peri T, Kutz I, Zlotnick E, Zangen A. (2013) Effectiveness of deep transcranial magnetic stimulation combined with a brief exposure procedure in post-traumatic stress disorder--a pilot study. Brain Stimul 6:377–383. [DOI] [PubMed] [Google Scholar]
- SJ Iwabuchi, Raschke F, Auer DP, Liddle PF, Lankappa ST, Palaniyappan L. (2017) Targeted transcranial theta-burst stimulation alters fronto-insular network and prefrontal GABA. Neuroimage 146:395– 403. [DOI] [PubMed] [Google Scholar]
- Janes AC, Frederick Bd, Richardt S, Burbridge C, Merlo-Pich E, Renshaw PF, Evins AE, Fava M, Kaufman MJ. (2009) Brain fMRI reactivity to smoking-related images before and during extended smoking abstinence. Exp Clin Psychopharmacol 17:365–373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janes AC, Pizzagalli DA, Richardt S, deB Frederick B, Chuzi S, Pachas G, Culhane MA, Holmes AJ, Fava M, Evins AE, et al. (2010) Brain reactivity to smoking cues prior to smoking cessation predicts ability to maintain tobacco abstinence [published correction appears in Biol Psychiatry (2010) 67:1002]. Biol Psychiatry 67:722–729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jorenby DE, Hays JT, Rigotti NA, Azoulay S, Watsky EJ, Williams KE, Billing CB, Gong J, Reeves KR, Varenicline Phase 3 Study Group (2006) Efficacy of varenicline, an alpha4beta2 nicotinic acetylcholine receptor partial agonist, vs placebo or sustained-release bupropion for smoking cessation: a randomized controlled trial [published correction appears in JAMA (2006) 296:1355]. JAMA 296:56–63. [DOI] [PubMed] [Google Scholar]
- Josselyn SA, Köhler S, Frankland PW. (2015) Finding the engram. Nat Rev Neurosci 16:521–534. [DOI] [PubMed] [Google Scholar]
- Kaarre O, Kallioniemi E, Könönen M, Tolmunen T, Kekkonen V, Kivimäki P, Heikkinen N, Ferreri F, Laukkanen E, Määttä S. (2018) Heavy alcohol use in adolescence is associated with altered cortical activity: a combined TMS-EEG study. Addict Biol 23:268–280. [DOI] [PubMed] [Google Scholar]
- Kaufman JN, Ross TJ, Stein EA, Garavan H. (2003) Cingulate hypoactivity in cocaine users during a GO-NOGO task as revealed by event-related functional magnetic resonance imaging. J Neurosci 23:7839–7843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Killen JD, Fortmann SP. (1997) Craving is associated with smoking relapse: findings from three prospective studies. Exp Clin Psychopharmacol 5:137–142. [DOI] [PubMed] [Google Scholar]
- Kilts CD, Schweitzer JB, Quinn CK, Gross RE, Faber TL, Muhammad F, Ely TD, Hoffman JM, Drexler KP. (2001) Neural activity related to drug craving in cocaine addiction. Arch Gen Psychiatry 58:334–341. [DOI] [PubMed] [Google Scholar]
- Kimiskidis VK, Papagiannopoulos S, Kazis DA, Sotirakoglou K, Vasiliadis G, Zara F, Kazis A, Mills KR. (2006) Lorazepam-induced effects on silent period and corticomotor excitability. Exp Brain Res 173:603–611. [DOI] [PubMed] [Google Scholar]
- Ko JH, Monchi O, Ptito A, Bloomfield P, Houle S, Strafella AP. (2008) Theta burst stimulation-induced inhibition of dorsolateral prefrontal cortex reveals hemispheric asymmetry in striatal dopamine release during a set-shifting task: a TMS-[(11)C]raclopride PET study. Eur J Neurosci 28:2147–2155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Korchounov A, Ilic TV, Schwinge T, Ziemann U. (2005) Modification of motor cortical excitability by an acetylcholinesterase inhibitor. Exp Brain Res 164:399–405. [DOI] [PubMed] [Google Scholar]
