

Abstract
Background
The relatively low efficiency of biolistic transformation and subsequent integration of multiple copies of the introduced gene/s significantly complicate the genetic modification of wheat (Triticum aestivum) and other plant species. One of the key factors contributing to the reproducibility of this method is the uniformity of the DNA/gold suspension, which is dependent on the coating procedure employed. It was also shown recently that the relative frequency of single copy transgene inserts could be increased through the use of nanogram quantities of the DNA during coating.
Results
A simplified DNA/gold coating method was developed to produce fertile transgenic plants, via microprojectile bombardment of callus cultures induced from immature embryos. In this method, polyethyleneglycol (PEG) and magnesium salt solutions were utilized in place of the spermidine and calcium chloride of the standard coating method, to precipitate the DNA onto gold microparticles. The prepared microparticles were used to generate transgenics from callus cultures of commercial bread wheat cv. Gladius resulting in an average transformation frequency of 9.9%. To increase the occurrence of low transgene copy number events, nanogram amounts of the minimal expression cassettes containing the gene of interest and the hpt gene were used for co-transformation. A total of 1538 transgenic wheat events were generated from 15,496 embryos across 19 independent experiments. The variation of single copy insert frequencies ranged from 16.1 to 73.5% in the transgenic wheat plants, which compares favourably to published results.
Conclusions
The DNA/gold coating procedure presented here allows efficient, large scale transformation of wheat. The use of nanogram amounts of vector DNA improves the frequency of single copy transgene inserts in transgenic wheat plants.
Keywords: Biolistic transformation, DNA/gold coating, Immature embryo, Single copy events, Transformation frequency, Triticum aestivum
Background
Plant genetic transformation using biolistic microprojectile bombardment is broadly applied in the generation of transgenic plants, including important cereal crops, such as rice (Oryza sativa), maize (Zea mays) and wheat (Triticum aestivum) [1–3]. As one of the main industrial crops, wheat has attracted considerable attention in the establishment and optimization of efficient transformation methods [3–15]. In most of these publications, the DNA coating preparation remained largely unchanged from the original protocol based on the formation of an unstable DNA/spermidine/Ca2+ complex [16]. Despite some optimization [17, 18], the reproducibility of this method remains quite variable in large-scale stable transformation experiments.
Another major concern of stable biolistic transformation is random, multi-copy transgene integration, commonly resulting in irregular compositions of inverted repeats or transgene rearrangements. These can lead to transgene silencing [19, 20], aberrant transgene expression in subsequent generations [21, 22] and even loss of the transgenes [23]. Transgenic plants with single/low copy inserts are less affected by these problems [24], and thus are desirable for functional genomic studies or the production of marker free plants after genetic segregation. Using minimal transgene cassettes instead of bombarding with whole plasmid DNA can increase the transformation frequency (TF) with simple transgene integration patterns [13, 25, 26]. To date, this strategy has been applied in the transformation of several monocotyledonous species such as wheat [13], maize [26], pearl millet (Pennisetum glaucum) [27], and sugarcane (Saccharum officinarum) [28]. Furthermore, low transgene copy number is found to be inversely correlated with the amount of the DNA cassette used in the biolistic transformation [26]. Quantities as low as 2.50–2.73 ng of DNA cassette per shot have been recommended for the efficient production of single copy events in maize [26] and in sugarcane [29].
In this paper, we present a biolistic transformation method that uses a reduced load of DNA expression cassettes in combination with the microprojectile particles coating procedure based on PEG/MgCl2. This process results in relatively high transformation frequency and greater representation of single transgene copy events, averaging 38.2% as deduced from the examination of 1538 transformed wheat plants in 19 independent experiments.
Methods
Plant material and explants
The spring wheat cv. Gladius was utilised throughout this research. Gladius is an elite Australian cultivar with the pedigree RAC875/Krichauff//Excalibur/Kukri/3/RAC875/Krichauff/4/RAC875//Excalibur/Kukri. Donor plants were maintained in a glasshouse (23 °C, 16/8 h day/night). Immature grains were collected 11–14 days after flowering and surface-sterilized in 70% (v/v) ethanol for 2 min, then for 20 min in 1% NaOCl (sodium hypochlorite), followed by three rinses in sterile water. The excised embryos were incubated, scutellum side up, on solidified callus induction MS [30] medium containing 500 mg/L casein hydrolysate, 30 mg/L CP (centrophenoxine), 30% sucrose, and 5 g/L gelrite (MS30-CP) for 7–14 days before transformation [15]. T. monococcum L. suspension cells [31] were propagated in 250 ml flasks with weekly subcultures in liquid basal MS2–2,4D medium with 2 mg/L 2,4D and 30 g/L sucrose.
DNA constructs used in transformation
The GUS (β-glucuronidase A) gene driven by the P-Ubi promoter (1986 bp) [32] was used for transient expression analysis (Fig. 1). For stable transformation, the ~ 3.0 Kb selectable marker DNA cassette, containing the P-Ubi promoter, hpt, hygromycin resistance gene, and cauliflower mosaic virus 35S Terminator, was cut out with PmeI and XmnI restriction enzymes (New England Biolabs) (Fig. 1). Plasmids carrying the Genes of Interest (GOI), represented by 19 new wheat genes isolated at the University of Adelaide, were digested with restriction enzymes to release the DNA cassettes, and isolated by electrophoresis on 1% agarose gels. The gel pieces bearing the minimal DNA cassettes were processed with the GenElute Gel Extraction Kit (Sigma Aldrich).
