


Abstract
Plasminogen activator inhibitor type 2 (PAI-2) is a serine protease inhibitor traditionally regarded as a regulator of fibrinolysis and extracellular matrix degradation. More recently, PAI-2 has been implicated in diverse processes such as keratinocyte differentiation, cell death and viral pathogenesis. The PAI-2 promoter tightly regulates PAI-2 gene expression in a cell-specific manner and this control is mediated, in part, by the upstream silencer element, PAUSE-1. Here we have defined PAUSE-1 and investigated its activity as a silencer. A series of mutations were generated within the PAUSE-1 element and analysed for transcription factor binding and transcriptional silencing activity. These studies have defined the minimal functional PAUSE-1 element as TCTNxAGAN3T4, where x = 0, 2 or 4. Examination of related elements present in other promoters, such as the human IFNβ promoter, suggests that PAUSE-1 is a member of a family of universal silencers with the consensus sequence TCTNxAGA. UV crosslinking analyses determined that the PAUSE-1 binding protein was ∼67 kDa. Insertion of PAUSE-1 into the heterologous (SV40) or the minimal PAI-2 promoters silenced transcription by 2.5-fold. These data show that PAUSE-1 acts as a powerful silencer of PAI-2 gene transcription and is likely to be important in the silencing of other genes as well.
INTRODUCTION
Plasminogen activator inhibitor type 2 (PAI-2) was one of the first identified members of the structurally conserved but functionally diverse family of serine protease inhibitors (serpins) known as ovalbumin-like serpins or ov-serpins (1). While ov-serpins may inhibit serine protease activities, they have clearly evolved additional functions and as yet unidentified intracellular targets (2–6). PAI-2 was originally identified as an inhibitor of urokinase-plasminogen activator (uPA) (7,8) and as such PAI-2 has been shown to modulate uPA-mediated adhesion and migration as well as uPA-dependent lysis of the extracellular matrix (8). Furthermore, PAI-2-transfected melanoma cells demonstrate an impaired degradation of extracellular matrix and reduced metastasis (9). In addition to its ability to inhibit uPA-mediated proteolysis, PAI-2 has been implicated in protecting cells from inappropriately timed apoptosis and cell death (2,10–13), potentially a critical role in activated haematopoietic cells and differentiating keratinocytes.
Under physiological conditions, PAI-2 expression is limited to a select number of cell types, which include differentiated keratinocytes, activated monocytes and macrophages, placental trophoblasts and some tumour cell lines (8). However, high levels of PAI-2 gene expression are rapidly achieved in a cell-specific manner upon stimulation with several factors, including phorbol esters (PMA), lipopolysaccharide, tumour necrosis factor-α, retinoic acid, lipoprotein (a), interferon-γ and viral RNA (8,10,14,15). Several elements, of varying functional relevance, have been identified in the PAI-2 promoter (16–21). It is clear that PAI-2 gene expression is tightly regulated on several levels, potentially via powerful transcriptional repression, which in turn is regulated by specific transcriptional activators.
Gene expression may be actively repressed through negative regulatory genetic elements called ‘silencers’ (22). Two classes of silencer exist, the ‘silencer element’ and the ‘negative regulatory element’ (22). Previously we identified a silencer in the PAI-2 promoter and experimentally defined it as a ‘silencer element’ or classical silencer. This silencer down-regulated PAI-2 gene expression in vitro and was called PAUSE-1, for PAI-2 upstream silencer element 1 (23). PAUSE-1 was originally identified within a 300 bp PAI-2 promoter fragment that retained silencer activity in functional reporter gene assays using both HeLa cells, which do not express PAI-2, and U937 cells, which can be induced to express PAI-2 in response to PMA (23). A minimal 28 bp silencer element was located within a palindromic XbaI restriction enzyme site, at a position 1.832 kb upstream of the transcription initiation site (23). This element was shown to be a ‘silencer element’ and was proposed to be a key regulatory element of the PAI-2 promoter (23). Here we have defined the critical nucleotides required for PAUSE-1-mediated repression and characterised the PAUSE-1 binding protein (PAUSE-1 BP) complex. The data suggest that PAUSE-1 is a member of a family of transcriptional regulators which direct an active mechanism of transcriptional repression.
MATERIALS AND METHODS
Cell culture
Cell cultures were maintained in RPMI 1640 medium (Gibco BRL, Gaithersberg, MD) supplemented with 10% foetal calf serum, 25 mM HEPES, 200 µg/ml penicillin and 100 µg/ml streptomycin sulphate and were incubated in a 5% CO2/95% air atmosphere at 37°C. The cell lines were: HeLa (human cervical adenocarcinoma, ATCC no. CCL-2.2); SW480 (human colorectal adenocarcinoma, ATCC no. CCL-228); SW620 (human lymph node metastasis derived from SW480, ATCC no. CCL-227); MCF7 (human mammary gland adenocarcinoma, ATCC no. HTB-22); MCF7M (human lymph node metastasis derived from MCF7); HaCaT (human keratinocyte; 24); MonoMac6 (human macrophage; 25); U937 (human histiocytic lymphoma, ECACC no. 85011440); 293 (adenovirus 5-transformed human kidney, ATCC no. CRL-1573); MM253-3D (human axillary lymph node melanoma metastasis; 26); MM170 (human inguinal lymph node melanoma metastasis; 26); MM229 (human lymph node melanoma metastasis; 26); MM418c1 (human melanoma; 26); MM418c5 (human melanoma; 26); and MM96L (xenograft of human lymph node melanoma metastasis; 26). PMA was added to cultures at 40 ng/ml.
Nuclear extract preparation
Nuclear extracts were prepared from ∼1 × 107 cultured cells as described by Antalis and Godbolt (27). The human hematopoietic cells, U937 and MonoMac6 were lysed using a lysis buffer containing 0.05% NP-40 (Sigma, St Louis, MO), while the remaining cell lines were lysed using a lysis buffer containing 0.15% NP-40. Protein concentrations were determined using the Bio-Rad Protein Microassay (Bio-Rad, Hercules, CA).
