

Abstract
The androgen receptor (AR) has context-dependent roles in breast cancer growth and progression. Overall, high tumor AR levels predict a favorable patient outcome, but several studies have established a tumor promotional role for AR, particularly in supporting the growth of estrogen receptor positive (ER-positive) breast cancers after endocrine therapy. Our previous studies have demonstrated that obesity promotes mammary tumor progression after ovariectomy (OVX) in a rat model of postmenopausal breast cancer. Here, we investigated a potential role for AR in obesity-associated post-OVX mammary tumor progression following ovarian estrogen loss. In this model, we found that obese but not lean rats had nuclear localized AR in tumors that progressed 3 weeks after OVX, compared to those that regressed. AR nuclear localization is consistent with activation of AR-dependent transcription. Longer-term studies (8 weeks post-OVX) showed that AR nuclear localization and expression were maintained in tumors that had progressed, but AR expression was nearly lost in tumors that were regressing. The anti-androgen enzalutamide effectively blocked tumor progression in obese rats by promoting tumor necrosis and also prevented the formation of new tumors after OVX. Neither circulating nor mammary adipose tissue levels of the AR ligand testosterone were elevated in obese compared to lean rats; however, IL-6, which we previously reported to be higher in plasma from obese versus lean rats, sensitized breast cancer cells to low levels of testosterone. Our study demonstrates that, in the context of obesity, AR plays a role in driving ER-positive mammary tumor progression in an environment of low estrogen availability, and that circulating factors unique to the obese host, including IL-6, may influence how cancer cells respond to steroid hormones.
Electronic supplementary material
The online version of this article (doi:10.1007/s12672-017-0302-9) contains supplementary material, which is available to authorized users.
Keywords: Obesity, Breast cancer, Androgen receptor, Adipose tissue, Steroid hormone, Estradiol, Testosterone, IL-6
Introduction
Obesity, which now affects 40% of US adult women [1], increases the incidence of and worsens the outcomes for breast cancer [2, 3]. Some controversy exists regarding the effect of obesity on breast cancer risk before menopause, but studies have definitively shown elevated risk in obese, postmenopausal women (reviewed in [4]), particularly for estrogen receptor (ER) positive disease [2, 5]. Obese women typically present with larger tumors and more frequent lymph node involvement [6], and are less likely to benefit from traditional anti-hormonal therapies, such as tamoxifen or aromatase inhibitors [7]. Further, regardless of menopausal status, obese women with breast cancer are more likely to die from the disease compared to their lean counterparts [2]. These observations strongly suggest that obesity-related breast cancer is unique and implicate a role for altered steroid receptor signaling in the obesogenic setting. Given that over 200,000 women are diagnosed with ER-positive breast cancers annually [8] and undergo treatment with agents that aim to reduce estrogen production or interfere with ER activity, collectively referred to as endocrine therapy, the high prevalence of obesity argues that it is critical that we understand how obesity acts as a biological variable in treatment modalities and therapeutic outcomes.
Recently, the androgen receptor (AR) has emerged as a potential driver of breast cancer resistance to endocrine therapy in preclinical studies [9–11], despite a documented link to a favorable patient outcome for those with ER-positive breast cancer [12–14]. Overall, AR is more frequently expressed in breast tumors than ER or the progesterone receptor (PR), occurring in up to 80% of invasive and approximately 85% of in situ cancers [12, 13]. Like ER, AR is inversely associated with features of malignancy including tumor size, Ki67 index, lymph node metastasis, and histological grade [13]. While it seems counter-intuitive that AR can be tumor-promotional, the same inverse relationships exist for ER. Preclinical studies show that the function of AR in breast tumor growth and progression depends on the cellular context, the activation state of the receptor, the levels of ER, and the hormonal milieu of the host [15, 16]. For example, in ER-positive tumors, a high ratio of AR compared to ER (percent cells positive by IHC) indicated a greater than fourfold increased risk for failure while on tamoxifen [15]. In triple negative breast cancer (TNBC), the prognostic significance of AR also remains somewhat controversial (reviewed in [17]). TNBC with high AR are more indolent and slow growing, and, likely because they are slower growing, they are also less likely to achieve pathological complete response to chemotherapy than most other subtypes of TNBC [18]. Overall, recent preclinical and clinical studies suggest AR is a viable therapeutic target for a subset of patients with TNBC and recurrent ER-positive breast cancer [15, 19–21], an avenue being investigated in ongoing clinical trials.
Here, we investigated a potential role for AR in obesity-associated tumor progression using a preclinical rat model of obesity and postmenopausal ER-positive breast cancer. In our model, a high fat diet (HFD) is used to separate obesity-prone and -resistant females, representing the top and bottom tertiles of body fat percentage, respectively [22]. N-methyl-N-nitrosourea (MNU) is used to initiate mammary tumors and ovariectomy (OVX) simulates a postmenopausal environment. After OVX, obese rats have more tumors that progress and fewer tumors that regress and develop more new tumors compared to lean rats [23]. In a more recent study, we identified a 3-week window following OVX as a period of rapid weight gain that associates with tumor progression and high tumor PR expression [24]. Here, we extend these studies to demonstrate that AR nuclear localization, which is indicative of AR transcriptional activation, is associated with early post-OVX tumor progression only in obese rats, and that treating these rats with the anti-androgen enzalutamide reduced PR expression and prevented tumor growth by stimulating tumor necrosis. We also identified a role for IL-6, a pro-inflammatory obesogenic cytokine, in sensitizing breast cancer cells to low levels of testosterone. Altogether, these data indicate that the obese environment enhances AR-dependent mammary tumor progression after the loss of ovarian estrogen production. These data support therapeutic targeting of AR to circumvent breast cancer resistance to ER-directed endocrine therapy and argue that this approach may be particularly efficacious in the context of obesity. Indeed, such clinical trials are underway which may be able to address whether obesity is a factor in the response to targeting AR in breast cancer.
Methods
Animal Care and Treatment
Female Wistar rats (5 weeks of age) were purchased from Charles River Laboratories (Wilmington, MA) and individually housed in metabolic caging at the University of Colorado Denver Center for Comparative Medicine and the Center for Human Nutrition Satellite Facility (22–24 C; 12:12 h light-dark cycle) with free access to food and water. To induce obesity, rats were maintained on a purified high-fat diet (46% kcal fat; Research Diets, New Brunswick, NJ; RD# D12344) for the duration of the study. All procedures were approved by the Institutional Animal Care and Use Committee.
OP-OR/OVX Model of Obesity and Postmenopausal Breast Cancer
This study used our previously characterized model of obesity and postmenopausal breast cancer [22, 25]. Briefly, rats were given a single injection of N-methyl-N-nitrosourea (MNU, 50 mg/kg) at 50 ± 2 days of age to induce mammary tumor formation. MNU was purchased by the National Cancer Institute’s Chemical Carcinogen Reference Standards Repository operated under contract by Midwest Research Institute, Kansas City, MO (N02-CB-07008). The outbred Wistar strain of rats used have a variable response to HFD, and rats were ranked by percent body fat at 14–18 weeks of age as previously described [24]. Rats in the top and bottom tertiles of body fat were considered obese and lean, respectively; rats from the middle tertile were not used for this study. Rats were palpated weekly for tumor formation beginning 3 weeks after MNU injection, and tumors were measured in three dimensions using digital calipers. Rats entered the treatment phase of the study in cohorts based on tumor burden (≥1 cm3), at which time rats were ovariectomized (OVX) under isoflurane anesthesia. Body weight and food intake were monitored weekly before surgery and daily thereafter [26, 27]. Body composition was determined at OVX and every other week thereafter for the duration of the study by quantitative magnetic resonance (qMR; Echo MRI Whole Body Composition Analyzer; Echo Medical Systems, Houston, TX). Enzalutamide was provided to Research Diets, Inc. by Astellas, Inc. and Medivation, Inc., (Medivation, Inc. was acquired by Pfizer, Inc. in September 2016) and incorporated into the purified HFD (Research Diets, Inc.; D12344) at a concentration of 0.43 mg/g food, for oral administration [15, 16]. Control rats were fed the same HFD without enzalutamide. Rats were assigned to either the control or enzalutamide group based on several characteristics, including tumor burden, tumor multiplicity, body weight, and body fat percent at the time of OVX [24, 28].
Plasma Measurements
Blood was drawn under isoflurane anesthesia at the time of euthanasia from the anterior vena cava using a heparinized needle, and plasma was collected. All blood draws were made during the latter part of the light cycle. Insulin, leptin, cholesterol, non-esterified fatty acids (NEFA), triglycerides, and glucose were measured using commercially available assays as previously described [28]. ELISA was used for testosterone measurements (IBL America).
Immunohistochemistry
IHC was performed on formalin-fixed, paraffin-embedded tissue sections using standard histologic procedures. Antibodies against ER (Leica Microsystems; clone 6F11; 1:100), PR (Dako North America; A0098; 1:750), AR (Abcam; ab133273; 1:1000), Ki67 (Thermo Scientific; clone SP6, 1:400), and TUNEL (ApopTag Plus InSitu Apoptosis Detection Kit; EMD Millipore) were purchased commercially. Target retrieval solution, protein blocking solution, and secondary antibodies were all purchased from Dako North America. Steroid hormone receptor levels were quantified manually in blinded samples (ER, PR, or AR) or using the Aperio Digital Pathology System and ImageScope software (Leica Biosystems; Ki67, TUNEL). For ER and PR, the Allred scoring system was used as we have done previously [24]. For AR, stained tumor sections were individually evaluated for percentage of strong, medium, and weak positive cells, and these data were analyzed using a scoring system based on that reported by Remmele et al. [29] and that has been previously described in detail for primary breast tumors [30]. The scoring criteria for AR are as follows, according to Feng et al. [30]:
-
Staining Intensity (SI):
0: no staining,
weak staining,
2: moderate staining,
-
3:
strong staining;
-
Percentage of Positive Cells (PP):
0: no positive cells
fewer than 10% positive,
11-50% positive,
51-80% positive,
>80% positive;
Score for either nuclear or cytoplasmic AR = SI × PP; AR localization score = AR nuclear score − AR cytoplasmic score.
