

Keywords: nerve regeneration, puerarin, autophagy, cerebral ischemia/reperfusion, AMPK-mTOR-ULK1 signaling pathway, light chain 3, p62, ischemic stroke, AMPK/mTOR, traditional Chinese medicine, middle cerebral artery occlusion, neural regeneration
Abstract
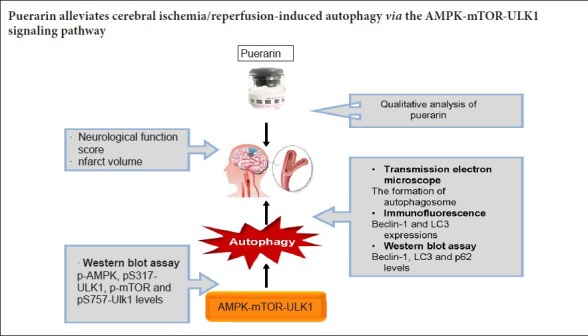
Puerarin suppresses autophagy to alleviate cerebral ischemia/reperfusion injury, and accumulating evidence indicates that the AMPK-mTOR signaling pathway regulates the activation of the autophagy pathway through the coordinated phosphorylation of ULK1. In this study, we investigated the mechanisms underlying the neuroprotective effect of puerarin and its role in modulating autophagy via the AMPK-mTOR-ULK1 signaling pathway in the rat middle cerebral artery occlusion model of cerebral ischemia/reperfusion injury. Rats were intraperitoneally injected with puerarin, 50 or 100 mg/kg, daily for 7 days. Then, 30 minutes after the final administration, rats were subjected to transient middle cerebral artery occlusion for 90 minutes. Then, after 24 hours of reperfusion, the Longa score and infarct volume were evaluated in each group. Autophagosome formation was observed by transmission electron microscopy. LC3, Beclin-1 p62, AMPK, mTOR and ULK1 protein expression levels were examined by immunofluorescence and western blot assay. Puerarin substantially reduced the Longa score and infarct volume, and it lessened autophagosome formation in the hippocampal CA1 area following cerebral ischemia/reperfusion injury in a dose-dependent manner. Pretreatment with puerarin (50 or 100 mg/kg) reduced Beclin-1 expression and the LC3-II/LC3-I ratio, as well as p-AMPK and pS317-ULK1 levels. In comparison, it increased p62 expression. Furthermore, puerarin at 100 mg/kg dramatically increased the levels of p-mTOR and pS757-ULK1 in the hippocampus on the ischemic side. Our findings suggest that puerarin alleviates autophagy by activating the APMK-mTOR-ULK1 signaling pathway. Thus, puerarin might have therapeutic potential for treating cerebral ischemia/reperfusion injury.
Introduction
Ischemic stroke is a very complex pathophysiological process and is a major cause of mortality and disability worldwide (Hankey, 2012). The most effective treatment for acute ischemic stroke is thrombolytic therapy with recombinant tissue plasminogen activator soon after the ischemic event. However, this approach may lead to reperfusion injury and a series of adverse reactions, including inflammation, calcium overload, glutamate excitotoxicity, oxidative stress, apoptosis and autophagy (Chen et al., 2012; Lanzillotta et al., 2013; Yin et al., 2013; Zhang et al., 2016). In addition, only a small percentage of patients are eligible for recombinant tissue plasminogen activator therapy because of the narrow therapeutic time window, high risk of hemorrhagic transformation, and other multiple contraindications, such as symptomatic intracranial hemorrhage (Culman et al., 2012; Jickling et al., 2014; Acampa et al., 2017). Thus, it is essential to explore an alternative and complementary medicine which not only rapidly restores oxygen supply to ischemic tissues, but also reduces reperfusion injury and improves the benefit-to-risk ratio for patients with ischemic stroke. The root of Pueraria lobata (kudzu root, Gegen in Chinese), a common traditional Chinese herb, has been shown to alleviate hangover, alcoholism, measles, menopausal symptoms, diabetes mellitus, fever, diarrhea and migraine for over 2000 years (Wong et al., 2011). Puerarin (C21H20O9), one of the main isoflavones in the extract of Pueraria lobata, has been used for the prevention and treatment of cardio-cerebrovascular diseases for over 20 years in mainland China (Tang et al., 2007; Zheng et al., 2017). Our previous study, a meta-analysis, showed that puerarin injection might be effective and safe for ischemic stroke in the clinic (Zheng et al., 2017). In an earlier study (Liu et al., 2013), we found that puerarin attenuates cerebral ischemia/reperfusion (I/R) injury by modulating the anti-inflammatory response. Furthermore, accumulating evidence suggests that puerarin alleviates cerebral I/R injury by improving the microcirculation, and by attenuating inflammation, apoptosis and autophagy (Lim et al., 2013; Wu et al., 2014; He et al., 2017; Liu et al., 2017). However, the underlying mechanisms remain unclear.
Autophagy is a regulated process for the breakdown and recycling of cellular constituents, such as damaged organelles, damaged and misfolded proteins, and even intracellular pathogens (Rabinowitz and White, 2010). Adenosine monophosphate-activated protein kinase (AMPK) is a pivotal energy sensor in maintaining metabolic homeostasis, and has been shown to initiate autophagy. Mammalian target of rapamycin (mTOR) is a conserved serine/threonine kinase that regulates cell growth and autophagy (Alers et al., 2012; Fu et al., 2016; Dai et al., 2017; Shen et al., 2017). AMPK and mTOR regulate global ischemia-induced autophagy through the coordinated phosphorylation of Unc-51-like kinase 1 (ULK1). AMPK phosphorylates ULK1 at Ser317 to activate autophagy, but the activity of AMPK can be suppressed by mTOR via increased phosphorylation of ULK1 at Ser757 (Kim et al., 2011; Tian et al., 2015; Hwang et al., 2017). Therefore, the AMPK-mTOR-ULK1 signaling pathway may be a novel therapeutic target for treating cerebral I/R injury by modulating autophagy.
Recent studies show that puerarin protects against cardiomyocyte hypertrophy and ethanol-induced liver damage by restoring autophagy through the AMPK-mTOR signaling pathway (Noh et al., 2011; Liu et al., 2015). He et al. (2017) showed that puerarin reduces ischemia-induced cerebral edema, neurological deficits and infarct volume to confer neuroprotection against cerebral ischemic injury, and this neuroprotection was achieved by inhibiting autophagy. Therefore, in the present study, we investigate whether puerarin protects against cerebral I/R injury by modulating the AMPK-mTOR-ULK1 pathway in the rat.
Materials and Methods
Animals
Forty specific-pathogen-free male Sprague-Dawley rats aged 10 weeks and weighing 250–280 g were purchased from the Laboratory Animal Center of China Three Gorges University (Yichang, China; license No. SCXK (E) 2011-0012). All rats were maintained under a 12-hour light-dark cycle, with 60 ± 5% humidity and a temperature of 22 ± 3°C, and were allowed free access to water and food. The animal experiments were conducted in accordance with the guidelines of the Laboratory Animal Management Committee of Three Gorges University, China (No. 2016101A) and the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH publication number 85-23, revised 1985).
