

Abstract
Colistin remains one of the few antibiotics effective against multi-drug resistant (MDR) hospital pathogens, such as Klebsiella pneumoniae. Yet resistance to this last-line drug is rapidly increasing. Characterized mechanisms of colR in K. pneumoniae are largely due to chromosomal mutations in two-component regulators, although a plasmid-mediated colR mechanism has recently been uncovered. However, the effects of intrinsic colistin resistance are yet to be characterized on a whole-genome level. Here, we used a genomics-based approach to understand the mechanisms of adaptive colR acquisition in K. pneumoniae. In controlled directed-evolution experiments we observed two distinct paths to colistin resistance acquisition. Whole genome sequencing identified mutations in two colistin resistance genes: in the known colR regulator phoQ which became fixed in the population and resulted in a single amino acid change, and unstable minority variants in the recently described two-component sensor crrB. Through RNAseq and microscopy, we reveal the broad range of effects that colistin exposure has on the cell. This study is the first to use genomics to identify a population of minority variants with mutations in a colR gene in K. pneumoniae.
Subject terms: Bacterial genetics, Antimicrobial resistance
Introduction
Antimicrobial resistance is an urgent threat to human health worldwide, and current treatment options are dwindling. Multi-drug resistant Klebsiella pneumoniae (MDR-Kp) was flagged by the World Health Organization and the Centre for Disease Control as “an urgent threat to human health” as it causes a range of high-mortality illnesses, including pneumonia, urinary tract infections and bloodstream infections, which are only treatable with a handful of last-line antibiotics, such as colistin. Colistin is a positively charged polypeptide antibiotic of the polymyxin class that binds to lipid A on the negatively charged bacterial cell surface, lowering the net charge, permeabilising the membrane and culminating in cell lysis1. Colistin was discovered in 1947 and first used as an antimicrobial in the late 1950s for the treatment of Gram-negative infections, but its use diminished in the 1970s due to nephrotoxicity and was replaced largely by aminoglycosides2–4. Recently, colistin was reintroduced into the clinical setting because it remains largely effective against MDR-Kp, as well as other Gram-negative bacilli including Acinetobacter baumannii and Pseudomonas aeruginosa, particularly as a component of combination therapy3,5. Worldwide colistin resistance (colR) rates are around 1.5% for K. pneumoniae, however, in some high-use countries this can reach up to 40% for MDR-Kp6.
In 2015, the first non-chromosomal colR gene, mcr-1, was identified on plasmids from food-associated Escherichia coli throughout China and now is also found in Europe7,8. Mobile genetic elements (MGEs) have also been associated with patient-to-patient spread of colR in hospitals9,10. Increased capsule production plays a role in providing antibiotic resistance in K. pneumoniae by acting as a physical barrier11,12. However, the major mechanisms of colR in K. pneumoniae are due to intrinsic mutations, which can be selected by exposure to colistin. They usually involve SNPs in the two-component system (TCS) genes phoPQ or pmrAB, where the sensor component (PhoQ/PmrB) activates the regulator component (PhoP/PmrA) via phosphorylation in response to environmental signals13. In K. pneumoniae, the regulator component of both systems control expression of the arnBCADTEF (or pmr) operon which decreases the net charge of lipid A by attaching additional sugars, specifically 4-amino-4-deoxy-L-arabinose (L-Ara4N), thus preventing the positively charged colistin from binding lipid A14–16. Similarly, the product of the PmrAB-controlled gene pmrC was shown to decorate lipid A with phosphoethanolamine to provide tolerance to colistin17. In addition, colR is also conferred by inactivation of the small lipoprotein MgrB, which represses phoPQ expression13,18–21. Multiple mutations in the sensory component of the TCS crrAB22, were recently shown to confer high level colistin resistance in Klebsiella pneumoniae23. The crrAB system acts via a modulator named crrC that interacts with PmrAB to alter arn expression24, and recently it was proposed that crrB mutations also activate a putative efflux pump to remove antibiotics from the cell25.
Heteroresistance can be defined as subsets of an otherwise isogenic bacterial population that display a range of susceptibilities to an antibiotic. More recently, a review by El-Halfawy and Valvano26 described the use of the term ‘heteroresistance’ to denote genetic changes in subpopulations of bacteria in acquired resistance to antimicrobials. Heteroresistance has been recognized as a characteristic marker of rapid resistance gain, often resulting in therapeutic failure27. However, its full impact on bacterial antibiotic resistance is poorly understood, because of the difficulty detecting bacterial subpopulations using standard laboratory methods26. Colistin heteroresistance has been phenotypically shown to occur in K. pneumoniae28 and the genetic basis of heteroresistance has been the subject of several of studies which show mutations (SNPs or tandem gene duplications) in TCSs confer colR in Gram-negative strains21,22,29,30.
Here, we explore the genetic basis of intrinsic colistin heteroresistance, which has not yet been investigated on a whole-genome level. Taking a holistic approach, due to the complex nature of colistin resistance and its significance in treatment of MDR-Kp infections, we used multiple genomic approaches to examine the molecular changes associated with spontaneous colR gain and maintenance in populations of K. pneumoniae. We combined whole genome sequencing, RNA sequencing (RNAseq) and electron microscopy to uncover the effects of colistin stress on these spontaneously colR populations to better understand the impact of increasing colistin tolerance on cells grown in vitro. We also examined the phylogenetic distribution of the crrB gene across K. pneumoniae to investigate whether it is horizontally acquired.
Results and Discussion
Directed evolution experiments reveal multiple phenotypic and genotypic paths to colR
To induce spontaneous colR, two biological replicate cultures (denoted C1 and C2) of the colistin susceptible K. pneumoniae strain Ecl831 were grown on solid media with increasing concentrations of colistin sulphate. Whole community growth (>20 colonies) was taken from the highest concentration of colistin for which there were colonies and re-plated on selective media containing a range of colistin concentrations (from 1–128 μg/ml in two-fold dilutions). This was repeated every day for five days and growth recorded (Table 1). We observed the emergence of two distinct patterns of colistin resistance (Fig. 1). Ecl8 Culture 1 (C1) became resistant to the highest colistin concentration tested (128 μg/ml) after one day of growth under colistin selection (Fig. 1A), whereas resistance for Culture 2 (C2) increased to the highest concentration (128 mg/L) over 4 days (Fig. 1B).