Fig. 1.
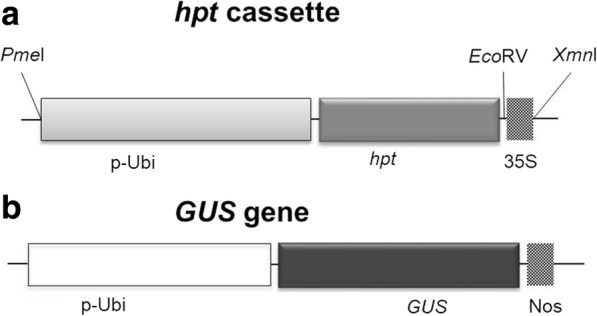
Schematic representation of the hpt and GUS gene constructs used in the transformation experiments. P-Ubi, ubiquitin promoter (1.6 Kb); hpt, hygromycin resistance gene (1 Kb); GUS, gusA gene (1.8 Kb); 35S, 35S terminator (200 bp); Nos, Nos terminator (258 bp). About ~ 3 Kb hpt DNA cassette was used for stable transformation
DNA/gold coating procedure
The DNA/gold coating procedure corresponding to the initial Bio-Rad protocol [16] was carried out as follows: 50 μl aliquots of gold microparticles (0.6 μm, 30 mg/ml) suspended in 50% (v/v) sterile glycerol solution were combined with DNA (5 μg in 5 μl of sterile water), 2.5 M CaCl2 (50 μl) and 0.1 M spermidine, Spd (20 μl) under constant vortexing. After vortexing for several minutes, the suspension was incubated at room temperature for 20 min. The coated DNA was then pelleted by 1 min centrifugation at 3920×g. The pelleted DNA was washed with 100 μl of 70% (v/v) ethanol, followed by 100 μl of absolute ethanol and finally resuspended in 60 μl of absolute ethanol. Six microliter aliquots of the coated DNA were used per bombardment.
The PEG/Mg2+ (PM) coating procedure was executed as follows: 50 μl aliquots of gold microparticles were combined with DNA (10 μl) and supplemented under vortexing with 10 μl of PM solution (42% PEG 2000 and 560 mM MgCl2). The stock solutions of 52.5% PEG 2000 (Sigma-Aldrich) and 2.8 M MgCl2 (filter sterilized) were mixed at a ratio of 4:1. The suspension was vortexed for 1 min, followed by 20 min incubation at room temperature and a 1–5 min centrifugation (longer for diluted DNA) to pellet the coated DNA. The pelleted DNA was washed once with 100 μl of absolute ethanol and resuspended in 60 μl of absolute ethanol (6 μl per bombardment). All steps were carried out at room temperature. In stable transformation experiments, 60 ng of selective marker gene (Fig. 1a) and 60 ng of GOI DNA cassettes were used for DNA/gold coating (50 μl gold + 10 μl of DNA + 10 μl PM solution).
Plant transformation
Particle bombardment was conducted using the Particle Delivery System PDS-1000/He (Bio-Rad). Embryogenic callus was osmotically pre-treated on MS5-CP medium [15] containing 100 g/L sucrose for 4 h. Callus pieces (ca. 1–2 mm) placed in the centre of the plate to form a circle with a diameter of 25–30 mm were bombarded at 900 psi, with the 15 mm flight distance and a 60 mm target distance. The bombarded calli were transferred to MS5-CP medium 16 h after the treatment and were grown in the dark for 1 week. Plantlet selection and regeneration procedures were as described earlier [15].
Transient GUS assay
The uncut plasmid DNA pAHC25 [32] was used only for transient expression analysis in suspension cells which were treated 4 days after subculture. Five ml of well-mixed suspension adjusted to OD = 0.1–0.2 was spread on Ruled Qualitative Filter Paper discs with a 5 mm Ruled Grid, diameter 55 mm (Whatman, USA). A vacuum was applied to get an even spread of a very thin layer of cells on the surface of the filter paper. The coated papers were subsequently transferred to MS2–2,4D medium with 100 g/L sucrose for osmotic pre-treatment for 4 h. Bombardment conditions for the callus tissues and the suspension cells were the same as described above. The transformed cells were maintained on the MS2 medium for 48 h. GUS staining with X-gluc (0.5 mg/mL) was as described in [33]. Filter disks with the treated cells were carefully transferred into 60 mm plastic Petri dishes with 550 μl of the X-gluc solution and incubated overnight at 37 °C. For further details see [34].
To quantify GUS expression in the T. monococcum suspension cells, the number of cells with blue staining was counted on the same Ruled Qualitative Filter Papers under a Binocular microscope (Nikon, Japan), and recorded as described earlier [34, 35]. Each treatment was repeated at least three times, with mean and variability calculated.