Puerarin injection was purchased from North China Pharmaceutical Co., Ltd. (FALT150901; Shijiazhuang, China). Freshly prepared puerarin injection was dissolved in physiological saline to a final concentration of 20 mg/mL. Standard purified puerarin (purity: 99.71%) was purchased from Chengdu Mann Stewart Biological Technology Co. Ltd. (Chengdu, China; 15012916).
Rats were randomly divided into four groups after seven days of adaptation, as follows: sham group, I/R group, puerarin (PU) (50 mg/kg) group and PU (100 mg/kg) group. Based on the study of Wu et al. (2014), we selected 50 and 100 mg/kg doses of puerarin for administration. Before surgery, the rats in the sham and I/R groups were given physiological saline (100 mg/kg) once daily for 7 days. The rats in the PU (50 and 100 mg/kg) groups were given intraperitoneal injection of puerarin at 50 or 100 mg/kg, once daily for 7 days. On day 7, 30 minutes after the puerarin administration, rats were subjected to middle cerebral artery occlusion (MCAO).
High performance liquid chromatography (HPLC)
To evaluate the quality of puerarin for injection, HPLC was performed using an Alliance 2695 HPLC system (Waters Corporation, Milford, MA, USA) and a Welchrom C18 column (4.6 mm × 250 mm, 5 μm) (Welch Materials, Inc., Austin, TX, USA). The mobile phase consisted of 0.2% phosphoric acid in water (A) and acetonitrile (B). The following gradient program was used: 0–60 minutes, 10–65% acetonitrile. The flow rate was 1 mL/min. The ultraviolet wavelength was set at 254 nm, and the sample injection volume was 10 μL. The chromatographic data were recorded and processed with Empower 3.0 software (Waters Corporation).
Surgical procedures for MCAO
MCAO surgery was carried out as previously described (Longa et al., 1989). Rats were fasted overnight, but allowed free access to water. Before surgery, rats were intraperitoneally anesthetized with 10% chloral hydrate, 350 mg/kg (Sinopharm Chemical Reagent Co., Ltd., Shanghai, China). A monofilament nylon suture (4-0) with a spherical tip was inserted into the internal carotid artery through the external carotid artery stump and gently advanced into the internal carotid artery, approximately 18–22 mm towards the circle of Willis. The nylon suture was slowly pulled out after 90 minutes of transient ischemia to allow reperfusion. Rats in the sham group only underwent vessel separation without filament introduction. Rats were considered to have undergone successful induction of focal cerebral ischemia when their neurological deficit scores were between 1 and 3.
Neurological deficits
As in the study by Longa et al. (1989), neurological deficits were measured after 24 hours of reperfusion using the following scale: Normal, 0 = no motor deficits; mild, 1 = torso turning to the ipsilateral side when held by the tail and exhibiting forelimb weakness; moderate, 2 = circling to the contralateral side, but normal posture at rest; severe, 3 = the affected side is unable to bear weight at rest; critical, 4 = no spontaneous locomotor activity or barrel rolling.
Infarct volume
After 24 hours of reperfusion, infarct volume was assessed by 2,3,5-triphenyl tetrazolium chloride (TTC) staining. Rats were anesthetized with 10% chloral hydrate (350 mg/kg), and the brains were rapidly removed and frozen for 15 minutes at −20°C. Each brain was sliced into five 2-mm-thick sections and stained with 2% TTC for 30 minutes at 37°C. Image-pro plus 6.0 (Media Cybernetics, Maryland, MD, USA) was used to scan and analyze the images. Infarct volume was calculated by taking the average of the infarct area on both sides of the slice and multiplying it by the section thickness.
Transmission electron microscopy
After 24 hours of reperfusion, rats were perfused with saline solution followed by 4% paraformaldehyde/2.5% glutaraldehyde in phosphate buffer. The hippocampal CA1 area was cut into blocks of 1 mm × 1 mm × 1 mm and postfixed in 2.5% glutaraldehyde at 4°C. The pieces were rinsed three times with 0.1 M phosphate buffer, and then immersed in 1% osmium tetroxide in 0.1 M phosphate buffer at 4°C for 2 hours. Thereafter, the tissue blocks were dehydrated in graded ethanol solutions and embedded in epoxy resin, followed by polymerization at −80°C for 24 hours. Blocks were cut on a Leica ultramicrotome (Leica, Wetzlar, Germany) into ultrathin sections (60−70 nm). These sections were stained with 3% lead citrate, and subsequently examined using a transmission electron microscope (Hitachi, Tokyo, Japan).
Immunofluorescence
After 24 hours of reperfusion, total brain tissues were perfused with saline solution and 4% paraformaldehyde in phosphate buffer. The hippocampi were rapidly removed and postfixed overnight at 4°C in 4% paraformaldehyde. The hippocampi were cut into 20-μm coronal sections, which were then preincubated in 0.3% Triton X-100 in phosphate buffer for 30 minutes, and then incubated overnight with light chain 3 (LC3) primary antibody (1:200; Cell Signaling Technology, Danvers, MA, USA) at 4°C or Beclin-1 primary antibody (1:200; Cell Signaling Technology). After three rinses with phosphate buffer, sections were incubated with goat anti-rabbit IgG (H+L) secondary antibody (1:200; Boster Biological Technology, Wuhan, China) at room temperature for 2 hours, and then washed three times with phosphate buffer. Nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI). The sections were mounted on a coverslip with an antifade mounting medium (Beyotime, Shanghai, China). Cells were observed at 400× magnification under a fluorescence microscope (Leica, Wetzlar, Germany).
Western blot assay
Hippocampi were lysed in ice-cold lysis buffer for 10 minutes and centrifuged at 12,000 × g for 15 minutes. Equal amounts of total protein were separated by 10% (w/v) sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Afterwards, proteins were transferred to polyvinylidene difluoride membranes (Millipore, Bedford, MA, US). Membranes were blocked in 5% (w/v) non-fat milk at room temperature for 1 hour, and then incubated overnight with anti-rat LC3, Beclin-1 polyclonal antibody (1:1000; Abcam, Cambridge, UK), anti-rat AMPK, p-AMPK, mTOR, p-mTOR, ULK1, p-ULK1 (S317), p-ULK1 (S757) monoclonal antibody, anti-rat p62 polyclonal antibody (1:1000; Cell Signaling Technology) or goat anti-rat GAPDH antibody (1:2000; Boster) at 4°C. Membranes were rinsed twice in Tris-buffered saline/Tween-20 (10 minutes each). The blots were then incubated with a peroxidase-conjugated goat anti-rabbit IgG (1:3000; Boster) for 1 hour at room temperature, and signals were visualized with Immobilon Western Chemiluminescent HRP Substrate (Boster). GAPDH was used as the loading control, and the optical density of the target protein to GAPDH was measured using Image J software (Wayne Rasband, National Institutes of Health, USA).
