

Abstract
Activity in medial prefrontal cortex (mPFC) during persuasive messages predicts future message-consistent behavior change, but there are significant limitations to the types of persuasion processes that can be invoked inside an MRI scanner. For instance, real world persuasion often involves multiple people in conversation. Functional near infrared spectroscopy (fNIRS) allows us to move out of the scanner and into more ecologically valid contexts. As a first step, the current study used fNIRS to replicate an existing fMRI persuasion paradigm (i.e. the sunscreen paradigm) to determine if mPFC shows similar predictive value with this technology. Consistent with prior fMRI work, activity in mPFC was significantly associated with message-consistent behavior change, above and beyond self-reported intentions. There was also a difference in this association between previous users and non-users of sunscreen. Activity differences based on messages characteristics were not observed. Finally, activity in a region of right dorsolateral PFC (dlPFC), which has been observed with counterarguing against persuasive messages, correlated negatively with future behavior. The current results suggest it is reasonable to use fNIRS to examine persuasion paradigms that go beyond what is possible in the MRI scanner environment.
Keywords: fNIRS, persuasion, replication, mPFC, dlPFC, health behavior
Introduction
Persuasion is the art of soft power—using words and images to convince others to change their beliefs and behavior. Although the study of persuasion has an incredibly long history, our understanding of the brain’s role in persuasion processes is incredibly short. It is only in the last decade that a concerted effort has begun to localize some of the brain regions that contribute to persuasion and its behavioral consequences. Critically, researchers have used a ‘brain-as-predictor’ approach (Falk et al., 2015a) to predict whether an individual or even a mass audience will respond in the desired way to a persuasive message. In the current research, we extend this work to the use of functional near infrared spectroscopy (fNIRS), which allows for more portable, mobile and flexible neuroimaging than what is possible using functional magnetic resonance imaging (fMRI). Our goal was to replicate a recent fMRI persuasion paradigm using fNIRS to see whether the brain-as-predictor approach would succeed with this method as well.
Past fMRI persuasion research
Over the past decade, a number of studies have identified medial prefrontal cortex (mPFC) as a central predictor of whether persuasive messages will be successful in changing behavior. In a number of studies aimed at persuading behavior changes, ventromedial prefrontal cortex (vmPFC) or mPFC activity during message exposure was associated with smoking cessation (Chua et al., 2011; Falk et al., 2011; Cooper et al., 2015; Riddle et al., 2016), increased sunscreen use (Falk et al., 2010a; Vezich et al., 2017) and decreased sedentary behavior (Falk et al., 2015a; Cooper et al., 2017). In fact, when controlling for prior behavior and self-reported intentions to follow the messages, mPFC activity still adds significantly more predictive ability to persuasion outcome models (Falk et al., 2011; Riddle et al., 2016), accounting for as much as 23% more variability in behavior (Falk et al., 2010a).
Additionally, the predictive utility of mPFC responses is not restricted to the individual level in persuasion research. Other studies have also investigated the neural correlates of persuasion in focus groups, and then used that information to predict population-level behaviors. Activity in mPFC areas of participant samples has been used to predict the performance of large-scale phone (Falk et al., 2012) and e-mail anti-smoking campaigns (Falk et al., 2016), the future sales of new music (Berns and Moore, 2012) and amount of message propagation between people (Falk et al., 2013).
However, all of the above results come from fMRI research. There are many benefits to using fMRI technology for studying the neural correlates of persuasion, such as differentiating between the roles of prefrontal subdivisions in persuasion (Cooper et al., 2017) or analysing the interaction between cortical and subcortical areas in message processing (Ramsay et al., 2013). But fMRI is also a highly expensive apparatus that restricts the movement of research participants and is not transportable. These constraints limit the types of real world persuasion that can be studied with fMRI. For instance, populations in rural areas or who are mobility impaired cannot be easily recruited to university laboratories, and fMRI cannot be brought to them. Persuasion is also a highly contextualized and social phenomenon, but fMRI requires that participants be removed from the stimulus-rich environment they normally experience and disconnected from real-time interaction with other people. Some studies have studied real-time social influence with fMRI by virtually connecting participants to an outside agent or another participant in a separate MRI scanner (Montague et al., 2002; Schilbach et al., 2006; Saito et al., 2010; Wilms et al., 2010), but these paradigms are still much more artificial than everyday interpersonal interaction where influence often occurs.
In contrast, functional near infrared spectroscopy (fNIRS) is a similar neuroimaging method to fMRI that has not been used to study persuasion neuroscience before, but has important advantages over fMRI for this purpose (Cui et al., 2011; Ferrari and Quaresima, 2012). Both imaging modalities indirectly measure brain activity via the concentration of oxygenated and deoxygenated hemoglobin (HbO and HbR) in the cortex. But while fMRI uses the magnetic properties of HbO to make these measurements, fNIRS relies on the optical properties. Because skin and bone are relatively transparent to near infrared light, while HbO is not, researchers can measure HbO concentration changes in the cortex by affixing light emitters and detectors to the scalp, measuring the intensity of light that propagates through the head, and then calculating HbO concentration changes via the Modified Beer-Lambert Law (for more information on the biophysics of fNIRS, see Ferrari et al., 2004; Scholkmann et al., 2014). Because of this, a simple cap or headband can be worn to hold the optodes to the head. This means participants are free to sit up and move around during experiments. This considerably widens the range of experimental designs researchers may use, enabling them to test how neural activity during persuasion occurs in more naturalistic situations than laying in a dark and loud MRI scanner. Additionally, fNIRS is much less expensive to acquire and operate than fMRI, making it a more feasible option for policy makers or community researchers interested in applying neuroscience techniques to their work.