Table 1.
Growth patterns observed on colistin plates for Ecl8 cultures C1 and C2.
| Culture | Day | Colistin concentration (µg/ml)a | SNPs detected compared to D0 (grown + col) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | Gene | Position in gene | Ref | SNP | aa change | Proportion population (%)b | |||
| C1 | +col | D0 | +++ | +++ | ++ | − | − | − | − | − | − | − | − | − | − | − |
| D1 | +++ | +++ | +++ | +++ | +++ | +++ | +++ | + | ||||||||
| D2 | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | ||||||||
| D3 | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | phoQ c | 136 | T | C | K46Q | 100 | ||
| D4 | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | ||||||||
| D5 | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | phoQ c | 136 | T | C | K46Q | 100 | ||
| C1 | no col | D6 | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | ||||||
| D7 | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | ||||||||
| D8 | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | ||||||||
| D9 | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | ||||||||
| D10 | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | phoQ c | 136 | T | C | K46Q | 100 | ||
| C2 | + col | D0 | +++ | +++ | ++ | − | − | − | − | − | − | − | − | − | − | − |
| D1 | +++ | +++ | +++ | +++ | + | − | − | − | ||||||||
| D2 | +++ | +++ | +++ | + | − | − | − | − | ||||||||
| D3 | +++ | +++ | +++ | +++ | +++ | +++ | − | − | crrB d,e | 278 | G | A | S93N | 71.8, 71.9 | ||
| D4 | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | ||||||||
| D5 | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | crrB d | 470 | G | A | G157A | 61.5, 62.2 | ||
| crrB d | 23 | G | C | S8N | 23.8, 37.1 | |||||||||||
| C2 | no col | D6 | +++ | +++ | +++ | +++ | +++ | +++ | − | − | ||||||
| D7 | +++ | +++ | ++ | − | − | − | − | − | ||||||||
| D8 | +++ | +++ | ++ | − | − | − | − | − | none | |||||||
aBacterial growth levels in 10 μl spot denoted by: +++ = lawn growth; ++ = 20–100 colonies; + = 4–20 colonies.
bResults for SNP analysis of DNA from the duplicate cultures are given.
cAlso detected when grown without colistin selection.
dNot detected when grown without colistin selection.
eAnnotated as “rstA regulator” or BN373_26561 (from positions 2777167–2778228 in Ecl8).
NB: The bold represent when the samples were extracted for sequencing.
Figure 1.
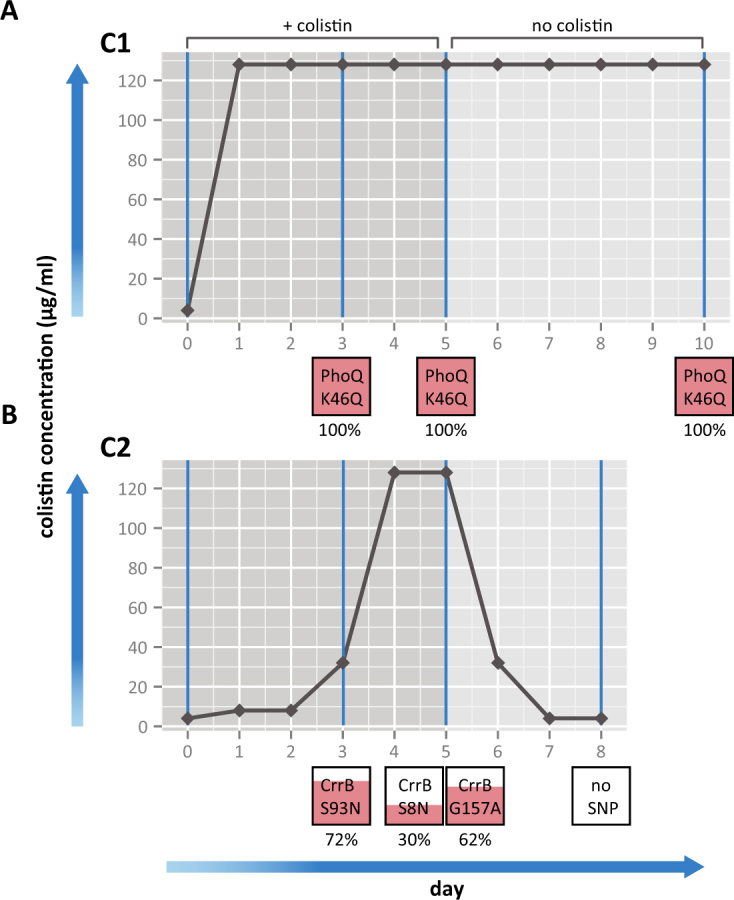
Bacterial growth with exposure to colistin sulfate and SNPs obtained at each day. The amount of bacterial growth on solid media was observed over a range of colistin concentrations (0–128 μg/ml) for 2 cultures of Ecl8 over 8–10 days for C1 (A) and C2 (B). The horizontal black lines represent bacterial growth on different concentrations of colistin over a 5 day time period, then growth after colistin selection is removed for another 5 days or until colistin sensitivity was reached. The boxes below represent the SNPs identified in each culture at D3, D5 or D10 and the amount of pink fill represents the proportion of the population harboring the SNP and this number is displayed below.
Mutations identified in phoQ and crrB by whole-genome sequencing
To understand the genotypic factors underlying colR gain, we sequenced genomic DNA to identify SNPs in the Ecl8 C1 and C2 cultures at specific time points, day 3 (D3) and day 5 (D5). DNA was extracted from duplicate liquid cultures grown for a further day with and without colistin, and compared to the colistin naïve starting cultures at day 0 (D0) grown without selection (See Fig. 1 and Table 1). It was apparent that culture C1 possessed a SNP within phoQ (an A to G substitution at base position 136) at both time points (D3 and D5), with or without selection. This SNP represents a novel non-conservative amino acid change in PhoQ, with a positively charged lysine replacing a negatively charged glutamine at codon 46 (K46Q). This K46Q substitution is in close proximity to a mutation (T48I) previously shown to provide polymyxin resistance via loss of function of the PhoQ phosphatase domain, eliminating the ability of PhoQ to inactivate the PhoP activator32,33. We hypothesize that this K46Q substitution in PhoQ similarly disrupts the phosphatase domain, allowing continuous PhoP activation of downstream genes such as those that modify lipid A, thereby providing colistin resistance.