Electrophoretic mobility shift assay (EMSA)
Oligonucleotides were synthesised by Bresatec Pty Ltd (Adelaide, Australia). Double-stranded oligonucleotides used as probe or competitor were prepared by annealing each oligonucleotide listed in Table 1 with a corresponding 6mer complementary oligonucleotide and blunt-ending using Klenow DNA polymerase (New England Biolabs, Beverly, MA). Radiolabelled DNA was prepared by incorporating [α-32P]-labelled dCTP (3000 Ci/mmol) (Amersham Pharmacia Biotech, Little Chalfont, UK) and separated from the unincorporated [α-32P]dCTP using a Sephadex G-25 column (NAP-5) (Amersham Pharmacia Biotech) with 10 mM Tris, pH 8.0, 1 mM EDTA (TE) as the mobile phase. The efficiency of radiolabelling was analysed by thin layer chromatography on PEI cellulose (Merck, Darmstadt, Germany), using a separation buffer of 0.75 M KH2PO4.
Table 1. Nucleotide sequences of oligonucleotides used to investigate PAUSE-1 sequence specificity.
 |
1Bold, capital letters represent specific sequence homology to PAUSE-1.
Binding buffers and reaction conditions were optimised for specific PAUSE-1/PAUSE-1 BP protein binding. The reactions contained 2 µl of 10× GIa buffer (100 mM Tris–HCl, pH 8.0, 500 mM NaCl, 100 mM MgCl2, 1 mM EDTA, pH 8.0, 20% glycerol, 10 mM DTT), 2 µg poly(dI·dC) (Boehringer Mannheim, Mannheim, Germany), 5 µg nuclear extract, 2 µl (∼100 c.p.s., ∼100 pg) of 32P-labelled DNA and H2O to 20 µl. Unless otherwise stated DNA–protein binding reactions were carried out for 10 min at room temperature. Competition reactions were performed by the addition of non-radiolabelled double-stranded DNA oligonucleotides at the onset of the binding reaction. Following electrophoresis on a 5% non-denaturing polyacrylamide gel (acrylamide:bis-acrylamide 29:1) in 100 mM Tris–HCl, pH 8.3, 100 mM boric acid, 2 mM EDTA (TBE), the gel was dried under vacuum and subjected to autoradiography.
The stability of the PAUSE-1 BP complex was investigated by incubating HeLa nuclear extracts with radiolabelled PAUSE-1 for 15, 30 and 60 min and 2 and 24 h at both 4 and 25°C. The binding affinity of the PAUSE-1 BP complex was investigated using standard conditions except employing a range of reaction incubation times (from 30 s to 20 min).
UV crosslinking experiments
For UV crosslinking experiments, Br-dUTP (Boehringer Mannheim) was incorporated into the radiolabelled double-stranded oligonucleotide probes during Klenow-mediated end-filling of annealed oligonucleotides. The DNA–protein binding reactions were performed under the standard conditions described above or with the addition of calf thymus DNA (2 µg) as non-specific competitor DNA. The reactions were then subjected to UV light at 312 nm (Gene Pulser; Bio-Rad) for varying times. The covalently linked DNA–protein complexes were separated by SDS–PAGE. The complexes were visualised by autoradiography.
Reporter plasmid construction
Isolation and cloning of an 8.8 kb fragment spanning the PAI-2 promoter to generate the plasmid denoted pBS8.8pro has been described (16). Similarly, engineering of the PAI-2 promoter reporter constructs pCAT5′-5.1 (extending to 5.1 kb 5′ of the transcription initiation site), pCAT5′-1.9 (extending to –1.9 kb and containing the PAUSE-1 element) and pCAT5′-1.7 (extending to –1.7 kb and not containing the PAUSE-1 element) has been described previously (23). The constructs denoted pCAT5′-1.7+PAUSE-1, +M1, +M2, +M3, +M4, +M5, +M6, +M7, +M8, +M9 and +M10 were constructed as follows. Blunt-ended double-stranded oligonucleotides corresponding to the sequences listed in Table 1 were generated as described for EMSAs and phosphorylated using T4 PNK (New England Biolabs). The DNA was ligated into the dephosphorylated (thermostable alkaline phosphatase; Gibco BRL) EcoRV restriction enzyme site of pCAT5′-1.7. Sequence and orientation were verified by DNA sequence analysis using the PAI-2 promoter-specific primer EC-1 (5′-GTG AGA AAG AGT AGC ACC TG-3′).
pCAT Control+PAUSE-1 was constructed by ligating the blunt-ended double-stranded PAUSE-1 oligonucleotide into the end-filled, dephosphorylated BglII restriction site of the pCAT Control vector (Promega, Madison, WI). Sequence and orientation were verified using the pCAT sequencing primer, CAT Primer 3 (5′-CGG ATG AGC ATT CAT CAG GC-3′).
Transient transfection and reporter gene assays
Choloramphenicol acetyltransferase (CAT) reporter plasmids were purified on CsCl gradients and transfected into HeLa and U937 cells by electroporation as previously described (23). The plasmid pCAT Control (Promega), comprising a CAT reporter gene driven by the SV40 promoter and enhancer, was used as the positive control. The promoterless pCAT Basic (Promega) was used as a negative control. Electroporated cells were diluted into 5 ml of medium and allowed to incubate for 18 h in the absence or presence of 40 ng/ml PMA. Cells were harvested and CAT activities were assayed as described (23). CAT activities are presented as % conversion/mg total protein for each construct and represent averages ± SE of a minimum of three separate transfections.
Northern blot analysis
Total RNA was isolated from ∼1 × 107 cells by the guanidinium isothiocyanate method of Chomczynski and Sacchi (28). RNA (10 µg) was electrophoresed on denaturing agarose gels containing 1.1% formaldehyde, transferred to nylon membranes and hybridised as described elsewhere (29). Blots were probed with a plasmid containing PAI-2 cDNA, pBTA447 (7), labelled with [32P]dCTP using the Megaprime DNA labelling system (Amersham Pharmacia Biotech) and purified by chromatography as above. To standardise for loading, the blots were reprobed with a double-stranded oligonucleotide against human 18S rRNA (5′-GAT GGT AGT CGC CGT GCC TAC CAT G-3′) (29).
