


Abstract
The c-myc protooncogene plays an important role in the abnormal growth pattern of melanoma cells. In an attempt to inhibit c-Myc expression and the growth of an established murine melanoma cell line, we targeted homopurine sequences within the mouse myc mRNA with modified antisense oligonucleotides (AS ODNs). Psoralen was conjugated to the 5′-end of these clamp-forming oligonucleotides (clamp ODNs). Gel mobility shift analysis demonstrated a sequence-specific interaction between the active clamp ODNs (Myc-E2C and Myc-E3C) and the 1.4 kb c-myc mRNA, but no interaction with the control clamp ODN (SCR**). This association was further confirmed by thermal denaturation studies. In vitro translation assays demonstrated that both Myc-E2C and Myc-E3C at 5 µM inhibited c-Myc expression >99% after UV activation at 366 nm. Immunostaining of B16-F0 cells with a c-Myc monoclonal antibody revealed a significant reduction in c-Myc after clamp ODN treatment compared with the untreated or SCR** control-treated cells. This result was corroborated by western blot analysis. Utilizing the MTT assay to determine the effects of ODN-mediated c-Myc reduction on B16-F0 growth, we observed 60 and 64% reductions in growth after treatment with 5 µM Myc-E3C and Myc-E2C, respectively. We attribute the enhanced effectiveness of the clamp ODNs to psoralen activation. Our preliminary data suggest that inhibiting c-Myc overexpression results in a significant reduction in abnormal proliferation of B16-F0 melanoma cells and that the increased efficiency of clamp ODNs may provide an important advantage for their use in antisense therapies.
INTRODUCTION
Under normal conditions c-myc contributes to a number of pathways that govern cellular proliferation and differentiation (1). It is classified as an ‘immediate early response’ gene because of its role in cell cycle control (2). Rapid cellular proliferation appears to be directly related to induced overexpression of myc, while decreased expression decreases proliferation (3). Activated c-myc may be expressed at a higher level as a result of amplification, proviral insertion or chromosomal translocation and is consequently implicated in a wide variety of neoplasms (4,5). For example, c-myc overexpression correlates with relapse risk in uterine cervical carcinoma (6), whereas amplification in breast cancer correlates with poor prognosis (7). Increased Myc oncoprotein levels have been documented in head and neck cancers as well (8). Nonetheless, the cause(s) of c-myc overexpression in melanoma is currently unknown (9). To determine the effect of antisense inhibition of c-myc expression in melanoma cells, we have targeted exons 2 and 3 of the c-myc message with antisense (AS) ODNs in the B16-F0 mouse melanoma cell line (10).
Several alterations of AS ODNs have been utilized in attempts to increase their potential as inhibitors of cellular gene expression. We have initially employed three such modifications in the design of our ODNs, the first being the use of phosphorothioate linkages, which provide enhanced nuclease resistance, and the second being the combination of two covalently linked ODNs, allowing target mRNA hybridization to occur by both Watson–Crick and Hoogsteen hydrogen bonds. This ODN–target complex forms a ‘hairpin’ interaction or bimolecular triplex (11) in which the ODN acts as a ‘molecular clamp’ (12) around the target mRNA. The final addition of a photoreactive psoralen moiety conjugated to the 5′-end of the homopyrimidine ODN enables it to form a covalent adduct (crosslink) with the homopurine target strand after UV exposure at 366 nm, facilitating the formation of an irreversible, covalent adduct between the mRNA and clamp ODN. This irreversible process renders the message inactive for translation of full-length, functional protein products (13).
In addition to enhancing modifications, we considered the fundamentals of the ODN structure itself in our design. Based on the stability findings of Shimizu et al. (14), we chose to use an interlinking loop of four thymine bases, versus six, eight or more. Taking into account the body of data surrounding the increased stabilization of a pyrimidine·purine·pyrimidine DNA parallel triplex motif, one may wonder why we designed these clamp ODNs without the substitution of 5-methylcytosine in the Hoogsteen base pairing fragment to alleviate the problem of C+ protonation (15–17). Our initial plan was to design the most efficient AS ODN with the highest biological activity and lowest toxicity. We began by examining what we perceived as the major obstacles when composing these molecules: cellular entry, protection, specificity and persistence. We felt that this initial modification blueprint of our first generation clamp ODNs would overcome all these obstacles to achieve the desired effect without the compromise of increasing generalized toxicity and thus chose a simple, but calculated, approach to ODN design.
The goals of this study were three-fold: (i) to determine whether phosphorothioate clamp-forming AS ODNs (clamp ODNs) had enhanced efficiency compared to standard AS ODNs targeting the same sequences; (ii) to evaluate the ability of clamp ODNs to effectively lower in vitro and cellular c-Myc levels; and (iii) to ascertain the effects that reducing c-Myc expression would have on the proliferation rate of melanoma cells. We hypothesized that the clamp ODN may inhibit complete translation machinery assembly or structurally block polypeptide elongation (18,19). Gel mobility shift assays and thermal denaturation verified the ability of the active clamp ODNs to specifically hybridize with the c-myc message targets. These fundamental studies corroborated an initial in vitro experiment, which demonstrated increased efficiency of Myc-E2C and Myc-E3C to completely inhibit detectable c-Myc protein at 5 µM in a rabbit reticulocyte lysate system. By comparison, there was <5% inhibition of c-Myc translation with the standard AS ODNs (Myc-E2 and Myc-E3) or the control clamp ODN (SCR**). Immunohistochemistry revealed that only the active clamp ODNs are more efficient at reducing detectable cellular c-Myc levels, while western blot analysis confirmed a specific diminished cellular expression of the protein. In addition, Myc-E2C and Myc-E3C demonstrated an elevated potential to reduce B16-F0 cellular proliferation in a MTT cytotoxicity assay. Our data confirm that these clamp-forming ODNs show greater promise for use as therapeutic antisense agents and that c-Myc overexpression plays an important role in the abnormal growth of melanoma cells.
MATERIALS AND METHODS
Cell culture
B16-F0 is an established murine melanoma cell line obtained from the American Type Culture Collection (CRL-6322_FL). Cells were routinely passaged in Dulbecco’s modified minimal essential medium (DMEM) supplemented with 10% heat-inactivated fetal bovine serum and 1% Antibiotic-Antimycotic (10 000 IU/ml penicillin G, 10 000 µg/ml streptomycin sulfate, 25 µg/ml amphotericin B in 0.85% saline) (Gibco BRL). Cells were grown at 37°C in a water-jacketed CO2 incubator (Nuaire).
Oligonucleotide synthesis
Phosphorothioate oligonucleotides were prepared in an Applied Biosystems 394 automated DNA/RNA synthesizer using standard phosphoramidite chemistry and purchased from the University of Alabama at Birmingham Oligonucleotide Core Facility or Oligos Etc. The sequences of all ODNs are shown in Table 1.