The consensus sequence derived from pooled cells of culture C2 from D3 and D5 was identical to that of the sequenced D0 colistin naïve cells for both of the duplicate extractions when using standard SNP calling methods based on mapping (present in >75% of reads, quality score >30; as in34). However, when alternative de novo assembly-based methods were used, three SNPs were identified in a subset of the sequence reads from the C2 cultures at D3 and D5 grown in the presence of colistin (see methods). When the mapping-based SNP detection cutoffs were relaxed (bases mismatching the reference and present in >5% of reads were considered), these three additional SNPs could be confirmed. However, these SNPs were not detected in the same cultures re-grown in the absence colistin selection by either variant calling method, nor were they present at any detectable level in the D0 starting cultures. All three SNPs detected in C2 fell within a single gene (locus ID BN373_26561; Table 1). The gene, BN373_26561, was 99% identical to crrB, previously shown to confer colR22,23,25.
In C2 at D3, one SNP at position 278 bp within crrB gene (amino acid change S93N) was present in ~72% of the sequence reads (Fig. 2). Two other SNPs were detected in the C2 cells collected at D5. These were located at positions 23 and 470 bp of the crrB gene and were represented in 30% and 62% of reads and resulted in the non-synonymous changes G157A and S8N, respectively (Table 1; Fig. 2). Due to the short sequence read length, it was not possible to determine whether these 2 SNPs were present together in the same cell. None of these 3 SNPs in crrB have previously been associated with colR and so we cannot be certain they are responsible for the colR phenotype. However, this would be consistent with earlier findings that SNPs throughout the crrB gene can confer colR24.
Figure 2.
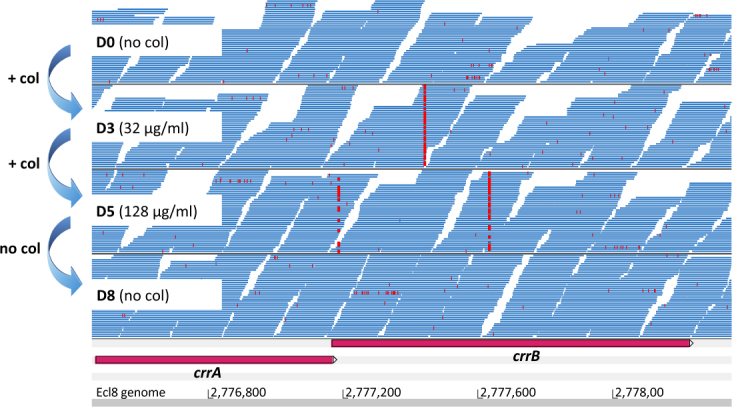
SNPs found in reads from heteroresistant C2 population in crrB. Screen shot of Artemis display with Bam files of the DNA sequencing reads, shown in blue, from D0 (colistin naïve), D3, D5 (colistin resistant) and D8 (colistin resistance lost) C2 cultures where bases differing from the reference (SNPs) are marked in red. The bp positions of the Ecl8 genome are shown below.
Single colonies were isolated from the stored bulk C2 cultures from D3 and subjected to PCR and DNA sequencing of the crrB gene. Two colonies containing the SNP at 278 bp in crrB were identified and these were subjected to whole genome sequencing. Both colonies had a single non-synonymous SNP in crrB (S93N) compared to the Ecl8 wild-type and no additional mutations. These 2 colonies were tested for colistin susceptibly, both had an MIC of >128 μg/ml, which is 16x greater than the wild-type. Thus, we have confirmed that one of these single SNPs in crrB it is associated with a high level of colistin resistance in this K. pneumoniae strain. One isogenic mutant (“colony 9”) was used for further experimentation (see methods).
Unfortunately, after screening over 100 single colonies from stored frozen C2 cultures taken at D5, using PCR and DNA sequencing, we were unable to retrospectively recover any additional individual mutants: all possessed the wild type allele and not the crrB SNPs associated with resistance at D5 from the pooled cells. Thus, we could not perform additional experiments to prove that the minority populations of crrB mutants observed are causative of the colistin resistance phenotype observed. However, taken together with the evidence that mutations throughout crrB gene are proven to confer colistin resistance in K. pneumoniae23 and that a heteroresistant population of the phoQ mutation has also been shown to give a colR phenotype in K. pneumoniae29, it is likely that this is the case.
Stability of mutant populations associated with colR
To assay for the stability of the phoQ and crrB mutations in both the C1 and C2 colR cultures, cells taken at D5 were re-passaged 5 times without antibiotic selection (Fig. 1). Susceptibility assays revealed C1 cultures to be phenotypically resistant (at 128 μg/ml) until D10. Whole genome sequencing (WGS) confirmed the presence of the mutation at bp 136 in phoQ, indicating that the phoQ mutation is stably maintained without selection.
Conversely, cells taken from C2 became fully susceptible to colistin after a single passage in the absence of selection at D6 (Fig. 1). None of the mutations seen previously in crrB were identified in the sequence of C2 cells cultures grown in the absence of selection when subjected to WGS. To determine whether the mutant was simply growing more slowly than wild type, a comparison of growth rates was performed using the method described in Hall et al.35. There was no significant difference (two-tailed student t-test, P < 0.1) in growth rate between the single crrB (S93N) mutant “colony 9” (3.38 × 10−3 OD/min, maxOD600: 0.665) and the wild-type Ecl8 (3.37 × 10−3 OD/min, maxOD600: 0.646). Thus, the reason that the crrB mutants were undetectable from culture after selection remains unclear and was not investigated further.