DNA isolation, qPCR analysis and Southern hybridisation
DNA was extracted from leaf tissue using the freeze-dry method for qPCR analysis [36] and using the phenol-chloroform method for the Southern hybridisation [37]. Total DNA was quantified using a Nano-Drop spectrophotometer (ND-2000, NanoDrop Technologies Inc., USA). Transgene copy numbers were determined by Real-time quantitative PCR using two reference genes, Cyclophilin and Glyceraldehyde-3-phosphate dehydrogenase, as described in [38], and confirmed with a cross-test of the same individuals by Southern autoradiography hybridisation. For the Southern analysis, DNA samples were digested with EcoRV at 37 °C for 5 h, run on a 1.0% (v/v) agarose gel, blotted onto a Hybond N+ nylon membrane (GE Healthcare Life Sciences, NSW, Australia) and subjected to hybridisation with a 32P-labelled DNA probe specific to the transgene following standard methods. The DNA probe was labelled using the random primer method with the Ready-to-Go DNA labelling kit beads (GE Healthcare Life Sciences, NSW, Australia). An ‘in-house’ probe (35S, 354 bp) targeting part of the backbone of the 4237 bp transformed DNA fragment was used. Primers for amplifying this probe by PCR were as follows: F: 5’-CAACATGGTGGAGCACGAC-3′ and R: 5-GCGTCATCCCTTACGTCAGTGGAG-3′.
Statistical analysis
Mean and standard errors were calculated with ANOVA using standard Excel software. Probabilities for significance were calculated using Student’s t-test.
Results
PEG/Mg2+ coating procedure
PEG and Mg2+ solutions are known to allow efficient DNA precipitation at room temperature [39, 40]. We found that they can be applied successfully in DNA/gold coating (Fig. 2). In transient transformation experiments with 6% PEG / 80 mM MgCl2, the relative numbers of GUS-positive cells were, on average, between 1.2-fold [41] and 1.6-fold (Fig. 3a) higher compared to the Spd/CaCl2 coating method. It was also found that the PEG/Mg2+ DNA/gold coating mixture remains relatively stable for 3 h at room temperature providing for additional flexibility in long experiments and implying that this method offers better DNA protection. The GUS transient expression, based on relative numbers of GUS-positive cells, was lower by 3.9-fold after 3 h of incubation at room temperature for the original Spd/Ca2+ coating mixture (Fig. 3a). Magnesium chloride (MgCl2, pH = 5.5) and magnesium acetate (MgAc, pH = 6.5) at 80 mM concentration had a comparable effect on the successful DNA/gold co-precipitation although there was an average of 1.14-fold improvement with MgCl2 compared to MgAc (Fig. 3b). Therefore, in the stable transformation experiments, PEG-MgCl2 was used for coating as the optimal method.
Fig. 2.
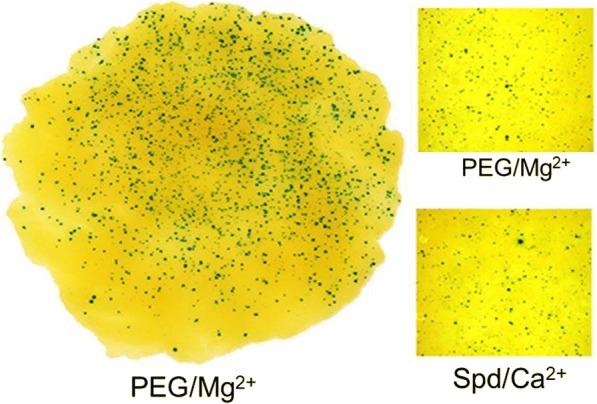
The bombarded T. monococcum suspension cells. The treated cultures exhibit relatively uniform distribution of the X-Gluc stained cells over an area of 30–40 mm in diameter
Fig. 3.
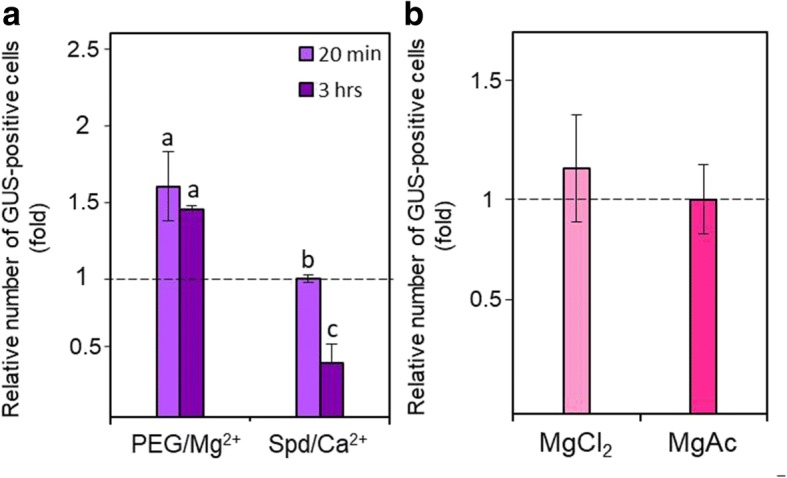
Relative number of GUS-positive cells in wheat cell suspension cultures bombarded under different DNA/gold coating conditions. a Incubation time (20 min and 3 h, light and dark purple, respectively) for different coating methods. b Sources of ions: 80 mM MgCl2 or MgAc. P-Ubi:GUS DNA cassette (300 ng per shot) was used for each treatment. The Spd/Ca2+ method with 20 min incubation was set as one unit. Bars represent means ± standard errors for three replicates. Different letters above the bars represent significant differences (P < 0.05) using Student’s t-test
Efficient regeneration of single copy T0 transgenic wheat plants
To obtain high numbers of single copy events, a ratio of 1:1 of 6 ng/per shot each of the GOI and hpt cassette was used for the large-scale generation of transgenic wheat plants. In total, we isolated 15,496 immature embryos for 19 independent biolistic transformation experiments (Table 1). The average transformation frequency was 7.4% for GOI and 9.9% (between 3.1 and 20.3%) for hpt with a total of 1538 hygromycin resistant transgenic plants.