Table 1. 5′→3′ sequences of non-clamp and clamp antisense oligonucleotides.
 |
Oligonucleotide sequences are complementary to sequences in exon 2 (Myc-E2 and Myc-E2C) or exon 3 (Myc-E3 and Myc-E3C) of the c-myc transcript.
apsor represents the 5′-end-conjugated psoralen moiety in clamp ODNs.
b(T4) represents the four thymine base pairs forming the loop region in the clamp ODNs.
cODNs are non-clamp-forming (NC), clamp-forming (CF) or scrambled clamp-forming (SC).
dSchematic of clamp ODN–mRNA target complexes at sequences within the second and third exons.
RNA isolation and northern blot analysis
Total RNA was isolated from a 70% confluent culture of cells harvested with 1% trypsin–EDTA in Hank’s balanced salt solution (HBSS) according to the outlined protocol for the RNeasy Total RNA kit (Qiagen). Total RNA concentration and purity were determined by spectrophotometric measurement at A260 and A280. Aliquots of 20, 10 and 5 µg of RNA were loaded onto a 1% formaldehyde gel, transferred to a neutral nitrocellulose membrane overnight and probed with a 32P-labeled 1.8 kb mouse c-myc cDNA probe (20).
Western blot analysis
A 65% confluent T-150 culture flask of B16-F0 cells was harvested in a 1:10 solution of trypsin–EDTA and HBSS, centrifuged and resuspended in 350 µl of 1× lysis buffer (5X; Promega), incubated for 20 min, the supernatant recentrifuged and transferred to a fresh tube. An aliquot of 35 µl of the cell lysate was mixed with 30 µl of SDS gel loading buffer and resolved by 10% SDS–PAGE. The gel was transferred to a PDVF membrane and western blot analysis was performed according to the procedure provided with the primary antibody. Briefly, the membrane was incubated at room temperature for 1 h in a 5% non-fat milk powder solution in 0.05% PBS-T (Sigma) to block non-specific reactivity. The blot was then washed three times for 5 min each time with PBS-T alone. It was then incubated for 1 h at room temperature in PBS-T containing a 1:100 dilution of mouse-reactive c-Myc monoclonal antibody c8 (Santa Cruz Biotechnology Inc.). After another series of 3–5 min washes, the blot was incubated for 45 min with a 1:500 dilution of horseradish peroxidase-conjugated goat anti-mouse secondary antibody (Dako). The blot was again washed three times in PBS-T then once in PBS alone for 5 min. Protein detection was performed using ECL chemiluminescent reagents (Amersham), followed by rapid autoradiography at room temperature.
Plasmid construction and clone isolation
The pUC18 plasmid was digested with XhoI to generate a 1.4 kb mouse c-myc cDNA fragment (21). This insert was subcloned into the 2.9 kb pBluescript SK(+/–) phagemid vector (Stratagene) at the compatible restriction site relative to the T7 promoter by ligation in the presence of T4 DNA ligase and ligase buffer (Gibco BRL) for 18 h at 15°C. The DNA mixture was used to transform 50 µl of Escherichia coli strain DH5α competent cells (Gibco BRL), plated onto ampicillin (100 mg/ml) + LB agar plates and grown overnight for 16 h at 37°C. Colonies were grown up in 3 ml of LB with ampicillin and plasmid DNAs were purified using a standard alkaline lysis miniprep kit (Qiagen), followed by purification with Quantum Prep Freeze ’n Squeeze DNA Gel Extraction Spin Columns (Bio-Rad) from a 1% TAE–agarose gel. After verifying the presence of the 4.3 kb resultant plasmid on a 1% TBE–agarose gel, a large-scale plasmid preparation (Qiagen) was performed to retrieve ample DNA for in vitro transcription experiments. The pUC18 parental plasmid was a generous gift from Dr David Jones (Moffat Cancer Research Center, University of South Florida).
In vitro transcription and rabbit reticulocyte lysate translation
The pBS:M-c-myc plasmid was linearized with HindIII and prepared for template use in an in vitro run-off transcription reaction using T7 polymerase in Novagen’s Large Scale Transcription kit. Briefly, the following kit components were assembled at room temperature with the template: 5× Transcription Buffer, 20 mM ATP, CTP, GTP and UTP, 1 M DTT, RNase-free water and 100 U T7 RNA polymerase. The reactions were incubated at 37°C for 2 h. Next, 5 U DNase I was added to stop the transcription reaction and remove the template. mRNAs were repeatedly purified by phenol:chloroform extraction and ethanol precipitation, quantitated at A260, then analyzed on a 1% TAE (DEPC)–agarose gel. Each initial translation reaction contained the pH 7.6 Novagen Translation Mix (25 mM DTT, 250 mM HEPES, 100 mM creatine phosphate and 312.5 µM all 19 amino acids except methionine), 0.5 mM MgOAc, RNase-free water and 0.6 µg 1.4 kb c-myc mRNA template incubated with ODN at 1, 2.5, 5 and 7.5 µM for 30 min at 37°C, then UV-crosslinked (366 nm) for 5 min on ice. After prehybridization, 100 mM KOAc and nuclease-treated rabbit reticulocyte lysate were added with 1.0 mCi/ml [35S]methionine and samples were incubated for 1 h at 30°C. Although this lysate system allows for cap-dependent in vitro translation, with the suggested use of the pCITE vector and KCl to achieve higher fidelity with capped mRNAs, we had no need for these proteins in the translation of our myc insert and thus used KOAc in our reactions. Unincorporated label was removed from 15 µl of each reaction mixture by acetone precipitation (Novagen protocol), then dissolved in 55 µl of SDS gel loading buffer. Samples were heated at 90°C for 3 min, cooled, then resolved by 10% SDS–PAGE. Gels were dried onto Whatmann paper and exposed to BioMax MR autoradiography film (Kodak) at –70°C (22).
Gel mobility shift analysis
Clamp ODNs Myc-E2C, Myc-E3C, SCR** (control) and HaeII-cleaved pBR322 (marker) were 3′-end-labeled with deoxyadenosine 5′-triphosphate (ddATP), 3′-[α-32P]cordycepin 5′-triphosphate [5000 Ci (185 TBq)/mmol; NEN] according to the protocol outlined in the DNA 3′-End Labeling kit (Boehringer Mannheim). Non-incorporated [α-32P]ddATP was removed using STE Select-D G-25 spin columns (5 Prime→3 Prime Inc.). Each ODN (50 000 c.p.m.) was added to a reaction mixture containing 25 mM DTT, 250 mM HEPES, 25 mM magnesium acetate, 100 mM creatine phosphate, pH 7.6, and 2.5 µg in vitro transcribed c-myc mRNA, in a total volume of 30 µl. The reactions were incubated at 37°C for 30 min then UV-crosslinked (366 nm) for 5 min on ice. Samples were electrophoresed on a 12% native polyacrylamide gel at 350 V in 1× TBE buffer. The gel was exposed to Biomax MR autoradiography film (Kodak) for 18 h at –80°C, then processed.