Transcriptomic and morphological responses to colistin in heterogeneous populations
To further characterize the overall responses of the C2 heterogeneous populations, we grew the C2 cultures in ¼ MIC of colistin (D3; 16 mg/L, MIC: 64 mg/L and D5 64 mg/L, MIC; ≥128 mg/L) to mid-log phase to extract the RNA. Due to the remarkable out competition of the crrB gene SNP variant by wild-type genotypes we could not directly assay the overall transcriptional effect of the heterogeneous population without colistin stress. Therefore, we performed RNAseq of C2 at D3 and D5 exposed to colistin as well as the D0 colistin naïve starting culture without colistin. Growing the cells at ½ MIC (D3; 32 mg/L and D5; 128 mg/L) severely retarded growth of the cultures in broth, we assume due to the heterogenous nature of this resistance mechanism, therefore we opted to grow the cultures in ¼ MIC to obtain enough cell numbers for RNA extraction.
We compared the transcriptomic differences between the heterogeneous C3 and C5 populations and the colistin-naïve Ecl8 starting population (D0). Of the 4202 total Ecl8 genes assayed in C2, we detected 389 and 226 differentially expressed genes at D3 and D5, respectively, compared to D0. (Table S1, Dataset S1). Genes showing increased expression in the C2 heterogeneous populations at D3 and D5 included all the genes in the arnA-T operon involved in lipid-A modification. Other notable increases were four genes surrounding crrB (Fig. S1), including a response regulator (crrA), crrC (BN373_26541), a glycosyl transferase (BN373_26571) and an acridine efflux pump (BN373_26531) all had a log2FC of up to 9 (over 500-fold). Of the 27 known efflux systems in Ecl8, 4 showed increased expression under colistin selection: BN373_11321 (RND-family), BN373_15271 (MacA) and BN373_36071 (RND-family). These efflux systems all have been shown to act as multi-drug transporters36. Conversely, genes encoding the porins OmpA and OmpC, which allow passive antibiotic uptake37 showed decreased levels of expression in C2 at D3 and D5 (log2FC of −1.5 and −2 respectively). The European Nucleotide Accession (ENA) accessions for raw data for all genes and conditions can be found in Table S2.
Gene Ontology (GO) classification38 was used to summarize global changes in the transcriptome during colistin exposure. The most enriched GO term for increased expression in these cultures (summarised in Fig. 3A) was the “lipopolysaccharide biosynthetic process” (GO:0009103) and “quinone binding” (GO:0048038). The latter is consistent with a recently described secondary mechanism of action of polymyxins against Gram-negative bacteria which involves the inhibition of essential inner membrane respiratory enzymes such as NADH-quinone oxidoreductase39. Core metabolic functions showed a decreased expression in the “cobalamin biosynthetic process” (GO:0009236), relating to genes associated with vitamin B12 biosynthesis. Some unexpected functional groups were affected, spanning housekeeping, metabolic and morphological function, including: “regulation of cell shape” (GO:0008360), “cell cycle” (GO:0007049), “fatty acid biosynthetic process” (GO:0006633) and “ribosome” (GO:0005840), suggesting a broad downstream effect on the cell beyond specific colistin resistance mechanisms.
Figure 3.

Overall effects of colistin exposure on colistin resistance Ecl8 cultures. (A) GO Terms derived from total differentially expressed genes in colR heteroresistant poplution of C2. C2 D3, D5 grown with colistin selection compared to D0 colistin naïve cultures to give colistin exposure and resistance, and D3, D5 grown with colistin compared to the same day grown without colistin selection to give colistin stress effects. The represented GO terms had a GSEA FDR-corrected p-value less than 0.01 in any comparison. Heatmap colors represent −log10 p-values obtained from GSEA analysis of differentially expressed genes. Red colors indicate GO terms with higher expression relative to the comparator, blue colors lower expression. (B) Scatter plot of expression changes compared to average expression of single the SNP crrB mutant grown with and (C) without colistin exposed. Each black dot represents a gene, and genes belonging to relevant enriched pathways are marked in the colour given by the key. The fold change cut-offs used are marked with a black line.
Morphological changes during colistin exposure
To further elucidate how the C2 colistin heterogeneous populations responded to colistin exposure, cells were viewed under transmission electron microscopy (TEM) to look for alterations in cell morphology (Fig. S2). Cells from the C2 heterogeneous population, taken from the plate with the highest concentration colistin at D5 were compared to colistin naïve cells from the D0 inoculation plate. C2 heterogeneous colR populations when exposed to colistin revealed loss of membrane integrity and membrane blebbing. Other signs of stress were also apparent in these cells, including the presence of phage particles in the media and surrounding the bacterial cell membrane in addition to fimbriae. Also of note were the dark ion-dense granules in the colistin exposed C2 heterogeneous colR. These granules are known to accumulate at the end of active growth and are used for storage of excess sulphur, phosphate, carbohydrates and metal ions during phosphate or nitrogen starvation40. In addition the outline of extracellular polysaccharide (EPS) was observed more frequently in colR cells.
Many of these TEM observations correlated with the RNA-seq data (see Fig. S2). This included increased expression of the Type 1 fimbriae biosynthetic cluster fimACDFG and genes from two of the three potential Ecl8 prophage (lambda and P2-like phages (BN373_1491-921 and BN373_33401-961 respectively;41–43), up to a log2FC of 7, or 128 fold increase (Dataset S1)44. Ion transport genes such as mgtA (BN373_02651) for magnesium transport or iron-chelate ABC transporters fepAC, and 4 genes of a FeCT family iron-chelate uptake transporter operon (BN373_10811-41) displayed increased expression, potentially related to the storage granules seen under TEM. As expected, OM proteins, involved in maintaining membrane integrity and stability during membrane stress, such as slyB and yfgL also showed increased expression, as well as general stress response genes, such as dnaJK. Overexpression of slyB, results in increased membrane permeability and the uptake of non-specific siderophores, a response related to stress response45,46. Although EPS production was observed in the TEM images, no transcriptomic changes were observed in the locus responsible for K2-like capsule formation (BN373_30991 to 31181).