Table 1.
Summary of T0 transgenic wheat plants regenerated in 19 independent events of biolistic transformations
| Experiment | No. trans-formed embryos | Transgene copy No. | No. hygromycin resistant plants (HmR) | No. GOI transformants | GOI with one copy transformants (%) | Transformation frequency, TF, HmR (%) | Co-transformation, Co-TF (%) | |||
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | ≥3 | |||||||
| T01 | 857 | 18 | 19 | 4 | 17 | 58 | 40 | 47.5 | 6.8 | 69.0 |
| T02 | 998 | 36 | 29 | 7 | 40 | 112 | 76 | 38.2 | 11.2 | 67.9 |
| T03 | 1005 | 21 | 22 | 5 | 29 | 77 | 56 | 39.3 | 7.7 | 72.7 |
| T04 | 912 | 21 | 20 | 2 | 56 | 99 | 78 | 25.6 | 10.8 | 78.8 |
| T05 | 899 | 14 | 23 | 7 | 44 | 88 | 74 | 31.1 | 9.8 | 84.1 |
| T06 | 753 | 18 | 15 | 0 | 13 | 46 | 28 | 53.6 | 6.1 | 60.9 |
| T07 | 950 | 37 | 60 | 31 | 65 | 193 | 156 | 38.5 | 20.3 | 80.8 |
| T08 | 914 | 62 | 48 | 14 | 28 | 152 | 90 | 53.3 | 16.6 | 59.2 |
| T09 | 936 | 38 | 51 | 16 | 6 | 111 | 73 | 69.9 | 11.9 | 65.8 |
| T10 | 735 | 10 | 25 | 14 | 38 | 87 | 77 | 32.5 | 11.8 | 88.5 |
| T11 | 729 | 20 | 19 | 7 | 82 | 128 | 108 | 17.6 | 17.6 | 84.4 |
| T12 | 662 | 14 | 25 | 8 | 1 | 48 | 34 | 73.5 | 7.2 | 70.8 |
| T13 | 620 | 2 | 15 | 6 | 72 | 95 | 93 | 16.1 | 15.3 | 97.9 |
| T14 | 708 | 11 | 30 | 2 | 38 | 81 | 70 | 42.9 | 11.4 | 84.6 |
| T15 | 847 | 33 | 6 | 2 | 7 | 48 | 15 | 40.0 | 5.7 | 31.3 |
| T16 | 856 | 16 | 8 | 5 | 7 | 36 | 20 | 40.0 | 4.2 | 55.6 |
| T17 | 777 | 5 | 8 | 4 | 7 | 24 | 19 | 42.1 | 3.1 | 79.2 |
| T18 | 681 | 1 | 7 | 4 | 10 | 22 | 21 | 33.3 | 3.2 | 94.5 |
| T19 | 657 | 12 | 9 | 1 | 11 | 33 | 21 | 42.9 | 5.0 | 63.6 |
| Total | 15,496 | 389 | 439 | 139 | 571 | 1538 | 1149 | |||
| Transformation, % | 100 | 9.9 | 7.4 | 74.7 | ||||||
| GOI transformation, % | – | 38.2 | 12.1 | 49.7 | 100 | |||||
Data summarized for 19 stable transformation experiments. The transgene copy numbers of GOI in each event were determined by qPCR and confirmed by the Southern blot
The transgene copy numbers in each T0 plant were determined by qPCR and confirmed with a cross-examination of the same individuals using the Southern hybridisation. The example of the Southern blot is present in Fig. 4, where four out of eight T0 transgenic plants showed unique pattern of a single copy of the transgene insertion.
Fig. 4.
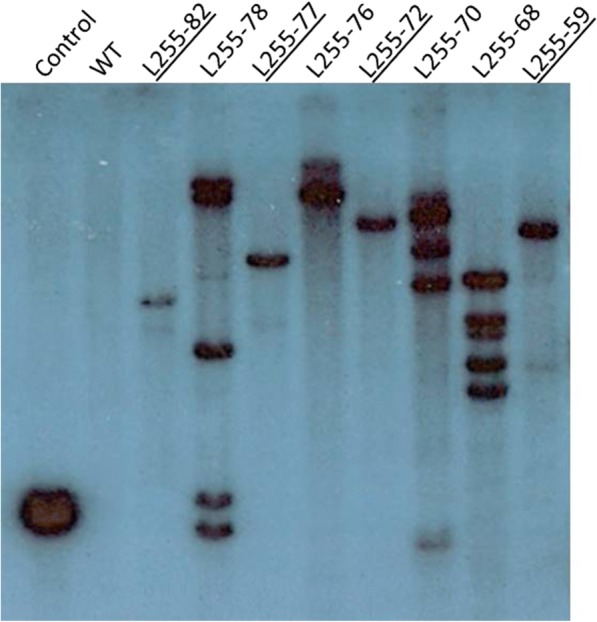
Example of Southern blot of transgenic wheat plants with different copy numbers of the transgene. Four out of the eight transgenic lines had single transgene insertions (underlined). Four other transgenic lines (not underlined) had multiple copy numbers of the transgene. A positive control, consisting of DNA from a barley plant with a single transgene insertion is indicated as the ‘Control’. The negative control is represented by wild-type (WT) wheat DNA (cv. Gladius)
Of all the transgenic plants, the frequency of single copy events varied from 16.1% in T13 to 73.5% in T12 with an average of 38.2%. In experiment T07, the number of single copy events reached 60, in comparison to 15 in experiments T06 and T13. Interestingly, the total number of two copy events was 139, which was three-fold lower than the number of single copy events (439). The in vitro tissue culture period from the isolation of embryos to the regeneration of transgenic plants was between 140 and 190 days (Fig. 5).