RESULTS
Definition of the PAUSE-1 binding sequence
The 28 bp PAUSE-1 element (23) contains a palindromic sequence with three repeated pyrimidine dinucleotide (CT) and three repeated purine dinucleotide (AG) motifs centred on a XbaI restriction enzyme site, within the context of 5′ and 3′ flanking sequences (Table 1). EMSA experiments using a synthetic 28 bp double-stranded oligonucleotide have demonstrated one complex indicating the presence of a DNA-binding protein (PAUSE-1 BP) in HeLa cell nuclear extracts (23). To examine the nucleotide specificity of the interaction between PAUSE-1 and the PAUSE-1 BP (PAUSE-1/PAUSE-1 BP) complex, a panel of double-stranded oligonucleotides containing mutations in the PAUSE-1 sequence were synthesised as listed in Table 1. The mutant oligonucleotides M1–M7 and M9 were designed with specific mutations relating to the palindromic XbaI restriction enzyme site within the PAUSE-1 sequence. M8 represented a PAUSE-1-like sequence present in the interferon β (IFNβ) promoter (23) and M10 was based on the PAUSE-1-like Ski-binding element, GTCT/2 (30).
The ability of the mutant oligonucleotides to specifically compete for PAUSE-1 complex binding was investigated by competitive EMSA using HeLa nuclear extracts (Fig. 1A). EMSA analysis using 1000-fold competitor PAUSE-1 resulted (23) in a decrease in the intensity of the PAUSE-1/PAUSE-1 BP complex by ∼95%. This level of competition was also observed using M10, indicating that this oligonucleotide contains elements that compete strongly for PAUSE-1 BP binding. Competition using M1–M3, M5 and M8 resulted in a decrease in PAUSE-1/PAUSE-1 BP complex association of ∼85%. M4, M7 and M9 competed relatively poorly with the PAUSE-1/PAUSE-1 BP complex (∼60%) and M6 failed to compete. These data suggest that a common motif in the mutant oligonucleotides M1–M4, M8 and M10, potentially the TCT and/or AGA motif, play a critical role in PAUSE-1/PAUSE-1 BP association.
Figure 1.

Competition of PAUSE-1/PAUSE-1 BP complex binding by mutant PAUSE-1 oligonucleotides. (A) EMSA using ∼100 pg 28 bp 32P-labelled PAUSE-1 as probe and 5 µg HeLa nuclear extract in GIa buffer. PAUSE-1/PAUSE-1 BP complex binding was competed out, to varying degrees, by addition of ∼100 ng (1000-fold) non-radiolabelled competitor oligonucleotide as designated in Table 1. (B) Direct binding of mutant PAUSE-1 oligonucleotides to the PAUSE-1 BP complex. EMSA using ∼100 pg 32P-labelled PAUSE-1 mutants as designated in Table 1 and 5 µg HeLa nuclear extract in GIa buffer. Bands due to non-specific binding are indicated (NS).
To further investigate the binding characteristics of the mutant oligonucleotides, each of the oligonucleotides was radiolabelled and tested for direct nuclear protein binding by EMSA (Fig. 1B). All mutant oligonucleotides formed complexes of similar electrophoretic mobility to the PAUSE-1/PAUSE-1 BP complex and M9 bound an additional faster migrating complex. M3, M8 and M10 each formed complexes with signal intensities of ∼125% compared to PAUSE-1. The increased efficiency of M3 binding, in which the thymidine repeat had been substituted, may indicate an inhibitory role for this conserved sequence. M1–M4 and M9 formed complexes with reduced relative signal intensities of ∼80%, while M6 and M7 failed to form any DNA–protein complexes. Each of the observed DNA–protein complexes, with the exception of the faster migrating M9 complex, was effectively competed with 1000-fold excess cold competitor PAUSE-1 (data not shown), indicating that the DNA–protein bands likely represent PAUSE-1 BP-mediated complexes. The additional faster migrating protein complex formed by M9 was poorly competed out by competitor PAUSE-1 and is likely to be unrelated to PAUSE-1 BP binding.
These data show that the mutant oligonucleotides M3, M8 and M10 contain DNA binding elements, which promote a strong interaction with the PAUSE-1 BP complex. The mutant oligonucleotides M1–M4 also contain sequences that bind to the PAUSE-1 BP complex but with reduced efficiency. Taken together, these data indicate a consensus DNA binding site for PAUSE-1 BP of TCTNxAGA, where x = 0, 2 or 4. However, a single TCT or AGA motif also retains significant binding affinity for the PAUSE-1 BP complex and the thymidine repeat may also influence binding activity.
Functionality of PAUSE-1 as a silencer
The EMSA data demonstrate the ability of the PAUSE-1 BP complex to bind to its recognition sequence but do not provide information with respect to its functionality in relation to transcription. To investigate this, each mutant oligonucleotide was inserted into a plasmid containing a transcriptionally active PAI-2 promoter and the effects on gene transcription investigated by reporter gene assay. The PAI-2 reporter gene constructs used in these experiments are shown in Figure 2A. Insertion of PAUSE-1 into the PAI-2 promoter construct pCAT5′-1.7 has been demonstrated to restore silencer function in vitro in both HeLa and U937 cells (23). Therefore, the PAUSE-1 oligonucleotides listed in Table 1 were inserted into pCAT5′-1.7 and evaluated for their efficacy in restoring silencer function in the context of the PAI-2 promoter.
Figure 2.