Statistical analysis
Quantitative data were expressed as the mean ± SEM and analyzed with SPSS 19.0 software (IBM, Armonk, NY, USA). All data were analyzed using one-way analysis of variance followed by the least significant difference test. A P-value < 0.05 was considered statistically significant.
Results
Puerarin purity analysis by HPLC
By comparing the retention times and the spectra of the reference standard, puerarin was definitively identified in the puerarin injection (Figure 1).
Figure 1.
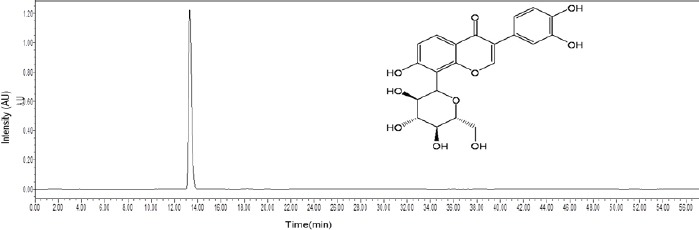
High performance liquid chromatography fingerprint chromatogram of puerarin injection at 254 nm.
Puerarin alleviated neurological deficits and reduces infarct volume in rats with cerebral I/R injury
We evaluated neurological scores after 24 hours of reperfusion. Rats in the sham group did not exhibit any symptoms of neurobehavioral dysfunction (Longa's score 0), while rats in the I/R group displayed significant neurobehavioral dysfunction (P < 0.01, vs. sham group). The scores were significantly better in the PU (100 mg/kg) group than in the I/R group (Figure 2A; P < 0.01). Neurological deficits were also alleviated in the PU (50 mg/kg) group compared with the I/R group (Figure 2A; P < 0.05).
Figure 2.

Effects of puerarin on neurological deficits and infarct volume of rats after 24 hours of reperfusion.
Neurological deficits were assessed using Longa's score, and infarct volume was assessed using TTC staining. (A) Neurological scores. (B) Images of TTC-stained brain sections. Red areas indicate non-infarcted tissue, and white areas indicate infarcted tissue. (C) Infarct volume ratio. Data are expressed as the mean ± SEM (n = 3 rats per group) and were analyzed by one-way analysis of variance followed by the least significant difference test. **P < 0.01, vs. sham group; #P < 0.05, ##P < 0.01, vs. I/R group. PU: Puerarin; I/R: ischemia/reperfusion; TTC: 2,3,5-triphenyl tetrazolium chloride.
Infarct volume was measured by TTC staining after 24 hours of reperfusion. Red areas indicate non-infarcted regions, while white areas indicate infarcted brain tissue (Figure 2B). In comparison with the sham group, infarct volume was significantly increased in the I/R group (P < 0.01). Pretreatment with puerarin at either dose significantly reduced infarct volume compared with the I/R group (Figure 2C; P < 0.05 and P < 0.01). The protective effect of 100 mg/kg puerarin was more robust than that of 50 mg/kg puerarin; however, the difference was not statistically significant.
Puerarin reduced the formation of autophagosomes following cerebral I/R injury
Detection of autophagic vesicles by electron microscopy is considered the gold standard for identifying autophagy. Therefore, we assessed the formation of autophagosomes in neurons in the hippocampal CA1 by electron microscopy. As shown in Figure 3, neurons in the sham group appeared relatively normal, with comparative healthy-looking nuclei, mitochondria, lysosomes and endoplasmic reticulum. After 24 hours of reperfusion, many lysosomes and autophagosomes could be found in neurons. Cellular morphology was heavily impacted. Neurons were moderately vacuolated, and the mitochondria had lost their integrity. Puerarin pretreatment decreased the number of lysosomes and autophagosomes and alleviated the neuronal injury. The 100 mg/kg dose had a more robust neuroprotective effect than the 50 mg/kg dose.
Figure 3.
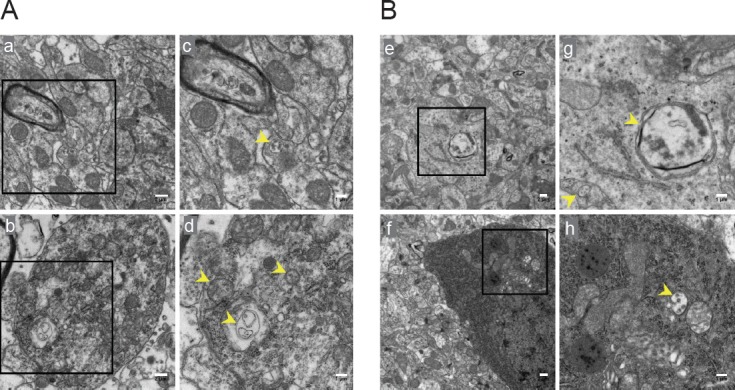
The formation of autophagosomes assessed by electron microscopy in the hippocampal CA1 area of rats.
(A) Transmission electron microscopy showing autophagosomes (yellow arrow) in the CA1 in the sham and I/R groups. (B) Transmission electron microscopy showing autophagosomes (yellow arrow) in the CA1 in the PU (50 mg/kg) and PU (100 mg/kg) groups. The squares in a, b, e and f indicate the area enlarged in c, d, g and h, respectively. Original magnification: a, b, 5000×; c, d, g, h; 8000×; e, f, 3000×. Scale bars: a, b, e, f, 1 μm; c, d, g, h, 2 μm. PU: Puerarin; I/R: ischemia/reperfusion.
Puerarin reduced hippocampal expression of LC3 and Beclin-1 after cerebral I/R
To examine autophagic activity after cerebral I/R and to determine whether puerarin inhibits the activation of autophagy, immunofluorescence was performed for LC3 and Beclin-1 in the sham, I/R, PU (50 mg/kg) and PU (100 mg/kg) groups. Both LC3 and Beclin-1 have been used as specific markers for autophagy. As shown in Figure 4, the expression of LC3 and beclin-1 proteins in the CA1 area were significantly increased in the I/R group compared with the sham group (P < 0.01). Pretreatment with puerarin at 100 mg/kg significantly attenuated this increase in LC3 and Beclin-1 levels. Pretreatment with puerarin at 50 mg/kg also significantly decreased beclin-1 expression (Figure 4B, D; P < 0.05), without significantly affecting LC3 expression.
Figure 4.

Effects of puerarin on LC3 and Beclin-1 expression in the hippocampal CA1 area after 24 hours of reperfusion.
(A) LC3-positive (red) neurons. The blue is DAPI staining. (B) Mean fluorescence intensity for LC3. (C) Beclin-1-positive (red) neurons. The blue is DAPI staining. (D) Mean fluorescence intensity for Beclin-1. Data are expressed as the mean ± SEM (n = 3 rats per group) and were analyzed by one-way analysis of variance followed by the least significant difference test. **P < 0.01, vs. sham group; #P < 0.05, vs. I/R group. Images were captured at 400× magnification. Scale bars: 20 μm. PU: Puerarin; I/R: ischemia/reperfusion, LC3: light chain 3; DAPI: 4′,6-diamidino-2-phenylindole.