RNAseq on the crrB SNP mutant illustrates the colR mechanism
To determine the specific effects of the crrB colR SNP, we compared the transcriptome of the single crrB mutant, “colony 9” (crrB::S93N) to the wild type Ecl8, both grown in the presence and absence of selection (Fig. 3B,C). The only gene to have highly increased expression (>256 fold) in the crrB::S93N mutant without colistin exposure was crrC (Fig. 3C). This supports the colR mechanism proposed by Cheng and colleagues24, where crrB activates the adjacent gene, crrC, and subsequently downstream lipidA modification genes via the pmrAB pathway. We also observed an increase in expression of between 2–4 fold of the arn lipid A modification genes as well as the other genes proximal to crrB (BN373_26531 and BN373_26571) predicted to encode an acridine efflux pump and glycosyltransferase, showing that these genes are activated in the crrB::S93N mutant. Non-synonymous mutations in crrB have previously described the increase in expression of lipid A modification genes, crrC and a putative glycosyltransferase, which were attributed to contribute to colR 22,24.
Using GO terms to summarise these data: Lipid A biosynthesis genes showed increased expression (boxed in Fig. 3B) and conversely, a decreased expression in genes involved in siderophore transport and cobalamin biosynthesis. The specific reasons why these pathways were affected remains unknown, although recent work has shown that acquisition of colR via pmrB mutations in A. baumannii results in decreased growth in iron limiting conditions47 and also that cobalamin synthesis and siderophore pathways interact as they compete for TonB-mediated transport across the outer membrane (OM)48.
When the crrB::S93N mutant was exposed to colistin (at 128 μg/ml) and compared to Ecl8 (grown in the absence of selection), crrC and a neighboring gene (BN373_26571) predicted to encode a glycostransferase, had increased expression (~256 fold). Also 9/16 genes in the ethanolamine utilization (eut) operon, encoding the ethanolamine catabolic pathway, showed decreased expression. This potentially makes more phosphoethanolamine available to be added to Lipid A by PmrC to change the cell charge49 associated with polymyxin resistance. Consistent with this pmrC also showed an increase in expression (9.8 fold).
Consistent with data from the heterogeneous culture, RNAseq of the crrB::S93N mutant showed an overall increase in Lipid A biosynthesis genes, as well as more pleiotropic effects including oxidative stress, changes in ribosomal genes as well as on core cellular processes.
Phylogenetic distribution of crrB within Klebsiella pneumoniae
Previously crrB was found in only in a subset of K. pneumoniae24 contrasting with other TCS sensors that mediate colR, like phoQ, present in all K. pneumoniae. Moreover, it was hypothesized that crrB was acquired by K. pneumoniae via lateral gene transfer, due to its relatively low GC content22. To understand the significance of the crrB gene at the population level, we screened a global collection of representative Klebsiella isolates from a range of niches and sample types, including K. pneumoniae and the closely related species K. quasipneumoniae and K. variicola (dataset described in50).
The crrB gene was detected in 54% (151/280) of all genomes screened (Dataset S2) including community and hospital isolates. However, of these crrB-containing isolates, 94% (142/151) belonged to K. pneumoniae sensu stricto (KpI) the species most often associated with human infection (50; Dataset S2). The frequency K. quasipneumoniae and K. variicola in crrB-containing isolates was only 5% and 1%, respectively. The phylogenetic distribution of crrB in the K. pneumoniae is shown in Fig. 4A, and reveals that the vast majority of strains carried this gene, including hospital-associated sequence types ST258/11, ST65 and ST48. However, crrB is only sporadically present in a single clade, including clinically important STs (ST23, ST14/15 and ST43), due to what appears to be a common basal deletion (shaded in Fig. 4A), indicating that crrB is not universally present in important disease causing STs.
Figure 4.

Phylogenetic tree of a KpI from a Global Klebsiella collection. (A) The dots on the ends of the branches represent whether the crrB gene is present in each isolate, red denotes lineages A and blue denotes lineage B. The boxed numbers on the outer ring shows the sequence type. The clade with KpI that has lost the crrB gene is shaded in beige. The grey lines represent an identified transfer of crrB and the arrow shows the inferred direction. (B) The 2 separate lineages of the crrB gene, based on SNP differences within the gene, are marked with blue and red colouring.
In Ecl8, the crrAB operon is located within an ~11 kb region with a skewed GC-content (44.1% compared to the average of 56.9%; Fig. S1). This region includes an insertion sequence element, IS101 as well as additional genes that appear transcriptionally coupled to crrB. Across the different lineages, it appears that this crr locus was exchanged as a single unit between lineages having been repeatedly lost and re-acquired. However, there are no mobility functions, apart from the transposase associated with IS101 and so it is likely that exchange occurs through homologous recombination between the conserved regions flanking this locus.
The phylogeny of the crrB gene itself shows there are two major clades of the crrB gene (Fig. 4B) which is incongruent with the K. pneumoniae phylogeny (Fig. 4A) suggesting that additionally crrB alone is broadly exchanged across distant K. pneumoniae lineages by recombination. In support of this, we identified likely donors for two distinct horizontal gene transfer events that may explain the re-introduction of the crrB gene into the ostensibly crrB-negative clade (Fig. 4A). Taken together, these data suggest that crrB is ancestral to K. pneumoniae, was subsequently lost by deletion in one clade and the subject of intra-species recombination. No evidence of lateral gene transfer via mobile genetic elements was observed.