Given these advantages, fNIRS has the potential to be extremely useful in both the continued development of persuasion neuroscience and in the application of these findings. However, there are downsides to using fNIRS for neuroimaging—namely, its signal cannot penetrate more than a few centimeters into the cortex, its spatial resolution is not as fine as fMRI (fNIRS signal correlates best with MRI when an ROI radius of five voxels is used, Cui et al., 2011), and there is no structural brain image generated so the localization of activity must be inferred through the 10–20 external positioning system. Thus, before fNIRS can seriously be used to further our understanding of the brain during persuasion in naturalistic contexts, it is important to empirically test whether or not the brain-as-predictor findings from fMRI can replicate with fNIRS as well.
Current study
Given the need for testing the potential of fNIRS in persuasion neuroscience, the main goal of this study was to replicate our prior fMRI research. Specifically, we replicated the procedure of Vezich et al. (2017). In this work, participants viewed persuasive paragraphs about using sunscreen everyday, and then were surveyed about their intentions for future sunscreen use. A week later, they were re-contacted about their sunscreen use following the imaging session. They found a significant relationship between activity in the mPFC during message exposure and later sunscreen use after the scan. Our main goal then was to replicate these results in this a priori region of interest. While this particular study design does not capitalize on the advantages fNIRS poses for naturalistic experiments, a close replication of an fMRI paradigm will hopefully bolster confidence in using fNIRS for persuasion work in these other contexts.
The Vezich et al. study also probed the kind of persuasive messaging that elicited the best predictive neural activity, and whether or not this predictive ability varied as a function of individual differences across participants. These questions help clarify why the mPFC works as a predictor, and in what contexts its activity is a reliable indicator of later behavior. Specifically, the researchers tested the extent to which ‘gain’ vs ‘loss’ framing of a message would elicit differential mPFC activity, considering that factually equivalent information presented as a gain can increase motivation and self-relevant valuation more than loss framing (Kahneman and Tversky, 1979; Salovey and Wegener, 2003). In other words, it was predicted that describing the advantages of using sunscreen would elicit greater persuasion-related brain activity than describing the consequences of not using sunscreen. They found that mPFC activity in response to gain messages was significantly greater than activity during loss messages, and that the extent of this difference in each individual predicted amount of sunscreen use.
In addition, the activation difference between previous users and non-users of sunscreen was investigated (i.e. participants who either did or did not use sunscreen prior to participation in the experiment). Psychological models of persuasion and health behavior make a distinction between the processes necessary for initial behavior enactment and subsequent behavior maintenance (Weinstein et al., 1998; Miilunpalo et al., 2000; Fogg, 2009), and Vezich and colleagues indeed found that mPFC activity was associated with future sunscreen use in previous non-users of sunscreen, but not in existing sunscreen users.
Finally, Vezich et al. tested messages about ‘why’ to use sunscreen vs ‘how’ to use sunscreen in order to investigate the differential effects of intent and action planning, but the results for this contrast were located in the rostral inferior parietal lobule and posterior inferior frontal gyrus, which are areas that our fNIRS set up was not designed to measure. So in the current work, we repeated the contrasts of gain vs loss and users vs non-users to test whether the fMRI results in the mPFC generalize to fNIRS. It is important to note that while the general MPFC-behavior effects have been observed numerous times, the mPFC effects due to gain vs loss were novel to the Vezich study and user vs non-user investigations are also few in number (Weber et al., 2015).
This work will also investigate one more hypothesis that was not examined in Vezich et al. (2017)—whether or not the right dorsolateral prefrontal cortex (right dlPFC) might also be associated with behavior change after persuasive messaging—specifically, negatively associated. In Falk et al. (2010b), an area of the right dlPFC activated for unpersuasive messages more than persuasive messages, when viewing arguments about activities that people would have few strong opinions about. In other studies, dlPFC activity was associated with arguing against previously held attitudes (Kato et al., 2009; Ramsay et al., 2013). Additionally, in pilot work, our group has found that counterarguing against persuasive messaging was associated with both increased right dlPFC and decreased message consistent behavior change (Falk et al., unpublished data). Based on these findings, increased right dlPFC activity during persuasive messaging might reflect counterarguing and rejection of persuasive messages. If paired with a measure of mPFC activity, data from these two regions might give a more robust prediction of future behavior after persuasive message exposure. We therefore investigated whether decreased activity in the right dlPFC will also be a predictor of persuasion, and if combining the right dlPFC and mPFC into one model will be an even better predictor than either region alone.
Materials and methods
Participants
A total of 84 participants were recruited for participation in this study (37 sunscreen users, 47 non-users). This is roughly double the sample size of Vezich et al., because it was unknown how large the effect size would be for this study in the fNIRS modality. Of these participants, 3 did not complete the study, and the data from 12 more participants was deemed unusable due to poor neuroimaging data quality. The final sample consisted of 69 participants—29 sunscreen users and 40 non-users. All were female right-handed undergraduate students in university (M age= 21.25, s.d. age = 2.66). Written informed consent was obtained and all participants were paid for their participation. The study protocol was approved by the University of California—Los Angeles Institutional Review Board.
Pre-study questionnaire
In this questionnaire prior to completing the experimental task, participants were asked to report how many days in the previous 7 days they wore sunscreen, in addition to six distractor questions such as how many days they flossed or exercised. Finally, participants reported how many days over the next 7 days they intended to use sunscreen.
Experimental persuasion task
The persuasion task consisted of 32 different persuasive messages about using sunscreen. These messages were divided into four conditions, such that the messages were framed to describe either the benefits of using sunscreen (‘Gain’ messages), the risks of not using sunscreen (‘Loss’), how to use sunscreen appropriately (‘How’) and neutral facts about sunscreen formulation (‘Fact’). Examples of messages in each of these conditions can be found in Table 1. Participants read each message one at a time on a computer screen, while a pre-recorded voice read the message aloud at the same time. Each message was presented for 16–20 s, and appeared in a pseudo-random order such that one message from each condition was presented in random order within a block, and eight blocks total of that same ordering was shown to the participant. A jittered rest period separated each message, showing just a small cross hair on the screen. After four blocks, participants were given a brief break before continuing on to the final four blocks when they were ready. The entire task lasted ∼18 min.
Table 1.