Puerarin decreased the LC3-II/LC3-I ratio and Beclin-1 expression, while increasing p62 expression
To further clarify the impact of puerarin on autophagy, LC3, p62 and Beclin-1 protein levels were evaluated after 24 hours of reperfusion by western blot assay. As shown in Figure 5, the LC3-II/LC3-I ratio and the expression of Beclin-1 significantly increased, while p62 levels decreased, after 24 hours of reperfusion. In comparison, pretreatment with puerarin reduced the LC3-II/LC3-I ratio and Beclin-1 levels, and increased p62 levels after 24 hours of reperfusion (Figure 5; P < 0.05 or P < 0.01). The PU (50 mg/kg and 100 mg/kg) groups showed a dose-dependent trend, but no significant difference in expression was observed between the two PU groups.
Figure 5.

Effects of puerarin on the expression of LC3-II/LC3-I, p62 and Beclin-1 in the hippocampus after 24 hours of reperfusion.
(A–C) Western blot assay for LC3, P62 and Beclin-1. Data are expressed as the mean ± SEM (n = 3 per group) and were analyzed by one-way analysis of variance followed by the least significant difference test. **P < 0.01, vs. sham group; #P < 0.05, ##P < 0.01, vs. I/R group. PU: Puerarin; I/R: ischemia/reperfusion; LC3: light chain 3.
Puerarin reduced p-AMPK and pS317-ULK1 levels
To investigate the mechanisms underlying the anti-autophagic effect of puerarin, we examined its effect on hippocampal p-AMPK and pS317-ULK1 levels in the ischemic brain. Figure 6 shows that p-AMPK and pS317-ULK1 levels were significantly increased in the I/R group compared with the sham group after 24 hours of reperfusion (P < 0.01). Pretreatment with puerarin at 50 mg/kg or 100 mg/kg significantly decreased the levels of p-AMPK and pS317-ULK1 (P < 0.05 and P < 0.01).
Figure 6.

Effects of puerarin on the levels of AMPK and S317-ULK1 in the hippocampus after 24 hours of reperfusion.
(A, B) Western blot assay for AMPK and S317-ULK1. GAPDH was used as a loading control. Data are expressed as the mean ± SEM (n = 3 per group) and were analyzed by one-way analysis of variance followed by the least significant difference test. **P < 0.01, vs. sham group; #P < 0.05, ##P < 0.01, vs. I/R group. PU: Puerarin; I/R: ischemia/reperfusion; AMPK: adenosine monophosphate-activated protein kinase; ULK1: uncoordinated 51-like kinase 1.
Puerarin increased p-mTOR and pS757-ULK1 levels
AMPK phosphorylates ULK1 at Ser317 to activate autophagy, while AMPK activity is suppressed by mTOR via increased phosphorylation of ULK1 at Ser757 (Kim et al., 2011; Tian et al., 2015; Hwang et al., 2017). Thus, mTOR, p-mTOR, ULK1 and pS757-ULK1 protein levels after 24 hours of reperfusion were further investigated by western blot assay. p-mTOR and pS757-ULK1 levels were significantly decreased in the I/R group compared with the sham group (Figure 7; P < 0.01). Puerarin at 100 mg/kg significantly increased p-mTOR and pS757-ULK1 levels compared with the I/R group (Figure 7; P < 0.05). Pretreatment with puerarin at 50 mg/kg significantly increased pS757-ULK1 levels (Figure 7; P < 0.05). p-mTOR levels were lower in the PU (50 mg/kg) group than in the I/R group (Figure 7; P > 0.05).
Figure 7.

Effects of puerarin on the levels of mTOR and S757-ULK1 in the hippocampus after 24 hours of reperfusion.
(A, B) Western blot assay for mTOR and S757-ULK1. GAPDH was used as a loading control. Data are expressed as the mean ± SEM (n = 3 per group) and were analyzed by one-way analysis of variance followed by the least significant difference test. *P < 0.05, **P < 0.01, vs. sham group; #P < 0.05, vs. I/R group. PU: Puerarin; I/R: ischemia/reperfusion; mTOR: mammalian target of rapamycin; ULK1: uncoordinated 51-like kinase 1.
Discussion
Intravenous administration of recombinant tissue plasminogen activator is currently the only Food and Drug Administration-approved thrombolytic therapy for ischemic stroke (Lin et al., 2016). However, only a small proportion of patients are given this treatment because of the narrow time window of drug administration and the risk of hemorrhagic transformation. Therefore, it is of utmost importance to discover alternative or complementary medicines with a wide therapeutic time window, which can inhibit hemorrhagic transformation, and which can protect the brain against reperfusion damage. In China, traditional Chinese medicine has been widely used for the prevention or treatment of cerebral ischemia or ischemic stroke and secondary complications. The China Food and Drug Administration (http://www.sda.gov.cn) lists a variety of traditional Chinese medicinal preparations, such as Buyang Huanwu decoction (Pan et al., 2017), Xiaoxuming decoction (Fu et al., 2013), Neuroaid (Chen et al., 2013), Xuesaitong injection (Wang et al., 2015), resveratrol (Li et al., 2016) and acanthopanax (Li et al., 2009), as well as puerarin (Chang et al., 2009; Wu et al., 2014), which is a therapeutically active isoflavone isolated from kudzu (Pueraria lobata) (Wong et al., 2011). Kudzu root is one of the oldest medicinal herbs, and has been used since ancient times in China and Southeast Asia. Kudzu root has antipyretic, antidiarrhetic, diaphoretic and antiemetic properties (Prasain et al., 2003). Numerous basic studies and clinical trials show that puerarin exhibits therapeutic effectiveness in various diseases, such as coronary heart disease, angina pectoris (Luo et al., 2012), cardiac infarction (Ma et al., 2016), obstruction of retinal arteries and veins (Hao et al., 2014), sudden deafness (Liu et al., 2002), diabetes (She et al., 2014) and ischemic stroke (Xu et al., 2005; Wang et al., 2014). Our previous study, a meta-analysis of 35 randomized controlled trials involving a total of 3,224 participants with ischemic stroke, showed that puerarin injection might be effective and safe for ischemic stroke (Zheng et al., 2017). To explore the underlying mechanisms, animal experiments were carried out in our previous study (Liu et al., 2013), which indicated that puerarin reduces the expression of inflammatory cytokines, such as IL-6, IL-1β and TNF-α, to attenuate cerebral I/R injury. Xu et al. (2005) reported that puerarin treatment in rats with MCAO reduced apoptosis and necrosis by 38.6% and 28.5%, respectively, and substantially inhibited caspase-3 activity, a biochemical marker of apoptosis, suggesting that the neuroprotection afforded by puerarin against cerebral ischemia involves an anti-apoptotic effect. Wu et al. (2014) showed that puerarin increases cerebral blood perfusion both by arteriolar relaxation and p42/44 MAPK-mediated angiogenesis, suggesting that puerarin exerts anti-hypertensive and anti-stroke actions by improving microcirculation. Recently, He et al. (2017) demonstrated that puerarin downregulates LC3 and beclin-1 expression, thereby suppressing cerebral I/R-induced autophagy. However, the mechanisms underlying the beneficial effects of puerarin remain unclear, which prevents the widespread utilization of puerarin and kudzu root worldwide as an alternative or complementary medicine.