Tissue Hormone Measurements: Sample Preparation
Pulverized tissue samples (100 mg) were spiked with 10 μL of internal standards (10 ng/mL Testosterone-D3). Two milliliters of methanol were added to the sample, sonicated for 20 min at 50 °C, and then placed in a -20 °C freezer for 1 hour. Following the freezer incubation, the sample was centrifuged at 2000 rpm for 20 min, and the supernatant was collected in a clean conical glass vial. The supernatant was evaporated to dryness with nitrogen using a N-EVAP with a 50 °C water bath and reconstituted with 500 μL of acetonitrile. The sample was further cleaned up with dispersive solid phase extraction. Fifty milligrams of Universal dSPE (Agilent Technologies) was added, vortexed for 30 s, and centrifuged at 2000 rpm for 20 min. The supernatant was collected, evaporated to dryness with nitrogen using a N-EVAP with a 50 °C water bath, and reconstituted with 200 μL of 50:50 acetonitrile:water for analysis of testosterone by LC-MS/MS. For analysis of estradiol, dry samples were reconstituted in 100 μL of 1 mg/mL dansyl chloride and 100 μL of 1 mg/mL sodium carbonate. Samples were then incubated for 15 min at 60 °C and then analyzed by LC-MS/MS.
Liquid Chromatography-Mass Spectrometry
The instrument used in the analysis was an Agilent 1290 UPLC coupled to an Agilent 6460 triple quadruple mass spectrometer, which was equipped with an ESI source using Agilent Jet Stream Technology (Agilent, Santa Clara, CA). Estradiol and testosterone were separated on an Agilent Eclipse Plus C18 column (2.1 × 100 mm, 3.5 μm). A sample volume of 10 μL was injected and a mixture of water with 0.1% Formic acid (A) and acetonitrile with 0.1% formic acid (B) at a flow rate of 0.4 mL/min. The gradient used was 20% B increasing to 50% B at 5 min, followed by and increase to 80% B at 7 min, and held at 80% B for 3 min. The column temperature was held at 40 °C. The ionization source conditions used were as follows: nebulizer 35 psi, gas flow of 10 L/min at 350 °C, sheath gas flow of 11 L/min at 350 °C, and the capillary voltage (+)3500 V. Ionization polarity was set to positive. The ion transitions (m/z) monitored were 506→171 and 506→156 for estradiol, 289→109/97 for testosterone, and 292→109/97 for testosterone-D3. Compound identifications were confirmed by retention time, and the product ion ratio correlation between the sample peaks and corresponding standards (±20%). The data collection and processing were performed by using Agilent MassHunter Quantitative software (v.B.06.01). Quantitation was performed with linear regression using a 6-point calibration curve from 50 to 3000 pg/mL for estradiol and 10–600 pg/mL for testosterone.
Patient Samples
Normal breast adipose tissue samples were collected under an approved Colorado Institutional Review Board protocol from patients undergoing mastectomy surgery at the University of Colorado Hospital. Tissues were from postmenopausal women. Immediately after removal, tissue samples were cut into ~100 mg pieces, weighed, snap frozen in liquid nitrogen, and stored at −80C for later analysis.
Rat Tissue Q-PCR Analysis
Total RNA was isolated from rat ovary, mammary adipose (MG), or mammary tumor tissue using Trizol, followed by column clean-up (Qiagen). cDNA was prepared using the Verso kit (Thermo Scientific), and qPCR was performed using Taqman reagents, with commercially available predesigned Cyp19a1 primer/probe set (Rn00567222_m1; Life Technologies).
Adipose Cellularity
Adipose tissue cell size distribution was determined using the Adiposoft ImageJ plug-in on one full H&E stained section of subcutaneous/mammary or retroperitoneal adipose tissue. Frequency distributions and ANOVA were calculated using Prizm software.
Cell Culture Studies
Cell lines were obtained from the University of Colorado Cancer Center Protein Production, Monoclonal Antibody and Tissue Culture Shared Resource, and authenticity was validated using STR analysis within 2 months of conducting experiments. For standard growth, cells were cultured in DMEM supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin. During experiments, cells were cultured in phenol red-free, glucose-free DMEM (Gibco, A14430–01) supplemented with 1% penicillin/streptomycin, 5 mM glucose, 2 mM L-glutamine, 5 ng/mL human insulin, 1 mM sodium pyruvate, and 10 μM HEPES buffer. Cells were plated in 6-well plates and allowed to adhere overnight in standard growth medium. The next morning, medium was aspirated, cells were washed, and then incubated for 3 h in experimental medium with or without 1 ng/mL recombinant human IL-6 (R&D Systems). Cells were then treated with varying doses of testosterone (Sigma Aldrich), or EtOH vehicle, in the presence or absence of IL-6 (1 ng/mL; R&D Systems) for 24 h. For inhibitor studies, cells were also treated with either fulvestrant (ICI 182780; 100 nM) or enzalutamide (Selleck Chemicals; 10 μM). RNA was isolated using Qiazol Lysis Reagent (Qiagen), and cDNA was synthesized using the Verso kit (Thermo Scientific). Q-PCR analysis was performed using Sybr Green (Fkbp5, Klk3, Tmprss2) or Taqman (Polr2a, Cyp19a1, Esr1) reagents. Primers for Fkbp5 were (forward) 5′ TGA GCA GGG AGA GGA TAT TAC C and (reverse) 5′ TCT CCA ATC ATC GGC GTT TC. Primers for Klk3 were (forward) 5′ GAT GCT GTG AAG GTC ATG GA and (reverse) 5′ GAG GTC CAC ACA CTG AAG TT. Primers for Tmprss2 were (forward) 5′ AGA CGA CTG GAA CGA GAA CTA and (reverse) 5′ GAT CCG CTG TCA TCC ACT ATT C. Pre-designed primers and FAM-labeled probe for RNA-polymerase II (Polr2a; Hs00172187_m1), aromatase (Cyp19a1; Hs00903410_m1), and ERα (Esr1; Hs00174860_m1) were purchased commercially (Life Technologies). Gene expression was analyzed as transcript copies based on a standard curve method as we have previously described [31, 32].
Human Breast Cancer Sample Analysis
The dataset used for primary breast tumor analysis was previously published by others [33]. Normalized gene expression data from primary ER-positive human breast tumors were downloaded from NCBI Gene Expression Omnibus (GEO; GSE54430). Samples were sorted by patient serum IL-6 level, categorized as high (≥10 pg/mL) or low (<10 pg/mL). AR and Fkbp5 expression is reported as mean ± SEM log2-transformed values. The data can be accessed through the following link: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE54330
Statistical Analysis
Statistical analyses were performed using Graph Pad Prizm 6.0 software. Student’s t tests or ANOVA tests were used for normally distributed data, and Mann-Whitney tests were used for non-normal data. Unless otherwise indicated, data are presented as mean ± standard error of the mean. A chi-square test was used to compare the number of regressing, progressing, and newly formed tumors between control and enzalutamide-treated rats. Significance level was set at less than or equal to 0.05.
Results
AR Nuclear Localization Associates with Post-OVX Tumor Progression in Obese Rats
We previously showed in our preclinical rat model that obesity in combination with OVX-induced weight gain supports tumor progression, which was associated with elevated PR expression, a glycolytic gene expression profile, and elevated tumor glucose uptake [24]. In that study, juvenile rats were placed on a high fat diet (HFD); MNU was used to induce mammary tumorigenesis, and mature rats were separated into obesity resistant (lean) and obesity prone (obese) phenotypes [24]. At a defined tumor burden (≥1 cm3), rats underwent OVX, and tumors were monitored for three additional weeks (Fig. 1a). To examine the role of AR in tumor progression following OVX, tumor sections from lean and obese rats were stained for AR using IHC. We found AR was expressed in most tumors. However, subcellular localization of AR showed two dominant patterns, either primarily localized to the nucleus (Fig. 1b) or distributed throughout the nucleus and cytoplasm (Fig. 1c). AR nuclear localization reflects its activation as a transcription factor [16], and others have demonstrated nuclear localization upon treatment of breast cancer cells with androgen ligands [15, 19]. We next assigned AR nuclear and cytoplasmic scores for each tumor and then combined these data to generate an AR localization score, for each mammary tumor according to a previously published method [30]. According to this method, a higher score indicates nuclear localized AR, while a lower score reflects cytoplasmic AR. Tumor AR scores were then sub-categorized based on whether the tumor was regressing or progressing after OVX and determined by change in tumor size (Fig. 1d) and also on the adiposity status of the host (lean versus obese). In lean rats, AR was distributed throughout the nucleus and cytoplasm in all tumors (Supplemental Fig. 1A), and no differences in AR localization scores between regressing and progressing tumors were found (Fig. 1e). Conversely, in obese rats, there was a slight, but non-significant increase in the AR nuclear score, and a significant decrease in the AR cytoplasmic score (Supplemental Fig. 1B). Further, the overall AR localization score was significantly higher in progressing compared to regressing tumors (Fig. 1e). ER was not different between regressing and progressing tumors from obese rats (Fig. 1f), but PR was significantly elevated in progressing compared to regressing tumors (Fig. 1g). As would be expected, Ki67 tended to be higher in progressing than in regressing tumors from the obese rats (Fig. 1h). Overall, AR was predominantly localized to the nucleus rather than the cytoplasm in progressing tumors from obese rats compared to those that were regressing, or compared to any tumor from lean rats, suggesting that AR activation preferentially supports obesity-associated tumor growth after OVX in this model.
Fig. 1.