- Krawczyk DC. (2002) Contributions of the prefrontal cortex to the neural basis of human decision making. Neurosci Biobehav Rev 26:631–664. [DOI] [PubMed] [Google Scholar]
- Kringelbach ML, Rolls ET. (2004) The functional neuroanatomy of the human orbitofrontal cortex: evidence from neuroimaging and neuropsychology. Prog Neurobiol 72:341–372. [DOI] [PubMed] [Google Scholar]
- Kübler A, Murphy K, Garavan H. (2005) Cocaine dependence and attention switching within and between verbal and visuospatial working memory. Eur J Neurosci 21:1984–1992. [DOI] [PubMed] [Google Scholar]
- Kujirai K, Kujirai T, Sinkjaer T, Rothwell JC. (2006) Associative plasticity in human motor cortex during voluntary muscle contraction. J Neurophysiol 96:1337–1346. [DOI] [PubMed] [Google Scholar]
- Lang N, Hasan A, Sueske E, Paulus W, Nitsche MA. (2008) Cortical hypoexcitability in chronic smokers? A transcranial magnetic stimulation study. Neuropsychopharmacology 33:2517–2523. [DOI] [PubMed] [Google Scholar]
- Lee WH, Lisanby SH, Laine AF, Peterchev AV. (2016) Comparison of electric field strength and spatial distribution of electroconvulsive therapy and magnetic seizure therapy in a realistic human head model. Eur Psychiatry 36:55–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li X, Hartwell KJ, Owens M, Lematty T, Borckardt JJ, Hanlon CA, Brady KT, George MS. (2013a) Repetitive transcranial magnetic stimulation of the dorsolateral prefrontal cortex reduces nicotine cue craving. Biol Psychiatry 73:714–720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li X, Malcolm RJ, Huebner K, Hanlon CA, Taylor JJ, Brady KT, George MS, See RE. (2013b) Low frequency repetitive transcranial magnetic stimulation of the left dorsolateral prefrontal cortex transiently increases cue-induced craving for methamphetamine: a preliminary study. Drug Alcohol Depend 133:641–646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li X, Sahlem GL, Badran BW, McTeague LM, Hanlon CA, Hartwell KJ, Henderson S, George MS. (2017) Transcranial magnetic stimulation of the dorsal lateral prefrontal cortex inhibits medial orbitofrontal activity in smokers. Am J Addict 26:788–794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liebetanz D, Nitsche MA, Tergau F, Paulus W. (2002) Pharmacological approach to the mechanisms of transcranial DC-stimulation-induced after-effects of human motor cortex excitability. Brain 125:2238–2247. [DOI] [PubMed] [Google Scholar]
- Loheswaran G, Barr MS, Rajji TK, Blumberger DM, Le Foll B, Daskalakis ZJ. (2016) Alcohol intoxication by binge drinking impairs neuroplasticity. Brain Stimul 9:27–32. [DOI] [PubMed] [Google Scholar]
- Lücke C, Heidegger T, Röhner M, Toennes SW, Krivanekova L, Müller-Dahlhaus F, Ziemann U. (2014) Deleterious effects of a low amount of ethanol on LTP-like plasticity in human cortex. Neuropsychopharmacology 39:1508–1518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lusted A, Roerecke M, Goldner E, Rehm J, Fischer B. (2013) Prevalence of pain among nonmedical prescription opioid users in substance use treatment populations: systematic review and meta-analyses. Pain Physician 16:E671–E684. [PubMed] [Google Scholar]
- Maas LC, Lukas SE, Kaufman MJ, Weiss RD, Daniels SL, Rogers VW, Kukes TJ, Renshaw PF. (1998) Functional magnetic resonance imaging of human brain activation during cue-induced cocaine craving. Am J Psychiatry 155:124–126. [DOI] [PubMed] [Google Scholar]
- Malenka RC, Bear MF. (2004) LTP and LTD: an embarrassment of riches. Neuron 44:5–21. [DOI] [PubMed] [Google Scholar]