In the present study, we investigated the beneficial effects of puerarin pretreatment against cerebral I/R injury and the mechanisms underlying the anti-autophagic action of puerarin. Puerarin alleviated neurological deficits and reduced infarct volume in a dose-dependent manner, suggesting that the 7-day pretreatment provides robust neuroprotection against focal cerebral I/R injury. Furthermore, pretreatment with puerarin at 50 or 100 mg/kg decreased the formation of autophagosomes in the CA1 area, suppressed the increase in the LC3-II/LC3-I ratio, and increased p62 levels in the ischemic hippocampus, suggesting direct physical interaction between p62 and LC3. Pretreatment with puerarin also downregulated Beclin-1 in the ischemic hippocampus.
p62/SQSTM1 plays a critical role in the formation of intracellular ubiquitin-related protein aggregates caused by deficient autophagy, and it binds directly to LC3 and is degraded through the autophagic-lysosomal pathway, implying complete autophagic flux (Kihara et al., 2001; Klionsky et al., 2012). LC3 is present as two forms; LC3-I (cytoplasmic form) and LC3-II (processed form), which is the only protein marker reliably associated with mature autophagosomes. Beclin-1 is critical for the nucleation phase of autophagy, and it promotes the conversion of LC3-I to LC3-II to induce autophagy (Komatsu et al., 2007; Kuma et al., 2007). These findings suggest that that the neuroprotective effect of puerarin is associated with the suppression of autophagy, consistent with the study of He et al. (2017). Our findings are also consistent with other previous studies (Koike et al., 2008; Baek et al., 2014), which show that suppression of autophagy ameliorates ischemic brain damage after 24 hours of reperfusion. However, in contrast, Carloni et al. (2008) and Buckley et al. (2014) reported that induction of autophagy protects against ischemic stroke damage after 24 hours of reperfusion. This lack of consensus on the role of autophagy in ischemic stroke injury may in large part be caused by differences in the critical therapeutic time window, when autophagy was assessed and methodological approaches (Buckley et al., 2014). A recent study suggests that autophagy peaks at 12−24 hours in a cerebral ischemic animal model (Tian et al., 2010). Moderate autophagy may exert a neuroprotective effect, while excessive autophagy could induce cell death. Hence, a potential therapeutic strategy for ischemic stroke is to optimize the level of autophagy, and puerarin may help to achieve this.
Accumulating evidence demonstrates that the AMPK-mTOR signaling pathway (Liang et al., 2007; Wang et al., 2012) can modulate autophagic activation following cerebral I/R through the coordinated phosphorylation of ULK1 (Hwang et al., 2017). A recent study suggests that AMPK is an upstream regulator of mTOR in mammalian cells, and it is well known that autophagy is promoted by AMPK and inhibited by mTOR (Kim et al., 2011). Indeed, in the present study, we found that p-AMPK levels were dramatically upregulated, while p-mTOR levels were remarkably downregulated, in the I/R group (Figures 6 and 7). Generally, activated AMPK can inhibit mTOR to reduce phosphorylation of ULK1 on Ser757. AMPK then phosphorylates ULK1 on Ser317, leading to ULK1–AMPK interaction, and ultimately, the induction of autophagy (Kim et al., 2011; Russell et al., 2013; Tian et al., 2015; Hwang et al., 2017).
In our present study, pS317-ULK1 levels were substantially increased, while pS757-ULK1 levels were decreased, in the I/R group. These results are in line with those of Hwang et al. (2017), who used a global ischemic rat model. As in our present study, these investigators found that AMPK inhibits mTOR to reduce S757-ULK1 phosphorylation, and then phosphorylates S317-ULK1 to induce autophagy during cerebral I/R injury. Here, puerarin dramatically decreased the levels of p-AMPK and pS317-ULK1, while it increased the levels of p-mTOR and pS757-ULK1. There was a tendency towards dose-dependency (50 vs. 100 mg/kg), although it was not statistically significant. This may be a result of the limited sample size, which will be optimized in our future studies.
Taken together, our findings suggest that puerarin inhibits cerebral I/R-induced autophagy. This effect of puerarin may involve the suppression of AMPK expression and inhibition of the phosphorylation of ULK1 at S317, as well as an enhancement of mTOR expression and phosphorylation of ULK1 at Ser757 (Figure 8). However, it is still unclear whether puerarin also regulates other pathways, such as the c-Jun N-terminal kinase pathway, to inhibit autophagy. Moreover, further investigation is needed to clarify whether puerarin widens the therapeutic time window for tissue plasminogen activator administration and whether it inhibits hemorrhagic transformation. We will tackle these unresolved issues in future studies, with the aim of optimizing treatment for ischemic stroke.
Figure 8.
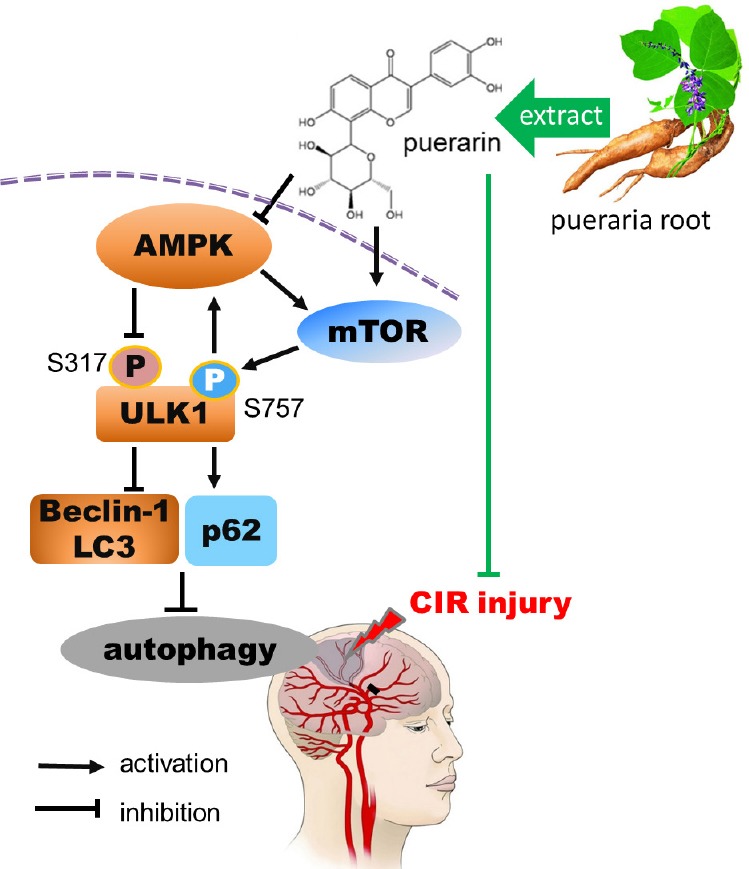
Proposed scheme showing how puerarin suppresses CIR-induced autophagy via the AMPK-mTOR-ULK1 signaling pathway.