Thermal denaturation
Short 35 bp RNA oligomers containing either the exon 2 or exon 3 target were commercially synthesized by Oligos Etc. Inc. Each target ODN was combined with its respective clamp ODN (E2C or E3C) at an equimolar ratio (3 µM each) in a 50 mM Tris–acetate buffer containing 10 mM MgCl2 and 50 mM NaCl, pH 5.5. The reaction mixtures, as well as each ODN component alone, were incubated at 37°C for 30 min, then slowly cooled overnight at 4°C. The non-clamp E2 and E3 and clamp SCR** ODNs were also combined with the respective target RNA ODNs (both with clamp SCR**) to serve as controls for specificity. Melting studies were conducted using the Swift thermo-program on a Pharmacia Biotech Ultraspec 2000 spectrophotometer, in Teflon stoppered 1 cm path length quartz cells. Absorbance (260 nm) was measured while the temperature incrementally increased from 20 to 85°C at a rate of 0.5°C/min. The melting temperature (Tm) was calculated by computer fit of the first derivative of the absorbance with respect to 1/T, for two or three consecutively repeated runs.
Immunohistochemical quantitation of c-Myc
B16-F0 cells were seeded at a density of 104 cells/well in 4-chamber slide plates and incubated for 18 h at 37°C. Cells were transiently transfected with 5 µM ODN in Opti-MEM I medium (Gibco BRL) for 6 h. Plates were UV-irradiated (366 nm) on ice for 5 min, then supplemented DMEM (Gibco BRL) was added for 18 h of incubated growth. The treatment regimen was repeated three more times. Twenty hours after the final treatment, cells were fixed with cold acetone/methanol/37% formalin (19:19:2 v/v/v) for 30 min at –70°C and immunostained according to a modification of the procedures described by Ebbinghaus et al. (23). Slides were incubated with 200 ng/ml mouse c-Myc monoclonal antibody (Santa Cruz Biotechnology Inc.) at room temperature for 1 h. Immunodetection was performed with the ABC system (Santa Cruz Biotechnology Inc.) and HRP–DAB reaction. Slides were alcohol dehydrated, fixed with Permount solution (Sigma), coverslipped and photographed on an Olympus microscope.
Western blot analysis of c-Myc expression
B16-F0 cells were seeded at 105 in 60 mm culture dishes. Cells were treated according to the previously outlined transfection procedure, with one modification. Cell lysates were harvested 30 min after the final treatment and DMEM replenishment rather than at 20 h (24). Cells were trypsin–EDTA harvested (1:10 in HBSS), centrifuged and resuspended in 25 µl of 1× lysis buffer, incubated for 20 min, recentrifuged and the supernatant transferred to a fresh tube. Total protein extracts were loaded with an equal volume of SDS gel loading buffer and fractionated by 10% SDS–PAGE. The gel was transferred to a nitrocellulose membrane and western blot analysis was carried out according to the procedure previously outlined for both c-Myc and β-actin expression.
Effect of ODNs on cellular proliferation
B16-F0 cells were seeded at 103 cells/well in 96-well plates in triplicate and incubated for 18 h at 37°C. Cells were treated as in the immunhistochemistry experiment. Twenty hours after the final treatment, 10 µl of MTT reagent (BMB; Sigma) was added to each sample for 4 h, then 100 µl of solubilization solution (0.01 HCl in 10% SDS) was added for 18 h. Plates were analyzed on a microplate reader at 595 nm to generate the graphical data.
RESULTS AND DISCUSSION
Northern and western blot analyses to measure c-myc mRNA and c-Myc protein expression in B16-F0 cells
We first evaluated the baseline level of c-myc expression in the B16-F0 mouse melanoma cell line. Northern analysis confirmed a significant level of expression of the 2.4 kb c-myc mRNA target, after hybridization to a 1.8 kb, 32P-labeled mouse myc probe. Figure 1A shows the transcript expression level in 20, 10 and 5 µg samples of total RNA taken from a 70% confluent flask of cells. The lower panel shows expression of β-actin message for relative comparison with expression of the target transcript. We next used western blot analysis to examine the level of c-Myc protein as an indicator of endogenous c-myc expression. As seen in Figure 1B, the B16-F0 cells produce a significant amount of c-Myc protein. This level of expression was adequate to allow detection of the reduction in c-Myc protein that results after ODN treatment. Based on these data, we selected the B16-F0 line as a suitable cell system in which to study both clamp ODN activity and the effects on cellular proliferation after a significant reduction in endogenous c-Myc levels (Fig. 1).
Figure 1.

Endogenous c-myc mRNA and c-Myc protein expression in the B16-F0 cell line. (A) Detection of the 2.4 kb c-myc mRNA by northern blot analysis with a 1.8 kb cDNA probe. The lower autoradiogram is the same blot reprobed with a 2.1 kb β-actin probe to compare relative levels of expression. (B) Western blot detection of the c-Myc 67 kDa primary protein product with the c8 c-Myc monoclonal antibody.
Inhibition of in vitro translated c-Myc in the presence of oligonucleotide
Cell-free assays provide an effective means with which to determine ODN binding affinity, target recognition and the optimal effective concentration. We subcloned exons 2 and 3 of the c-myc cDNA coding region (nucleotides 545–1937) into the pBluescript SK(+/–) RNA expression vector (Stratagene). The in vitro generated run-off transcripts were used as the mRNA target pool for the rabbit reticulocyte lysate translation study in this report. AS ODNs were designed which were complementary to nucleotides 685–695 of exon 2 and 1381–1390 of exon 3.
To determine the inhibitory effects of the Myc-E2C and Myc-E3C ODNs on c-myc translation, four concentrations of ODN (1, 2.5, 5 and 7.5 µM) were tested in this in vitro system. Prior to translation, mRNA (0.6 µg) and ODNs were preincubated with all translation reagents except KOAc (100 mM), [35S]methionine and lysate for 30 min at 37°C, as previously outlined in Materials and Methods. Psoralen-conjugated clamp ODNs and control reactions (no ODN) were then UV exposed at 366 nm for 5 min on ice. The remaining reagents were added and the samples were incubated for 60 min at 30°C. The 35S-labeled acetone precipitates were then analyzed by SDS–PAGE. As seen in Figure 2, Myc-E2C and Myc-E3C completely inhibit Myc translation at 5 and 7.5 µM, while there was no significant inhibition observed with SCR**, the control clamp ODN, nor the standard AS ODNs Myc-E2 and Myc-E3 (<5%) at the same concentrations.
Figure 2.