Examples of the four types of messages shown to participants about using sunscreen
| Message type | Example |
|---|---|
| Fact | ‘In USA, sunscreen products are regulated as over-the-counter (OTC) drugs by the U.S. Food and Drug Administration (FDA). The FDA has several safety and effectiveness regulations in place that govern the manufacture and marketing of all sunscreen products, including safety data on its ingredients.’ |
| How | ‘Apply liberally and evenly to all exposed skin. The average adult in a bathing suit should use approximately one ounce of sunscreen per application. Not using enough will reduce the product’s SPF and the protection you get. Be sure to cover often-missed spots: lips, ears, around eyes, neck, scalp if hair is thinning, hands and feet.’ |
| Gain | ‘Daily application of broad spectrum sunscreen with SPF 15 or higher has been clinically demonstrated to keep skin looking younger, more elastic and healthier. Maintaining good habits about using sunscreen is crucial for having beautiful skin for years to come, that not only looks better but is more likely to remain healthy.’ |
| Loss | ‘Studies have found that inconsistent use of sunscreen is associated with a number of skin issues. These include, but are not limited to, wrinkling, sagging, splotchy, leathery, uneven skin. To avoid these issues, you should apply broad-spectrum sunscreen with SPF 15 or higher to any and all skin that will be exposed to the sun.’ |
Post-study questionnaire
After the experimental task, participants again reported how many days and how many times over the next 7 days they intended to use sunscreen. Participants also answered questions about how confident they were in their ability to use sunscreen more often, their beliefs on the potential benefits of increasing sunscreen use and the persuasiveness of the messages they viewed as well as their openness to persuasive messaging in general. These ratings were provided on a sliding scale between 0 and 100.
One-week follow-up questionnaire
Eight days after completing the in-lab session, participants were emailed a follow-up questionnaire and were asked to report on their sunscreen use over the past 7 days as well as their intention to use sunscreen over the next 7 days, in terms of number of days they used sunscreen over the previous week and number of days per week they now intend to use sunscreen in general. Before leaving the lab after the scanning session, participants consented to being contacted again for general research inquiries, but were not expecting to be re-contacted about their sunscreen use specifically.
fNIRS neural data acquisition
Neural data during the persuasion task was measured using near infrared spectroscopy (fNIRS). The fNIRS system used in this study was the fNIR Imager 1000 from fNIR Devices (fnirdevices.com). This system measured the relative changes in oxygenated hemoglobin (HbO) and deoxygenated hemoglobin (HbR) in the prefrontal cortex while participants proceeded through the experimental persuasion task, using near infrared wavelengths of 730 and 850 nm. A diagram of the source and detector optode layout can be seen in Figure 1. This layout is fixed within a semi-rigid headband device, so the optode spacing of 3 cm was standardized across all participants. To ensure that the headband was positioned in the same place for each participant, a small indicator line was located in the middle of the bottom edge of the headband, and the headband was secured to the head so that this indicator line rested just above the nasion point (Figure 1A). This provided ample coverage of the medial, ventromedial and lateral prefrontal cortices (Figure 1B).
Fig. 1.

(A) Placement of the fNIRS headband device on the participant’s head. Positioning across participants was standardized using the 10–20 external landmark system. (B) Approximate location of each channel of data (numbered), projected downward onto the surface of the MNI standard brain cortex from the 10–20 positions.
After securing the fNIRS headband to participants’ heads, experimenters checked the signal quality coming from each data channel by running a test data acquisition session in COBI Studio acquisition software (BIOPAC Systems, Inc.). If the measured intensity of light signal received by detector optodes was >3500 V or <200 V, efforts were made to bring the signal into the acceptable range. This includes changing the gain level of the detectors, tightening or loosening the headband on the participant’s head and pinning any hair out of the way of the optodes. Once all possible improvements were made to the signal strength, the experimenters then initiated the data recording session in the acquisition software and turned off the lights in the experimenting room to reduce ambient light noise.
Procedure
The procedure for this study involved two sessions—one in the lab and one a week later online. In the first week, upon arrival, participants were given information about the study and asked to provide informed consent. Then, participants filled out the pre-study questionnaire in privacy. After completing the questionnaire, experimenters attached the fNIRS headband to their head, checked and adjusted signal levels, initiated fNIRS data acquisition and then began the experimental persuasion task. At this point experimenters then turned out the lights and left the room, so that participants could read the experimental messages in private for the entirety of the task. Once this task was finished, experimenters returned to the room to turn on the lights, stop neural data acquisition, remove the fNIRS headband and open the post-study questionnaire for participants to fill out before again leaving the room. As soon as participants indicated that they were finished with this questionnaire, they were paid for their participation in this session and allowed to leave.
One week after the laboratory session, participants were e-mailed a link to the follow-up questionnaire in order to gauge their sunscreen use and intentions over time. If participants completed this survey, they were invited to return to the lab to receive payment for this final part.
fNIRS data analysis
Neural fNIRS data were first analysed for signal-to-noise ratio to determine its usability. If the amplitude of the light intensity in a channel fell <500 V or >4200 V, indicating that a good connection between the scalp and optode was not achieved, then that channel of data was removed. If more than half of a participant’s data channels were over or under saturated in this way, then that entire participant was removed from analysis. Thirteen participants were removed from analysis for this reason, and the majority of other participants had at least one channel removed.
Raw light intensity fNIRS data were then preprocessed in nirsLAB (http://nirx.net/nirslab-1/) using a band-pass filter of 0.01–0.2 Hz to remove cardiac fluctuation and slow signal drift. Motion artifacts were also identified and removed if they took the form of spikes or discontinuities that exceeded 5 s.d. from the variance of the rest of the data. Light intensity values were then converted to HbO and HbR hemoglobin concentration using the Modified Beer Lambert Law.