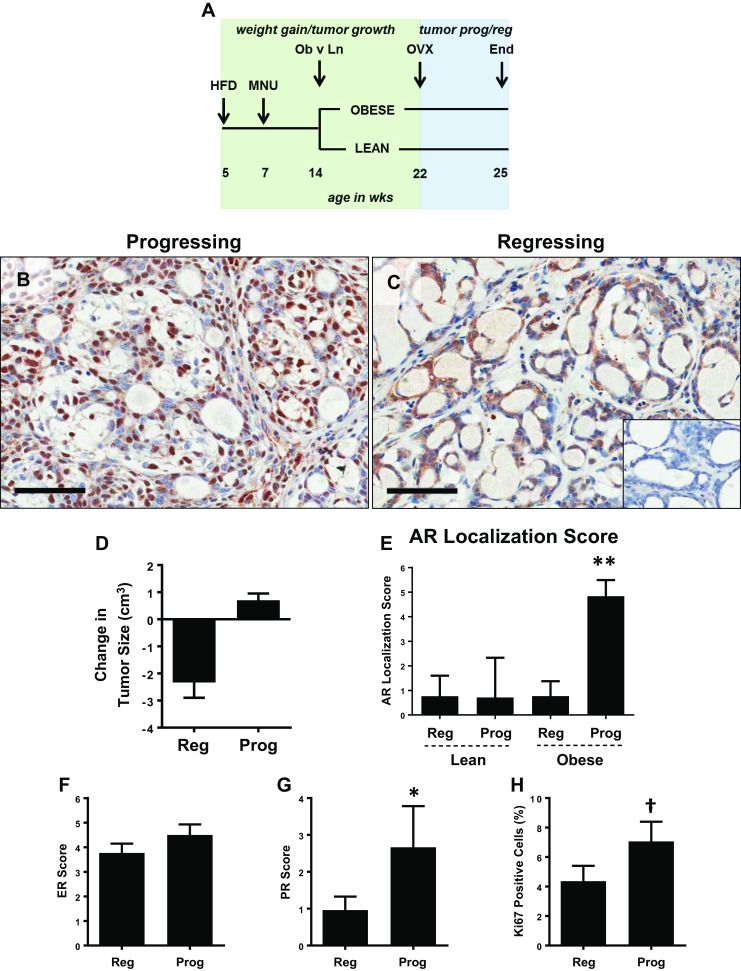
AR is localized to the nucleus in progressing tumors from obese rats. a Schematic study design. HFD high fat diet, MNU N-methyl-N-nitrosourea, OVX ovariectomy. b–c Representative images of AR immunostaining showing b nuclear and c cytoplasmic localization. Scale bars = 60 μm. Inset in c shows an AR negative (0% cells positive) tumor. d Change in size of regressing (shrinking) and progressing (growing) tumors from OVX until the end of study (3 weeks post OVX) in lean and obese rats. Regressing were defined as those with a size change <0 cm3, and progressing were defined as those with a size change >0 cm3, measured from the time of OVX until the end of the study (3 weeks post OVX). e AR localization score, calculated as Nuclear AR Score – Cytoplasmic AR Score in regressing (reg) and progressing (prog) tumors from lean and obese rats. The maximum cytoplasmic or nuclear AR score is 12, so the AR localization score can range from −12 (strong AR intensity, exclusively cytoplasmic) to 12 (strong AR intensity, exclusively nuclear). T test determined statistical significance. N = 17 regressing and 7 progressing tumors from lean, and 26 regressing and 6 progressing tumors from obese rats. **p < 0.01 f–h IHC scores for ER (f), PR (g) based on the Allred system, and percent Ki67 positive cells (h) in tumors from obese rats, categorized as regressing (reg) or progressing (prog). Mann-Whitney test determined statistical significance. N = 26 regressing and 6 progressing tumors. *p < 0.05; †p < 0.1
Circulating and Adipose Tissue Hormone Measurements in Post-OVX Rats
Differences in AR nuclear localization between lean and obese post-OVX rats could be due to differences in plasma or adipose tissue levels of steroid hormone receptor ligands. To address this question, we measured circulating levels of testosterone, as well as mammary adipose tissue testosterone and estradiol in lean and obese rats 3 weeks after OVX. We found no significant differences in plasma levels of testosterone (0.23 ± 0.02 versus 0.28 ± 0.04 ng/mL) between lean and obese rats, respectively. Further, in a previous study, we found no differences in plasma estradiol between lean and obese rats 3 weeks after OVX, and overall, the levels of estradiol in that study were at the lowest limit of detection for the assay [21]. Analysis of hormone levels in mammary adipose tissue revealed that estradiol was below the limit of detection in both lean and obese OVX rats (Fig. 2a); whereas, testosterone was present but was not different between lean and obese rats (Fig. 2b). As a positive estrogen control, we included human breast adipose tissue samples from post-menopausal women. In all but two human adipose tissue samples, estradiol was readily detectable by LC-mass spectrometry (LC-MS/MS; Fig. 2a); however, only two human breast tissue samples had detectable levels of testosterone (Fig. 2b). The conversion of androgens to estrogens in breast tissue of postmenopausal women results from expression and activity of the aromatase enzyme [34, 35]. Therefore, we evaluated expression of the aromatase gene, Cyp19a1, in rat mammary adipose and tumor tissue, using rat ovary tissue as a positive control. Surprisingly, we were unable to detect Cyp19a1 in either mammary or tumor tissue from lean or obese rats (Fig. 2c). Importantly, the testosterone levels in lean and obese rat mammary adipose tissue do not explain the difference in the observed localization of AR described in Fig. 1. Furthermore, our data suggest that testosterone rather than estrogen could be driving tumor progression in this model.
Fig. 2.
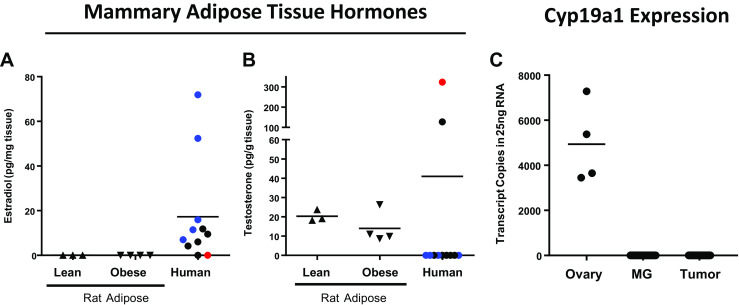
Adipose tissue hormone measurements at 3 weeks post OVX. LC-mass spectrometry was used to measure a estradiol and b testosterone in mammary/breast adipose tissue from postmenopausal women (N = 3 women, 1–5 pieces of tissue each) or from lean or obese rats taken 3 weeks post OVX. For human samples, circles of the same color are from the same patient. c Q-PCR analysis of Cyp19a1 (aromatase) in rat ovary (N = 4), rat mammary adipose (MG; N = 29), and rat mammary tumor (N = 37). Bars represent means
Inhibiting AR Prevents Obesity-Associated Tumor Progression After OVX
To determine the role of AR activation in post-OVX tumor growth, we utilized the anti-androgen, enzalutamide [23, 24]. Enzalutamide is now FDA approved for prostate cancer [36] and has efficacy against triple-negative and ER-positive breast cancers in preclinical studies [15, 19]. To test the effect of enzalutamide on the post-OVX growth of mammary tumors, we used the same general experimental design that we have used previously and focused on the impact of enzalutamide on obese rats (Fig. 3a and [24]), since we observed differences in AR localization between regressing and progressing tumors in this group (Fig. 1e). OVX was performed when mammary tumor burden reached ≥1 cm3, at approximately 22 weeks of age. Obese rats were randomized to control HFD or enzalutamide treatment for 8 weeks, at which time the study was terminated (Fig. 3a).
Fig. 3.
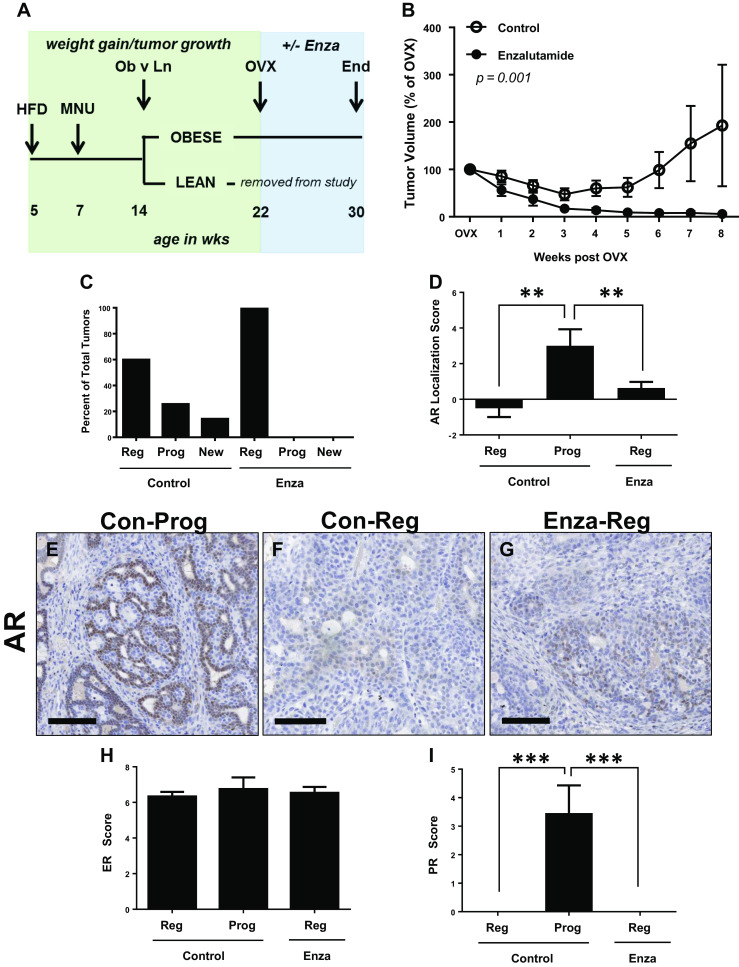
Enzalutamide treatment inhibits tumor progression in obese rats after OVX. a Schematic study design. HFD high fat diet, MNU N-methyl-N-nitrosourea, OVX ovariectomy. Lean rats were used for another study. Enzalutamide was administered for 8 weeks beginning at OVX, indicated by the blue box. b Tumor volume from control (empty circle) and enzalutamide-treated (filled circle) rats, represented as percent of that at OVX; p = 0.001 by ANOVA. c Percent of total tumors from the end of study that were regressing (reg) or progressing (prog) from the time of OVX, or that were newly formed (new) after OVX in Control (N = 19) and enzalutamide-treated (N = 11) rats; p = 0.007 by chi-square analysis. d AR localization scores in regressing and progressing tumors (N = 4–11 tumors per group). Representative images of AR immunostaining in e progressing and f regressing tumors from control rats, and in a g regressing tumor from enzalutamide-treated rat (g). Scale bars = 100 μm. h ER scores, and i PR scores in regressing (reg) and progressing (prog) tumors from control and enzalutamide-treated rats (N = 11–12 tumors per group). Mann-Whitney tests determined statistical significance for steroid hormone receptor levels. **p < 0.01; ***p < 0.001
At the time of randomization, tumor burden per rat was identical in both the control and enzalutamide-treated groups (1.13 ± 0.24 versus 1.13 ± 0.24 cm3 for control and enzalutamide groups, respectively). Consistent with our previous study [24], most tumors began to regress shortly after OVX (Fig. 3b), demonstrating the estrogen dependence of mammary tumors in this model. Approximately 5 weeks after OVX, some of the tumors that existed at the time of OVX, but that regressed shortly after, began to re-grow in control rats and were classified as progressing (Fig. 3b, c; N = 19 rats). At the end of the study, five new tumors had formed during the post-OVX period in the control group (Fig. 3c). Overall, 25.7% (9/35) of tumors in the control rats were progressing, 60% (21/35) of tumors were regressing, and 14.3% (5/35) of all tumors were newly emerged after OVX (Fig. 3c). In contrast, 100% (20/20) of tumors from enzalutamide-treated rats (N = 11 rats) completely regressed by the end of the study, and no new tumors formed (chi-square test, p = 0.007; Fig. 3c).