Inhibition of in vitro translation of c-myc mRNA following treatment with AS and clamp ODNs in a cell-free rabbit reticulocyte lysate system. Lane 1, lysate only control, containing neither mRNA nor ODN; lane 2, c-myc mRNA alone translation control; lanes 3–6, treated with ODN Myc-E2 at 1, 2.5, 5 and 7.5 µM concentrations, respectively; lanes 7–10, treated with ODN Myc-E3 at 1, 2.5, 5 and 7.5 µM concentrations, respectively; lanes 11–14, treated with ODN Myc-E2C at 1, 2.5, 5 and 7.5 µM concentrations, respectively; lanes 15–18, treated with ODN Myc-E3C at 1, 2.5, 5 and 7.5 µM concentrations, respectively; lanes 19–22, treated with SCR**, the control clamp ODN, at 1, 2.5, 5 and 7.5 µM concentrations, respectively. c-Myc translation after Myc-E2 and Myc-E3 treatment (non-clamp AS ODNs) was not affected at any concentration (lanes 3–10). Compared to control translation of the 1.4 kb transcript, both Myc-E2C and Myc-E3C exerted concentration-dependent translation inhibition on c-Myc expression (lanes 11–18). After treatment with 5 µM of either clamp ODN (lanes 13 and 17) Myc translation was no longer detected, suggesting 100% inhibition. SCR** treatment produced no effect on the translation of Myc protein in this system (lanes 19–22).
Proposing the main mechanism of action as translation inhibition by blocking elongation of the nascent transcript into functional protein, the presence of prematurely terminated polypeptides might be expected as confirmation. The fact that none were detected in this system does not nullify the proposed mechanism. If any were to be present, they would result from the E2C clamp ODN treatment, because the exon 2 target sequence starts 75 bp downstream of the endogenous c-myc AUG codon (nucleotide 607), to generate a polypeptide chain of ~25 amino acids. A chain of this size would have a mass of only ∼2.7 kDa. The Kaleidoscope prestained molecular weight standards (Bio-Rad) used in this experiment would not be able to detect a polypeptide chain this small, since the lowest limit of detection is 8 kDa. Concerning the E3C clamp ODN, no premature termination polypeptide would be expected because the target sequence includes the G of the start codon and extends to the 9 bases following the AUG.
In addition to providing an effective treatment dose, these results show that at a low concentration maximum inhibition of translation of Myc can be achieved and is dependent on sequence specificity and a mechanistic advantage of the active clamp ODNs. We directly attribute this enhanced effectiveness of Myc-E2C and Myc-E3C to the ability of psoralen to intercalate in a site-specific manner and form an irreversible covalent adduct with the target mRNA. This observation agrees with other studies that conjugate moieties provide an advantage for ODN efficacy and stability, especially when targeting regions outside or downstream of the translation initiation site (18,19,25,26). Additionally, the structure of clamp ODNs provide increased stability by the dual interaction of two hybridizing fragments, which engage almost all possible hydrogen bonds within the homopurine target sequence (13). In the absence of UV activation there was no significant inhibitory difference between the non-clamp and active clamp ODNs at 5 µM. At 25 µM, without exposure to UV E2C inhibited Myc synthesis by 40% and E3C by 50%, while there was no significant inhibition after SCR** treatment up to 50 µM (data not shown).
Although we feel that the clamp ODNs primarily act as structural impediments to elongation, it is also feasible that when only the Watson–Crick base pairing ODN has hybridized to the target mRNA RNase H is activated, degrading the message. Because this may be occurring in this system, it is necessary that we mention it, but it should be noted that this is most likely a negligible occurrence. If such an occurrence greatly interfered with the results of protein synthesis it would be most apparent in lysate reactions containing the non-clamp ODNs (Fig. 2, lanes 3–10). With no evidence of inhibition of Myc synthesis in any of these samples, it can only be concluded that RNase H activation is minimal at these ODN concentrations in this system (Novagen).
Gel mobility shift analysis of clamp ODNs with c-myc mRNA
In order to confirm that the inhibition of c-Myc protein expression was in fact due to a sequence-specific interaction between the c-myc message and the clamp ODNs, we performed a gel mobility shift assay. This experiment was designed to establish the ability of Myc-E2C and Myc-E3C to form a molecular clamp (12) with the single-stranded myc targets in exons 2 and 3, as well as to confirm the resultant inhibitory effects on c-Myc expression. Figure 3 demonstrates sequence-specific binding of the 1.4 kb c-myc mRNA by both the active Myc-E2C and Myc-E3C clamp ODNs, whereas no interaction was detected with the control clamp ODN SCR**.
Figure 3.
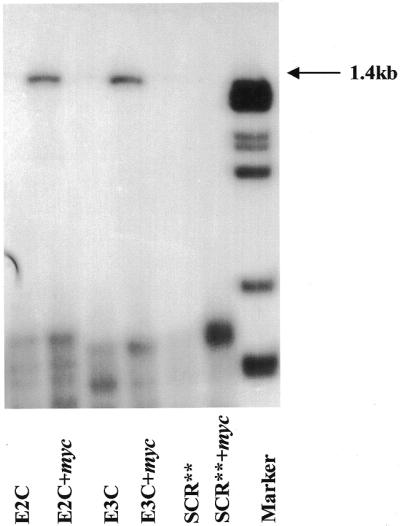
Gel mobility shift analysis of clamp ODNs with c-myc mRNA. Gel mobility shift analysis demonstrates sequence-specific binding of Myc-E2C and Myc-E3C to the 1.4 kb c-myc mRNA. The first and third lanes are the labeled active clamp ODNs alone. No shift occurred between the control clamp ODN, SCR**, which migrates at the same mobility in the presence of the mRNA as it does alone. The last lane is HaeII-cleaved pBR322 marker DNA.
Thermal dissociation of clamp ODNs with exon 2 and exon 3 c-myc RNA target oligomers
To further confirm the association of the active clamp ODNs with their targeted sequences in the myc mRNA we utilized short (35mer) RNA oligomers containing the exon 2 or exon 3 targets (Oligos Etc.). Figure 4 shows the melting curves and an average of two or three Tm values for Myc-E2C alone, its controls and the clamp ODN–RNA complex. None of the clamp ODNs were thermally stable alone (Fig. 4A, 0°C). In contrast, the target RNA oligomers did have some secondary structure, as predicted by Oligos Etc. (Fig. 4B, 47.2°C). Melting curves for the RNA oligomers with non-clamp ODNs were essentially superimposable with curves for the RNA oligomers alone (data not shown). It was again confirmed that SCR** could not associate with either myc sequence by the fact that both curves (Fig. 4D, 47.0°C) could be superimposed on those for the respective RNA oligomers alone (Fig. 4B). Only Myc-E2C (Fig. 4C, 49.7°C) and Myc-E3C had stability transitions and higher melting temperatures indicative of a more complex structure. Tm measurements for Myc-E3C were 0°C (Myc-E3C alone), 46.5°C (E3R alone), 44.7°C (SCR** + E3R) and 50.8°C (Myc-E3C + E3R) (data not shown). The monophasic transition seen only in the melting curve of the active clamp ODN (E2C) with its complementary RNA oligomer target is consistent with other data signifying a simultaneous dissociation of an intramolecular triplex-like complex (27). All thermal denaturation studies were conducted in a pH 5.5 buffer based on the knowledge that C-G-C triplets associate more strongly at acidic pH (28). This is most likely due to the increased stabilization after protonation of the cytosines in the Hoogsteen base pairing ODN domain (16).