Conclusions
In vitro induction of colR resistance in K. pneumoniae identified known and novel SNPs in two genes known to confer colistin resistance. At the genotypic level we identified a novel SNP in, phoQ, as well as mutations in the crrB gene. One SNP, in phoQ, was present in 100% of the population, and was stable after selection was removed. This contrasted with the heterogeneous cultures, which carried mutations in the crrB gene. The crrB mutants were present in a maximum of ~70% of the total population under selection, but importantly never reached fixation and were not maintaining in the population after selection was removed. We show that exposing the crrB heterogeneous cultures to increasingly higher concentrations of colistin resulted in a range of transcriptional changes in core cellular functions, which correlated with morphological changes in the cells themselves. These far-reaching effects may explain why we do not observe colR populations harboring crrB SNPs reaching fixation, presumably being outcompeted by isolates with a wild type crrB genotype in the absence of colistin. Certainly in other systems, susceptible wild-type genotypes were shown to outcompete the colR variants in a heterogeneous population, including pmrAB variants in Acinetobacter spp.51–53. However, in our study, we found no significant difference in growth between the isogenic crrB SNP colR mutant compared to a wild-type crrB carrying isolate. Further work is required to determine why these mutants are not maintained in the population and appear to be out-competed by the wild-type without selection in culture. The genetic basis of heteroresistance has only recently been described in several publications which show that mutations (SNPs or gene duplication) in TCSs, in a subset of the population, result in a heteroresistance phenotype. In most cases these result from alterations in expression in the lipid A modification genes29,30,54, whilst in another study no genetic basis was found to explain heteroresistance55. However in the later case, Band et al.55 observed a loss of colR upon removal of selection thereby concluding it to be due to an unstable mutation. We propose that the heteroresistance mechanisms observed here mediated by mutations in crrB in only a subset of the population, may offer a new mechanism of achieving resistance during colistin exposure, whereby the mutant subpopulation would be quickly replaced by the wild-type strains in the absence of selection resulting in a reversion to pre-treatment sensitive levels as was observed by Band et al.55. We however could not determine the exact mechanism of heteroresistance, the increase in expression of lipid-A modification genes in the crrB heterogenous populations may explain the reason for the apparent loss of genotype and subsequent phenotype in the absence of selection. In addition, we observed the production of an EPS structures in the TEM images but no genetic mechanism to explain this phenotype. Perhaps the EPS provides a physical barrier, however further work will have to be done to prove this.
Although the phenotype is unstable, the potential to gain complete colR upon exposure to the drug have further clinical implications with respect to the rapid nature of colR acquisition by this mechanism.
Phylogenetic analysis of the distribution of crrB indicated that it was acquired earlier due to its basal position in K. pneumoniae sensu stricto, the principle species associated with human disease, but was lacking from the ancestors of the closely related species K. variicola and K. quasipneumoniae. Moreover, crrB was lost from one K. pneumoniae clade that includes important sequence types associated with liver abscesses and neonatal sepsis (ST23) as well as hospital associated MDR sequence types (ST14, ST15). Unlike phoQ, whilst there may be an ancestral function of crrB, it is not currently part of the core K. pneumoniae genome.
Finally, this work complements the recent discovery that a colistin resistance gene (mcr-1) can be carried on mobile extrachromosomal plasmids7,55 and illustrates that homologous recombination is another important mechanism facilitating the transfer of colR genes (such as crrAB). This finding is particularly worrying and should reinforce the need to preserve our current effective antibiotics and dedicate resources to developing new ones.
One major limitation of our study is that because we did not to perform single gene knockouts and complementation studies, we could not definitively prove that the crrB minority variants were causative of the colistin resistance phenotype observed. Further, the instability of the crrB mutants without selection limited our ability to test the dynamics of these mutants in a population. However, we are continuing work to fully investigate the effect of crrB mutations on heteroresistant populations.
Methods
Inducing colistin resistance
Minimum Inhibitory Concentrations (MIC) of duplicate cultures of a colistin-sensitive ampicillin-resistant non-mucoid Klebsiella pneumoniae strain, Ecl8 (31; GenBank accession number HF5364824) and also two single colonies of the 258 bp crrB SNP mutant was determined using the British Society for Antimicrobial Chemotherapy (BSAC) agar dilution method56. To induce colistin resistance, duplicate cultures of Ecl8 were passaged daily in increasing concentrations of colistin sulfate from 1–128 μg/ml in serial dilutions on iso-sensitest agar (Oxoid), until they reached clinical colR (128 μg/ml) over 5 days (~150 generations). We used iso-sensitest media throughout, as it has found to be most suitable for detecting colistin heteroresistance57. Approximately 109 cells of initial inoculum was resuspended in 100 μl 0.9% saline, and 3 μl of this was spotted onto the colistin-containing agar plates which were grown overnight at 37 °C, and 10 μl was used to inoculate iso-sensitest broth for initial nucleic acid extraction (D0). Growth was taken from the highest concentration colistin plate where at least 20 colonies had growth and the entire colony spot was resuspended in 100 μl 0.9% saline and respotted onto another set of plates. At nucleic acid extraction days (D3, D5), 10 μl of the saline resuspension was also used to inoculate iso-sensitest broth + 1/2 MIC colistin and RNA was extracted mid-log phase, grown overnight, and DNA extracted. We also included positive and negative growth controls, to ensure viability of the selection.
Electron Microscopy
The morphology of C2 colR cells, taken from the 128 μg/ml colistin D5 plate, were compared to colistin naïve cells, taken from the colistin naïve starting plate at D0, by negative staining and fine structure analysis of ultrathin sections using transmission electron microscopy (TEM), on a Spirit Biotwin 120-kV TEM (FEI) and imaging with a F415 charge-coupled-device (CCD) camera (Tietz) For negative staining, cells were mixed with water and applied to a freshly glow-discharged, Formvar/carbon-coated EM grid and an equal volume of 3% ammonium molybdate with 1% trehalose. For the ultrathin sectional stains, cells were fixed with ruthenium hexamine trichloride (RHT) for 1 hour at room temperature in 2.5% glutaraldehyde in 0.05 M sodium cacodylate buffer with 0.7% RHT and 50 mM L-lysine monohydrochloride, followed by 3 buffered rinses, then re-fixed in 1% osmium tetroxide, dehydrated in an ethanol series (staining with 2% uranyl acetate at the 20% stage) and embedded in Epon araldite resin (Sigma). 50 μm ultrathin sections were cut on a UCT ultramicrotome EM (Leica), contrasted with uranyl acetate and lead citrate and analysed.