To determine the relationship between AR cellular localization and tumor progression, we evaluated AR using IHC and the same scoring system described in Fig. 1. In control rats, the AR localization score was significantly higher in progressing versus regressing tumors (Fig. 3d), and this was driven by a higher nuclear AR score (Supp Fig. 2A), rather than a lower cytoplasmic score (Supp Fig. 2B). We found that enzalutamide treatment reduced overall AR levels and the localization score significantly, compared to progressing tumors from control rats (Fig. 3d, g). As we saw previously, AR was predominantly nuclear in progressing tumors (Fig. 3d–e), while regressing tumors contained weak AR that was localized to the cytoplasm (Fig. 3d, f). ER levels were similar in tumors from control and enzalutamide-treated rats, regardless of whether they were progressing or regressing (Fig. 3h). Consistent with Fig. 1, PR levels were highest in progressing versus regressing tumors from control rats, and were significantly reduced by enzalutamide treatment (Fig. 3i). Although the PR gene (Pgr) is a classical ER target, AR can interact with ER at the Pgr promoter [16]. Our data indicate that, in the absence of estradiol, AR may play a role in regulating PR levels in tumors from obese rats.
The effect of enzalutamide on breast cancer cell proliferation and survival has been reported to depend on the relative levels of ER and steroid hormone ligands (estrogens and androgens) [15, 16]; therefore, we evaluated cell proliferation and death in tumors from control and enzalutamide-treated rats. Surprisingly, the percentages of tumor Ki67 (Fig. 4a) and TUNEL (Fig. 4b) positive cells were equivalent between control and enzalutamide groups; however, histological examination revealed extensive and significantly increased necrosis (Fig. 4c) in enzalutamide-treated tumors compared to controls (Fig. 4d–g). These observations suggest that the tumor-inhibitory effects of enzalutamide in post-OVX obese rats is due to suppression of cell survival rather than proliferation. This is consistent with the observation that, in androgen-driven tumors, AR antagonism induced cell death, while it suppressed cell proliferation in estrogen-dependent tumors [16].
Fig. 4.
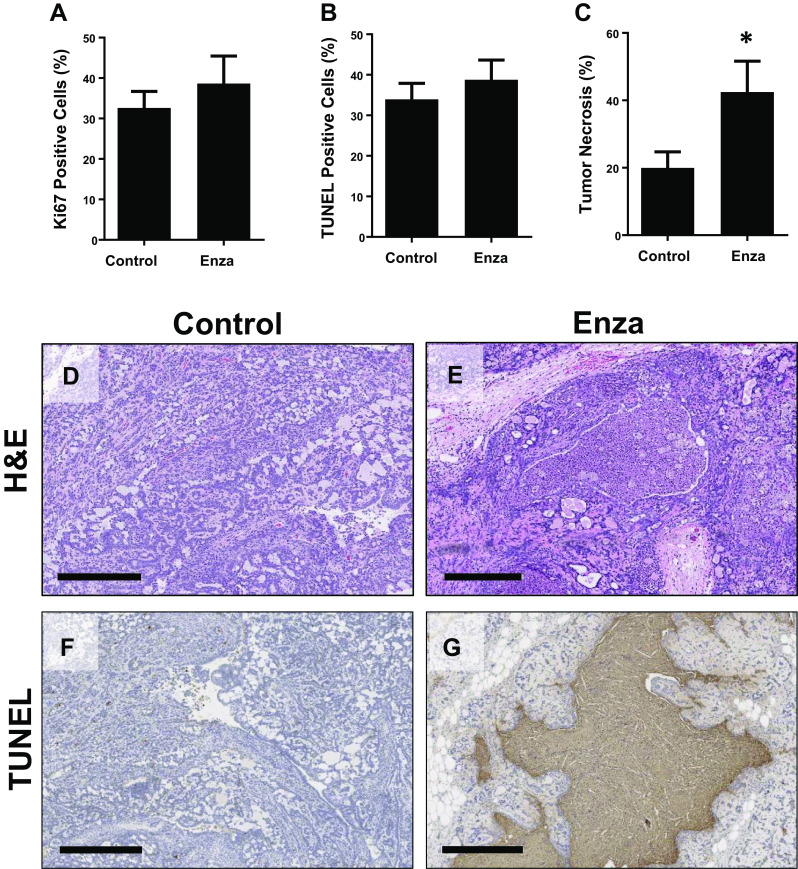
Enzalutamide inhibits tumor cell survival in obese rats. Percent of a Ki67 and b TUNEL positive cells in tumors from control and enzalutamide-treated rats. c Percent of tumor area that was necrotic in control and enzalutamide-treated rats. Representative H&E (d, e) and TUNEL (f, g) stained images of tumors from control (d , f) and enzalutamide-treated (e , g) rats. For TUNEL analysis, only the cells staining positive were quantified. The necrotic areas were not counted. Scale bars for d and e are 300 μm, and for f and g are 500 μm. T tests determined statistical significance. N = 11–25 tumors per group. *p < 0.05
Plasma and Morphometric Characteristics of Enzalutamide-Treated Rats
Steroid hormone ablation for the treatment of both prostate and breast cancer leads to metabolic dysfunction and negative sequela for many cancer patients [37–40]. However, our observations showing enhanced-mammary tumor AR signaling under an obesogenic environment, combined with high efficacy of enzalutamide in suppressing these mammary tumors, suggest a potential interaction between whole body metabolism and enzalutamide efficacy in postmenopausal breast cancer. Our previous study implicated peripheral insulin resistance as one factor contributing to increased tumor growth in obese, OVX rats [21]. To determine whether enzalutamide treatment affected metabolism in the host environment, we evaluated body composition and plasma levels of metabolic hormones in control and enzalutamide-treated rats. At the time of OVX, body weights were similar in both groups (Table 1). At the study end, enzalutamide-treated rats showed no evidence of toxicity, gaining equivalent weight after OVX as control rats (Supp Fig. 3A). We found that enzalutamide treatment did not affect body fat or liver fat contents (Table 1). In addition, differences were not found in gonadal, mesenteric, or retroperitoneal fat pad weights between control and enzalutamide-treated rats at the end of the study (Table 1). Plasma levels of non-esterified fatty acids (NEFA), insulin, leptin, and triglycerides (TAG) also were not different between control and enzalutamide-treated rats (Table 2); however, enzalutamide treatment was associated with modestly but significantly higher plasma cholesterol and fasting glucose (Table 2). Overall, AR antagonism by enzalutamide did not appear to adversely affect host metabolic function and was not toxic over the 8-week treatment period.
Table 1.
Morphometric characteristics of control and enzalutamide-treated rats
| Characteristic | Control | Enzalutamide | P value | ||||
|---|---|---|---|---|---|---|---|
| Mean | SEM | N | Mean | SEM | N | ||
| Body weight at OVX (g) | 319 | 13.9 | 22 | 312 | 5.9 | 11 | not significant |
| Body weight at study end (g) | 404 | 20.6 | 19 | 403 | 6.1 | 10 | not significant |
| Body fat at study end (%) | 34 | 1.6 | 19 | 33.4 | 1.3 | 11 | not significant |
| Liver fat (%) | 13.2 | 2.1 | 19 | 12.2 | 0.7 | 11 | not significant |
| Gonadal fat mass (g) | 15.8 | 1.2 | 19 | 16.3 | 1.2 | 11 | not significant |
| Mesenteric fat mass (g) | 8.6 | 0.9 | 19 | 6.9 | 0.5 | 11 | not significant |
| Retroperitoneal fat mass (g) | 6.7 | 0.5 | 19 | 6.4 | 0.5 | 11 | not significant |
Table 2.
Serum characteristics of control and enzalutamide-treated rats
| Serum component | Control | Enzalutamide | P value | ||||
|---|---|---|---|---|---|---|---|
| Mean | SEM | N | Mean | SEM | N | ||
| NEFA (uM) | 705 | 36.4 | 19 | 743 | 50.8 | 11 | not significant |
| TAG (mg/dL) | 46.5 | 2.9 | 19 | 55.8 | 3.8 | 11 | not significant |
| Cholesterol (mg/dL) | 80.8 | 5.8 | 19 | 99.8 | 4.6 | 11 | 0.02 |
| Glucose (mM) | 11.3 | 0.4 | 19 | 13.5 | 0.4 | 11 | 0.002 |
| Insulin (ng/mL) | 0.58 | 0.08 | 19 | 0.49 | 0.08 | 10 | not significant |
| Leptin (ng/mL) | 5.6 | 0.9 | 19 | 5.6 | 0.6 | 11 | not significant |
Because we observed differences in circulating lipids and glucose between control and enzalutamide-treated rats, we assessed the profile of adipocyte sizes, which imparts insulin sensitivity, metabolic function, and also associates with chronic inflammation. Large, hypertrophic adipocytes are frequently insulin resistant and display elevated lipolysis, while small adipocytes are more insulin sensitive and readily take up glucose and circulating lipids [37]. Moreover, a profile of hypertrophic adipocytes is often associated with the presence of crown-like structures, which are dying cells engulfed by macrophages [41]. We measured adipocyte cellularity in retroperitoneal (abdominal) and mammary (subcutaneous) fat pads from obese-control rats and from obese rats treated with enzalutamide and compared these profiles to those from lean rats. As expected, the median adipocyte size was lower in lean versus obese rats in both the subcutaneous and abdominal depots (Supp Fig. 3B,C). In the abdominal fat pad, obese rats had a greater prevalence of very small adipocytes (20 μm diameter), but they also had more large adipocytes (>100 μm diameter; Supp Fig. 3B) than lean rats. The peak in small adipocytes seen in this depot could reflect an expanded preadipocyte pool in response to signals from hypertrophic, lipid-engorged adipocytes. Enzalutamide treatment had effects on adipocyte size in both depots, with the greatest effect in the abdominal fat pad. Specifically, enzalutamide treatment was associated with a decrease in the median adipocyte size in abdominal (Supp Fig. 3B) and subcutaneous (Supp Fig. 3C) fat pads, such that there was a greater prevalence of smaller adipocytes in enzalutamide-treated compared to obese-control rats. The difference in adipose depot cell sizes between control and enzalutamide-treated rats suggests that AR plays a role in adipocyte lipid storage and synthesis after the loss of ovarian estrogens. Nonetheless, after 8 weeks of treatment enzalutamide does not appear to strongly influence body composition.