Figure 4.

Thermal denaturation curves of clamp ODNs and c-myc RNA oligomers. Melting curve of Myc-E2C alone (A), 35 bp RNA oligomer E2R alone (B), Myc-E2C with E2R (C) and SCR** with E2R (D). Melting studies were done in 50 mM Tris–acetate buffer (10 mM MgCl2 and 50 mM NaCl), pH 5.5, with an equimolar amount of each ODN (3 µM). The absorbance (OD) was measured at 260 nm.
Immunodetection of c-Myc after ODN treatment in B16-F0 cells
To assess the ability of the active clamp ODNs to generate a reduction in cellular c-Myc protein expression, we employed a c-Myc-specific murine monoclonal antibody (Santa Cruz Biotechnology Inc.) in an immunohistochemical analysis. B16-F0 cells were fixed to the slide plate after ODN treatment and c-Myc detection was performed according to a modified procedure discussed in Materials and Methods. The expression of c-Myc protein was easily detected by strong nuclear staining in untreated cells (Fig. 5b). Treatment with 5 µM Myc-E2C or Myc-E3C clamp ODNs considerably reduced this expression (Fig. 5e and f), whereas the same concentration of non-clamp (Myc-E2 and Myc-E3) and control clamp ODN (SCR**) essentially had no effect (Fig. 5c, d and g). There was no staining for c-Myc detected in the control cells serving as a background for immunostaining (Fig. 5a). Additionally, the cells treated with the active clamp ODNs displayed a greater degree of cell death compared to the untreated and normal AS ODN-treated cells. Cells treated with SCR** also exhibited cellular death, but to a lesser degree. To determine whether this observed death was a clamp ODN effect versus a specific consequence of c-Myc reduction after Myc-E2C and Myc-E3C treatment we performed western blot analysis, to specifically monitor Myc protein levels after each ODN treatment, and cytotoxicity–proliferation experiments.
Figure 5.

Representative photographs of B16-F0 cells after ODN treatment and immunohistochemical analysis with the c8 c-Myc monoclonal antibody. Cells were treated in slide plates for 4 days, 6 h each day at 37°C, with Opti-MEM medium (a and b) or with 5 µM Myc-E2 (c), Myc-E3 (d), Myc-E2C (e), Myc-E3C (f) or SCR** (g). Endogenous c-Myc expression is detected as a brown stain against a blue hematoxylin counterstain. The cells in (a) were treated with medium alone and were not stained with primary Myc antibody, thus serving as a background control for antibody reactivity. The cells in (b) were also treated with medium only, but were stained with primary antibody to serve as the non-ODN-treated control. The cells in (c) and (d) were treated with Myc-E2 and Myc-E3, respectively, and both show immunoreactivity consistent with the untreated control cells (b). In contrast, the cells treated with Myc-E2C (e) and Myc-E3C (f) exhibit a drastic reduction in cellular Myc detection, as well as marked changes in cell morphology and cell death. The cells treated with the control clamp ODN, SCR** (g), also underwent some cell death, but maintained immunoreactivity for the Myc antibody comparable to the untreated control (b), which implies non-specific effects. The results of the immunohistochemical analysis further substantiate that Myc-E2C and Myc-E3C can specifically target the c-myc mRNA to reduce cellular c-Myc expression, an effect also confirmed in the in vitro translation assay.
Western analysis of cellular c-Myc expression after ODN treatment
Figure 6 demonstrates the ability of Myc-E2C and Myc-E3C to reduce the cellular levels of Myc in the B16-F0 cell line. Here, cells were treated for a total of 4 days with 5 µM concentrations of Myc-E2C, Myc-E3C, Myc-E2, Myc-E3, SCR** or medium alone, for 6 h every day. The cells were harvested 30 min following the last medium replenishment. Western blot analysis was performed, as described in Materials and Methods, using the c8 c-Myc antibody, as well as a monoclonal antibody against β-actin to serve as an internal control for global protein expression. After ECL detection and rapid autoradiography there was clear visualization of the antibody-bound c-Myc bands. By comparison of c-Myc levels by densitometry (Fig. 6) to untreated cells, it was found that Myc-E2C treatment reduced B16-F0 cellular c-Myc expression by 78%, Myc-E3C by 65%, and the non-clamp ODNs Myc-E2 and Myc-E3 by 35 and 7%, respectively. There was no significant change in Myc expression after treatment with the control clamp ODN SCR** nor any non-specific changes in β-actin expression. The only change in actin was observed after E2C treatment, a consequence of the increased cell death and resulting reduction in total cell volume relative to the controls.
Figure 6.

ECL detection of western blot analyzed B16-F0 cellular lysates treated with clamp ODNs and UV irradiation (366 nm). After four 5 µM ODN treatments total cell lysates were harvested, resolved by 10% SDS–PAGE and underwent western analysis using the c8 c-Myc and β-actin monoclonal antibodies (Santa Cruz Biotechnology Inc.). The blot was exposed to ECL reagents (Amersham) for c-Myc and actin protein detection. The lanes are labeled with the treatment (i.e. lane 1, CTL, is the UV alone control for B16-F0 cellular c-Myc expression). Myc-E2C treatment resulted in >78% reduction in c-Myc protein expression, while Myc-E3C resulted in >65% reduction. The lanes treated with Myc-E2 and Myc-E3 resulted in 35 and 7% reductions in c-Myc expression, respectively. There was no change in Myc expression after treatment with SCR**.
ODN effects on B16-F0 proliferation
The purpose of this experiment was to evaluate the ability of clamp ODNs to inhibit cellular proliferation and to determine the effective ODN concentration. We also wanted to determine whether the cell death observed in the immunohistochemistry experiment was the result of Myc-E2C and Myc-E3C specifically targeting c-myc message or a general, non-specific effect produced by clamp ODN treatment.
B16-F0 cells were plated in triplicate and treated by the same transfection procedure used in the immunohistochemical analysis. Alternatively, 20 h after final treatment we used the MTT assay instead of the trypan blue dye exclusion method because it allows a screen of the complete sample field in the experimental vessel. This essentially eliminates the error associated with experimental manipulation when removing a sample and provides a more exclusive measure of experimental deviation.
Figure 7 demonstrates that a 5 min UV exposure is not cytotoxic to these cells when compared to the non-ODN-treated control and that four treatments with ODN at 5 µM gave the maximal effect. We found that UV exposure for 10 or 20 min resulted in significant cellular cytotoxicity and 2 day or 1 µM ODN treatments were not as effective, while 10 µM treatments exhibited more non-specific effects with the control ODN (data not shown). In contrast, the shorter UV exposure time appeared to enhance the ability of Myc-E2C and Myc-E3C (5 µM) to reduce cellular proliferation by 64 and 60%, respectively. We attribute this effect to activation of the photoreactive reaction whereby psoralen forms an irreversible covalent crosslink with the target mRNA (18,19). Neither Myc-E2, Myc-E3, SCR** nor UV alone reduced the proliferation rate of this established melanoma cell line by more than 10%. Thus, these experiments demonstrate that psoralen-conjugated ODNs targeting c-myc mRNA are capable of inhibiting cellular proliferation by as much as 64%, while control ODNs inhibit by no more than 10%.