Whole Genome Sequencing and SNP-calling
Whole community growth spots were resuspended in 100 µl PBS. 10 µl of this was used to inoculate cultures of 10 ml of iso-sensitest with 1/4 MIC colistin. DNA was extracted using a Promega Wizard Extraction Kit (Promega) and sequenced on an Illumina HiSeq2000 sequencer with 150 cycle paired-end runs. FASTQ files were deposited in the ENA, for accession number see Table S2. Sequences of isolates from D3, D5, and D10 were compared against their parental isolate D0 for C1 and C2, respectively to identify genomic variants conferring colistin resistance. SNP and indel variants were detected as previously described58 via a de novo assembly using SGA v0.9.1959 with built-in variant calling as well as a mapping approach using SMALT v0.7.4 with subsequent SNP calling34. To compare results between both methods variants were mapped back against Ecl8. Resulting variants were checked manually for accuracy. Single colonies of C2 from D3 were plated onto colistin containing agar and use as template DNA a PCR for the crrB gene (Primers in Table S3). We sequenced PCR products from 20 colonies, and 2 colonies containing the crrB SNP were subjected to whole genome sequencing on a 150 bp paired-end MiSeq run (Accession numbers in Table S1).
Phylogenetic characterisation of crrB within K. pneumoniae
A phylogenetic analysis of crrB was performed to identify lineages of the gene. Sequence reads of a set of 328 population-wide isolates from Holt et al.60 were assembled de novo using an in-house assembly pipeline utilising Velvet v1.0.1261 and Velvet optimizer for initial contig assembly, scaffolding of contigs using SSPACE v2.062, filling of gaps using GapFiller v1.1163, and a final mapping of reads against the scaffolds using SMALT v0.75 (H. Ponstingl and Z. Ning, manuscript in preparation; http://www.sanger.ac.uk/resources/software/smalt/). The presence of crrB was determined using a BLASTn search with a 98% cutoff as well as an in-house script which uses SMALT v0.75 to map the sequence of crrB back to the contigs using a 90% nucleotide acid identity cutoff. To detect recombination events in the crrB gene region, genomes were de novo assembled using SPAdes64 and suspected recombinant sequences were queried in against all genome assemblies using BLASTn65. High identity alignments from phylogenetically distant isolates were used to identify candidate recombinant DNA donors. Candidate donor sequences were then aligned to the query sequence using NUCmer66, SNP density was established, and the results were analysed with the Artemis Comparison Tool67. Alignments containing regions of very low SNP density compared to that across the rest of the genome were taken as evidence of recombination. Loss of the crrB gene due to recombination was further confirmed using the SPAdes assembly graphs. Bandage68 was used to view the assembly graphs and locate insertion sequences within crrB genetic context. Losses of the crrB gene due to nonsense mutation were identified using SeaView69. Phylogenetic analysis of the crrB gene was performed using RAxML70 to identify lineages (Fig. 4B). The two major clades of crrB were then plotted against the K. pneumoniae core genome phylogenetic tree generated previously50 and annotated with the inferred recombination events (Fig. 4A).
RNAseq
RNA was extracted at OD 0.5 +/− 0.05, from the same D3, D5 and D0 C2 cultures as used in DNA extraction for SNP calling. Duplicate independent cultures were grown for each condition and time point to give technical replicates. RNA was extracted using a modified chloroform-based method (as previously described71) and depleted for ribosomal RNA (Ribo-ZeroTM Magnetic kit, Epicentre) prior to non-stranded sequencing using the TruSeq Illumina protocol. Samples were subsequently run on an Illumina HiSeq2500 sequencer for which an average of 1.75 Gb of raw reads in 100 base read pairs was obtained for each sample. The resulting FASTQ files were mapped against the Klebsiella pneumonia Ecl8 genome31. QPCR was used to confirm the trajectory of the fold-changes of 11 genes observed by RNAseq with primers listed in Table S3, as previously described72.
RNA was extracted at mid-log growth, from the same cultures as used for DNA extraction for whole genome sequencing. Due to the remarkable out competition of the crrB gene SNP variant by wild-type genotypes we could not directly assay the overall transcriptional effect of the heterogeneous population without colistin stress. Therefore we performed RNAseq of C2 at D3 and D5 exposed to colistin as well as the D0 colistin naïve starting culture. Transcriptomic analysis was also performed on the single crrB SNP mutant to determine the colR mechanism, this mutant, grown in colistin, was compared to the wild-type strain grown in sub-inhibitory (1/4 MIC) concentrations of colistin.
Differential expression analysis
Read counts per feature were aggregated and further analysis was conducted in R. Read counts were normalized using the TMM normalization implemented in the EdgeR package version 3.4.273. The Voom transformation74 was applied to the normalized read counts and differential expression analysis was performed using the Limma package version 3.18.1375. Read counts from C2 colR cultures (D3 and D5) were compared to those from colistin naïve cultures (D0) to determine changes in gene expression after SNP acquisition and during colistin stress. RNA from each culture grown with and without colistin selection was also compared. Differentially expressed genes with a log2 fold change (logFC) of >1 (increased expression) or a logFC < −1 (decreased expression) and a Q-value < 0.05 were considered significant for this analysis.
Functional analysis of differentially expressed genes
Slimmed gene ontology (GO) terms were obtained for K. pneumoniae str. Ecl8 using the QuickGO interface76,77. Redundant entries were removed and Gene Set Enrichment Analysis (GSEA) analysis was then performed38 using GSEA version 2.1.0, run in ranked-list mode with enrichment statistic weighted on the per-gene logFCs calculated by voom, and GO-terms with a FDR corrected p-value less than 0.01 in any comparison are displayed in Fig. 3A.
Availability of data and material
All sequence data is available at the European Nucleotide Archive (ENA) (http://www.ebi.ac.uk/ena) under the study accession number ERP004913. Specific sample accession numbers are given in Table S2. Other data generated or analysed during this study are included in this published article and its supplementary information files.
Electronic supplementary material
Acknowledgements
We would like to thank and acknowledge Thamarai Schneiders for her kind gift of the Ecl8 strain. This work was supported by the Wellcome Trust grant number WT098051. The salaries of AKC and CJB were supported by the Medical Research Council [grant number G1100100/1] and MJE is supported by Public Health England. LB is supported by a research fellowship from the Alexander von Humboldt Stiftung/Foundation. KEH is supported by the NHMRC of Australia (Fellowship #1061409).
Author Contributions
A.K.C. wrote the manuscript with support from C.J.B., L.B., J.D., J.P. and N.R.T. and all authors contributed to editing. A.K.C., C.J.B., L.B., J.P., M.E. and N.R.T. were involved in the study design. A.K.C., C.J.B., M.M., M.F., D.P. and D.G. performed the laboratory experiments. L.B., J.D., K.E.H., R.R.W., C.J.B. and A.K.C. performed the WGS, phylogenetic and RNAseq data analysis.