- Marhe R, Luijten M, van de Wetering BJ, Smits M, Franken IH. (2013) Individual differences in anterior cingulate activation associated with attentional bias predict cocaine use after treatment. Neuropsychopharmacology 38:1085–1093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matochik JA, London ED, Eldreth DA, Cadet JL, Bolla KI. (2003) Frontal cortical tissue composition in abstinent cocaine abusers: a magnetic resonance imaging study. Neuroimage 19:1095–1102. [DOI] [PubMed] [Google Scholar]
- Mattick RP, Kimber J, Breen C, Davoli M. (2004) Buprenorphine maintenance versus placebo or methadone maintenance for opioid dependence. Cochrane Database Syst Rev (3):CD002207. [DOI] [PubMed] [Google Scholar]
- Maxwell J. (1861) On physical lines of force. Philos Mag 1:451–513. [Google Scholar]
- McDonnell MN, Orekhov Y, Ziemann U. (2007) Suppression of LTP-like plasticity in human motor cortex by the GABAB receptor agonist baclofen. Exp Brain Res 180:181–186. [DOI] [PubMed] [Google Scholar]
- McHugh MJ, Demers CH, Braud J, Briggs R, Adinoff B, Stein EA. (2013) Striatal-insula circuits in cocaine addiction: implications for impulsivity and relapse risk. Am J Drug Alcohol Abuse 39:424–432. [DOI] [PubMed] [Google Scholar]
- Michael N, Gosling M, Reutemann M, Kersting A, Heindel W, Arolt V, Pfleiderer B. (2003) Metabolic changes after repetitive transcranial magnetic stimulation (rTMS) of the left prefrontal cortex: a sham-controlled proton magnetic resonance spectroscopy (1H MRS) study of healthy brain. Eur J Neurosci 17:2462–2468. [DOI] [PubMed] [Google Scholar]
- Miller EK, Cohen JD. (2001) An integrative theory of prefrontal cortex function. Annu Rev Neurosci 24:167–202. [DOI] [PubMed] [Google Scholar]
- Minozzi S, Amato L, Vecchi S, Davoli M, Kirchmayer U, Verster A. (2011) Oral naltrexone maintenance treatment for opioid dependence. Cochrane Database Syst Rev (4):CD001333. [DOI] [PubMed] [Google Scholar]
- Mishra BR, Nizamie SH, Das B, Praharaj SK. (2010) Efficacy of repetitive transcranial magnetic stimulation in alcohol dependence: a sham-controlled study. Addiction 105:49–55. [DOI] [PubMed] [Google Scholar]
- Moeller FG, Dougherty DM, Barratt ES, Schmitz JM, Swann AC, Grabowski J. (2001) The impact of impulsivity on cocaine use and retention in treatment. J Subst Abuse Treat 21:193–198. [DOI] [PubMed] [Google Scholar]
- Moeller FG, Steinberg JL, Schmitz JM, Ma L, Liu S, Kjome KL, Rathnayaka N, Kramer LA, Narayana PA. (2010) Working memory fMRI activation in cocaine-dependent subjects: association with treatment response. Psychiatry Res 181:174–182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moll GH, Heinrich H, Rothenberger A. (2003) Methylphenidate and intracortical excitability: opposite effects in healthy subjects and attention-deficit hyperactivity disorder. Acta Psychiatr Scand 107:69–72. [DOI] [PubMed] [Google Scholar]
- Moran-Santa Maria MM, Hartwell KJ, Hanlon CA, Canterberry M, Lematty T, Owens M, Brady KT, George MS. (2015) Right anterior insula connectivity is important for cue-induced craving in nicotine-dependent smokers. Addict Biol 20:407–414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nahas Z, Lomarev M, Roberts DR, Shastri A, Lorberbaum JP, Teneback C, McConnell K, Vincent DJ, Li X, George MS, et al. (2001) Unilateral left prefrontal transcranial magnetic stimulation (TMS) produces intensity-dependent bilateral effects as measured by interleaved BOLD fMRI. Biol Psychiatry 50:712–720. [DOI] [PubMed] [Google Scholar]