Figure 7.

B16-F0 murine melanoma growth inhibition by c-myc non-clamp and clamp AS ODNs, as measured by the MTT assay. Cells were treated with ODN (filled square) or a combination of ODN and UV irradiation (open square). Myc-E2C and Myc-E3C resulted in 64 and 60% reductions in cellular proliferation, respectively. All other ODNs resulted in no significant effects, as compared to control growth. Means ± SE, defined by bars, of triplicate wells for all data points, representative of four independent experiments, are given.
Although the exact mechanism of action has not yet been defined, we suggest that the ODNs either impede proper or complete assembly of the translation machinery or stop the translating ribosome at the point of the psoralen crosslink, thus inhibiting elongation. Both the gel shift and thermal denaturation studies confirmed that these DNA oligonucleotides bind their intended c-myc mRNA target sequences by a specific interaction, compared to controls. Immunohistochemistry and western blot analysis confirmed that the active clamp ODNs target the endogenous c-myc message in these melanoma cells and reduce c-Myc protein expression. Finally, we suggest that the cell death observed in the immunostaining experiment and mirrored in the MTT assay are both a consequence of suppressed Myc levels, and not a result of UV exposure or non-specific ODN effects. These data therefore validate the ability of the Myc-E2C and Myc-E3C clamp ODNs to target c-myc mRNA and inhibit cellular growth as a result of reduced Myc oncoprotein expression in B16-F0 melanoma cells.
CONCLUSIONS
The use of AS ODNs to inhibit abnormal expression of oncogenic proteins at the level of their mRNA precursors was initially utilized in 1978 (29). Since that time enormous strides have been made within this field. Many studies have demonstrated the ability of AS ODNs to specifically block gene expression with therapeutic significance, resulting in their progression to the clinical trial stage (30–35). We have targeted two coding regions of the c-myc oncogene primary transcript in the B16-F0 mouse melanoma cell line. Psoralen-conjugated, clamp-forming AS ODNs were used alone at 5 µM to significantly reduce expression of the c-Myc primary protein product. The use of cationic lipids as the vehicle for delivery was eliminated in this study based on previous studies in our laboratory. In particular, we have observed that using such tools with the intention of enhancing uptake and lowering the necessary concentration of ODN required to achieve the desired effect does not always occur when working with phosphorothioate ODNs. Contrary to the desired outcome, we have experienced the need to increase the ODN concentration when combined with certain lipids, to overcome the tendency of these reagents to bind the ODN too tightly, not releasing enough into the cell cytoplasm to result in significant inhibition of c-Myc expression or cell proliferation. Further confirmed by immunohistochemical observation, there was an increase in punctate staining, indicating lysosomal or other vesicle trapping of the ODN when lipids were used. Also, some transfection reagents may increase the possibility of non-specific cellular toxicity.
This study provides further evidence to support the hypothesis that c-Myc overexpression may contribute to the malignant proliferation of melanoma. More specifically, our data implicate probable deregulation of one or more of the mechanisms that govern in vivo c-Myc expression. In particular, the management of mRNA stability or translation or protein stability, three key levels at which Myc activity is tightly controlled (36). We propose that the mechanism through which these clamp ODNs act is translation inhibition, by functioning as structural blocks to the elongating ribosomal machinery. In addition, they may generate substrates for RNase H, prior to complete formation of the clamp ODN–mRNA target complex.
A number of studies have shown that inhibiting myc by the antisense therapeutic approach resulted in beneficial malignant growth inhibition (37–40). In addition to regarding Myc levels as a ‘prognostic marker’ for this cancer, we have sought to further establish it as a key contributor to this disease state (17). Whether it is causative or resultant in melanoma remains to be determined, especially as more information on the complete function and regulation and its upstream and downstream effectors becomes available. Particularly interesting is the recent work of Bazarov et al. (41) regarding a 10-fold reduction in Ras-dependent transformation in response to only a moderate reduction in c-Myc expression. Using c-myc+/+, c-myc+/– and c-myc–/– (42–44) cell lines, they found that the ability of Ha-Ras(G12V) (45) to negate the expression of anchorage deprivation-induced p27 (46,47) was itself nullified by the absence of c-Myc expression (41). Because a 50% reduction in Myc was found to only modestly affect proliferation and apoptosis in their system, this study has blazed a new trail for the therapeutic use of highly active AS ODNs (41). Where the main focus of most antisense research has been to significantly inhibit the overexpression of oncogenic proteins, with the goal of reversing the malignant phenotype, another avenue for achieving this objective has been paved. In our melanoma system, where Myc is clearly overexpressed, we have obtained a substantial reduction in cell growth after treatment with myc-targeted clamp ODNs; the proposed therapeutic outcome is obvious. Based on these observations that down-regulation of Myc diminishes malignant melanoma proliferation, future studies should focus on c-myc transcription-specific regulation and the factors both directly or cooperatively involved, to delineate whether myc is exerting causative effects, or on translation-specific regulation and its factors, if resultant effects are in operation. Even so, a further therapeutic potential is the application of these ODNs to systems where Myc expression is known to be a resultant effect, yet key for the propagation of disease by a primary or cooperating, more causal oncogene (41). It is important that systems previously unattractive for antisense-based therapy be re-examined from this alternative perspective of, rather than working to eliminate the ‘boss’, cutting off the ‘right-hand’ man.
We are currently testing these ODNs in mice and exploring measures for enhanced in vivo administration, with the anticipation that as better delivery systems are developed the most active ODNs will serve as viable options for therapy in the clinic. Simultaneously, we are continuing efforts to design the most active antisense molecules. With this goal in mind, future modifications under consideration include lengthening the loop by one base, because of the study conducted by Prakash and Kool (48) indicating that 5 nt loops offer even greater stability to bimolecular and circular ODNs than those composed of 4, 6 or 8 nt, especially when the first and fifth bases are purines (49). We also intend to utilize the finding that using loop-end bases complementary to each other and/or the nucleotide directly adjacent to the 5′-end of the target strand increases both binding affinity and sequence selectivity (50). Finally, our second generation clamp ODNs will most likely contain 5-methylcytosines in place of the Hoogsteen fragment cytosines to determine whether the documented stability would increase the overall anti-proliferative efficacy of these ODNs (51).
Our study confirms the necessity of ODN modifications, particularly when targets are within the coding region of the message, since it has been shown that standard AS ODNs are unable to stop actively translating ribosomal complexes (52). Thus, when considering mRNA target sites, especially those located downstream of the translation initiation region, these data support further development of AS ODNs with maximum efficiency, having the capability to serve as potent inhibitors of oncogene expression and specifically impact the transformed phenotype. Finally, it must be recognized that the use of clamp ODNs reiterates Hélène’s idea that a triplex-mediated mechanism can facilitate strong, sequence-specific target recognition in the ‘antisense field’ (52).