Fig. 5.

Biolistic wheat transformation process with hygromycin selection. a Freshly isolated immature embryo; (b) 2 week-old embryogenic callus before bombardment; (c) Shoot regeneration on 50 mg/L hygromycin selection medium; (d) Shoot and root regeneration on 50 mg/L hygromycin selection medium; (e) Plantlet recovery in 1/2 MS medium; (f) T0 transgenic plants in the greenhouse
Discussion
The PEG/Mg2+ coating procedure in the current study showed improved results compared to the standard Spd/Ca2+ protocol in transient transformation. The Bio-Rad coating method relies on high quality spermidine which can differ from batch to batch. Spermidine solutions are hydroscopic, oxidisable, deaminated with time and, therefore, the frozen aliquots should be made fresh at least once a month [16, 20]. The stock solutions of CaCl2 and spermidine cannot be combined together in one solution and must be used separately. In contrast, PEG and magnesium salts can be conveniently made in a single stock solution that remains stable for years when stored at − 20 °C. Gold/DNA coating in transformation experiments is done by simple mixing 50 μl gold suspension with 10 μl DNA and 10 μl of PM solution under vortexing at room temperature. To date, the PEG/Mg2+ procedure described here has been applied successfully in stable transformation of spring wheat cvs. Bobwhite, Akadaruma [41] and Gladius using embryogenic callus cultures as well as in producing transgenic winter wheat, durum wheat, maize, sorghum, pearl millet and eastern gamagrass (data not shown).
A study in rice found that 80% of transgenic lines transformed with minimal cassette DNA showed simple integration patterns, compared with 20–30% of lines transformed with plasmid DNA [25]. The use of minimal DNA cassettes instead of plasmid DNA in biolistic transformation almost tripled the frequency of wheat stable transformation from 0.4 to 1.1% [42]. Furthermore, when nanogram amounts of DNA cassette were employed (25 ng and 2.5 ng per shot [26], the regeneration of low copy transgenic maize plants became more frequent. In sugarcane, bombardment with 6.6 ng per shot of the minimal cassettes resulted in over 30% of low copy integration events [28]. In the present study, co-transformation frequencies (co-TFs) of the GOI and selective marker minimal expression cassettes were between 31.3 and 97.9% with an average of 74.7%, which is comparable to the results in sugarcane using 6.6 ng of DNA/shot (79–84%) [28] and in wheat using 200 ng of DNA/shot (91.7%) [42]. The large difference in co-TFs in our experiments is likely due to the impact of various GOIs on co-integration and plant regeneration.
The efficiency of wheat biolistic transformation remains relatively low at 1–5% [43]. In recent publications, the use of 5 ng per shot of the expression cassettes in the model wheat cv. Bobwhite S26 resulted in 50–60% of transformants with simple integration pattern at a TF of 2.7–4.0% [44]. In our experiments with the commercial wheat cv. Gladius, the TFs ranged from 3.1 to 20.3%. TFs below 5% were observed only in three experiments (T16-T18) out of 19. Variations in TFs between experiments can be attributed to seasonal fluctuations in the quality of immature embryos, to GOI effects on plant transformation, to changes in tissue culture conditions due to batch-to-batch differences in reagent quality, etc.
We produced between 16.1–73.5% single copy events (Table 1) by using 6 ng of minimal GOI DNA cassettes. This is comparable to the 41–80% reported in transformation of maize (2.5 ng DNA cassette per shot) [45], and to the 49.2% for sugarcane transformation (2.73 ng DNA cassette per shot) [29]. The results suggest that the use of nanogram amounts of DNA cassette is desirable when seeking to regenerate plants with low copy insertion of the transgene. However, the optimal quantity for each plant species in biolistic transformation needs more investigation [46].
Conclusions
The use of PEG and Mg2+ instead of Spd and Ca2+ in the conventional biolistic coating procedure was developed to transform wheat and other plant species in both transient and stable transformation. In bombarding a low quantity of minimal DNA cassette was reported to efficiently regenerate low copy transgenic plants. Nanogram amounts of the minimal expression cassettes of the GOI and the hpt gene were routinely used in high-throughput experiments to generate single copy transgenic plants of commercial wheat at a high frequency.
Acknowledgments
This manuscript is presented dedicated to thein memory of Dr. Ainur Ismagul. Ainur died tragically in early 2015 and is sorely missed by her family and friends. She played a key role in developing a highly efficient cereal transformation capability and in building research ties between Australia and Kazakhstan. Her contribution lives on. We thank Yuan Li and Hui Zhou for copy number analysis, Peter Quail for kindly providing the pAHC25 genetic construct and Ursula Langridge for work with plants in the greenhouse. We also thank Carly Schramm for critical comments in the manuscript.