- Naim-Feil J, Bradshaw JL, Rogasch NC, Daskalakis ZJ, Sheppard DM, Lubman DI, Fitzgerald PB. (2016) Cortical inhibition within motor and frontal regions in alcohol dependence post-detoxification: a pilot TMS-EEG study. World J Biol Psychiatry 17:547–556. [DOI] [PubMed] [Google Scholar]
- Naqvi NH, Rudrauf D, Damasio H, Bechara A. (2007) Damage to the insula disrupts addiction to cigarette smoking. Science 315:531–534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nardone R, Bergmann J, De Blasi P, Kronbichler M, Kraus J, Caleri F, Tezzon F, Ladurner G, Golaszewski S. (2010a) Cholinergic dysfunction and amnesia in patients with Wernicke-Korsakoff syndrome: a transcranial magnetic stimulation study. J Neural Transm (Vienna) 117:385–391. [DOI] [PubMed] [Google Scholar]
- Nardone R, Bergmann J, Kronbichler M, Caleri F, Lochner P, Tezzon F, Ladurner G, Golaszewski S. (2010b) Altered motor cortex excitability to magnetic stimulation in alcohol withdrawal syndrome. Alcohol Clin Exp Res 34:628–632. [DOI] [PubMed] [Google Scholar]
- Nitsche MA, Fricke K, Henschke U, Schlitterlau A, Liebetanz D, Lang N, Henning S, Tergau F, Paulus W. (2003) Pharmacological modulation of cortical excitability shifts induced by transcranial direct current stimulation in humans. J Physiol 553:293–301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nitsche MA, Grundey J, Liebetanz D, Lang N, Tergau F, Paulus W. (2004a) Catecholaminergic consolidation of motor cortical neuroplasticity in humans. Cereb Cortex 14:1240–1245. [DOI] [PubMed] [Google Scholar]
- Nitsche MA, Jaussi W, Liebetanz D, Lang N, Tergau F, Paulus W. (2004b) Consolidation of human motor cortical neuroplasticity by D-cycloserine. Neuropsychopharmacology 29:1573–1578. [DOI] [PubMed] [Google Scholar]
- Nitsche MA, Paulus W. (2000) Excitability changes induced in the human motor cortex by weak transcranial direct current stimulation. J Physiol 527:633–639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Connell KA, Shiffman S, Decarlo LT. (2011) Does extinction of responses to cigarette cues occur during smoking cessation? Addiction 106:410–417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orth M, Amann B, Robertson MM, Rothwell JC. (2005) Excitability of motor cortex inhibitory circuits in Tourette syndrome before and after single dose nicotine. Brain 128:1292–1300. [DOI] [PubMed] [Google Scholar]
- Pascual-Leone A, Valls-Solé J, Wassermann EM, Hallett M. (1994) Responses to rapid-rate transcranial magnetic stimulation of the human motor cortex. Brain 117:847–858. [DOI] [PubMed] [Google Scholar]
- Passik SD, Hays L, Eisner N, Kirsh KL. (2006) Psychiatric and pain characteristics of prescription drug abusers entering drug rehabilitation. J Pain Palliat Care Pharmacother 20:5–13. [PubMed] [Google Scholar]
- Pierantozzi M, Marciani MG, Palmieri MG, Brusa L, Galati S, Caramia MD, Bernardi G, Stanzione P. (2004) Effect of Vigabatrin on motor responses to transcranial magnetic stimulation: an effective tool to investigate in vivo GABAergic cortical inhibition in humans. Brain Res 1028:1–8. [DOI] [PubMed] [Google Scholar]
- Politi E, Fauci E, Santoro A, Smeraldi E. (2008) Daily sessions of transcranial magnetic stimulation to the left prefrontal cortex gradually reduce cocaine craving. Am J Addict 17:345–346. [DOI] [PubMed] [Google Scholar]
- Priori A, Berardelli A, Inghilleri M, Accornero N, Manfredi M. (1994) Motor cortical inhibition and the dopaminergic system: pharmacological changes in the silent period after transcranial brain stimulation in normal subjects, patients with Parkinson’s disease and drug-induced parkinsonism. Brain 117:317–323. [DOI] [PubMed] [Google Scholar]