Acknowledgments
ACKNOWLEDGEMENTS
We thank Scott Blume, Paula Bates, Vincenzo Guarcello and Scott Ebbinghaus for their vital input and expertise on this project and also Nadarajah Vigneswaran for his help with the immunohistochemical procedures. This work was supported by NCI grants R0ICA42664 and R0ICA54380 and grants from the VA Medical Research Service and US Army Prostate Research Initiative PC970218; DVIC.
References
- 1.Kelly K. and Siebenlist,U. (1986) The regulation and expression of c-myc in normal and malignant cells. Annu. Rev. Immunol., 4, 317–338. [DOI] [PubMed] [Google Scholar]
- 2.Marcu K.B., Bossone,S.A. and Patel,A.J. (1992) Myc function and regulation. Annu. Rev. Biochem., 61, 809–860. [DOI] [PubMed] [Google Scholar]
- 3.Kim H.-G. and Miller,D.M. (1995) Inhibition of in vitro transcription by a triplex-forming oligonucleotide targeted to human c-myc P2 promoter. Biochemistry, 34, 8165–8171. [DOI] [PubMed] [Google Scholar]
- 4.Cole M.D. (1986) The myc oncogene: its role in transformation and differentiation. Annu. Rev. Genet., 20, 361–384. [DOI] [PubMed] [Google Scholar]
- 5.Bishop J.M. (1983) Cellular oncogenes and retroviruses. Annu. Rev. Biochem., 52, 301–354. [DOI] [PubMed] [Google Scholar]
- 6.Riou G., Barrois,M., Le,M.G., George,M., LeDoussal,V. and Haie,C. (1987) C-myc proto-oncogene expression and prognosis in early carcinoma of the uterine cervix. Lancet, ii, 761–763. [DOI] [PubMed] [Google Scholar]
- 7.Varley J.M., Swallow,J.W., Breammar,W.J., Whittaker,J.L. and Walker,R.A. (1987) Alterations to either c-erb B-2(neu) or c-myc proto-oncogenes in breast carcinomas correlate with poor short-term prognosis. Oncogene, 1, 423–430. [PubMed] [Google Scholar]
- 8.Field J.K., Spandidos,D.S., Stell,P.M., Vaughan,E.D., Evan,G.I. and Moore,J.P. (1989) Elevated expression of the c-myc oncoprotein correlates with poor prognosis in head and neck squamous cell carcinoma. Oncogene, 4, 11463–1468. [PubMed] [Google Scholar]
- 9.Grover R., Ross,D.A., Wilson,G.D. and Sanders,R. (1997) Measurement of c-myc oncoprotein provides an independent prognostic marker for regional metastatic melanoma. Br. J. Plast. Surg., 50, 478–482. [DOI] [PubMed] [Google Scholar]
- 10.Fidler I.J. (1973) Selection of successive tumour lines for metastasis. Nature New Biol., 242, 148. [DOI] [PubMed] [Google Scholar]
- 11.Kool E. (1991) Molecular recognition by circular oligonucleotides: increasing the selectivity of DNA binding. J. Am. Chem. Soc., 113, 6265–6266. [Google Scholar]
- 12.Giovannangeli C., Thuong,N.T. and Hélène,C. (1993) Oligonucleotide clamps arrest DNA synthesis on a single-stranded DNA target. Proc. Natl Acad. Sci. USA, 90, 10013–10017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cimino G.D., Gamper,H.B., Isaacs,S.T. and Hearst,J.E. (1985) Psoralens as photoactive probes of nucleic acid structure and function: organic chemistry, photochemistry and biochemistry. Annu. Rev. Biochem., 54, 1151–1193. [DOI] [PubMed] [Google Scholar]
- 14.Shimizu M., Hanvey,J.C. and Wells,R.D. (1989) Intramolecular DNA triplexes in supercoiled plasmids. I. Effect of loop size on formation and stability. J. Biol. Chem., 264, 5944–5949. [PubMed] [Google Scholar]
- 15.Lipsett M.N. (1964) Complex formation between polycytidylic acid and guanine oligonucleotides. J. Biol. Chem., 239, 1256–1260. [PubMed] [Google Scholar]
- 16.D’Souza D.J. and Kool,E.T. (1994) Solvent, pH and ionic effects on the binding of single-stranded DNA by circular oligodeoxynucleotides. Bioorg. Med. Chem. Lett., 4, 965–970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wang S., Xu,Y. and Kool,E.T. (1997) Recognition of RNA by triplex formation: divergent effects of pyrimidine C-5 methylation. Bioorg. Med. Chem. Lett., 5, 1043–1050. [DOI] [PubMed] [Google Scholar]
- 18.Giovannangeli C., Montenay-Garestier,T., Rougée,M., Chassignol,M., Thuong,N.T. and Hélène,C. (1991) Single-stranded DNA as a target for triplex formation. J. Am. Chem. Soc., 113, 7775–7777. [Google Scholar]
- 19.Hélène C. (1991) Rational design of sequence-specific oncogene inhibitors based on antisense and antigene oligonucleotides. Eur. J. Cancer, 27, 1466–1471. [DOI] [PubMed] [Google Scholar]
- 20.Sambrook J., Fritsch,E.F. and Maniatis,T. (1989) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
- 21.Stanton L.W., Fahrlander,P.D., Tesser,P.M. and Marcu,K.B. (1984) Nucleotide sequence comparison of normal and translocated murine c-myc genes. Nature, 310, 423–425. [DOI] [PubMed] [Google Scholar]
- 22.Brown D., Yu,Z., Miller,P., Blake,K., Wei,C., Kung,H., Black,R.J., Ts’o,P.O.P and Chang,E.H. (1989) Modulation of ras expression by anti-sense, nonionic deoxyoligonucleotide analogs. Oncogene Res., 4, 243–252. [PubMed] [Google Scholar]
- 23.Ebbinghaus S.W., Vigneswaran,N., Miller,C.R., Chee-Awai,R.A., Mayfield,C.A., Curiel,D.T. and Miller,D.M. (1996) Efficient delivery of triplex forming oligonucleotides to tumor cells by adenovirus-polylysine complexes. Gene Ther., 3, 287–297. [PubMed] [Google Scholar]