Puerarin pretreatment alleviates neurological deficits, decreases infarct volume, upregulates p62 and lowers the LC3-II/LC3-I ratio and Beclin-1 expression. These effects involve suppression of AMPK expression and the inhibition of phosphorylation of ULK1 at S317, as well as the enhancement of mTOR expression and the phosphorylation of ULK1 at Ser757. CIR: Cerebral ischemia/reperfusion; AMPK: adenosine monophosphate-activated protein kinase; mTOR: mammalian target of rapamycin; ULK1: uncoordinated 51-like kinase 1; LC3: light chain 3.
Footnotes
Conflicts of interest: The authors declare that they have no conflicts of interest.
Financial support: This work was supported by the National Natural Science Foundation of China, No. 81202625; the Open Fund of Key Laboratory of Cardiovascular and Cerebrovascular Diseases Translational Medicine, China Three Gorges University, China, No. 2016xnxg101. The funders did not participate in the study design, in the collection, analysis and interpretation of data, in the writing of the paper, and in the decision to submit the paper for publication.
Institutional review board statement: The animal experiments were executed in accordance with the Laboratory Animal Management Committee of Three Gorges University (approval No. 2016101A) and the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH Publication number 85-23, revised 1985).
Copyright license agreement: The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement: Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Funding: This work was supported by the National Natural Science Foundation of China, No. 81202625; the Open Fund of Key Laboratory of Cardiovascular and Cerebrovascular Diseases Translational Medicine, China Three Gorges University, China, No. 2016xnxg101.
(Copyedited by Wang J, Li CH, Qiu Y, Song LP, Zhao M)
References
- Acampa M, Camarri S, Lazzerini PE, Guideri F, Tassi R, Valenti R, Cartocci A, Martini G. Increased arterial stiffness is an independent risk factor for hemorrhagic transformation in ischemic stroke undergoing thrombolysis. Int J Cardio. 2017;243:466–470. doi: 10.1016/j.ijcard.2017.03.129. [DOI] [PubMed] [Google Scholar]
- Alers S, Loffler AS, Wesselborg S, Stork B. Role of AMPK-mTORUlk1/2 in the regulation of autophagy: cross talk, shortcuts, and feedbacks. Mol Cell Biol. 2012;32:2–11. doi: 10.1128/MCB.06159-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baek SH, Noh AR, Kim KA, Akram M, Shin YJ, Kim ES, Yu SW, Majid A, Bae, ON Modulation of mitochondrial function and autophagy mediates carnosine neuroprotection against ischemic brain damage. Stroke. 2014;45:2438–2443. doi: 10.1161/STROKEAHA.114.005183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buckley KM, Hess DL, Sazonova IY, Periyasamy-Thandavan S, Barrett JR, Kirks R, Grace H, Kondrikova G, Johnson MH, Hess DC, Schoenlein PV, Hoda MN, Hill WD. Rapamycin up-regulation of autophagy reduces infarct size and improves outcomes in both permanent MCAL, and embolic MCAO, murine models of stroke. Exp Transl Stroke Med. 2014;6:8. doi: 10.1186/2040-7378-6-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carloni S, Buonocore G, Balduini W. Protective role of autophagy in neonatal hypoxia-ischemia induced brain injury. Neurobiol Dis. 2008;32:329–339. doi: 10.1016/j.nbd.2008.07.022. [DOI] [PubMed] [Google Scholar]
- Chang Y, Hsieh CY, Peng ZA, Yen TL, Hsiao G, Chou DS, Chen CM, Sheu JR. Neuroprotective mechanisms of puerarin in middle cerebral artery occlusion-induced brain infarction in rats. J Biomed Sci. 2009;16:9. doi: 10.1186/1423-0127-16-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen CL, Young SH, Gan HH, Singh R, Lao AY, Baroque AC, 2nd, Chang HM, Hiyadan JH, Chua CL, Advincula JM, Muengtaweepongsa S, Chan BP, de Silva HA, Towanabut S, Suwanwela NC, Poungvarin N, Chankrachang S, Wong KS, Eow GB, Navarro JC, Venketasubramanian N, Lee CF, Bousser MG. Chinese medicine neuroaid efficacy on stroke recovery: a double-blind, placebo-controlled, randomized study. Stroke. 2013;44:2093–2100. doi: 10.1161/STROKEAHA.113.002055. [DOI] [PubMed] [Google Scholar]
- Chen L, Wang L, Zhang X, Cui L, Xing Y, Dong L, Liu Z, Li Y, Zhang X, Wang C, Bai X, Zhang J, Zhang L, Zhao X. The protection by octreotide against experimental ischemic stroke: up-regulated transcription factor Nrf2, HO-1 and down-regulated NF-kappaB expression. Brain Res. 2012;1475:80–87. doi: 10.1016/j.brainres.2012.07.052. [DOI] [PubMed] [Google Scholar]
- Culman J, Nguyen-Ngoc M, Glatz T, Gohlke P, Herdegen T, Zhao Y. Treatment of rats with pioglitazone in the reperfusion phase of focal cerebral ischemia: a preclinical stroke trial. Exp Neurol. 2012;238:243–253. doi: 10.1016/j.expneurol.2012.09.003. [DOI] [PubMed] [Google Scholar]
- Dai SH, Chen T, Li X, Yue KY, Luo P, Yang LK, Zhu J, Wang YH, Fei Z, Jiang XF. Sirt3 confers protection against neuronal ischemia by inducing autophagy: Involvement of the AMPK-mTOR pathway. Free Radic Biol Med. 2017;108:345–353. doi: 10.1016/j.freeradbiomed.2017.04.005. [DOI] [PubMed] [Google Scholar]
- Fu DL, Lu L, Zhu W, Li JH, Li HQ, Liu AJ, Xie C, Zheng GQ. Xiaoxuming decoction for acute ischemic stroke: a systematic review and meta-analysis. J Ethnopharmacol. 2013;148:1–13. doi: 10.1016/j.jep.2013.04.002. [DOI] [PubMed] [Google Scholar]
- Fu L, Huang L, Cao C, Yin Q, Liu J. Inhibition of AMP-activated protein kinase alleviates focal cerebral ischemia injury in mice: Interference with mTOR and autophagy. Brain Res. 2016;1650:103–111. doi: 10.1016/j.brainres.2016.08.035. [DOI] [PubMed] [Google Scholar]
- Hankey GJ. Anticoagulant therapy for patients with ischaemic stroke. Nat Rev Neurol. 2012;8:319–328. doi: 10.1038/nrneurol.2012.77. [DOI] [PubMed] [Google Scholar]