Funding
This work research and publication costs were supported by funding to the Australian Centre for Plant Functional Genomics (ACPFG), the University of Adelaide, from the Australian Research Council and the Grains Research and Development Corporation Australia and by Research program BR05236500 of the Ministry of Education and Science, Kazakhstan.
Availability of data and materials
All data generated or analysed during this study are included in this published article.
Abbreviations
- CoTF
Co-transformation frequency
- CP
Centrophenoxine
- GOI
Gene of Interest
- GUS
β-glucuronidase A gene
- Hpt
Hygromycin resistance gene
- MS
Murashige and Skoog medium
- PEG
Polyethyleneglycol
- psi
Pounds per square inch, a unit of pressure
- Spd
Spermidine
- TF
Transformation frequency
- X-gluc
5-Bromo-4-chloro-3-indolyl-β-D-glucuronide, a reagent to detect β-glucuronidase
Authors’ contributions
AI organised and coordinated all experiments, NY and YSk carried out bombardment experiments, HB made Southern hybridisation and autoradiography, EM, GI and IM analysed wheat transformants, NY and SE made mathematical treatment and first manuscript writing, YSh and SJ checked data, prepared Figures and managed with the manuscript, NB prepared genetic constructs, PL supervised the project. All authors read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Footnotes
Ainur Ismagul is deceased. This paper is dedicated to his memory.
Contributor Information
Ainur Ismagul, Email: ainur.ismagul@mail.ru.
Nannan Yang, Email: nannan.yang@dpi.nsw.gov.au.
Elina Maltseva, Email: elina_m@inbox.ru.
Gulnur Iskakova, Email: g.iskakova83@gmail.com.
Inna Mazonka, Email: imazonka@gmail.com.
Yuri Skiba, Email: yuriy.skiba@gmail.com.
Huihui Bi, Email: bihuihui826@126.com.
Serik Eliby, Email: serik.eliby@mail.ru.
Satyvaldy Jatayev, Email: satidjo@gmail.com.
Yuri Shavrukov, Email: yuri.shavrukov@flinders.edu.au.
Nikolai Borisjuk, Email: nborisjuk@yahoo.com.
Peter Langridge, Phone: +61-8-8313-7468, Email: peter.langridge@adelaide.edu.au.
References
- 1.Cho MJ, Yano H, Okamoto D, Kim HK, Jung HR, Newcomb K, et al. Stable transformation of rice (Oryza sativa L.) via microprojectile bombardment of highly regenerative, green tissues derived from mature seed. Plant Cell Rep. 2004;22(7):483–489. doi: 10.1007/s00299-003-0713-7. [DOI] [PubMed] [Google Scholar]
- 2.Wang K, Frame B. Biolistic gun-mediated maize genetic transformation. Methods Mol Biol. 2009;526:29–45. doi: 10.1007/978-1-59745-494-0_3. [DOI] [PubMed] [Google Scholar]
- 3.Zhang K, Liu J, Zhang Y, Yang Z, Gao C. Biolistic genetic transformation of a wide range of Chinese elite wheat (Triticum aestivum L.) varieties. J Genet Genomics. 2015;42:39–42. doi: 10.1016/j.jgg.2014.11.005. [DOI] [PubMed] [Google Scholar]
- 4.Vasil V, Castillo AM, Fromm ME, Vasil IK. Herbicide resistant fertile transgenic wheat plants obtained by microprojectile bombardment of regenerable embryogenic callus. Bio/Technol. 1992;10:667–674. [Google Scholar]
- 5.Vasil V, Srivastava V, Castillo AM, Fromm ME, Vasil IK. Rapid production of transgenic wheat plants by direct bombardment of cultured immature embryos. Bio/Technol. 1993;11:1553–1558. [Google Scholar]
- 6.Weeks JT, Anderson OD, Blechl AE. Rapid production of multiple independent lines of fertile transgenic wheat (Triticum aestivum L.) Plant Physiol. 1993;102:1077–1084. doi: 10.1104/pp.102.4.1077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Nehra NS, Chibbar RN, Leung N, Caswell K, Mallard C, Steinhauer L, et al. Self-fertile transgenic wheat plants regenerated from isolated scutellar tissues following microprojectile bombardment with two distinct gene constructs. Plant J. 1994;5:285–297. doi: 10.1046/j.1365-313X.1994.05020285.x. [DOI] [Google Scholar]
- 8.Zhou H, Arrowsmith JW, Fromm ME, Hironaka CM, Taylor ML, Rodriguez D, et al. Glyphosate-tolerant CP4 and GOX genes as a selectable marker in wheat transformation. Plant Cell Rep. 1995;15:159–163. doi: 10.1007/BF00193711. [DOI] [PubMed] [Google Scholar]
- 9.Altpeter F, Vasil V, Srivastava V, Stoger E, Vasil IK. Accelerated production of transgenic wheat (Triticum aestivum L.) plants. Plant Cell Rep. 1996;16:12–17. doi: 10.1007/BF01275440. [DOI] [PubMed] [Google Scholar]
- 10.Ortiz JPA, Reggiardo MI, Ravizzini RA, Altabe SG, Cervigni GDL, Spitteler MA, et al. Hygromycin resistance as an efficient selectable marker for wheat stable transformation. Plant Cell Rep. 1996;15:877–881. doi: 10.1007/BF00231579. [DOI] [PubMed] [Google Scholar]