IL-6 Sensitizes Breast Cancer Cells to Testosterone
Differences in circulating or adipose-derived steroid hormones did not appear to explain the difference in AR localization between lean and obese rats; however, obesity is associated with chronic low-grade inflammation [41]. The pro-inflammatory cytokine, IL-6, may drive prostate cancer progression and can activate AR in prostate cancer cells [42, 43]. We have previously reported that our obese rats have elevated circulating levels of IL-6 compared to lean rats [24]. Therefore, we postulated that IL-6 could play a role in the differential AR localization phenotype that we observed between tumors from lean and obese rats, potentially by enhancing the sensitivity of tumor cells to AR ligands. To examine this, we treated the ER/AR-positive MCF-7 human breast cancer cell line with increasing doses of testosterone in the presence and absence of IL-6, and the AR target gene, Fkbp5, was analyzed as a read-out of AR activation. By itself, IL-6 did not significantly increase Fkbp5 expression. However, in the presence of IL-6, MCF-7 cells were more sensitive to testosterone than those without IL-6, showing significant induction of Fkbp5 at 1 × 10−12 M (0.001 nM) testosterone (Fig. 5a). Similar effects of IL-6 were seen on AR targets Tmprss2 (Supp Fig 4A) and Klk3 (Supp Fig 4B); however, the sensitization to testosterone was not as great as with Fkbp5. In ER-positive MCF7 cells, the effect of testosterone could be blocked by the ER-downregulator, fulvestrant (ICI 182780), and by enzalutamide (Fig. 5b), suggesting that both ER and AR may be involved in the response to testosterone. We also found that IL-6 altered the sensitivity to testosterone in AR-positive/ER-negative MDA-453 cells, potentiating induction of Fkbp5 (Supp Fig 4C) and Tmprss2 (Supp Fig 4D), which could be blocked by enzalutamide (Supp Fig. 4E). Importantly, we did not observe any effects of IL-6, testosterone, or inhibitor treatment on expression of the control gene Polr2a (Fig. 5c–d; Supp Fig. 4E). To determine whether this relationship was found in primary human breast tumors, we evaluated AR and Fkbp5 in gene expression profiling data of ER-positive breast cancers with associated patient IL-6 measurements [33]. Expression of both AR and Fkbp5 were higher in primary ER-positive breast tumors from patients with high compared to low circulating IL-6 levels (Fig. 5e–f).
Fig. 5.
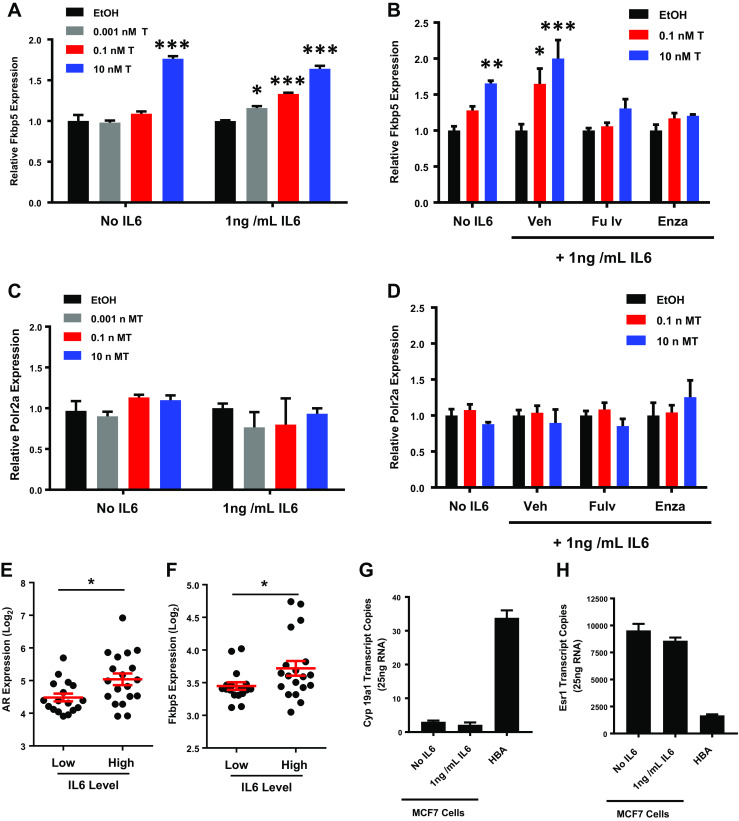
IL-6 sensitizes breast cancer cells to low levels of testosterone. a QPCR analysis of Fkbp5 expression in MCF7 cells treated with varying doses of testosterone in the presence or absence of IL-6. b QPCR analysis of Fkbp5 expression in MCF7 cells treated with low (0.1 nM) or high (10 nM) testosterone in the presence or absence of IL-6, and co-treated either with fulvestrant (Fulv; 100 nM) or enzalutamide (Enza; 10 μM). c–d Expression of Polr2a in cells treated with or without IL-6 and varying doses of testosterone (c), or in cells treated with IL-6, testosterone, or inhibitors (d) included as a control. e–f Expression of AR (e) and Fkbp5 (f) in primary breast tumor samples from women with high or low IL-6, as profiled in GSE54430. (g–h) Expression of Cyp19a1 (g) and Esr1 (h) in MCF7 cells treated with or without IL-6, and in human breast adipose tissue (HBA). T tests determined statistical significance. *p < 0.05; **p < 0.01; ***p < 0.001
The requirement of ER for the induction of Fkbp5 in MCF7 cells could suggest that the testosterone was aromatized to estradiol. Although cultured breast cancer cells are generally assumed not to synthesize estradiol, some studies have indicated that Cyp19a1 is expressed in some ER-positive human breast cancer cells (reviewed in [35]). We performed qPCR for the Cyp19a1 gene in our MCF7 cells, using human breast adipose as a positive control (Fig. 5g) and found that Cyp19a1 was at the lowest limit of detection and was not induced by IL-6 treatment. In contrast, Esr1, which encodes ERα, was highly expressed in these cells and was also not affected by IL-6 treatment (Fig. 5h). Altogether, these data suggest that factors unique to the host environment, such as IL-6, change how breast cancer cells respond to steroid hormones rather than impacting the levels of steroid receptor expression or ligands. Moreover, the observation that fulvestrant and enzalutamide can block testosterone-mediated AR activation indicates that, when both are present, ER and AR transcriptional activity may be elevated even in the absence of estradiol, and has implications for treating breast cancers resistant to aromatase inhibitors.
Discussion
We have established a rat model that allows us to investigate the effects of obesity upon mammary tumor growth in the post-menopausal setting through the use of OVX to remove ovarian production of estrogens. Using this model, we previously identified a critical window following OVX when obesity promotes the progression of existing tumors and also the development of new tumors [23, 24]. Here, we explored a role for AR in mammary tumor progression during this window, specifically in obese rats. Our data demonstrate that nuclear localization of AR, which indicates transcriptional activation, is increased in mammary tumors that progress in obese compared to lean rats shortly after estrogen loss (Fig. 1), and that AR nuclear localization and expression are maintained in progressing tumors up to 8 weeks after OVX (Fig. 3). The variability in AR localization across tumors between the lean and obese rats cannot be explained simply by differences in circulating or mammary adipose tissue testosterone between groups (Fig. 2), suggesting that other factors may contribute to activation of AR in tumors specifically in the obese. Regardless, treatment with the anti-androgen enzalutamide completely inhibited tumor progression by increasing tumor necrosis and prevented the formation of new tumors in obese rats (Fig. 3), which demonstrates a tumor-promotional role for AR in this context. Finally, we observed that IL-6, which we have previously demonstrated to be increased in the plasma of obese rats [24], potentiated the ability of low levels of testosterone to induce transcriptional activation of AR (Fig. 5). Together, our data suggest that obesity is a critical biological variable that modifies steroid hormone receptor action in mammary tumors and may support AR activation during mammary tumorigenesis and progression.
The role of AR continues to be investigated in breast cancer, and studies have suggested that AR can either be a positive or a negative prognostic factor [12, 17, 30, 44–46]. Recent reports show that AR drives breast cancer progression after treatment with either aromatase inhibitors or tamoxifen [9–11]. Higher levels of AR than ER were found in the primary tumors of patients that experienced disease recurrence on adjuvant tamoxifen [15] and were higher in tamoxifen-resistant metastatic breast cancers compared to primary tumors, as well as in a model of tamoxifen-resistant MCF7 breast cancer cells [9]. DHT stimulated proliferation in cell line models of aromatase inhibitor-resistant breast cancer [10], and AR overexpression was shown to confer aromatase-inhibitor resistance in breast cancer cells overexpressing Cyp19a1 [11]. These and other studies suggest that, in the absence of ER, or after the selective inhibition of ER, tumors can become dependent on AR [10, 15, 16]. Indeed, in AR-positive TNBC, AR has been shown to bind to the same regions of chromatin as ER in ER-positive breast cancer, indicating that AR can functionally substitute for ER [47].
In our study, we found predominant nuclear localization of AR in progressing mammary tumors from obese animals, which suggests that AR is a poor prognostic marker in the context of obesity, even though levels of testosterone are not elevated compared to levels in lean animals. These observations are consistent with a recently published study, which reported the percent AR positive nuclei, as is conventionally done for steroid hormone receptors, and also the relative levels of nuclear to cytoplasmic AR protein [30]. The authors found that the overall levels of AR had significant prognostic value only in TNBC, which has been shown [19]. Interestingly, the score that reflected AR nuclear localization, which they termed the AR-D value, significantly predicted a poor outcome for both TNBC and luminal (ER-positive) breast cancers [30]. Since AR translocates from the cytoplasm to the nucleus upon ligand binding and activation [16], this published study and our data highlight the importance of considering the AR activation and localization states when assessing its prognostic significance.
To determine whether increased testosterone in obese rats could explain the difference in AR localization, we measured this steroid hormone in circulation and in local mammary adipose tissue (Fig. 2). It is known that obese women have elevated levels of circulating estrogens and androgens compared to lean women after menopause [48, 49]; however, we did not find differences in testosterone in obese versus lean rats (Fig. 2). One limitation of this study is that we did not measure DHT in circulation or in adipose tissue. While DHT is a more potent AR ligand than testosterone, the ability to detect this hormone by mass spectrometry is poor; therefore, we cannot rule out the possibility that DHT was elevated in obese compared to lean rats, which could account for differences observed in AR localization. Although we did not confirm an increase in testosterone levels in obese rats (Fig. 2), we found that IL-6 sensitized human ER-positive and -negative breast cancer cells to low levels of testosterone (Fig. 5, Supp Fig. 4). IL-6 was shown to regulate AR activity in prostate cancer cells by inducing steroidogenic enzymes, including Cyp11a1, Akr1c3, and Hsd3b2 that all regulate de novo testosterone biosynthesis from cholesterol, and increasing intracellular testosterone production [50]. In addition, IL-6 has been shown to increase androgen-stimulated AR activity in prostate cancer cells [43]. While we did not observe a ligand-independent effect of IL-6 on AR-target gene expression, crosstalk between IL-6 and androgen signaling may play a role in breast cancer. The sensitization of breast cancer cells to testosterone in the presence of IL-6 demonstrates the importance of the obese environment in facilitating hormone-dependent breast cancer progression, potentially by changing the way tumor cells respond to available hormone ligands. Our studies employed a rat model of obesity that faithfully recapitulates many aspects of human breast cancer and its risk factors [22, 24]. In order to gain a more complete understanding about how obesity influences breast cancer progression, it will be important to study the variety of steroid receptor-positive breast tumors being made available as patient-derived xenograft specimens in transplant competent models of obesity. Clearly, obesity is a biological variable that merits consideration with regard to therapeutic approaches used to treat breast cancer.