Functional activities of the PAUSE-1 mutant oligonucleotides. (A) Diagram of the CAT reporter gene constructs. X denotes inserted oligonucleotides shown in Table 1. (B) Graphical representation of standardised HeLa and U937 CAT reporter gene assays. The transcriptional activities of the PAI-2 promoter constructs were analysed by CAT reporter gene activity in HeLa cells (yellow), untreated U937 cells (blue) and U937 cells treated with 40 ng/ml PMA (red). The data is presented as % conversion/mg total protein for each construct and are the averages ± SE of a minimum of three separate transfections. The pCAT Control and pCAT Control+PAUSE-1 constructs are represented separately using a different y-axis scale. (C) Graphical representation of the relative level of transcriptional repression. Data from (B) is expressed relative to the CAT reporter gene activity obtained using pCAT5′-1.7, such that the y-axis values represent the relative strength (fold) of silencer activity. Final values represent the average relative repression from HeLa, U937 and PMA-treated U937 cells.
Each construct was assayed for CAT reporter gene activity in HeLa cells and in U937 cells in the presence and absence of PMA. The results are summarised in Figure 2B. The reporter gene activities measured in HeLa cells were generally significantly lower than those measured for U937 cells, possibly reflecting differences in transfection efficiencies between the two cell lines. However, the relative silencer activities of each of the different PAUSE-1 mutants showed a very similar pattern in HeLa and U937 cells. This is represented in Figure 2C, wherein the CAT reporter gene activity is expressed relative to pCAT5′-1.7 as a direct measure of transcriptional repression.
Assay of HeLa cells transiently transfected with pCAT5′-1.9, containing the PAUSE-1 element in its native environment, repressed promoter gene activity ∼3.4-fold relative to the silencer-less construct pCAT5′-1.7 (Fig. 2B and C). Similarly, transient transfection of U937 cells with pCAT5′-1.9 repressed promoter activity ∼3.5-fold relative to pCAT5′-1.7 in both the presence and absence of PMA (Fig. 2B and C). When the 28 bp PAUSE-1 oligonucleotide was inserted into pCAT5′-1.7 (pCAT5′-1.7+PAUSE-1), PAI-2 promoter activity was repressed to a similar extent, ∼2.3-fold in both HeLa and U937 cells and 2.7-fold in PMA-treated U937 cells (Fig. 2C). When inserted into pCAT5′-1.7 in exactly the same context as PAUSE-1, M3 repressed transcription with an equal or better efficiency as PAUSE-1, with 3-fold repression in HeLa cells and ∼2.5-fold in U937 cells. The efficacy of M3 to repress transcriptional activity demonstrates that the thymidine repeat is not essential for silencer activity.
M10 was nearly as effective a silencer as PAUSE-1, consistent with EMSA data demonstrating efficient PAUSE-1 BP-binding activity for this oligonucleotide. The functional efficacy of M10 together with the EMSA data shows that the non-conserved 5′ (GTA TAG GGC) and 3′ (TTT TAG C) flanking sequences are not required for silencer activity. M8, although less effective than PAUSE-1 as a silencer, silences promoter activity at least 1.6-fold, relative to pCAT 5′-1.7. M8 contains the PAUSE-1 consensus nucleotides but, being derived from the IFNβ promoter, has an overall architecture which is markedly different to PAUSE-1. This altered architecture could explain the decrease in silencer activity that might be expected from this oligonucleotide. Therefore, the observed silencer activity of M8 is considered as good supporting evidence for a requirement for the CTC and AGA motifs of the PAUSE-1 consensus sequence. M1, M2 and M5 were only ∼50% as effective as PAUSE-1 in repressing transcription, indicating that the half-palindrome in the context of the flanking PAUSE-1 element has a significantly reduced silencer activity. M4 only showed silencer activity in HeLa cells. Although purely speculative, it is possible that M4 has been mutated in such a way that the cell specificity of the PAUSE-1 silencer is affected. M9 also only weakly repressed promoter activity. M6 and M7 failed to repress transcription when inserted into pCAT5′-1.7, consistent with the absence of critical nucleotides for PAUSE-1 BP complex association in these constructs.
In general, these data support the results of EMSA experiments, demonstrating that M3, M8 and M10 function effectively as PAUSE-1 homologues, both as binding elements for PAUSE-1 BP and as silencers of PAI-2 promoter-mediated transcription.
PAUSE-1-mediated repression of heterologous promoters
When inserted upstream of the SV40 promoter, the 28 bp PAUSE-1 element repressed promoter activity by ∼2-fold in both HeLa cells and U937 cells (Fig. 2B and C). This indicates that while PAUSE-1 can clearly function as an effective silencer when inserted into a heterologous environment (pCAT5′-1.7+PAUSE-1 and pCAT Control+PAUSE-1), it is a better repressor in its native environment (pCAT5′-1.9 and pCAT5′-5.1).
Cellular distribution of PAUSE-1 BP activity
To investigate the cellular distribution of PAUSE-1 BP complex activity, nuclear extracts from a range of cell lines derived from different tissues were examined by EMSA. As shown in Figure 3A, PAUSE-1-binding complexes were detected in all cell lines examined, indicating that PAUSE-1 BPs may be ubiquitously expressed. Intense bands were detected in extracts from HeLa, MM253-3D, MM170, MM418c5 and MM96L cells, indicating that these cells contained the highest PAUSE-1-associated binding activity. A single complex was detected in HeLa, SW480, SW620, MonoMac6, U937, 293, MM170, MM229, MM418c5 and MM96L cells, each having a similar mobility. Two complexes, one with a similar mobility to that detected in HeLa cells and a second faster migrating complex, were visualised in HaCaT, MM253-3D and MM418c1 nuclear extracts. The relative intensities of the two bands differed with each cell type. One complex was observed in both MCF7 and MCF7M cells, neither of which corresponded to the mobility of the complex detected in HeLa cells or to each other. These data suggest that the PAUSE-1 BP complex is widely expressed but complex members may differ between different cells.
Figure 3.

PAUSE-1 BP complex in nuclear extracts from cultured cell lines. (A) EMSA using ∼100 pg 28 bp 32P-labelled PAUSE-1 as probe and 5 µg nuclear extract from 15 cell lines in GIa buffer. Non-specific (NS) bands and free probe are indicated. (B) Expression of PAI-2 mRNA in cultured cell lines. Northern blot analysis of 10 µg total RNA isolated from 15 cultured cells lines and probed with 32P-labelled PAI-2 cDNA. The blot is overexposed to highlight the absence of PAI-2 mRNA in most of the cell lines. The blot was rehybridised with an 18S rRNA probe as a measure of RNA loading.