- Hao CH, Wang WT, Shao MX, Zhao ZY, Tang LD. Effects of puerarin on experimental model of retinal vein occlusion in rats. Zhong Cao Yao. 2014;6:110–114. [Google Scholar]
- He HY, Guo T, Zhang PY, Yang LQ, Deng YH. Puerarin provides a neuroprotection against transient cerebral ischemia by attenuating autophagy at the ischemic penumbra in neurons but not in astrocytes. Neurosci Lett. 2017;643:45–51. doi: 10.1016/j.neulet.2017.02.009. [DOI] [PubMed] [Google Scholar]
- Hwang JY, Gertner M, Pontarelli F, Court-Vazquez B, Bennett MV, Ofengeim D, Zukin RS. Global ischemia induces lysosomal-mediated degradation of mTOR and activation of autophagy in hippocampal neurons destined to die. Cell Death Differ. 2017;24:317–329. doi: 10.1038/cdd.2016.140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jickling GC, Liu D, Stamova B, Ander BP, Zhan X, Lu A, Sharp FR. Hemorrhagic transformation after ischemic stroke in animals and humans. J Cereb Blood Flow Metab. 2014;34:185–199. doi: 10.1038/jcbfm.2013.203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kihara A, Kabeya Y, Ohsumi Y, Yoshimori T. Beclin-phosphatidylinositol 3-kinase complex functions at the trans-Golgi network. EMBO Rep. 2001;2:330–335. doi: 10.1093/embo-reports/kve061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim J, Kundu M, Viollet B, Guan KL. AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat Cell Biol. 2011;13:132–141. doi: 10.1038/ncb2152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klionsky DJ, Abdalla FC, Abeliovich H, Abraham RT, Acevedo-Arozena A, Adeli K. Guidelines for the use and interpretation of assays for monitoring autophagy. Autophagy. 2012;8:445–544. doi: 10.4161/auto.19496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koike M, Shibata M, Tadakoshi M, Gotoh K, Komatsu M, Waguri S, Kawahara N, Kuida K, Nagata S, Kominami E, Tanaka K, Uchiyama Y. Inhibition of autophagy prevents hippocampal pyramidal neuron death after hypoxic-ischemic injury. Am J Pathol. 2008;172:454–469. doi: 10.2353/ajpath.2008.070876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Komatsu M, Waguri S, Koike M, Sou YS, Ueno T, Hara T, Mizushima N, Iwata J, Ezaki J, Murata S, Hamazaki J, Nishito Y, Iemura S, Natsume T, Yanagawa T, Uwayama J, Warabi E, Yoshida H, Ishii T, Kobayashi A, Yamamoto M, Yue Z, Uchiyama Y, Kominami E, Tanaka K. Homeostatic levels of p62 control cytoplasmic inclusion body formation in autophagy-deficient mice. Cell. 2007;131:1149–1163. doi: 10.1016/j.cell.2007.10.035. [DOI] [PubMed] [Google Scholar]
- Kuma A, Matsui M, Mizushima N. LC3, an autophagosome marker, can be incorporated into protein aggregates independent of autophagy: caution in the interpretation of LC3 localization. Autophagy. 2007;3:323–328. doi: 10.4161/auto.4012. [DOI] [PubMed] [Google Scholar]
- Lanzillotta A, Pignataro G, Branca C, Cuomo O, Sarnico I, Benarese M, Annunziato L, Spano P, Pizzi M. Targeted acetylation of NF-kappaB/RelA and histones by epigenetic drugs reduces post-ischemic brain injury in mice with an extended therapeutic window. Neurobiol Dis. 2013;49:177–189. doi: 10.1016/j.nbd.2012.08.018. [DOI] [PubMed] [Google Scholar]
- Li W, Liu M, Feng S, Wu B, Zhang S, Yang W, Liu GJ. Acanthopanax for acute ischaemic stroke. Cochrane Database Syst Rev. 2009;8:CD007032. doi: 10.1002/14651858.CD007032.pub2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Z, Fang F, Wang Y, Wang L. Resveratrol protects CA1 neurons against focal cerebral ischemic reperfusion-induced damage via the ERK-CREB signaling pathway in rats. Pharmacol Biochem Behav. 2016;146-147:21–27. doi: 10.1016/j.pbb.2016.04.007. [DOI] [PubMed] [Google Scholar]
- Liang J, Shao SH, Xu ZX, Hennessy B, Ding Z, Larrea M, Kondo S, Dumont DJ, Gutterman JU, Walker CL, Slingerland JM, Mills GB. The energy sensing LKB1-AMPK pathway regulates p27 (kip1) phosphorylation mediating the decision to enter autophagy or apoptosis. Nat Cell Biol. 2007;9:218–224. doi: 10.1038/ncb1537. [DOI] [PubMed] [Google Scholar]
- Lim DW, Lee C, Kim IH, Kim YT. Anti-inflammatory effects of total isoflavones from Pueraria lobata on cerebral ischemia in rats. Molecules. 2013;18:10404–10412. doi: 10.3390/molecules180910404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin L, Wang X, Yu Z. Ischemia-reperfusion injury in the brain: mechanisms and potential therapeutic strategies. Biochem Pharmacol. 2016 doi: 10.4172/2167-0501.1000213. doi:10.4172/2167-0501.1000213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu B, Wu Z, Li Y, Ou C, Huang Z, Zhang J, Liu P, Luo C, Chen M. Puerarin prevents cardiac hypertrophy induced by pressure overload through activation of autophagy. Biochem Biophys Res Commun. 2015;464:908–915. doi: 10.1016/j.bbrc.2015.07.065. [DOI] [PubMed] [Google Scholar]
- Liu JM, Ma L, He WP. Therapeutic effect of puerarin therapy on sudden deafness. Di Yi Jun Yi Da Xue Xue Bao. 2002;22:1044–1045. [PubMed] [Google Scholar]
- Liu X, Mei Z, Qian J, Zeng Y, Wang M. Puerarin partly counteracts the inflammatory response after cerebral ischemia/reperfusion via activating the cholinergic anti-inflammatory pathway. Neural Regen Res. 2013;8:3203–3215. doi: 10.3969/j.issn.1673-5374.2013.34.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Y, Tang Q, Shao S, Chen Y, Chen W, Xu X. Lyophilized powder of catalpol and puerarin protected cerebral vessels from ischemia by its anti-apoptosis on endothelial cells. Int J Biol Sci. 2017;13:327–338. doi: 10.7150/ijbs.17751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Longa EZ, Weinstein PR, Carlson S, Cummins R. Reversible middle cerebral artery occlusion without craniectomy in rats. Stroke. 1989;20:84–91. doi: 10.1161/01.str.20.1.84. [DOI] [PubMed] [Google Scholar]