- Pripfl J, Tomova L, Riecansky I, Lamm C. (2014) Transcranial magnetic stimulation of the left dorsolateral prefrontal cortex decreases cue-induced nicotine craving and EEG delta power. Brain Stimul 7:226–233. [DOI] [PubMed] [Google Scholar]
- Rapinesi C, Del Casale A, Di Pietro S, Ferri VR, Piacentino D, Sani G, Raccah RN, Zangen A, Ferracuti S, Vento AE, et al. (2016) Add-on high frequency deep transcranial magnetic stimulation (dTMS) to bilateral prefrontal cortex reduces cocaine craving in patients with cocaine use disorder. Neurosci Lett 629:43–47. [DOI] [PubMed] [Google Scholar]
- Reis J, Swayne OB, Vandermeeren Y, Camus M, Dimyan MA, Harris-Love M, Perez MA, Ragert P, Rothwell JC, Cohen LG. (2008) Contribution of transcranial magnetic stimulation to the understanding of cortical mechanisms involved in motor control. J Physiol 586:325–351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Risinger RC, Salmeron BJ, Ross TJ, Amen SL, Sanfilipo M, Hoffmann RG, Bloom AS, Garavan H, Stein EA. (2005) Neural correlates of high and craving during cocaine self-administration using BOLD fMRI. Neuroimage 26:1097–1108. [DOI] [PubMed] [Google Scholar]
- Rose JE, McClernon FJ, Froeliger B, Behm FM, Preud’homme X, Krystal AD. (2011) Repetitive transcranial magnetic stimulation of the superior frontal gyrus modulates craving for cigarettes. Biol Psychiatry 70:794–799. [DOI] [PubMed] [Google Scholar]
- Sachdeva A, Choudhary M, Chandra M. (2015) Alcohol withdrawal syndrome: benzodiazepines and beyond. J Clin Diagn Res 9:VE01–VE07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schacht JP, Anton RF, Myrick H. (2013) Functional neuroimaging studies of alcohol cue reactivity: a quantitative meta-analysis and systematic review. Addict Biol 18:121–133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheffer CE, Bickel WK, Brandon TH, Franck CT, Deen D, Panissidi L, Abdali SA, Pittman JC, Lunden SE, Prashad N, et al. (2018) Preventing relapse to smoking with transcranial magnetic stimulation: feasibility and potential efficacy. Drug Alcohol Depend 182:8–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen Y, Cao X, Tan T, Shan C, Wang Y, Pan J, He H, Yuan TF. (2016) 10-Hz repetitive transcranial magnetic stimulation of the left dorsolateral prefrontal cortex reduces heroin cue craving in long-term addicts. Biol Psychiatry 80:e13–e14. [DOI] [PubMed] [Google Scholar]
- Singer T, Critchley HD, Preuschoff K. (2009) A common role of insula in feelings, empathy and uncertainty. Trends Cogn Sci 13:334–340. [DOI] [PubMed] [Google Scholar]
- Stagg CJ, Best JG, Stephenson MC, O'Shea J, Wylezinska M, Kincses ZT, Morris PG, Matthews PM, Johansen-Berg H. (2009) Polarity-sensitive modulation of cortical neurotransmitters by transcranial stimulation. J Neurosci 29:5202–5206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stalnaker TA, Takahashi Y, Roesch MR, Schoenbaum G. (2009) Neural substrates of cognitive inflexibility after chronic cocaine exposure. Neuropharmacology 56 (Suppl 1):63–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strafella AP, Paus T, Barrett J, Dagher A. (2001) Repetitive transcranial magnetic stimulation of the human prefrontal cortex induces dopamine release in the caudate nucleus. J Neurosci 21:RC157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strafella AP, Paus T, Fraraccio M, Dagher A. (2003) Striatal dopamine release induced by repetitive transcranial magnetic stimulation of the human motor cortex. Brain 126:2609–2615. [DOI] [PubMed] [Google Scholar]