To test the role of AR in obesity-associated post-OVX tumor progression, we utilized enzalutamide, which inhibits AR nuclear translocation [15]. Enzalutamide effectively prevented the progression of tumors that existed at OVX and also prevented the formation of new tumors after OVX in obese rats, highlighting a critical tumor-promotional role for AR during the post-OVX period (Fig. 3). In general, enzalutamide treatment was not toxic and did not promote changes in body weight or body fat in obese rats after 8 weeks of treatment (Supp Fig. 3, Table 1). We found a modest, but significant, increase in fasting plasma glucose and cholesterol and also a decrease in the median adipocyte diameter with enzalutamide treatment, particularly in the abdominal depot (Table 2, Supp Fig. 3). Both AR and ER play important roles in glucose and lipid metabolism in men, and in pre- and postmenopausal women [51, 52]. Increases in plasma insulin and cholesterol have been reported in men taking enzalutamide [53]; however, no studies have reported changes in these metabolic parameters in females. In general, our data indicate that AR may play a role in metabolic homeostasis in females after the loss of ovarian function, but that short-term enzalutamide treatment does not dramatically alter metabolic function.
Recent studies have shown that AR and ER interact at target gene response elements and that disrupting AR with enzalutamide can prevent ER DNA binding [16]. Since the tumors from obese rats maintained ER after OVX (Figs. 1 and 3), we wished to determine whether ER activation played a role in AR nuclear localization. PR expression was elevated in tumors with nuclear-localized AR in short-term (Fig. 1) and long-term (Fig. 3) post-OVX studies, and enzalutamide treatment reduced PR levels, concomitant with tumor regression, without affecting ER levels (Fig. 3). PR is a well-known target of ligand-activated ER; however, we could not detect estradiol in mammary adipose tissue from either lean or obese rats (Fig. 2). This suggests that ligand-activated ER may not be solely driving the expression of PR. Indeed, a recent study showed that ER and AR act in concert to bind to the Pgr and other classical ER target gene promoters [16]. We found that the effects of IL-6 on ER-positive breast cancer cell sensitivity to testosterone could be blocked by fulvestrant or enzalutamide (Fig. 5), but that IL-6 also modified the response of ER-negative cells to testosterone, indicating that the effect is primarily through AR. Thus, while our data indicate that estradiol and ER may not be required for tumor progression in the obese rats after OVX, there may still be a role for ER in this process. In total, these observations suggest that our rat model of obese, postmenopausal breast cancer may be characterized by an increased dependence on AR, as is sometimes observed after endocrine therapy [10, 54].
Breast adipose tissue in women serves as a primary source of estradiol after menopause due to expression of Cyp19a1, which encodes the aromatase enzyme [55]. Using LC-mass spectrometry (LC-MS/MS), we were able to measure estradiol in the majority of breast adipose samples from postmenopausal women (Fig. 2), but not in mammary adipose tissue from lean or obese rats (Fig. 2). In addition, we could not detect Cyp19a1 in rat mammary adipose or tumor tissue (Fig. 2), suggesting that testosterone may not be converted into estradiol in this model. While LC-MS/MS is superior in sensitivity and specificity to ELISA, one caveat is that the lower limit of detection (~2 pM for estradiol) is above the amount sufficient to activate ER in breast cancer cells [56]. Thus, it is possible that estradiol is produced in the rat adipose tissue at levels below the limit of detection, but that, it is still functional due to coactivation of other signaling pathways. Testosterone was higher in rodent adipose tissue than it was in all but two human breast adipose tissue samples (Fig. 2). While the hormonal environment of a postmenopausal woman is very different from that of the post-OVX rat, these data indicate that the OVX female rat may serve as a valuable model of aromatase inhibition and could be used to identify novel regulators of acquired endocrine therapy resistance. In women, long term (>6 mos; [57–59]), but not short-term (<4 weeks; [60, 61]) aromatase inhibitor treatment can result in accumulation of androgens, which may support progression of aromatase inhibitor-resistant breast cancer through AR activation [10, 11, 54].
Concluding Remarks
Recent studies have demonstrated a role for AR in driving breast cancer growth and survival, particularly after endocrine therapy, and current clinical trials are focused on targeting AR in multiple breast tumor subtypes. Our study indicates that AR may translocate to the nucleus of tumor cells shortly after estrogen loss as an early indicator of tumor progression in a preclinical rat model of obesity and breast cancer. We also demonstrate a role for IL-6 in breast cancer cell AR activation, highlighting the importance of the host environment in dictating how mammary tumor cells respond to steroid hormones. Our data suggest that it may be important to assess AR activation as well as overall level in mammary tumors, and to consider that the host environment, either local or systemic, can influence how tumors respond to steroid hormones and endocrine therapies. Finally, this study may have implications in defining the population of women with ER-positive breast cancer most likely to benefit from anti-AR therapy.
Electronic supplementary material
(PDF 182 kb)
Acknowledgements
Enzalutamide was generously provided by Medivation, Inc./Astellas Pharmaceuticals. We thank Dr. Matthew Sikora for insightful discussions and critical review of the manuscript. We also thank Patricia Webb, Julie Houck, and Veronica Wessells for excellent technical assistance, and the Research Histology Shared Resource, the Protein Production, Monoclonal Antibody and Tissue Culture Shared Resource, and Biorepository Core, all funded in part by Cancer Center grant P30CA046934.
Compliance with Ethical Standards
Funding Sources
This work was supported by the Cancer League of Colorado (EAW), the University of Colorado Nutrition and Obesity Research Center Pilot Awards P30-DK048520 (EAW, EDG, PSM), NIH/NCI CA164166 (PSM, PS) and CA169430 (EDG), and the Komen Foundation KG081323 (SMA). EAW and EDG were supported by seed grants from the University of Colorado’s Center for Women’s Health Research. We also appreciate the generous support from both the Colorado Obesity Research Institute, and the Energy Balance and Metabolic Core Laboratories within the Colorado Nutrition Obesity Research Center.
Conflict of Interest
The authors declare that they have no conflict of interest.
Footnotes
Paul S. MacLean and Steven M. Anderson Co-senior authors
Contributor Information
Elizabeth A Wellberg, Phone: 303-724-3472, Email: Elizabeth.Wellberg@ucdenver.edu.
L Allyson Checkley, Email: Laura.Checkley@CoramHC.com.
Erin D Giles, Email: egiles@tamu.edu.
Stevi J Johnson, Email: Stevi.Johnson@ucdenver.edu.
Robera Oljira, Email: Robera.Oljira@ucdenver.edu.
Reema Wahdan-Alaswad, Email: Reema.Wahdan-Alaswad@ucdenver.edu.
Rebecca M Foright, Email: Rebecca.Foright@ucdenver.edu.
Greg Dooley, Email: Gregory.Dooley@colostate.edu.
Susan M. Edgerton, Email: Susan.Edgerton@ucdenver.edu
Sonali Jindal, Email: jindal@ohsu.edu.
Ginger C Johnson, Email: Ginger.Johnson@ucdenver.edu.
Jennifer K Richer, Email: Jennifer.Richer@ucdenver.edu.
Peter Kabos, Email: Peter.Kabos@ucdenver.edu.
Ann D Thor, Email: Ann.Thor@ucdenver.edu.
Pepper Schedin, Email: schedin@ohsu.edu.
Paul S MacLean, Phone: 303-724-3030, Email: Paul.MacLean@ucdenver.edu.
Steven M Anderson, Phone: 303-724-3742, Email: Steve.Anderson@ucdenver.edu.