- 11.Takumi S, Shimada T. Production of transgenic wheat through particle bombardment of scutellar tissues: frequency is influenced by culture duration. J Plant Physiol. 1996;149:418–423. doi: 10.1016/S0176-1617(96)80143-X. [DOI] [Google Scholar]
- 12.Barro F, Cannell ME, Lazzeri PA, Barcelo P. The influence of auxins on transformation of wheat and tritordeum and analysis of transgene integration patterns in transformants. Theor Appl Genet. 1998;97:684–695. doi: 10.1007/s001220050944. [DOI] [Google Scholar]
- 13.Yao Q, Cong L, Chang JL, Li KX, Yang GX, He GY. Low copy number gene transfer and stable expression in a commercial wheat cultivar via particle bombardment. J Exp Bot. 2006;57:3737–3746. doi: 10.1093/jxb/erl145. [DOI] [PubMed] [Google Scholar]
- 14.Kovalchuk N, Smith J, Pallotta M, Singh R, Ismagul A, Eliby S, et al. Characterization of the wheat endosperm transfer cell-specific protein TaPR60. Plant Mol Biol. 2009;71:81–98. doi: 10.1007/s11103-009-9510-1. [DOI] [PubMed] [Google Scholar]
- 15.Ismagul A, Iskakova G, Harris JC, Eliby S. Biolistic transformation of wheat with centrophenoxine as a synthetic auxin. In: Fleury D, Whitford R, editors. Crop breeding: methods and protocols, methods in molecular biology. New York: Humana Press; 2014. pp. 191–202. [DOI] [PubMed] [Google Scholar]
- 16.Sanford JC, Smith FD, Russel JA. Optimizing the biolistic process for different biological applications. Methods Enzymol. 1993;217:483–509. doi: 10.1016/0076-6879(93)17086-K. [DOI] [PubMed] [Google Scholar]
- 17.Kikkert JR. Biolistic transformation of plant cells. In: Celis JE, editor. Cell biology: a laboratory handbook, vol. 4. San Diego: Academic Press; 1998. pp. 157–161. [Google Scholar]
- 18.Rasco-Gaunt S, Riley A, Barcelo P, Lazzeri PA. Analysis of particle bombardment parameters to optimise DNA delivery into wheat tissues. Plant Cell Rep. 1999;19:118–127. doi: 10.1007/s002990050721. [DOI] [PubMed] [Google Scholar]
- 19.Kohli A, Gahakwa D, Vain P, Laurie DA, Christou P. Transgene expression in rice engineered through particle bombardment: molecular factors controlling stable expression and transgene silencing. Planta. 1999;208:88–97. doi: 10.1007/s004250050538. [DOI] [Google Scholar]
- 20.Sparks CA, Jones HD. Transformation of wheat by biolistics. In: Curtis IP, editor. Transgenic crops of the world – Essential Protocols. Dordrecht: Kluwer; 2004. pp. 19–35. [Google Scholar]
- 21.Srivastava V, Vasil V, Vasil IK. Molecular characterization of the fate of transgenes in transformed wheat (Triticum aestivum L.) Theor Appl Genet. 1996;92:1031–1037. doi: 10.1007/BF00224045. [DOI] [PubMed] [Google Scholar]
- 22.Stoger E, Williams S, Keen D, Christou P. Molecular characteristics of transgenic wheat and the effect on transgene expression. Transgenic Res. 1998;7:463–471. doi: 10.1023/A:1008833324193. [DOI] [Google Scholar]
- 23.Agrawal PK, Kohli A, Twyman RM, Christou P. Transformation of plants with multiple cassettes generates simple transgene integration patterns and high expression levels. Mol Breed. 2005;16:247–260. doi: 10.1007/s11032-005-0239-5. [DOI] [Google Scholar]
- 24.Jones HD. Wheat transformation: current technology and applications to grain development and composition. J Cereal Sci. 2005;41:137–147. doi: 10.1016/j.jcs.2004.08.009. [DOI] [Google Scholar]
- 25.Fu XD, Duc LT, Fontana S, Bong BB, Tinjuangjun P, Sudhakar D, et al. Linear transgene constructs lacking vector backbone sequences generate low-copy-number transgenic plants with simple integration patterns. Transgenic Res. 2000;9:11–19. doi: 10.1023/A:1008993730505. [DOI] [PubMed] [Google Scholar]
- 26.Lowe BA, Prakash NS, Way M, Mann MT, Spencer TM, Boddupalli RS. Enhanced single copy integration events in corn via particle bombardment using low quantities of DNA. Transgenic Res. 2009;18:831–840. doi: 10.1007/s11248-009-9265-0. [DOI] [PubMed] [Google Scholar]
- 27.O'Kennedy MM, Stark HC, Dube N. Biolistic-mediated transformation protocols for maize and pearl millet using pre-cultured immature zygotic embryos and embryogenic tissue. Methods Mol Biol. 2011;710:343–354. doi: 10.1007/978-1-61737-988-8_23. [DOI] [PubMed] [Google Scholar]
- 28.Jackson MA, Anderson DJ, Birch RG. Comparison of Agrobacterium and particle bombardment using whole plasmid or minimal cassette for production of high-expressing, low-copy transgenic plants. Transgenic Res. 2013;22(1):143–151. doi: 10.1007/s11248-012-9639-6. [DOI] [PubMed] [Google Scholar]