- Strube W, Bunse T, Nitsche MA, Wobrock T, Aborowa R, Misewitsch K, Herrmann M, Falkai P, Hasan A. (2015) Smoking restores impaired LTD-like plasticity in schizophrenia: a transcranial direct current stimulation study. Neuropsychopharmacology 40:822–830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sundaresan K, Ziemann U, Stanley J, Boutros N. (2007) Cortical inhibition and excitation in abstinent cocaine-dependent patients: a transcranial magnetic stimulation study. Neuroreport 18:289–292. [DOI] [PubMed] [Google Scholar]
- Taylor JJ, Borckardt JJ, Canterberry M, Li X, Hanlon CA, Brown TR, George MS. (2013) Naloxone-reversible modulation of pain circuitry by left prefrontal rTMS. Neuropsychopharmacology 38:1189–1197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teo JT, Swayne OB, Rothwell JC. (2007) Further evidence for NMDA-dependence of the after-effects of human theta burst stimulation. Clin Neurophysiol 118:1649–1651. [DOI] [PubMed] [Google Scholar]
- Terraneo A, Leggio L, Saladini M, Ermani M, Bonci A, Gallimberti L. (2016) Transcranial magnetic stimulation of dorsolateral prefrontal cortex reduces cocaine use: a pilot study. Eur Neuropsychopharmacol 26:37–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thickbroom GW. (2007) Transcranial magnetic stimulation and synaptic plasticity: experimental framework and human models. Exp Brain Res 180:583–593. [DOI] [PubMed] [Google Scholar]
- Thirugnanasambandam N, Grundey J, Adam K, Drees A, Skwirba AC, Lang N, Paulus W, Nitsche MA. (2011) Nicotinergic impact on focal and non-focal neuroplasticity induced by non-invasive brain stimulation in non-smoking humans. Neuropsychopharmacology 36:879–886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trojak B, Meille V, Achab S, Lalanne L, Poquet H, Ponavoy E, Blaise E, Bonin B, Chauvet-Gelinier JC. (2015) Transcranial magnetic stimulation combined with nicotine replacement therapy for smoking cessation: a randomized controlled trial. Brain Stimul 8:1168–1174. [DOI] [PubMed] [Google Scholar]
- Vicario CM, Komeilipoor N, Cesari P, Rafal RD, Nitsche MA. (2014) Enhanced corticobulbar excitability in chronic smokers during visual exposure to cigarette smoking cues. J Psychiatry Neurosci 39:232–238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vidal-Pineiro D, Martin-Trias P, Falcon C, Bargallo N, Clemente IC, Valls-Sole J, Junque C, Pascual-Leone A, Bartres-Faz D. (2015) Neurochemical modulation in posteromedial default-mode network cortex induced by transcranial magnetic stimulation. Brain Stimul 8:937–944. [DOI] [PubMed] [Google Scholar]
- Volkow ND, Fowler JS. (2000) Addiction, a disease of compulsion and drive: involvement of the orbitofrontal cortex. Cereb Cortex 10:318–325. [DOI] [PubMed] [Google Scholar]
- Volkow ND, Fowler JS, Wang GJ, Hitzemann R, Logan J, Schlyer DJ, Dewey SL, Wolf AP. (1993) Decreased dopamine D2 receptor availability is associated with reduced frontal metabolism in cocaine abusers. Synapse 14:169–177. [DOI] [PubMed] [Google Scholar]
- Volkow ND, Fowler JS, Wolf AP, Hitzemann R, Dewey S, Bendriem B, Alpert R, Hoff A. (1991) Changes in brain glucose metabolism in cocaine dependence and withdrawal. Am J Psychiatry 148:621–626. [DOI] [PubMed] [Google Scholar]
- Volkow ND, Hitzemann R, Wang GJ, Fowler JS, Wolf AP, Dewey SL, Handlesman L. (1992) Long-term frontal brain metabolic changes in cocaine abusers. Synapse 11:184–190. [DOI] [PubMed] [Google Scholar]