After converting the data to hemoglobin concentration, just the HbO chromophore was chosen for analysis. While HbR changes tend to be more spatially specific (Franceschini et al., 2000), HbO seems to have a relatively stronger correlation with the fMRI BOLD signal in the prefrontal cortex during task engagement (Cui et al., 2011), a comparatively stronger signal amplitude (Hoge et al., 2005) and slightly better signal-to-noise ratio (Strangman et al., 2002). Because the neural areas of interest in this study are larger cortical divisions, and the experimental goal is to investigate the ease of replicating prior fMRI research, HbO was the preferred signal to investigate.
Finally, first level statistical analysis was conducted by modeling the recorded brain data with a convolution of the message condition design matrix and a canonical HRF. Pre-coloring was used to account for serial correlations, as pre-coloring tends to estimate temporal correlation in fNIRS data better than pre-whitening (Ye et al., 2009). The resulting betas from this estimation step were then used to predict sunscreen behavior 1 week after the scan. Spatial localization of these effects were determined through a built-in nirsLAB process to convert 10–20 coordinate locations to MNI space.
Results
Behavioral results
Sunscreen use behavior and intentions were recorded in number of days per week that sunscreen was used/intended to be used. The distribution of this data was non-normal with clustering around both response boundaries, so a 10 000 iteration permutation test was used to shuffle the pre/post-label of paired scores or the user/non-user label of difference scores, then generate an experimental null distribution against which we tested our sample means without the assumption of normality. Table 2 reports the sample means and the significance of the pre-post differences. There was a significant increase in self-reported sunscreen use between pre-scan and 1 week after the scan for all participants (P < 0.001, Cohen’s d = 0.540). There was also a significant increase in intention to use sunscreen, both between pre-scan and post-scan (P < 0.001, d = 0.825) and between pre-scan and 1 week after the scan (P < 0.001, d = 0.688). Within user and non-user groups, these changes were also significant. Comparing pre-post sunscreen use difference scores between previous users and non-users of sunscreen revealed that non-users increased their sunscreen use significantly more than users (P = 0.0196, d = 0.586), although users still used sunscreen significantly more than non-users did at the 1 week follow-up (P < 0.001, d = 1.574). Non-users also had a larger increase in intention pre-post (P = 0.001, d = 1.078) and pre-week later (P = 0.001, d = 1.487). Users still intended to use sunscreen more than non-users did, both at post-scan (P = 0.001, d = 1.320) and the follow-up (P < 0.001, d = 1.359). Further breakdown of the behavioral data can be found in the Supplementary data.
Table 2.
Means and standard deviations (in parentheses) of behavioral results
| Sunscreen use |
Intention to use sunscreen |
||||
|---|---|---|---|---|---|
| Pre-scan | 1 week later | Pre-scan | Post-scan | 1 week later | |
| Everyone | 2.000 (2.797) | 3.203***(2.837) | 2.884 (3.183) | 5.087***(2.672) | 4.478***(2.769) |
| Users | 4.759 (2.325) | 5.241 (2.132) | 5.966 (2.009) | 6.724*(0.702) | 6.241 (1.504) |
| Non-users | 0 (0) | 1.725***(2.331) | 0.650 (1.610) | 3.900***(2.942) | 3.200***(2.785) |
Scores represent number of times a week participants intended to or engaged in sunscreen use. Bolded numbers signify a significant difference between scores relative to an experimental null created via 10 000 iteration permutation test.
P < 0.05,
P < 0.01,
P < 0.001.
fNIRS results
Relationship between mPFC activity and sunscreen use
To analyse whether or not mPFC activity predicted sunscreen behavior, Vezich et al. (2017) contrasted gain with fact messages as well as gain with loss messages and then used the resulting contrasts to predict sunscreen behavior (minus intention) in a correlation analysis. In this study, we chose to perform a binomial regression analysis controlling for intentions instead because that would better serve the bimodality of the behavior variable distribution. When we performed this test for the NIRS data channels in the mPFC region (lower half of Brodmann’s area 10), removing outlier data points for having extreme influence on the test results (DF Beta > 1), the results were similar to those seen in Vezich et al. Specifically, the gain > fact parameters in the channels over the right and left mPFC significantly predicted sunscreen use behavior, over and above the effect of self-reported intention, and accounted for significantly more variance than post-scan intentions alone [right mPFC: χ2(2, n = 51) = 10.705, P = 0.001, deviance change test D = 95.32, P < 0.001; left mPFC: χ2(2, n = 52) = 11.658, P < 0.001, deviance change test D = 94.03, P < 0.001] (Figure 2). McFadden’s R2 calculations showed that right mPFC activity accounted for 28.5% of the remaining variance in future behavior and left mPFC accounted for 28.1% of the remaining variance, over and above what participants themselves predicted in their self-reported intentions. For the gain > loss parameter, the right mPFC significantly predicted sunscreen use behavior over and above intention [χ2(2, n = 53) = 8.215, P = 0.004, deviance change test D = 71.93, P < 0.001, McFadden’s R2 = 0.215].
Fig. 2.
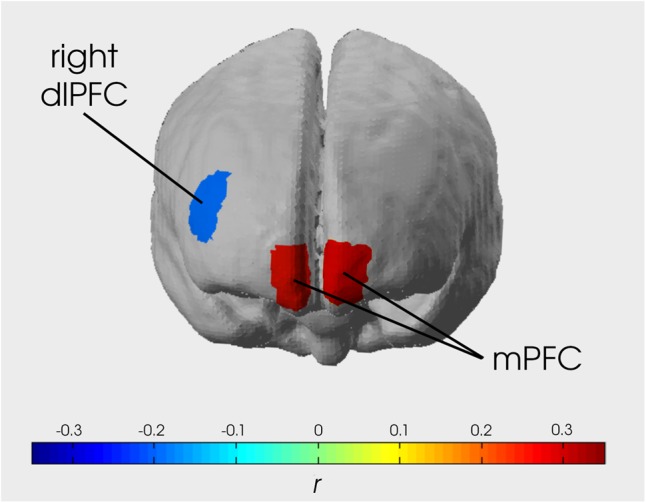
Image showing strength of correlation between activity in each significant data channel and sunscreen use behavior 1 week after the imaging session. Right and left mPFC were correlated r = 0.314 and 0.296, respectively, and right dlPFC was correlated r = −0.205. In a binomial regression model, right and left mPFC showed a significant positive association with future behavior over and above behavioral intentions [χ2(2, n = 51) = 31.093, P < 0.001, ΔR2 = 0.344; and χ2(2, n = 53) = 16.502, P < 0.001, ΔR2 = 0.261, respectively]. Right dlPFC was also significantly associated with future behavior [χ2(41) = 8.318, P = 0.0039, ΔR2 = 0.348]. Including both mPFC and right dlPFC in the model resulted in a better prediction than either region alone (Δdeviance P <0.001).