To investigate whether the presence of the PAUSE-1 BP correlated with PAI-2 mRNA expression, the relative levels of PAI-2 mRNA in each cell line were examined by northern blot. Significant levels of PAI-2 mRNA were detected only in HaCaT and MonoMac6 cells (Fig. 3B). These data demonstrate that the simple presence or absence of PAUSE-1 BP activity does not determine the expression pattern of PAI-2.
Properties of the PAUSE-1 BP complex
The interaction between PAUSE-1 and PAUSE-1 BP was examined by EMSA under a range of binding conditions. The intensity of the PAUSE-1/PAUSE-1 BP complex was not altered by incubation at 25°C for up to 2 h or by incubation at 4°C for up to 24 h (data not shown), suggesting that the PAUSE-1 BP complex is relatively stable. Similarly, the PAUSE-1/PAUSE-1 BP complex was unaffected by reducing the binding reaction time from 20 min to 30 s (data not shown), suggesting that the PAUSE-1 BP complex has a relatively high affinity of interaction for PAUSE-1.
The approximate molecular mass of the PAUSE-1 BP was investigated by UV crosslinking of [α-32P]-Br-dUTP-labelled PAUSE-1 in the presence of HeLa nuclear extracts (Fig. 4). PAUSE-1 specifically crosslinked to a band of 82 kDa. Minor additional bands were also present, however, these were determined to be non-specific as they disappeared in the presence of different non-specific competitor DNA (data not shown). Taking into account the molecular mass of the PAUSE-1 DNA of 15 kDa, these data indicate that the PAUSE-1 BP has a molecular mass of 67 kDa. The intensity of the 67 kDa protein did not increase with increased UV irradiation, suggesting that either the 67 kDa PAUSE-1 BP is present in low abundance or the PAUSE-1/PAUSE-1 BP interaction has a high binding affinity, or both.
Figure 4.
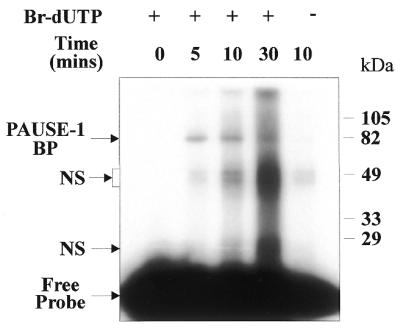
Determination of the molecular mass of PAUSE-1 BP by UV crosslinking. Experiments were performed with ∼100 pg 28 bp 32P-Br-dUTP-labelled PAUSE-1 or 32P-labelled PAUSE-1 in the absence of Br-dUTP (where indicated) and 5 µg HeLa nuclear extract in GIa buffer. Individual reactions were crosslinked for 0, 5, 10 or 30 min at 312 nm. Molecular mass markers are in kDa. Bands due to non-specific binding are indicated (NS). In the absence of Br-dUTP the band intensities were greatly reduced (lane 5).
UV crosslinking studies were also carried out using nuclear extracts from the 15 different cell lines. Figure 5 shows that PAUSE-1 directly binds to a single protein of ∼67 kDa (82 kDa protein + DNA) in each of the cell lines. These data indicate that the PAUSE-1/PAUSE-1 BP complexes of different mobilities observed in EMSAs may represent different complexes in association with the same DNA-binding protein, PAUSE-1 BP. This was confirmed directly by excision of gel slices of bands obtained by EMSA, followed by UV crosslinking and resolution by SDS–PAGE. The data show that shifted bands of different mobilities, including two distinct protein complexes from the same cell line (e.g. MM418c1), crosslinked to the single 67 kDa protein (data not shown).
Figure 5.

The presence of the PAUSE-1 BP in nuclear extracts from cultured cell lines. UV crosslinking experiments were performed using ∼100 pg 28 bp 32P-labelled PAUSE-1 and 5 µg nuclear extract in GIa buffer. Molecular mass markers are in kDa.
DISCUSSION
In this study we have defined the critical nucleotides required for protein binding and functional activity of PAUSE-1 and have characterised the PAUSE-1 BP complex.
Competitive EMSA experiments showed that M3, M8 and M10 competed as efficiently as PAUSE-1 for PAUSE-1 BP binding. Although flanking sequences are known to be crucial in determining the overall functionality of binding elements (22), the lack of 5′ sequence homology between PAUSE-1, M8 and M10 suggested that only the CT and/or AG repeats were essential for PAUSE-1/PAUSE-1 BP complex binding. In support of this, deletion of the CT and AG repeats (M6) resulted in loss of competition for binding to the PAUSE-1/PAUSE-1 BP complex. Competition with M1, M2, M4 and M5 was reduced in comparison to PAUSE-1, but only to ∼80%, showing that they retained a platform for PAUSE-1 BP binding. Confirming that a single TCT or AGA double-stranded site was sufficient to provide a reduced affinity binding site for the PAUSE-1 BP complex, direct binding experiments with M1, M2, M4 and M5 resulted in complexes with ∼80% of the intensity of PAUSE-1. In a double-stranded environment, M1 and M2, as well as M4 and M5, contain similar nucleotide motifs. Therefore, the results obtained using these oligonucleotides only indicate the significance of the double-stranded trinucleotide TCT, not TCT or AGA, per se.
M3 was designed to test the role of the thymidine repeat in the interaction of PAUSE-1 and PAUSE-1 BP. Similar thymidine repeats have been reported to be conserved in other negative regulatory elements containing PAUSE-1-like purine motifs (23). The increased competition of M3 for PAUSE-1 binding and increased efficiency of binding of M3 to the PAUSE-1 BP complex suggest that the presence of the thymidine repeat may reduce the efficiency of PAUSE-1 BP complex binding.