- Volkow ND, Wang GJ, Ma Y, Fowler JS, Wong C, Ding YS, Hitzemann R, Swanson JM, Kalivas P. (2005) Activation of orbital and medial prefrontal cortex by methylphenidate in cocaine-addicted subjects but not in controls: relevance to addiction. J Neurosci 25:3932–3939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vollstädt-Klein S, Loeber S, Richter A, Kirsch M, Bach P, von der Goltz C, Hermann D, Mann K, Kiefer F. (2012) Validating incentive salience with functional magnetic resonance imaging: association between mesolimbic cue reactivity and attentional bias in alcohol-dependent patients. Addict Biol 17:807–816. [DOI] [PubMed] [Google Scholar]
- Ward J, Bell J, Mattick RP, Hall W. (1996) Methadone maintenance therapy for opioid dependence: a guide to appropriate use. CNS Drugs 6:440–449. [Google Scholar]
- Wei XF, Grill WM. (2005) Current density distributions, field distributions and impedance analysis of segmented deep brain stimulation electrodes. J Neural Eng 2:139–147. [DOI] [PubMed] [Google Scholar]
- Werhahn KJ, Kunesch E, Noachtar S, Benecke R, Classen J. (1999) Differential effects on motorcortical inhibition induced by blockade of GABA uptake in humans. J Physiol 517:591–597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolters A, Sandbrink F, Schlottmann A, Kunesch E, Stefan K, Cohen LG, Benecke R, Classen J. (2003) A temporally asymmetric Hebbian rule governing plasticity in the human motor cortex. J Neurophysiol 89:2339–2345. [DOI] [PubMed] [Google Scholar]
- Wing VC, Barr MS, Wass CE, Lipsman N, Lozano AM, Daskalakis ZJ, George TP. (2013) Brain stimulation methods to treat tobacco addiction. Brain Stimul 6:221–230. [DOI] [PubMed] [Google Scholar]
- World Health Organization (2004) Global Status Report on Alcohol 2004, Department of Mental Health and Substance Abuse, Geneva, Switzerland. [Google Scholar]
- World Health Organization (2012) Global Report: Mortality Attributable to Tobacco, World Health Organization, Geneva, Switzerland. [Google Scholar]
- Zanardi R, Serretti A, Rossini D, Franchini L, Cusin C, Lattuada E, Dotoli D, Smeraldi E. (2001) Factors affecting fluvoxamine antidepressant activity: influence of pindolol and 5-HTTLPR in delusional and nondelusional depression. Biol Psychiatry 50:323–330. [DOI] [PubMed] [Google Scholar]
- Ziemann U. (2004) TMS and drugs. Clin Neurophysiol 115:1717–1729. [DOI] [PubMed] [Google Scholar]
- Ziemann U. (2011) Transcranial magnetic stimulation at the interface with other techniques: a powerful tool for studying the human cortex. Neuroscientist 17:368–381. [DOI] [PubMed] [Google Scholar]
- Ziemann U, Lönnecker S, Paulus W. (1995) Inhibition of human motor cortex by ethanol: a transcranial magnetic stimulation study. Brain 118:1437–1446. [DOI] [PubMed] [Google Scholar]
- Ziemann U, Netz J, Szelényi A, Hömberg V. (1993) Spinal and supraspinal mechanisms contribute to the silent period in the contracting soleus muscle after transcranial magnetic stimulation of human motor cortex. Neurosci Lett 156:167–171. [DOI] [PubMed] [Google Scholar]
- Ziemann U, Reis J, Schwenkreis P, Rosanova M, Strafella A, Badawy R, Müller-Dahlhaus F. (2015) TMS and drugs revisited 2014. Clin Neurophysiol 126:1847–1868. [DOI] [PubMed] [Google Scholar]
- Ziemann U, Rothwell JC. (2000) I-waves in motor cortex. J Clin Neurophysiol 17:397–405. [DOI] [PubMed] [Google Scholar]
- Ziemann U, Rothwell JC, Ridding MC. (1996) Interaction between intracortical inhibition and facilitation in human motor cortex. J Physiol 496:873–881. [DOI] [PMC free article] [PubMed] [Google Scholar]