- 24.Hanson K.D., Shichiri,M., Follansbee,M.F. and Sedivy,J.M. (1994) Effects of c-myc expression on cell cycle progression. Mol. Cell. Biol., 14, 5748–5755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Takasugi M., Guendouz,A., Chassignol,M., Decout,J.L., Lhomme,J., Thuong,N.T. and Hélène,C. (1991) Sequence-specific photo-induced cross-linking of the two strands of double-helical DNA by a psoralen covalently linked to a triple helix-forming oligonucleotide. Proc. Natl Acad. Sci. USA, 88, 5602–5606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Barre F.-X, Giovannangeli,C., Hélène,C. and Harel-Bellan,A. (1999) Covalent crosslinks introduced via a triple helix-forming oligonucleotide coupled to psoralen are inefficiently repaired. Nucleic Acids Res., 24, 743–749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Liquier J., Taillandier,E., Klinck,R., Guittet,E., Gouyette,C. and Huynh-Dinh,T. (1995) Spectroscopic studies of chimeric DNA-RNA and RNA 29-base intramolecular triple helices. Nucleic Acids Res., 23, 1722–1728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Xodo L.E., Manzini,G. and Quadrifoglio,F. (1990) Spectroscopic and calorimetric investigation on the DNA triplex formed by d(CTCTTCTTTCTTTTCTTTCTTCTC) and d(GAGAAGAAAGA) at acidic pH. Nucleic Acids Res., 18, 3557–3564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Grover R., Grobbelaar,A.O., Hudson,D.A., Forder,M., Wilson,G.D. and Sanders,R. (1997) The clinical significance of oncogene expression in subungual melanoma. Br. J. Plast. Surg., 50, 15–19. [DOI] [PubMed] [Google Scholar]
- 30.Stein C.A. and Cheng,Y.C. (1993) Antisense oligonucleotides as therapeutic agents: is the bullet really magical? Science, 261, 1004–1012. [DOI] [PubMed] [Google Scholar]
- 31.Gewirtz A.M. (1999) Myb targeted therapeutics for the treatment of human malignancies. Oncogene, 18, 3056–3062. [DOI] [PubMed] [Google Scholar]
- 32.Morgan R.A. (1997) Gene therapy for HIV infection. Clin. Exp. Immunol., 1, 41–44. [PubMed] [Google Scholar]
- 33.Stephan D. and Nabel,E.G. (1997) Gene and other biological therapies for vascular diseases. Fundam. Clin. Pharmacol., 11, 97–110. [DOI] [PubMed] [Google Scholar]
- 34.Holmes R.P. (1998) Pharmacological approaches in the treatment of primary hyperoxaluria. J. Nephrol., 1, 32–35. [PubMed] [Google Scholar]
- 35.Werner R.G. (1998) Identification and development of new biopharmaceuticals. Arzneimittelforschung, 48, 523–530. [PubMed] [Google Scholar]
- 36.Spencer C.A. and Groudine,M. (1991) Control of c-myc regulation in normal and neoplastic cells. Adv. Cancer Res., 56, 1–48. [DOI] [PubMed] [Google Scholar]
- 37.Collins J.F., Herman,P., Schuch,C. and Bagby,G.C.Jr (1992) c-myc antisense oligonucleotides inhibit the colony-forming capacity of Colo 320 colonic carcinoma cells. J. Clin. Invest., 89, 1523–1527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mizutani Y., Fukumoto,M., Bonavida,B. and Yoshida,O. (1994) Enhancement of sensitivity of urinary bladder tumor cells to cisplatin by c-myc antisense oligonucleotide. Cancer, 74, 2546–2554. [DOI] [PubMed] [Google Scholar]
- 39.Smith J.B. and Wickstrom,E. (1998) Antisense c-myc and immunostimulatory oligonucleotide inhibition of tumorigenesis in a murine B-cell lymphoma transplant model. J. Natl Cancer Inst., 90, 1146–1154. [DOI] [PubMed] [Google Scholar]
- 40.Warzocha K. (1999) Antisense strategy in hematological malignancies. Cytokines Cell. Mol. Ther., 5, 15–23. [PubMed] [Google Scholar]
- 41.Bazarov A.V., Adachi,S., Li,S.-F., Mateyak,M.K., Wei,S. and Sedivy,J.M. (2001) A modest reduction in c-Myc expression has minimal effects on cell growth and apoptosis but dramatically reduces susceptibility to Ras and Raf transformation. Cancer Res., 61, 1178–1186. [PubMed] [Google Scholar]
- 42.Shichiri M., Hanson,K.D. and Sedivy,J.M. (1993) Effects of c-myc expression on proliferation, quiescence and G0 to G1 transition in nontransformed cells. Cell Growth Differ., 4, 93–104. [PubMed] [Google Scholar]
- 43.Hanson K.D., Shichiri,M., Follansbee,M.R. and Sedivy,J.M. (1994) Effects of c-myc expression on cell cycle progression. Mol. Cell. Biol., 14, 5748–5755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Mateyak M.K., Obaya,A.J., Adachi,S. and Sedivy,J.M. (1997) Phenotypes of c-Myc-deficient rat fibroblasts isolated by targeted homologous recombination. Cell Growth Differ., 8, 1039–1048. [PubMed] [Google Scholar]
- 45.Miller A.D., Miller,D.G., Garcia,J.V. and Lynch,C.M. (1993) Use of retroviral vectors for gene transfer and expression. Methods Enzymol., 217, 581–599. [DOI] [PubMed] [Google Scholar]
- 46.Fang F., Orend,G., Watanabe,N., Hunter,T. and Ruoslahti,E. (1996) Dependence of cyclin E-cdk2 kinase activity on cell anchorage. Science, 271, 499–502. [DOI] [PubMed] [Google Scholar]
- 47.Zhu X.M., Ohtsubo,R.M., Bohmer,R.M., Roberts,J.M. and Assoian,R.K. (1996) Adhesion-dependent cell cycle progression linked to the expression of cyclin D1, activation of cyclin E-cdk2 and phosphorylation of the retinoblastoma protein. J. Cell Biol., 133, 391–403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Prakash G. and Kool,E.T. (1992) Structural effects in the recognition of DNA by circular oligonucleotides. J. Biol. Chem., 114, 3523–3527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wang S., Booher,M.A. and Kool,E.T. (1994) Stabilities of nucleotide loops bridging the pyrimidine strands in DNA pyrimidine·purine·pyrimidine triplexes: special stability of the CTTTG loop. Biochemistry, 33, 4639–4644. [DOI] [PubMed] [Google Scholar]
- 50.Booher M.A., Wang,S. and Kool,E.T. (1994) Base pairing and steric interactions between pyrimidine strand bridging loops and the purine strand in DNA pyrimidine·purine·pyrimidine triplexes. Biochemistry, 33, 4645–4651. [DOI] [PubMed] [Google Scholar]
- 51.Xodo L.E., Manzini,G., Quadrifoglio,F., van der Marel,G.A. and van Boom,J.H. (1991) Effect of 5-methylcytosine on the stability of triple-stranded DNA—a thermodynamic study. Nucleic Acids Res., 19, 5625–5631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Hélène C., Thuong,N.T. and Harel-Bellan,A. (1992) Control of gene expression by triple helix-forming oligonucleotides. The antigene strategy. Ann. N. Y. Acad. Sci., 660, 27–36. [DOI] [PubMed] [Google Scholar]