M9 reproducibly competed for PAUSE-1 BP binding approximately half as efficiently as PAUSE-1. M9 was originally designed as a mutant for PAUSE-1 with the central XbaI palindrome being disrupted (23). The XbaI site is destroyed in M9 but CTC and GAG sites are present with a gap of 6 nt between them. Therefore, M9 potentially provides a PAUSE-1-like consensus sequence, which may account for the competition observed with the double-stranded M9 oligonucleotide.
M8 and M10 bound with increased efficiency to the PAUSE-1 BP complex. The thymidine repeat is further downstream from the consensus PAUSE-1 BP complex-binding site in M8 compared to PAUSE-1, possibly reducing its putative negative binding influence. M10 potentially possesses two PAUSE-1-like binding sequences, which may increase its binding efficiency with the PAUSE-1 BP.
Functional studies showed that in its native environment (pCAT 5′-1.9) PAUSE-1 silenced PAI-2-directed transcription ∼3.4-fold. However, when cloned into pCAT 5′-1.7 or pCAT SV40, PAUSE-1 silenced transcription ∼2.6- and 2.1-fold, respectively. While PAUSE-1 clearly functions as an effective silencer, it may require other elements that are present in its native environment for maximal activity. Similar reductions in element potency compared to wild-type sequences have been reported elsewhere (31,32).
M3 behaved similarly to PAUSE-1 as a PAI-2-specific silencer. In fact, in the context of the PAI-2 promoter M3 was a stronger silencer than PAUSE-1 in HeLa cells. In conjunction with the EMSA data, this observation suggests that the four thymidine residues in PAUSE-1 may inhibit PAUSE-1 BP binding and therefore silencer activity. Thus, the thymidine repeat may actually play an important role in determining overall silencer function within the PAI-2 promoter.
M8 bound to the PAUSE-1 BP complex with similar affinity to PAUSE-1 but failed to silence PAI-2-directed transcription to the same level (1.6-fold). M8 contains a potential PAUSE-1 BP consensus binding site; a non-homologous 5′ flanking sequence and a 3′ thymidine repeat that is 4 nt downstream of the PAUSE-1-like consensus binding sequence. These results could suggest that the PAUSE-1 5′ flanking sequence plays a synergistic role in PAUSE-1 functionality by increasing silencer activity, without playing a role in PAUSE-1 BP binding. However, the overall architecture of M8 compared to PAUSE-1 is significantly altered and this could provide an alternative explanation for the decreased silencer activity observed.
In agreement with the EMSA data demonstrating absence of binding to PAUSE-1 BP, M6 and M7 fail to demonstrate any silencer activity in the context of the PAI-2 promoter. This is consistent with the proposed binding and functional sequence for PAUSE-1, as oligonucleotide M6 does not contain a CTC or AGA motif. Due to its diminished length, M7 may not be expected to bind to the PAUSE-1 BP complex. However, it was uncertain whether M7 would retain silencer activity. As M7 does not function as a PAUSE-1-like silencer, it is likely that either the 5′ or 3′ flanking sequence (or both) are important for PAUSE-1 function.
Taken together, the data demonstrate that the nucleotides present in the PAUSE-1 element which are critical for binding and functional activity in the context of the PAI-2 promoter were TCTNxAGAN3T4, where x = 0, 2 or 4.
mRNA expression may be controlled very simply by the differential expression of a strong silencer, as has been observed for NRSF in neuronal and non-neuronal cells (reviewed in 33). Thus, it could be hypothesised that PAUSE-1 BP activity would be observed in all PAI-2 non-expressing cell lines (e.g. HeLa) but not in PAI-2 expressing cell lines (e.g. HaCaT, MonoMac6 and U937). However, the PAUSE-1 BP complex was present in nuclear extracts from 15 different cell lines, indicating that the PAUSE-1 BP is widely expressed. Since PAI-2 mRNA expression is highly restricted among these cell lines, the mechanism of PAUSE-1 BP-mediated PAI-2 repression must be more complex, involving other cofactors, and not simply determined by the presence or absence of the PAUSE-1 BP complex.
Several PAUSE-1-associated complexes were observed in the various cell lines that were different to the PAUSE-1 BP complex observed in HeLa nuclear extracts with respect to mobility and/or the number of observed complexes. Several lines of evidence demonstrated, nonetheless, that these complexes associated with the 67 kDa PAUSE-1 BP. The relative mobilities of protein complexes on non-denaturing polyacrylamide gels are influenced by molecular mass, conformation and charge (34). Thus, while each cell line contained a PAUSE-1-specific DNA-binding complex, the PAUSE-1 BP complexes may be subtly different with respect to protein composition, conformation or redox state.
As noted by ourselves and others, several other genes in addition to PAI-2 contain PAUSE-1-like elements within their promoter regions (23,35). A search of the Eukaryotic Promoter Database (http://ww w.epd.isb-sib.ch /) revealed additional examples of PAUSE-1-like elements. A list of these genes and the respective elements are given in Table 2. Of these potential elements, only the ovalbumin PAUSE-1-like element has been characterised in vitro (36). This element was shown to silence transcription in the context of the homologous ovalbumin promoter and the heterologous thymidine kinase promoter (36). A second silencer with a similar binding sequence has subsequently been identified in the chicken ovalbumin gene (37). Although both of these elements bind to protein complexes as shown by EMSA, the identity of the specific DNA-binding proteins is not known. The sequence 5′-ANCCTCTCT-3′ was previously proposed as a consensus silencer element (36) and the PAUSE-1 element has been suggested to be a member of this ovalbumin-like silencer family (23). However, the data presented here and the presence of TCT and AGA motifs that are separated by 1–6 nt in all of these elements suggests a universal consensus sequence of TCTNxAGA.
Table 2. PAUSE-1-like promoter sequences.
 |
1Bold, capital letters represent specific sequence homology to PAUSE-1.