References
- 1.Flegal KM, Kruszon-Moran D, Carroll MD, Fryar CD, Ogden CL. Trends in obesity among adults in the United States, 2005 to 2014. JAMA. 2016;315(21):2284–2291. doi: 10.1001/jama.2016.6458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Chan DS, Vieira AR, Aune D, Bandera EV, Greenwood DC, McTiernan A, Navarro Rosenblatt D, Thune I, Vieira R, Norat T. Body mass index and survival in women with breast cancer-systematic literature review and meta-analysis of 82 follow-up studies. Annals of oncology : official journal of the European Society for Medical Oncology / ESMO. 2014;25(10):1901–1914. doi: 10.1093/annonc/mdu042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Munsell MF, Sprague BL, Berry DA, Chisholm G, Trentham-Dietz A. Body mass index and breast cancer risk according to postmenopausal estrogen-progestin use and hormone receptor status. Epidemiol Rev. 2014;36:114–136. doi: 10.1093/epirev/mxt010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Matthews SB, Thompson HJ (2016) The obesity-breast cancer conundrum: an analysis of the issues. Int J Mol Sci 17(6) [DOI] [PMC free article] [PubMed]
- 5.Neuhouser ML, Aragaki AK, Prentice RL, Manson JE, Chlebowski R, Carty CL, Ochs-Balcom HM, Thomson CA, Caan BJ, Tinker LF, Urrutia RP, Knudtson J, Anderson GL. Overweight, obesity, and postmenopausal invasive breast cancer risk: a secondary analysis of the Women’s Health Initiative randomized clinical trials. JAMA Oncol. 2015;1(5):611–621. doi: 10.1001/jamaoncol.2015.1546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Jiralerspong S, Kim ES, Dong W, Feng L, Hortobagyi GN, Giordano SH (2013) Obesity, Diabetes, and Survival Outcomes in a Large Cohort of Early-Stage Breast Cancer Patients. Annals of oncology: official journal of the European Society for Medical Oncology/ESMO [DOI] [PMC free article] [PubMed]
- 7.Ioannides SJ, Barlow PL, Elwood JM, Porter D. Effect of obesity on aromatase inhibitor efficacy in postmenopausal, hormone receptor-positive breast cancer: a systematic review. Breast Cancer Res Treat. 2014;147(2):237–248. doi: 10.1007/s10549-014-3091-7. [DOI] [PubMed] [Google Scholar]
- 8.ACS (2015-2016) Breast Cancer Facts & Figures
- 9.De Amicis F, Thirugnansampanthan J, Cui Y, Selever J, Beyer A, Parra I, Weigel NL, Herynk MH, Tsimelzon A, Lewis MT, Chamness GC, Hilsenbeck SG, Ando S, Fuqua SA. Androgen receptor overexpression induces tamoxifen resistance in human breast cancer cells. Breast Cancer Res Treat. 2010;121(1):1–11. doi: 10.1007/s10549-009-0436-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fujii R, Hanamura T, Suzuki T, Gohno T, Shibahara Y, Niwa T, Yamaguchi Y, Ohnuki K, Kakugawa Y, Hirakawa H, Ishida T, Sasano H, Ohuchi N, Hayashi SI (2014) Increased Androgen Receptor Activity and Cell Proliferation in Aromatase Inhibitor-Resistant Breast Carcinoma. J Steroid Biochem Mol Biol [DOI] [PubMed]
- 11.Rechoum Y, Rovito D, Iacopetta D, Barone I, Ando S, Weigel NL, O'Malley BW, Brown PH, Fuqua SA (2014) Ar Collaborates with Eralpha in Aromatase Inhibitor-Resistant Breast Cancer. Breast Cancer Res Treat [DOI] [PMC free article] [PubMed]
- 12.Collins LC, Cole KS, Marotti JD, Hu R, Schnitt SJ, Tamimi RM. Androgen receptor expression in breast cancer in relation to molecular phenotype: results from the Nurses’ Health Study. Mod Pathol. 2011;24(7):924–931. doi: 10.1038/modpathol.2011.54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ogawa Y, Hai E, Matsumoto K, Ikeda K, Tokunaga S, Nagahara H, Sakurai K, Inoue T, Nishiguchi Y. Androgen receptor expression in breast cancer: relationship with clinicopathological factors and biomarkers. Int J Clin Oncol. 2008;13(5):431–435. doi: 10.1007/s10147-008-0770-6. [DOI] [PubMed] [Google Scholar]
- 14.Peters AA, Buchanan G, Ricciardelli C, Bianco-Miotto T, Centenera MM, Harris JM, Jindal S, Segara D, Jia L, Moore NL, Henshall SM, Birrell SN, Coetzee GA, Sutherland RL, Butler LM, Tilley WD. Androgen receptor inhibits estrogen receptor-alpha activity and is prognostic in breast cancer. Cancer Res. 2009;69(15):6131–6140. doi: 10.1158/0008-5472.CAN-09-0452. [DOI] [PubMed] [Google Scholar]
- 15.Cochrane DR, Bernales S, Jacobsen BM, Cittelly DM, Howe EN, D'Amato NC, Spoelstra NS, Edgerton SM, Jean A, Guerrero J, Gomez F, Medicherla S, Alfaro IE, McCullagh E, Jedlicka P, Torkko KC, Thor AD, Elias AD, Protter AA, Richer JK. Role of the androgen receptor in breast cancer and preclinical analysis of enzalutamide. Breast Cancer Res. 2014;16(1):R7. doi: 10.1186/bcr3599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.D'Amato NC, Gordon MA, Babbs B, Spoelstra NS, Carson Butterfield KT, Torkko KC, Phan VT, Barton VN, Rogers TJ, Sartorius CA, Elias A, Gertz J, Jacobsen BM, Richer JK (2016) Cooperative Dynamics of Ar and Er Activity in Breast Cancer. Mol Cancer Res [DOI] [PMC free article] [PubMed]
- 17.Barton VN, D'Amato NC, Gordon MA, Christenson JL, Elias A, Richer JK. Androgen receptor biology in triple negative breast cancer: a case for classification as Ar+ or quadruple negative disease. Horm Cancer. 2015;6(5–6):206–213. doi: 10.1007/s12672-015-0232-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lehmann BD, Jovanovic B, Chen X, Estrada MV, Johnson KN, Shyr Y, Moses HL, Sanders ME, Pietenpol JA. Refinement of triple-negative breast cancer molecular subtypes: implications for neoadjuvant chemotherapy selection. PLoS One. 2016;11(6):e0157368. doi: 10.1371/journal.pone.0157368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Barton VN, D'Amato NC, Gordon MA, Lind HT, Spoelstra NS, Babbs BL, Heinz RE, Elias A, Jedlicka P, Jacobsen BM, Richer JK. Multiple molecular subtypes of triple-negative breast cancer critically rely on androgen receptor and respond to enzalutamide in vivo. Mol Cancer Ther. 2015;14(3):769–778. doi: 10.1158/1535-7163.MCT-14-0926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Caiazza F, Murray A, Madden SF, Synnott NC, Ryan EJ, O'Donovan N, Crown J, Duffy MJ. Preclinical evaluation of the Ar inhibitor enzalutamide in triple-negative breast cancer cells. Endocr Relat Cancer. 2016;23(4):323–334. doi: 10.1530/ERC-16-0068. [DOI] [PubMed] [Google Scholar]
- 21.Barton VN, Christenson JL, Gordon MA, Greene LI, Rogers TJ, Butterfield K, Babbs B, Spoelstra NS, D'Amato NC, Elias A, Richer JK (2017) Androgen Receptor Supports an Anchorage-Independent, Cancer Stem Cell-Like Population in Triple-Negative Breast Cancer. Cancer Res [DOI] [PMC free article] [PubMed]
- 22.Giles ED, Jackman MR, MacLean PS. Modeling diet-induced obesity with obesity-prone rats: implications for studies in females. Front Nutr. 2016;3:50. doi: 10.3389/fnut.2016.00050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Maclean PS, Giles ED, Johnson GC, McDaniel SM, Fleming-Elder BK, Gilman KA, Andrianakos AG, Jackman MR, Shroyer KR, Schedin PJ (2009) A Surprising Link between the Energetics of Ovariectomy-Induced Weight Gain and Mammary Tumor Progression in Obese Rats. Obesity (Silver Spring) [DOI] [PMC free article] [PubMed]
- 24.Giles ED, Wellberg EA, Astling DP, Anderson SM, Thor AD, Jindal S, Tan AC, Schedin PS, Maclean PS. Obesity and overfeeding affecting both tumor and systemic metabolism activates the progesterone receptor to contribute to postmenopausal breast cancer. Cancer Res. 2012;72(24):6490–6501. doi: 10.1158/0008-5472.CAN-12-1653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.MacLean PS, Giles ED, Johnson GC, McDaniel SM, Fleming-Elder BK, Gilman KA, Andrianakos AG, Jackman MR, Shroyer KR, Schedin PJ. A surprising link between the energetics of ovariectomy-induced weight gain and mammary tumor progression in obese rats. Obesity (Silver Spring) 2010;18(4):696–703. doi: 10.1038/oby.2009.307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.MacLean PS, Higgins JA, Johnson GC, Fleming-Elder BK, Peters JC, Hill JO. Metabolic adjustments with the development, treatment, and recurrence of obesity in obesity-prone rats. Am J Physiol Regul Integr Comp Physiol. 2004;287(2):R288–R297. doi: 10.1152/ajpregu.00010.2004. [DOI] [PubMed] [Google Scholar]
- 27.Giles ED, Jackman MR, Johnson GC, Schedin PJ, Houser JL, MacLean PS. Effect of the estrous cycle and surgical ovariectomy on energy balance, fuel utilization, and physical activity in lean and obese female rats. Am J Physiol Regul Integr Comp Physiol. 2010;299(6):R1634–R1642. doi: 10.1152/ajpregu.00219.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Checkley LA, Rudolph MC, Wellberg EA, Giles ED, Wahdan-Alaswad RS, Houck JA, Edgerton S, Thor AD, Schedin P, Anderson S, MacLean P (2017) Metformin accumulation correlates with organic cation transporter 2 protein expression and predicts mammary tumor regression in vivo. Cancer Prev Res (Phila) [DOI] [PMC free article] [PubMed]
- 29.Remmele W, Hildebrand U, Hienz HA, Klein PJ, Vierbuchen M, Behnken LJ, Heicke B, Scheidt E. Comparative histological, histochemical, immunohistochemical and biochemical studies on oestrogen receptors, lectin receptors, and Barr bodies in human breast cancer. Virchows Arch A Pathol Anat Histopathol. 1986;409(2):127–147. doi: 10.1007/BF00708323. [DOI] [PubMed] [Google Scholar]
- 30.Feng J, Li L, Zhang N, Liu J, Zhang L, Gao H, Wang G, Li Y, Zhang Y, Li X, Liu D, Lu J, Huang B (2016) Androgen and Ar Contribute to Breast Cancer Development and Metastasis: An Insight of Mechanisms. Oncogene [DOI] [PubMed]