Alternatively, if we contrast all persuasive messages (gain/loss/how conditions) with neural baseline like some other fMRI persuasion studies do (Falk et al., 2010, 2011), we also get significant results for both the right and left mPFC [right mPFC: χ2(2, n = 51) = 31.093, P < 0.001, deviance change test D = 115.05, P <0.001, McFadden’s R2 = 0.344; left mPFC: χ2(2, n = 53) = 16.502, P < 0.001, deviance change test D = 87.37, P <0.001, McFadden’s R2 = 0.261]. In both mPFC channels, the standardized coefficients for this contrast were greater than those for either gain > fact or gain > loss (Table 3), indicating that for the present dataset this contrast is the best predictor of future behavior. None of these results were affected by number of times rain or fog was recorded in the area in the week after the scan, which might be expected to influence sunscreen use.
Table 3.
Binomial regression results for models testing relationship between mPFC activity and future behavior, with self-reported intention included as a regressor
| Model | Parameter | Unstandardized coefficient | Standard error | Standardized coefficient | Wald’s χ2 | P value | |
|---|---|---|---|---|---|---|---|
| Gain > fact | |||||||
| Behavior = intention + right mPFC | Intercept | −3.3891 | 0.4681 | 52.410 | <0.001 | ||
| intention | 0.5980 | 0.0771 | 0.8917 | 63.439 | <0.001 | ||
| Channel 8 | 6363.2 | 1944.8 | 0.2375 | 10.705 | 0.001 | ||
| Behavior = intention + left mPFC | Intercept | −3.2799 | 0.4583 | 51.227 | <0.001 | ||
| intention | 0.5948 | 0.0735 | 0.8561 | 65.529 | <0.001 | ||
| Channel 10 | 8085.3 | 2368.0 | 0.2571 | 11.658 | <0.001 | ||
| Gain > loss | |||||||
| Behavior = intention + right mPFC | Intercept | −3.0960 | 0.4107 | 56.834 | <0.001 | ||
| intention | 0.5486 | 0.0655 | 0.8106 | 70.119 | <0.001 | ||
| Channel 8 | 5840.0 | 2037.6 | 0.2047 | 8.215 | 0.004 | ||
| Behavior = intention + left mPFC | Intercept | −3.0650 | 0.4256 | 51.870 | <0.001 | ||
| intention | 0.5374 | 0.0666 | 0.7620 | 65.175 | <0.001 | ||
| Channel 10 | 3142.8 | 1649.8 | 0.1273 | 3.629 | 0.057 | ||
| Gain/loss/how > baseline | |||||||
| Behavior = intention + right mPFC | Intercept | −3.6552 | 0.4855 | 56.674 | <0.001 | ||
| intention | 0.5761 | 0.0747 | 0.8591 | 59.543 | <0.001 | ||
| Channel 8 | 12220.3 | 2191.5 | 0.5610 | 31.093 | <0.001 | ||
| Behavior = intention + left mPFC | Intercept | −3.0077 | 0.4430 | 46.100 | <0.001 | ||
| intention | 0.5261 | 0.0698 | 0.7567 | 56.843 | <0.001 | ||
| Channel 10 | 7430.7 | 1829.2 | 0.3753 | 16.502 | <0.001 | ||
Coefficient values are reported in log odds, with unstandardized (relative to unit increase in predictors) and standardized (relative to standard deviation increase in predictors) values included.
Message and participant characteristics results
We proceeded with the following analyses using only the baseline contrast, as it produced the strongest effect in the first analysis and the message/participant characteristic and the right dlPFC hypotheses are less well replicated. Based on an analysis of variance of average activity levels in the mPFC channels for gain, loss and fact messages, there were no significant differences. This is a notable deviation from the original study. There were also no significant differences between average activity for gain and fact messages, or loss and fact messages.
Consistent with the prior study, there was a difference between sunscreen users and non-users in the right mPFC, such that sunscreen non-users had a stronger relationship between activity in the right mPFC and behavior than sunscreen users did (Table 4; Figure 3). This was revealed by a significant interaction term between user status and NIRS data channel in a binomial regression model [χ2(4, n = 53) = 4.679, P = 0.0305]. Investigating the interaction showed that non-users had a significant association between mPFC activity and future behavior [χ2(2, n = 30) = 11.36, P < 0.001, McFadden’s R2 = 0.281] while users did not [χ2(2, n = 23) = 1.22, P = 0.270, McFadden’s R2 = 0.179], replicating Vezich et al.’s findings.
Table 4.
Binomial regression results for models in users and non-users of sunscreen, in terms of the relationship between mPFC activity and future behavior
| Model | Parameter | Unstandardized coefficient | Standard error | Standardized coefficient | Wald’s χ2 | P value | |
|---|---|---|---|---|---|---|---|
| Gain/loss/how > baseline | |||||||
| Behavior = intention + user right mPFC | Intercept | −0.0304 | 2.1650 | 0.0002 | 0.989 | ||
| intention | 0.1578 | 0.3156 | 0.0492 | 0.250 | 0.617 | ||
| channel 8 | 2162.1 | 1959.2 | 0.1133 | 1.218 | 0.270 | ||
| Behavior = Intention + non-user right mPFC | Intercept | −3.2719 | 0.4901 | 44.564 | <0.001 | ||
| intention | 0.4032 | 0.0794 | 0.6405 | 25.809 | <0.001 | ||
| channel 8 | 10249.0 | 3040.5 | 0.5064 | 11.363 | <0.001 | ||
Coefficient values are reported in log odds, with unstandardized (relative to unit increase in predictors) and standardized (relative to standard deviation increase in predictors) values included.