The identification of several PAUSE-1-like sequences in various species and kingdoms suggests that the PAUSE-1 consensus sequence may be a universal transcriptional regulator. Although a potential DNA-binding partner is yet to be identified for PAUSE-1, several DNA-binding proteins have been reported to bind to PAUSE-1-like sequences. For example, mouse Pax-1 binds to TAT AGA TCT CGC CTT GCC (38), Ski binds to the sequence GTC TAG AC (30) and the Smad 3 and 4 binding site is GTC TAG AC (39). Subsequent experimentation will revolve around identification of the PAUSE-1 BP and its various binding partners. Identification of the PAUSE-1 BP will provide invaluable information regarding the regulation of PAI-2 gene expression as well as potentially the regulation of other genes containing PAUSE-1-like elements.
Acknowledgments
ACKNOWLEDGEMENTS
We wish to thank Dr Brett Stringer for helpful discussions. We also thank Dr Peter Parsons of the Queensland Institute of Medical Research, Brisbane, Dr R. Sutherland of the Garvan Institute, Sydney, Prof. H. Zieger-Heitbrock of the University of Munich, Germany, and Prof. N. E. Fusenig of the Deutsches Krebsforschungzentrum, Heidelberg, Germany, for use of their cell lines.
References
- 1.Remold O.D.E. (1993) The ovalbumin family of serpin proteins. FEBS Lett., 315, 105–108. [DOI] [PubMed] [Google Scholar]
- 2.Dickinson J.L., Bates,E.J., Ferrante,A. and Antalis,T.M. (1995) Plasminogen activator inhibitor type 2 inhibits tumor necrosis factor alpha induced apoptosis. Evidence for an alternate biological function. J. Biol. Chem., 270, 27894–27904. [DOI] [PubMed] [Google Scholar]
- 3.Grigoryev S.A., Bednar,J. and Woodcock,C.L. (1999) MENT, a heterochromatin protein that mediates higher order chromatin folding, is a new serpin family member. J. Biol. Chem., 274, 5626–5636. [DOI] [PubMed] [Google Scholar]
- 4.Sager R., Sheng,S., Pemberton,P. and Hendrix,M.J. (1997) Maspin. A tumor suppressing serpin. Adv. Exp. Med. Biol., 425, 77–88. [PubMed] [Google Scholar]
- 5.Schleef R.R. and Chuang,T.L. (2000) Protease inhibitor 10 inhibits tumor necrosis factor alpha-induced cell death. Evidence for the formation of intracellular high M(r) protease inhibitor 10-containing complexes. J. Biol. Chem., 275, 26385–26389. [DOI] [PubMed] [Google Scholar]
- 6.Young J.L., Sukhova,G.K., Foster,D., Kisiel,W., Libby,P. and Schonbeck,U. (2000) The serpin proteinase inhibitor 9 is an endogenous inhibitor of interleukin 1beta-converting enzyme (caspase-1) activity in human vascular smooth muscle cells. J. Exp. Med., 191, 1535–1544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Antalis T.M., Clark,M.A., Barnes,T., Lehrbach,P.R., Devine,P.L., Schevzov,G., Goss,N.H., Stephens,R.W. and Tolstoshev,P. (1988) Cloning and expression of a cDNA coding for a human monocyte-derived plasminogen activator inhibitor. Proc. Natl Acad. Sci. USA, 85, 985–989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kruithof E.K., Baker,M.S. and Bunn,C.L. (1995) Biological and clinical aspects of plasminogen activator inhibitor type 2. Blood, 86, 4007–4024. [PubMed] [Google Scholar]
- 9.Mueller B.M., Yu,Y.B. and Laug,W.E. (1995) Overexpression of plasminogen activator inhibitor 2 in human melanoma cells inhibits spontaneous metastasis in scid/scid mice. Proc. Natl Acad. Sci. USA, 92, 205–209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Antalis T.M., La Linn,M., Donnan,K., Mateo,L., Gardner,J., Dickinson,J.L., Buttigieg,K. and Suhrbier,A. (1998) The serine proteinase inhibitor (serpin) plasminogen activation inhibitor type 2 protects against viral cytopathic effects by constitutive interferon alpha/beta priming. J. Exp. Med., 187, 1799–1811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Gan H.X., Newman,G.W. and Remold,H.G. (1995) Plasminogen activator inhibitor type 2 prevents programmed cell death of human macrophages infected with Mycobacterium avium, serovar 4. J. Immunol ., 155, 1304–1315. [PubMed] [Google Scholar]
- 12.Kumar S. and Baglioni,C. (1991) Protection from tumor necrosis factor-mediated cytolysis by overexpression of plasminogen activator inhibitor type-2. J. Biol. Chem., 266, 20960–20964. [PubMed] [Google Scholar]
- 13.Lavker R.M., Risse,B., Brown,H., Ginsburg,D., Pearson,J., Baker,M.S. and Jensen,P.J. (1998) Localization of plasminogen activator inhibitor type 2 (PAI-2) in hair and nail: implications for terminal differentiation. J. Invest. Dermatol., 110, 917–922. [DOI] [PubMed] [Google Scholar]
- 14.Buechler C., Ullrich,H., Ritter,M., Porsch-Oezcueruemez,M., Lackner,K.J., Barlage,S., Friedrich,S.O., Kostner,G.M. and Schmitz,G. (2001) Lipoprotein (a) up-regulates the expression of the plasminogen activator inhibitor 2 in human blood monocytes. Blood, 97, 981–986. [DOI] [PubMed] [Google Scholar]
- 15.Ritchie H., Jamieson,A. and Booth,N.A. (1995) Peripheral blood monocyte synthesis of plasminogen activator inhibitor 2 in response to native and modified LDL. Thromb. Haemost., 74, 1521–1527. [PubMed] [Google Scholar]
- 16.Antalis T.M., Godbolt,D., Donnan,K.D. and Stringer,B.W. (1993) Southwestern blot mapping of potential regulatory proteins binding to the DNA encoding plasminogen activator inhibitor type 2. Gene, 134, 201–208. [DOI] [PubMed] [Google Scholar]