- 31.Rudolph MC, Wellberg EA, Lewis AS, Terrell KL, Merz AL, Maluf NK, Serkova NJ, Anderson SM. Thyroid hormone responsive protein Spot14 enhances catalysis of fatty acid synthase in lactating mammary epithelium. J Lipid Res. 2014;55(6):1052–1065. doi: 10.1194/jlr.M044487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Wellberg EA, Rudolph MC, Lewis AS, Padilla-Just N, Jedlicka P, Anderson SM. Modulation of tumor fatty acids, through overexpression or loss of thyroid hormone responsive protein spot 14 is associated with altered growth and metastasis. Breast Cancer Res. 2014;16(6):481. doi: 10.1186/s13058-014-0481-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Casneuf T, Axel AE, King P, Alvarez JD, Werbeck JL, Verhulst T, Verstraeten K, Hall BM, Sasser AK. Interleukin-6 is a potential therapeutic target in interleukin-6 dependent, estrogen receptor-alpha-positive breast cancer. Breast Cancer (Dove Med Press) 2016;8:13–27. doi: 10.2147/BCTT.S92414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.McTernan PG, Anwar A, Eggo MC, Barnett AH, Stewart PM, Kumar S. Gender differences in the regulation of P450 aromatase expression and activity in human adipose tissue. Int J Obes Relat Metab Disord. 2000;24(7):875–881. doi: 10.1038/sj.ijo.0801254. [DOI] [PubMed] [Google Scholar]
- 35.Miki Y, Suzuki T, Sasano H. Controversies of aromatase localization in human breast cancer—stromal versus parenchymal cells. J Steroid Biochem Mol Biol. 2007;106(1–5):97–101. doi: 10.1016/j.jsbmb.2007.05.007. [DOI] [PubMed] [Google Scholar]
- 36.Scher HI, Fizazi K, Saad F, Taplin ME, Sternberg CN, Miller K, de Wit R, Mulders P, Chi KN, Shore ND, Armstrong AJ, Flaig TW, Flechon A, Mainwaring P, Fleming M, Hainsworth JD, Hirmand M, Selby B, Seely L, de Bono JS, Investigators A. Increased survival with enzalutamide in prostate cancer after chemotherapy. N Engl J Med. 2012;367(13):1187–1197. doi: 10.1056/NEJMoa1207506. [DOI] [PubMed] [Google Scholar]
- 37.Basaria S. Androgen deprivation therapy, insulin resistance, and cardiovascular mortality: an inconvenient truth. J Androl. 2008;29(5):534–539. doi: 10.2164/jandrol.108.005454. [DOI] [PubMed] [Google Scholar]
- 38.Braga-Basaria M, Dobs AS, Muller DC, Carducci MA, John M, Egan J, Basaria S. Metabolic syndrome in men with prostate cancer undergoing long-term androgen-deprivation therapy. J Clin Oncol. 2006;24(24):3979–3983. doi: 10.1200/JCO.2006.05.9741. [DOI] [PubMed] [Google Scholar]
- 39.Rezaei MM, Rezaei MM, Ghoreifi A, Kerigh BF. Metabolic syndrome in patients with prostate cancer undergoing intermittent androgen-deprivation therapy. Can Urol Assoc J. 2016;10(9–10):E300–E305. doi: 10.5489/cuaj.3655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Lipscombe LL, Fischer HD, Yun L, Gruneir A, Austin P, Paszat L, Anderson GM, Rochon PA. Association between tamoxifen treatment and diabetes: a population-based study. Cancer. 2012;118(10):2615–2622. doi: 10.1002/cncr.26559. [DOI] [PubMed] [Google Scholar]
- 41.Iyengar NM, Gucalp A, Dannenberg AJ, Hudis CA. Obesity and cancer mechanisms: tumor microenvironment and inflammation. J Clin Oncol. 2016;34(35):4270–4276. doi: 10.1200/JCO.2016.67.4283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Culig Z, Puhr M. Interleukin-6: a multifunctional targetable cytokine in human prostate cancer. Mol Cell Endocrinol. 2012;360(1–2):52–58. doi: 10.1016/j.mce.2011.05.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Hobisch A, Eder IE, Putz T, Horninger W, Bartsch G, Klocker H, Culig Z. Interleukin-6 regulates prostate-specific protein expression in prostate carcinoma cells by activation of the androgen receptor. Cancer Res. 1998;58(20):4640–4645. [PubMed] [Google Scholar]
- 44.Castellano I, Allia E, Accortanzo V, Vandone AM, Chiusa L, Arisio R, Durando A, Donadio M, Bussolati G, Coates AS, Viale G, Sapino A. Androgen receptor expression is a significant prognostic factor in estrogen receptor positive breast cancers. Breast Cancer Res Treat. 2010;124(3):607–617. doi: 10.1007/s10549-010-0761-y. [DOI] [PubMed] [Google Scholar]
- 45.Hu R, Dawood S, Holmes MD, Collins LC, Schnitt SJ, Cole K, Marotti JD, Hankinson SE, Colditz GA, Tamimi RM. Androgen receptor expression and breast cancer survival in postmenopausal women. Clin Cancer Res. 2011;17(7):1867–1874. doi: 10.1158/1078-0432.CCR-10-2021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.McNamara KM, Moore NL, Hickey TE, Sasano H, Tilley WD. Complexities of androgen receptor signalling in breast cancer. Endocr Relat Cancer. 2014;21(4):T161–T181. doi: 10.1530/ERC-14-0243. [DOI] [PubMed] [Google Scholar]
- 47.Robinson JL, Macarthur S, Ross-Innes CS, Tilley WD, Neal DE, Mills IG, Carroll JS. Androgen receptor driven transcription in molecular apocrine breast cancer is mediated by Foxa1. EMBO J. 2011;30(15):3019–3027. doi: 10.1038/emboj.2011.216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Rinaldi S, Key TJ, Peeters PH, Lahmann PH, Lukanova A, Dossus L, Biessy C, Vineis P, Sacerdote C, Berrino F, Panico S, Tumino R, Palli D, Nagel G, Linseisen J, Boeing H, Roddam A, Bingham S, Khaw KT, Chloptios J, Trichopoulou A, Trichopoulos D, Tehard B, Clavel-Chapelon F, Gonzalez CA, Larranaga N, Barricarte A, Quiros JR, Chirlaque MD, Martinez C, Monninkhof E, Grobbee DE, Bueno-de-Mesquita HB, Ferrari P, Slimani N, Riboli E, Kaaks R. Anthropometric measures, endogenous sex steroids and breast cancer risk in postmenopausal women: a study within the epic cohort. International journal of cancer Journal international du cancer. 2006;118(11):2832–2839. doi: 10.1002/ijc.21730. [DOI] [PubMed] [Google Scholar]
- 49.Key TJ, Appleby PN, Reeves GK, Roddam A, Dorgan JF, Longcope C, Stanczyk FZ, Stephenson HE, Jr, Falk RT, Miller R, Schatzkin A, Allen DS, Fentiman IS, Key TJ, Wang DY, Dowsett M, Thomas HV, Hankinson SE, Toniolo P, Akhmedkhanov A, Koenig K, Shore RE, Zeleniuch-Jacquotte A, Berrino F, Muti P, Micheli A, Krogh V, Sieri S, Pala V, Venturelli E, Secreto G, Barrett-Connor E, Laughlin GA, Kabuto M, Akiba S, Stevens RG, Neriishi K, Land CE, Cauley JA, Kuller LH, Cummings SR, Helzlsouer KJ, Alberg AJ, Bush TL, Comstock GW, Gordon GB, Miller SR, Longcope C, Endogenous Hormones Breast Cancer Collaborative G Body mass index, serum sex hormones, and breast cancer risk in postmenopausal women. J Natl Cancer Inst. 2003;95(16):1218–1226. doi: 10.1093/jnci/djg022. [DOI] [PubMed] [Google Scholar]
- 50.Chun JY, Nadiminty N, Dutt S, Lou W, Yang JC, Kung HJ, Evans CP, Gao AC. Interleukin-6 regulates androgen synthesis in prostate cancer cells. Clin Cancer Res. 2009;15(15):4815–4822. doi: 10.1158/1078-0432.CCR-09-0640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Mauvais-Jarvis F, Clegg DJ, Hevener AL. The role of estrogens in control of energy balance and glucose homeostasis. Endocr Rev. 2013;34(3):309–338. doi: 10.1210/er.2012-1055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Navarro G, Allard C, Xu W, Mauvais-Jarvis F. The role of androgens in metabolism, obesity, and diabetes in males and females. Obesity (Silver Spring) 2015;23(4):713–719. doi: 10.1002/oby.21033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Tombal B, Borre M, Rathenborg P, Werbrouck P, Van Poppel H, Heidenreich A, Iversen P, Braeckman J, Heracek J, Baskin-Bey E, Ouatas T, Perabo F, Phung D, Hirmand M, Smith MR. Enzalutamide monotherapy in hormone-naive prostate cancer: primary analysis of an open-label, single-arm, phase 2 study. The lancet oncology. 2014;15(6):592–600. doi: 10.1016/S1470-2045(14)70129-9. [DOI] [PubMed] [Google Scholar]
- 54.Harvell DM, Richer JK, Singh M, Spoelstra N, Finlayson C, Borges VF, Elias AD, Horwitz KB. Estrogen regulated gene expression in response to neoadjuvant endocrine therapy of breast cancers: tamoxifen agonist effects dominate in the presence of an aromatase inhibitor. Breast Cancer Res Treat. 2008;112(3):489–501. doi: 10.1007/s10549-008-9923-6. [DOI] [PubMed] [Google Scholar]
- 55.Labrie F. Intracrinology in action: importance of extragonadal sex steroid biosynthesis and inactivation in peripheral tissues in both women and men. J Steroid Biochem Mol Biol. 2015;145:131–132. doi: 10.1016/j.jsbmb.2014.09.012. [DOI] [PubMed] [Google Scholar]
- 56.Sikora MJ, Cooper KL, Bahreini A, Luthra S, Wang G, Chandran UR, Davidson NE, Dabbs DJ, Welm AL, Oesterreich S. Invasive lobular carcinoma cell lines are characterized by unique estrogen-mediated gene expression patterns and altered tamoxifen response. Cancer Res. 2014;74(5):1463–1474. doi: 10.1158/0008-5472.CAN-13-2779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Kyvernitakis I, Albert US, Kalder M, Winarno AS, Hars O, Hadji P. Effect of anastrozole on hormone levels in postmenopausal women with early breast cancer. Climacteric. 2015;18(1):63–68. doi: 10.3109/13697137.2014.929105. [DOI] [PubMed] [Google Scholar]
- 58.Takagi K, Miki Y, Nagasaki S, Hirakawa H, Onodera Y, Akahira J, Ishida T, Watanabe M, Kimijima I, Hayashi S, Sasano H, Suzuki T. Increased intratumoral androgens in human breast carcinoma following aromatase inhibitor exemestane treatment. Endocr Relat Cancer. 2010;17(2):415–430. doi: 10.1677/ERC-09-0257. [DOI] [PubMed] [Google Scholar]
- 59.Rossi E, Morabito A, Di Rella F, Esposito G, Gravina A, Labonia V, Landi G, Nuzzo F, Pacilio C, De Maio E, Di Maio M, Piccirillo MC, De Feo G, D'Aiuto G, Botti G, Chiodini P, Gallo C, Perrone F, de Matteis A. Endocrine effects of adjuvant letrozole compared with tamoxifen in hormone-responsive postmenopausal patients with early breast cancer: the Hoboe trial. J Clin Oncol. 2009;27(19):3192–3197. doi: 10.1200/JCO.2008.18.6213. [DOI] [PubMed] [Google Scholar]
- 60.Bajetta E, Martinetti A, Zilembo N, Pozzi P, La Torre I, Ferrari L, Seregni E, Longarini R, Salvucci G, Bombardieri E. Biological activity of anastrozole in postmenopausal patients with advanced breast cancer: effects on estrogens and bone metabolism. Annals of oncology: official journal of the European Society for Medical Oncology /nnnnnnESMO. 2002;13(7):1059–1066. doi: 10.1093/annonc/mdf083. [DOI] [PubMed] [Google Scholar]
- 61.Stanway SJ, Palmieri C, Stanczyk FZ, Folkerd EJ, Dowsett M, Ward R, Coombes RC, Reed MJ, Purohit A. Effect of tamoxifen or anastrozole on steroid sulfatase activity and serum androgen concentrations in postmenopausal women with breast cancer. Anticancer Res. 2011;31(4):1367–1372. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(PDF 182 kb)