Fig. 3.
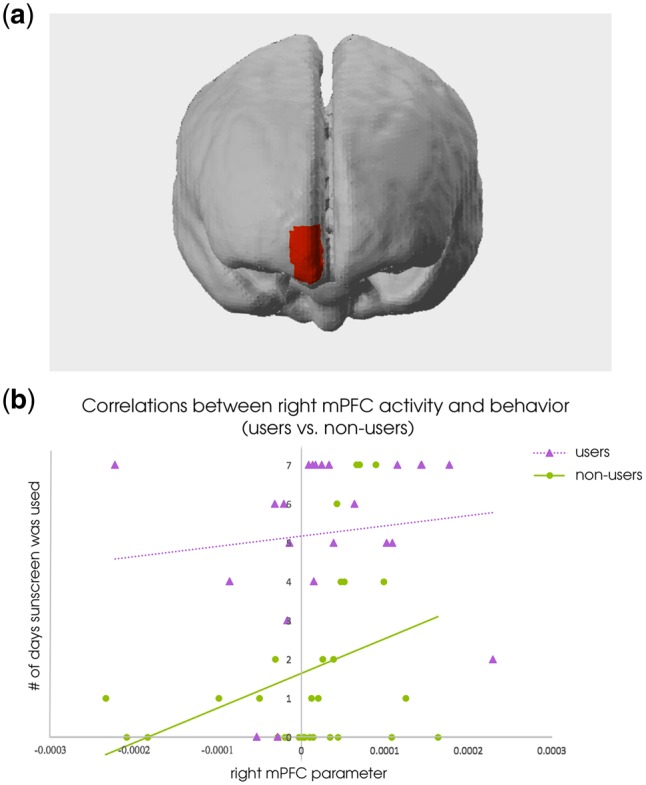
There was an interaction effect in the right mPFC, such that previous non-users of sunscreen showed a significant relationship between neural activity and future behavior while users did not. (A) The spatial location of this interaction. (B) Right mPFC parameter plotted with number of days sunscreen was used post-scan, separated into user/non-user distinction. Linear trend lines in each group are included for ease of visualizing the group difference.
Relationship between right dlPFC and persuasion
To evaluate how well right dlPFC could predict behavior change, we again ran a binomial regression predicting sunscreen use behavior from activity in the right dlPFC for gain, loss and how messages, incorporating post-scan sunscreen intentions as a regressor. There was a significant negative association between right dlPFC activity and sunscreen use behavior 1 week after the experiment [χ2(2, n = 44) = 8.318, P = 0.004, McFadden’s R2 = 0.348] (Table 5; Figure 2). A deviance change test showed that including this region also significantly improved model fit over and above the effect of self-reported intentions alone (D = 178.814, P < 0.001).
Table 5.
Binomial regression results for model testing relationship between right dlPFC activity and future behavior, as well as full model including both right dlPFC and bilateral mPFC
| Model | Parameter | Unstandardized coefficient | Standard error | Standardized coefficient | Wald’s χ2 | P value | |
|---|---|---|---|---|---|---|---|
| Gain/loss/how > baseline | |||||||
| Behavior = intention +right dlPFC | Intercept | −2.976 | 0.4691 | 40.242 | <0.001 | ||
| Intention | 0.4806 | 0.0756 | 0.7076 | 40.414 | <0.001 | ||
| Channel 1 | −3741.0 | 1297.1 | −0.246 | 8.318 | 0.004 | ||
| Behavior = intention + right dlPFC + right mPFC + left mPFC | Intercept | −2.967 | 0.5763 | 26.499 | <0.001 | ||
| Intention | 0.446 | 0.0909 | 0.650 | 24.110 | <0.001 | ||
| Channel 1 | −7877.9 | 2556.4 | −0.486 | 9.496 | 0.002 | ||
| Channel 8 | 28 130.6 | 5178.2 | 1.396 | 29.512 | <0.001 | ||
| Channel 10 | −17 290.9 | 4607.0 | −0.914 | 14.087 | <0.001 | ||
Coefficient values are reported in log odds, with unstandardized (relative to unit increase in predictors) and standardized (relative to standard deviation increase in predictors) values included.
Further, incorporating bilateral mPFC and right dlPFC into the model all at once created a better model fit than either of those regions alone. The deviance change test over and above just mPFC was D = 151.144, P < 0.001, and over and above right dlPFC was D = 121.404, P < 0.001. McFadden’s R2 showed that both regions accounted for an additional 58.46% of the variance beyond self-reported intentions alone.
It is worth noting, however, that because this model was composed of three separate data channels, missing data in any of these channels due to poor optode-scalp contact would result in a list-wise deletion of that participant’s data. Thus, out of 81 subjects that completed the study, only 27 had complete enough data for model inclusion. Thus, this reported effect size may be an inflated estimate.
Discussion
The immediate goal of this investigation was to determine whether previously observed persuasion effects examined with fMRI would replicate with fNIRS. The larger goal was to determine whether fNIRS is a useful tool for the study of persuasion that in the future can be used in novel contexts that cannot easily be studied with fMRI.
Two out of three of the original fMRI results were repeated in this study. Specifically, greater mPFC activity during persuasive messages about sunscreen use positively predicted future sunscreen use over and above self-reported intentions. There was also a difference between users and non-users of sunscreen, such that previous non-users’ mPFC activity significantly predicted future behavior, but users’ mPFC activity did not. Both of these findings were reported in Vezich et al., and our mPFC results had a similar effect size to previous fMRI findings (Falk et al., 2010, 2011).