- 17.Cousin E., Medcalf,R.L., Bergonzelli,G.E. and Kruithof,E.K. (1991) Regulatory elements involved in constitutive and phorbol ester-inducible expression of the plasminogen activator inhibitor type 2 gene promoter. Nucleic Acids Res., 19, 3881–3886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Dear A.E., Shen,Y., Ruegg,M. and Medcalf,R.L. (1996) Molecular mechanisms governing tumor-necrosis-factor-mediated regulation of plasminogen-activator inhibitor type-2 gene expression. Eur. J. Biochem., 241, 93–100. [DOI] [PubMed] [Google Scholar]
- 19.Mahony D., Kalionis,B. and Antalis,T.M. (1999) Plasminogen activator inhibitor type-2 (PAI-2) gene transcription requires a novel NF-kappaB-like transcriptional regulatory motif. Eur. J. Biochem., 263, 765–772. [DOI] [PubMed] [Google Scholar]
- 20.Samia J.A., Alexander,S.J., Horton,K.W., Auron,P.E., Byers,M.G., Shows,T.B. and Webb,A.C. (1990) Chromosomal organization and localization of the human urokinase inhibitor gene: perfect structural conservation with ovalbumin. Genomics, 6, 159–167. [DOI] [PubMed] [Google Scholar]
- 21.Ye R.D., Ahern,S.M., Le Beau,M.M., Lebo,R.V. and Sadler,J.E. (1989) Structure of the gene for human plasminogen activator inhibitor-2. The nearest mammalian homologue of chicken ovalbumin. J. Biol. Chem., 264, 5495–5502. [PubMed] [Google Scholar]
- 22.Ogbourne S. and Antalis,T.M. (1998) Transcriptional control and the role of silencers in transcriptional regulation in eukaryotes. Biochem. J., 331, 1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Antalis T.M., Costelloe,E., Muddiman,J., Ogbourne,S. and Donnan,K. (1996) Regulation of the plasminogen activator inhibitor type-2 gene in monocytes: localization of an upstream transcriptional silencer. Blood, 88, 3686–3697. [PubMed] [Google Scholar]
- 24.Boukamp P., Petrussevska,R.T., Breitkreutz,D., Hornung,J., Markham,A. and Fusenig,N.E. (1988) Normal keratinization in a spontaneously immortalized aneuploid human keratinocyte cell line. J. Cell Biol., 106, 761–771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ziegler-Heitbrock H.W., Thiel,E., Futterer,A., Herzog,V., Wirtz,A. and Riethmuller,G. (1988) Establishment of a human cell line (Mono Mac 6) with characteristics of mature monocytes. Int. J. Cancer, 41, 456–461. [DOI] [PubMed] [Google Scholar]
- 26.Maynard K. and Parsons,P.G. (1986) Cross-sensitivity of methylating agents, hydroxyurea and methotrexate in human tumor cells of the Mer– phenotype. Cancer Res., 46, 5009–5013. [PubMed] [Google Scholar]
- 27.Antalis T.M. and Godbolt,D. (1991) Isolation of intact nuclei from hematopoietic cell types. Nucleic Acids Res., 19, 4301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Chomczynski P. and Sacchi,N. (1987) Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem., 162, 156–159. [DOI] [PubMed] [Google Scholar]
- 29.Antalis T.M. and Dickinson,J.L. (1992) Control of plasminogen-activator inhibitor type 2 gene expression in the differentiation of monocytic cells. Eur. J. Biochem., 205, 203–209. [DOI] [PubMed] [Google Scholar]
- 30.Nicol R. and Stavnezer,E. (1998) Transcriptional repression by v-Ski and c-Ski mediated by a specific DNA binding site. J. Biol. Chem., 273, 3588–3597. [DOI] [PubMed] [Google Scholar]
- 31.Haniel A., Welge-Lussen,U., Kuhn,K. and Poschl,E. (1995) Identification and characterization of a novel transcriptional silencer in the human collagen type IV gene COL4A2. J. Biol. Chem., 270, 11209–11215. [DOI] [PubMed] [Google Scholar]
- 32.Shimokawa T. and Fujimoto,H. (1996) Identification of a transcriptional silencer in the protein-coding region of the mouse major inducible Hsp70 gene. Biochem. Biophys. Res. Commun., 221, 843–848. [DOI] [PubMed] [Google Scholar]
- 33.Jones F.S. and Meech,R. (1999) Knockout of REST/NRSF shows that the protein is a potent repressor of neuronally expressed genes in non-neural tissues. Bioessays, 21, 372–376. [DOI] [PubMed] [Google Scholar]
- 34.Orchard K. and May,G.E. (1993) An EMSA-based method for determining the molecular weight of a protein–DNA complex. Nucleic Acids Res., 21, 3335–3336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Baniahmad A., Muller,M., Steiner,C. and Renkawitz,R. (1987) Activity of two different silencer elements of the chicken lysozyme gene can be compensated by enhancer elements. EMBO J., 6, 2297–2303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Haecker S.A., Muramatsu,T., Sensenbaugh,K.R. and Sanders,M.M. (1995) Repression of the ovalbumin gene involves multiple negative elements including a ubiquitous transcriptional silencer. Mol. Endocrinol., 9, 1113–1126. [DOI] [PubMed] [Google Scholar]
- 37.Muramatsu T., Imai,T., Park,H.M., Watanabe,H., Nakamura,A. and Okumura,J. (1998) Gene gun-mediated in vivo analysis of tissue-specific repression of gene transcription driven by the chicken ovalbumin promoter in the liver and oviduct of laying hens. Mol. Cell. Biochem., 185, 27–32. [DOI] [PubMed] [Google Scholar]
- 38.Chalepakis G., Fritsch,R., Fickenscher,H., Deutsch,U., Goulding,M. and Gruss,P. (1991) The molecular basis of the undulated/Pax-1 mutation. Cell, 66, 873–884. [DOI] [PubMed] [Google Scholar]
- 39.Zawel L., Dai,J.L., Buckhaults,P., Zhou,S., Kinzler,K.W., Vogelstein,B. and Kern,S.E. (1998) Human Smad3 and Smad4 are sequence-specific transcription activators. Mol. Cell, 1, 611–617. [DOI] [PubMed] [Google Scholar]