- 29.Wu H, Awan FS, Vilarinho A, Zeng Q, Kannan B, Phipps T, et al. Transgene integration complexity and expression stability following biolistic or Agrobacterium-mediated transformation of sugarcane. In Vitro Cell Devel Biol Plant. 2015;51:603–611. doi: 10.1007/s11627-015-9710-0. [DOI] [Google Scholar]
- 30.Murashige T. Skoog F. A revised medium for rapid growth and bioassay with tobacco tissue culture. Physiol Plant. 1962;15:473–497. doi: 10.1111/j.1399-3054.1962.tb08052.x. [DOI] [Google Scholar]
- 31.Shimada T, Sasakuma T, Tsunewaki K. In vitro culture of wheat tissues. I. Callus formation, organ redifferention and single cell culture. Can J Genet Cytol. 1969;11:294–304. doi: 10.1139/g69-037. [DOI] [Google Scholar]
- 32.Christensen AH, Quail PH. Ubiquitin promoter-based vectors for high-level expression of selectable and/or screenable marker genes in monocotyledonous plants. Transgenic Res. 1996;5:213–218. doi: 10.1007/BF01969712. [DOI] [PubMed] [Google Scholar]
- 33.Jefferson RA. Assaying chimeric genes in plants: the GUS gene fusion system. Plant Mol Biol Rep. 1987;5(4):387–405. doi: 10.1007/BF02667740. [DOI] [Google Scholar]
- 34.Eini O, Yang N, Pyvovarenko T, Pillman K, Bazanova N, Tikhomirov N, et al. Complex regulation by Apetala2 domain-containing transcription factors revealed through analysis of the stress-responsive TdCor410b promoter from durum wheat. PLoS One. 2013;8:e58713. doi: 10.1371/journal.pone.0058713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kikkert JR, Vidal JR, Reisch BI. Stable transformation of plant cell by particle bombardment/biolistics. In: Peña L, editor. Transgenic plants: methods and protocols. Totowa: Humana Press; 2004. pp. 61–78. [Google Scholar]
- 36.Shavrukov Y, Gupta NK, Miyazaki J, Baho MN, Chalmers KJ, Tester M, et al. HvNax3 – a locus controlling shoot sodium exclusion derived from wild barley (Hordeum vulgare ssp. spontaneum) Funct Integr Genomics. 2010;10:277–291. doi: 10.1007/s10142-009-0153-8. [DOI] [PubMed] [Google Scholar]
- 37.Pallotta MA, Graham RD, Langridge P, Sparrow DHB, Barker SJ. RFLP mapping of manganese efficiency in barley. Theor Appl Genet. 2000;101:1100–1108. doi: 10.1007/s001220051585. [DOI] [Google Scholar]
- 38.Fletcher SJ. qPCR for quantification of transgene expression and determination of transgene copy number. In: Fleury D, Whitford R, editors. Crop breeding: methods and protocols, methods in molecular biology. New York: Humana Press; 2014. pp. 213–238. [DOI] [PubMed] [Google Scholar]
- 39.Maas C, Werr W. Mechanism and optimized conditions for PEG mediated DNA transfection into plant protoplasts. Plant Cell Rep. 1989;8:148–151. doi: 10.1007/BF00716828. [DOI] [PubMed] [Google Scholar]
- 40.Paithankar KR, Prasad KSN. Precipitation of DNA by polyethylene glycol and ethanol. Nucleic Acids Res. 1991;19:1346. doi: 10.1093/nar/19.6.1346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Eliby S, Kandzia R, Ismagul A, Karabaev M, Sasakuma T, Klimyuk V. Biolistic transformation of wheat utilizing a new DNA/gold coating procedure. Budapest, Hungary: Abstracts of the 6th International Wheat Conference; 2000. p. 313. [Google Scholar]
- 42.Yao Q, Cong L, He G, Chang J, Li K, Yang G. Optimization of wheat co-transformation procedure with gene cassettes resulted in an improvement in transformation frequency. Mol Biol Rep. 2007;34:61–67. doi: 10.1007/s11033-006-9016-8. [DOI] [PubMed] [Google Scholar]
- 43.Abdul R, Ma Z, Wang H. Genetic transformation of wheat (Titicum aestivum L.), a review. Triticeae Genomics Genet. 2010;1:1–7. [Google Scholar]
- 44.Tassy C, Partier A, Beckert M, Feuillet C, Barret P. Biolistic transformation of wheat: increased production of plants with single insertions and heritable transgene expression. Plant Cell Tissue Organ Cult. 2014;119:171–181. doi: 10.1007/s11240-014-0524-2. [DOI] [Google Scholar]
- 45.Prakash NS, Bhojaraja R, Shivbachan SK, Hari Priya GG, Nagraj TK, Prasad V, et al. Marker-free transgenic corn plant production through co-bombardment. Plant Cell Rep. 2009;28:1655–1668. doi: 10.1007/s00299-009-0765-4. [DOI] [PubMed] [Google Scholar]
- 46.Altpeter F, Springer NM, Bartley LE, Blechl AE, Brutnell TP, Citovsky V, et al. Advancing crop transformation in the era of genome editing. Plant Cell. 2016;28:1510–1520. doi: 10.1105/tpc.16.00196. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data generated or analysed during this study are included in this published article.