We also observed that right dlPFC activity during persuasive messages about sunscreen use was negatively associated with future sunscreen use, and information about the activity in this area paired with information from the mPFC offers better behavioral prediction than either area alone. The inspiration for examining this region of interest came from past studies where right dlPFC activity was associated with persuasion resistance and explicit counterarguing. While this study did not test whether counterarguing was the specific psychological phenomenon responsible for the dlPFC-behavior link, the results indicate that activity in this area is likely associated with some process that inhibits persuasion. The results also suggest that this process is somewhat independent from the self-integration occurring with mPFC activity, due to the model improvement that occurred when both areas were taken into account. Future research can thus examine in more detail the specific psychological mechanism responsible for activity in the right dlPFC and build a comprehensive account of the real time internal processes leading to persuasion and behavior change.
However, we did not find any activation differences between different kinds of persuasive messages. This is in contrast with the original study, which identified greater neural activation on an average for gain over fact and loss messages. In addition, the successful replication results above are the result of using a different neural activity parameter than in Vezich et al. (all persuasive messages over baseline, vs gain > fact and gain > loss). The original contrasts, gain > loss and gain > fact, did not replicate the user/non-user interaction and were weaker predictors of mPFC activity for the whole group than the baseline contrast was. It is not immediately clear why the different types of persuasive messages in this study did not produce differential neural responses. Perhaps it is because the original study had less statistical power than this one, making the effect estimates more unstable, and that any true within-subject difference between these message types is a relatively small effect that cannot be identified with fNIRS, which has a lower signal-to-noise ratio than fMRI. Our alternative approach of contrasting average message activation with baseline has precedent in previous neuroimaging studies of persuasive messaging (Falk et al., 2010, 2011, 2016; Cooper et al., 2015), so we believe our results still show a meaningful relationship between neural processing of persuasive information and downstream behavior. Further, it is useful to know what analysis approach is more appropriate for fNIRS specifically, for the purpose of future persuasion research efforts in this modality.
At the same time, a number of issues in this study limit how extensively the results can be interpreted. Firstly, there was quite a bit of neural data that were removed from analysis because of poor signal-to-noise. Twelve participants-worth of data had to be removed out of 81 total, and the majority of those remaining participants had at least one channel of data removed for poor quality. This did not severely limit our power in the analysis of mPFC activity, because the relative affordability of fNIRS allowed us to recruit many more subjects than in a typical fMRI experiment. It did however limit our power in the analysis of our more complex model that included both the mPFC and rdlPFC, because participants had to be excluded from this analysis if they were missing data in any of three different channels. This data loss is also a concern for possible applications of this approach that would aim to design personalized persuasive messages based on one person’s neural activity. The cause of this data loss is largely due to the specific fNIRS technology that we used. While the Biopac unit is one of the most affordable fNIRS units available, the rigidity of its headband device meant that accommodating variation in participants’ head shapes was difficult. Additionally, the flat and wide design of the optodes meant that light could not reach the scalp if there was any hair in the way. More advanced fNIRS units are now available that better address these issues, so data loss should be much less of a problem in the future. Further direct tests of the new technology’s capabilities to replicate fMRI findings should be performed to ensure that design improvements do solve the data loss issue and to identify what, if any, differences in effect sizes are possible to detect.
Taken together, these results provide evidence that fNIRS can replicate the reliable finding that signal from the mPFC during passive viewing of persuasive messages predicts later behavior. It can also distinguish between types of audience members, though not types of persuasive content framing in this particular study. This suggests that, while perhaps not as reliable for detecting small neurophysiological effects as fMRI, fNIRS can provide meaningful predictions about behavioral outcomes in response to persuasive messaging. This is especially valuable for use cases where fMRI is not practical or possible—for example when equipment must be transported to access remote populations; when large study samples are required with limited research budget or in applied research designs where participants need to move in and interact with a more ecologically valid environment than the fMRI scanner. Thus, the results here do indicate that fNIRS can be used toward the service of persuasion research, so long as research designs account for the identified limitations.
Supplementary data
Supplementary data are available at SCAN online.
Funding
This work was supported by the Minerva Initiative from the US Department of Defense (13RSA281, PI: M.D.L.).
Conflict of interest. None declared.
Supplementary Material
References
- Berns G.S., Moore S.E. (2012). A neural predictor of cultural popularity. Journal of Consumer Psychology, 22(1), 154–60. [Google Scholar]