- Luo ZK, Liu Y, Li HM. A clinical efficacy and safety study on coronary heart disease and angina treatment with Puerarin Injection. Zhonghua Liu Xing Bing Xue Za Zhi. 2012;33:614–616. [PubMed] [Google Scholar]
- Ma Y, Gai Y, Yan J, Li J, Zhang Y. Puerarin attenuates anoxia/reoxygenation injury through enhancing bcl-2 associated athanogene 3 expression, a modulator of apoptosis and autophagy. Med Sci Monit. 2016;22:977–983. doi: 10.12659/MSM.897379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noh BK, Lee JK, Jun HJ, Lee JH, Jia Y, Hoang MH, Kim JW, Park KH, Lee SJ. Restoration of autophagy by puerarin in ethanol-treated hepatocytes via the activation of AMP-activated protein kinase. Biochem Biophys Res Commun. 2011;414:361–366. doi: 10.1016/j.bbrc.2011.09.077. [DOI] [PubMed] [Google Scholar]
- Pan R, Cai J, Zhan L, Guo Y, Huang RY, Li X, Zhou M, Xu D, Zhan J, Chen H. Buyang Huanwu decoction facilitates neurorehabilitation through an improvement of synaptic plasticity in cerebral ischemic rats. BMC Complement Altern Med. 2017;17:173. doi: 10.1186/s12906-017-1680-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prasain JK, Jones K, Kirk M, Wilson L, Smith-Johnson M, Weaver C, Barnes S. Profiling and quantification of isoflavonoids in kudzu dietary supplements by high-performance liquid chromatography and electrospray ionization tandem mass spectrometry. J Agric Food Chem. 2003;51:4213–4218. doi: 10.1021/jf030174a. [DOI] [PubMed] [Google Scholar]
- Rabinowitz JD, White E. Autophagy and metabolism. Science. 2010;330:1344–1348. doi: 10.1126/science.1193497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russell RC, Tian Y, Yuan H, Park HW, Chang YY, Kim J, Kim H, Neufeld TP, Dillin A, Guan KL. ULK1 induces autophagy by phosphorylating Beclin-1 and activating VPS34 lipid kinase. Nat Cell Biol. 2013;15:741–750. doi: 10.1038/ncb2757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- She S, Liu W, Li T, Hong Y. Effects of puerarin in STZ-induced diabetic rats by oxidative stress and the TGF-beta1/Smad2 pathway. Food Funct. 2014;5:944–950. doi: 10.1039/c3fo60565e. [DOI] [PubMed] [Google Scholar]
- Shen P, Hou S, Zhu M, Zhao M, Ouyang Y, Feng J. Cortical spreading depression preconditioning mediates neuroprotection against ischemic stroke by inducing AMP-activated protein kinase-dependent autophagy in a rat cerebral ischemic/reperfusion injury model. J Neurochem. 2017;140:799–813. doi: 10.1111/jnc.13922. [DOI] [PubMed] [Google Scholar]
- Tang Y, Zhong ZY, Ge YZ, Sheng GT, Cao PL. Effects of puerarin on the expression of heat shock protein 70 in ischemia/reperfusion injured myocardium. Zhongguo Zuzhi Gongcheng Yanjiu. 2007;11:10265–10268. [Google Scholar]
- Tian F, Deguchi K, Yamashita T, Ohta Y, Morimoto N, Shang J, Zhang X, Liu N, Ikeda Y, Matsuura T, Abe K. In vivo imaging of autophagy in a mouse stroke model. Autophagy. 2010;6:1107–1114. doi: 10.4161/auto.6.8.13427. [DOI] [PubMed] [Google Scholar]
- Tian W, Li W, Chen Y, Yan Z, Huang X, Zhuang H, Zhong W, Chen Y, Wu W, Lin C, Chen H, Hou X, Zhang L, Feng D. Phosphorylation of ULK1 by AMPK regulates translocation of ULK1 to mitochondria and mitophagy. FEBS Lett. 2015;589:1847–1854. doi: 10.1016/j.febslet.2015.05.020. [DOI] [PubMed] [Google Scholar]
- Wang L, Yu Y, Yang J, Zhao X, Li Z. Dissecting Xuesaitong's mechanisms on preventing stroke based on the microarray and connectivity map. Mol Biosyst. 2015;11:3033–3039. doi: 10.1039/c5mb00379b. [DOI] [PubMed] [Google Scholar]
- Wang N, Zhang Y, Wu L, Wang Y, Cao Y, He L, Li X, Zhao J. Puerarin protected the brain from cerebral ischemia injury via astrocyte apoptosis inhibition. Neuropharmacology. 2014;79:282–289. doi: 10.1016/j.neuropharm.2013.12.004. [DOI] [PubMed] [Google Scholar]
- Wang P, Guan YF, Du H, Zhai QW, Su DF, Miao CY. Induction of autophagy contributes to the neuroprotection of nicotinamide phosphoribosyltransferase in cerebral ischemia. Autophagy. 2012;8:77–87. doi: 10.4161/auto.8.1.18274. [DOI] [PubMed] [Google Scholar]
- Wong KH, Li GQ, Li KM, Razmovski-Naumovski V, Chan K. Kudzu root: traditional uses and potential medicinal benefits in diabetes and cardiovascular diseases. J Ethnopharmacol. 2011;134:584–607. doi: 10.1016/j.jep.2011.02.001. [DOI] [PubMed] [Google Scholar]
- Wu XD, Wang C, Zhang ZY, Fu Y, Liu FY, Liu XH. Puerarin attenuates cerebral damage by improving cerebral microcirculation in spontaneously hypertensive rats. Evid Based Complement Alternat Med. 2014;2014:408–501. doi: 10.1155/2014/408501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu X, Zhang S, Zhang L, Yan W, Zheng X. The neuroprotection of puerarin against cerebral ischemia is associated with the prevention of apoptosis in rats. Planta Med. 2005;71:585–591. doi: 10.1055/s-2005-871261. [DOI] [PubMed] [Google Scholar]
- Yin J, Tu C, Zhao J, Ou D, Chen G, Liu Y, Xiao X. Exogenous hydrogen sulfide protects against global cerebral ischemia/reperfusion injury via its anti-oxidative, anti-inflammatory and anti-apoptotic effects in rats. Brain Res. 2013;1491:188–196. doi: 10.1016/j.brainres.2012.10.046. [DOI] [PubMed] [Google Scholar]
- Zhang P, Guo ZF, Xu YM, Li YS, Song JG. N-Butylphthalide (NBP) ameliorated cerebral ischemia reperfusion-induced brain injury via HGF-regulated TLR4/NF-kappaB signaling pathway. Biomed Pharmacother. 2016;83:658–666. doi: 10.1016/j.biopha.2016.07.040. [DOI] [PubMed] [Google Scholar]
- Zheng QH, Li XL, Mei ZG, Xiong L, Mei QX, Wang JF, Tan LJ, Yang SB, Feng ZT. Efficacy and safety of puerarin injection in curing acute ischemic stroke: a meta-analysis of randomized controlled trials. Medicine (Baltimore) 2017;96:e5803. doi: 10.1097/MD.0000000000005803. [DOI] [PMC free article] [PubMed] [Google Scholar]