- Chua H.F., Ho S.S., Jasinska A.J., et al. (2011). Self-related neural response to tailored smoking-cessation messages predicts quitting. Nature Neuroscience, 14(4), 426–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooper N., Bassett D.S., Falk E.B. (2017). Coherent activity between brain regions that code for value is linked to the malleability of human behavior. Scientific Reports, 7, 43250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooper N., Tompson S., O’Donnell M.B., Falk E.B. (2015). Brain activity in self- and value-related regions in response to online antismoking messages predicts behavior change. Journal of Media Psychology, 27(3), 93–108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cui X., Bray S., Bryant D.M., Glover G.H., Reiss A.L. (2011). A quantitative comparison of NIRS and fMRI across multiple cognitive tasks. NeuroImage, 54(4), 2808–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falk E.B., Berkman E.T., Lieberman M.D. (2012). From neural responses to population behavior: neural focus group predicts population-level media effects. Psychological Science, 23(5), 439–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falk E.B., Berkman E.T., Mann T., Harrison B., Lieberman M.D. (2010a). Predicting persuasion-induced behavior change from the brain. The Journal of Neuroscience, 30(25), 8421–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falk E.B., Berkman E.T., Whalen D., Lieberman M.D. (2011). Neural activity during health messaging predicts reudctions in smoking above and beyond self-report. Health Psychology, 30(2), 177–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falk E.B., Cascio C.N., Coronel J.C. (2015a). Neural prediction of communication-relevant outcomes. Communication Methods and Measures, 9(1-2), 30–54. [Google Scholar]
- Falk E.B., Morelli S.A., Welborn B.L., Dambacher K., Lieberman M.D. (2013). Creating buzz: the neural correlates of effective message propagation. Psychological Science, 24(7), 1234–42. [DOI] [PubMed] [Google Scholar]
- Falk E.B., O'Donnell M.B., Cascio C.N., et al. (2015b). Self-affirmation alters the brain’s response to health messages and subsequent behavior change. Proceedings of the National Academy of Sciences, 112(7), 1977–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falk E.B., O’Donnell M.B., Tompson S., et al. (2016). Functional brain imaging predicts public health campaign success. Social Cognitive and Affective Neuroscience, 11(2), 204–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falk E.B., Rameson L., Berkman E.T., et al. (2010b). The neural correlates of persuasion: a common network across cultures and media. Journal of Cognitive Neuroscience, 22(11), 2447–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrari M., Mottola L., Quaresima V. (2004). Principles, techniques, and limitations of near infrared spectroscopy. Canadian Journal of Applied Physiology, 29(4), 463–87. [DOI] [PubMed] [Google Scholar]
- Ferrari M., Quaresima V. (2012). A brief review on the history of human functional near-infrared spectroscopy (fNIRS) development and fields of application. NeuroImage, 63(2), 921–35. [DOI] [PubMed] [Google Scholar]
- Fogg B.J. (2009). The behavior grid: 35 ways behavior can change In: Chatterjee S., Dev P., editors. Proceedings of the 4th International Conference on Persuasive Technology .Claremont, CA, 26–29 April, New York: ACM, 42–6. [Google Scholar]
- Franceschini M.A., Toronov V., Filiaci M.E., Gratton E., Fantini S. (2000). On-line optical imaging of the human brain with 160-ms temporal resolution. Optics Express, 6(3), 49–57. [DOI] [PubMed] [Google Scholar]
- Hoge R.D., Franceschini M.A., Covolan R.J., Huppert T., Mandeville J.B., Boas D.A. (2005). Simultaneous recording of task-induced changes in blood oxygenation, volume, and flow using diffuse optical imaging and arterial spin-labeling MRI. NeuroImage, 25(3), 701–7. [DOI] [PubMed] [Google Scholar]
- Kahneman D., Tversky A. (1979). Prospect theory: an analysis of decision under risk. Econometrica, 47(2), 263–91. [Google Scholar]
- Kato J., Ide H., Kabashima I., Kadota H., Takano K., Kansaku K. (2009). Neural correlates of attitude change following positive and negative advertisements. Frontiers in Behavioral Neuroscience, 3, 6.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miilunpalo S., Nupponen R., Laitakari J., Marttila J., Paronen O. (2000). Stages of change in two modes of health-enhancing physical activity: methodological aspects and promotional implications. Health Education Research, 15(4), 435–48. [DOI] [PubMed] [Google Scholar]
- Montague P.R., Berns G.S., Cohen J.D., et al. (2002). Hyperscanning: simultaneous fMRI during linked social interactions. NeuroImage, 16(4), 1159–64. [DOI] [PubMed] [Google Scholar]
- Ramsay I.S., Yzer M.C., Luciana M., Vohs K.D., MacDonald A.W.I.I.I. (2013). Affective and executive network processing associated with persuasive antidrug messages. Journal of Cognitive Neuroscience, 25(7), 1136–47. [DOI] [PubMed] [Google Scholar]
- Riddle P.J. Jr, Newman-Norlund R.D., Baer J., Thrasher J.F. (2016). Neural response to pictorial health warning labels can predict smoking behavioral change. Social Cognitive and Affective Neuroscience, 11(11), 1802–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saito D.N., Tanabe H.C., Izuma K., et al. (2010). “Stay tuned”: Inter-individual neural synchronization during mutual gaze and joint attention. Frontiers in Integrative Neuroscience, 4, 127.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salovey P., Wegener D.T. (2003). Communicating about health: message framing, persuasion, and health behavior In: Suls J., Wallston K.A., editors. Social Psychological Foundations of Health and Illness .Malden, MA: Blackwell Publishing, 54–81. [Google Scholar]
- Schilbach L., Wohlschlaeger A.M., Kraemer N.C., et al. (2006). Being with virtual others: neural correlates of social interaction. Neuropsychologia, 44(5), 718–30. [DOI] [PubMed] [Google Scholar]
- Scholkmann F., Kleiser S., Metz A.J., et al. (2014). A review on continuous wave functional near-infrared spectroscopy and imaging instrumentation and methodology. NeuroImage, 85, 6–27. [DOI] [PubMed] [Google Scholar]
- Strangman G., Culver J.P., Thompson J.H., Boas D.A. (2002). A quantitative comparison of simultaneous BOLD fMRI and NIRS recordings during functional brain activation. NeuroImage, 17(2), 719–31. [PubMed] [Google Scholar]
- Vezich I.S., Katzman P.L., Ames D.L., Falk E.B., Lieberman M.D. (2017). Modulating the neural bases of persuasion: why/how, gain/loss, and users/non-users. Social Cognitive and Affective Neuroscience, 12(2), 283–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weber R., Huskey R., Mangus M., Westcott-Baker A., Turner B.O. (2015). Neural predictors of message effectiveness during counterarguing in antidrug campaigns. Communication Monographs, 82(1), 4–30. [Google Scholar]
- Weinstein N.D., Rothman A.J., Sutton S.R. (1998). Stage theories of health behavior: conceptual and methodological issues. Health Psychology, 17(3), 290–9. [DOI] [PubMed] [Google Scholar]
- Wilms M., Schilbach L., Pfeiffer U., Bente G., Fink G.R., Vogeley K. (2010). It’s in your eyes – using gaze-contingent stimuli to create truly interactive paradigms for social cognitive and affective neuroscience. Social Cognitive and Affective Neuroscience, 5(1), 98–107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ye J., C., Tak S., Jang K., Jung J., E., Jang J. (2009). NIRS-SPM: statistical parametric mapping for near-infrared spectroscopy. NeuroImage, 44(2), 428–47. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.