Competing Interests
The authors declare no competing interests.
Footnotes
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Amy K. Cain and Christine J. Boinett contributed equally to this work.
Change history
7/29/2021
The original online version of this Article was revised: In the original version of this Article the email addresses for authors Amy K. Cain and Christine J. Boinett were interchanged. Correspondence and requests for materials should be addressed to ac19@sanger.ac.uk for Amy K. Cain and to cb19@sanger.ac.uk for Christine J. Boinett.
Contributor Information
Amy K. Cain, Email: ac19@sanger.ac.uk
Christine J. Boinett, Email: cb19@sanger.ac.uk
Nicholas R. Thomson, Email: nrt@sanger.ac.uk
Electronic supplementary material
Supplementary information accompanies this paper at 10.1038/s41598-018-28199-y.
References
- 1.Velkov T, et al. Surface changes and polymyxin interactions with a resistant strain of Klebsiella pneumoniae. Innate Immun. 2014;20:350–63. doi: 10.1177/1753425913493337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Nation RL, et al. Consistent Global Approach on Reporting of Colistin Doses to Promote Safe and Effective Use. Clin Infect Dis. 2014;58:2013–2015. doi: 10.1093/cid/cit680. [DOI] [PubMed] [Google Scholar]
- 3.Falagas ME, Kasiakou SK. Colistin: the revival of polymyxins for the management of multidrug-resistant gram-negative bacterial infections. Clin. Infect. Dis. 2005;40:1333–1341. doi: 10.1086/429323. [DOI] [PubMed] [Google Scholar]
- 4.Li J, et al. Colistin: the re-emerging antibiotic for multidrug-resistant Gram-negative bacterial infections. Lancet Infectious Diseases. 2006;6:589–601. doi: 10.1016/S1473-3099(06)70580-1. [DOI] [PubMed] [Google Scholar]
- 5.Biswas S, Brunel J-M, Dubus J-C, Reynaud-Gaubert M, Rolain J-M. Colistin: an update on the antibiotic of the 21st century. Expert Review of Anti-infective Therapy. 2012;10:917–934. doi: 10.1586/eri.12.78. [DOI] [PubMed] [Google Scholar]
- 6.Capone, a. et al. High rate of colistin resistance among patients with carbapenem-resistant Klebsiella pneumoniae infection accounts for an excess of mortality. Clin. Microbiol. Infect. 19 (2013). [DOI] [PubMed]
- 7.Liu Y, et al. Articles Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect. Dis. 2015;3099:1–8. doi: 10.1016/S1473-3099(15)00424-7. [DOI] [PubMed] [Google Scholar]
- 8.Skov RL, Monnet DL. Plasmid-mediated colistin resistance (mcr-1gene): three months later, the story unfolds. Eurosurveillance. 2016;21:1–6. doi: 10.2807/1560-7917.ES.2016.21.9.30155. [DOI] [PubMed] [Google Scholar]
- 9.Marchaim D, et al. Outbreak of colistin-resistant, carbapenem-resistant Klebsiella pneumoniae in Metropolitan Detroit, Michigan. Antimicrob. Agents Chemother. 2011;55:593–599. doi: 10.1128/AAC.01020-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Arduino SM, et al. Transposons and integrons in colistin-resistant clones of Klebsiella pneumoniae and Acinetobacter baumannii with epidemic or sporadic behaviour. J. Med. Microbiol. 2012;61:1417–1420. doi: 10.1099/jmm.0.038968-0. [DOI] [PubMed] [Google Scholar]
- 11.Pan, Y. J. et al. Capsular types of Klebsiella pneumoniae revisited by wzc sequencing. PLoS One8 (2013). [DOI] [PMC free article] [PubMed]
- 12.Arakawa Y, et al. Genomic organization of the Klebsiella pneumoniae cps region responsible for serotype K2 capsular polysaccharide synthesis in the virulent strain chedid. J. Bacteriol. 1995;177:1788–1796. doi: 10.1128/jb.177.7.1788-1796.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Olaitan AO, Morand S, Rolain J-M. Mechanisms of polymyxin resistance: acquired and intrinsic resistance in bacteria. Front. Microbiol. 2014;5:1–18. doi: 10.3389/fmicb.2014.00643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Helander IM, et al. Characterization of lipopolysaccharides of polymyxin-resistant and polymyxin-sensitive Klebsiella pneumoniae O3. Eur. J. Biochem. 1996;237:272–278. doi: 10.1111/j.1432-1033.1996.0272n.x. [DOI] [PubMed] [Google Scholar]
- 15.Gunn JS, et al. PmrA-PmrB-regulated genes necessary for 4-aminoarabinose lipid A modification and polymyxin resistance. Mol. Microbiol. 1998;27:1171–1182. doi: 10.1046/j.1365-2958.1998.00757.x. [DOI] [PubMed] [Google Scholar]
- 16.Mitrophanov, A. Y., Jewett, M. W., Hadley, T. J. & Groisman, E. a. Evolution and dynamics of regulatory architectures controlling polymyxin B resistance in enteric bacteria. PLoS Genet. 4 (2008). [DOI] [PMC free article] [PubMed]
- 17.Kim SH, Jia W, Parreira VR, Bishop RE, Gyles CL. Phosphoethanolamine substitution in the lipid A of Escherichia coli O157: H7 and its association with PmrC. Microbiology. 2006;152:657–666. doi: 10.1099/mic.0.28692-0. [DOI] [PubMed] [Google Scholar]
- 18.Cannatelli A, et al. In vivo emergence of colistin resistance in Klebsiella pneumoniae producing KPC-type carbapenemases mediated by insertional inactivation of the PhoQ/PhoP mgrB regulator. Antimicrob. Agents Chemother. 2013;57:5521–5526. doi: 10.1128/AAC.01480-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Poirel L, et al. The mgrB gene as a key target for acquired resistance to colistin in Klebsiella pneumoniae. J. Antimicrob. Chemother. 2014;70:75–80. doi: 10.1093/jac/dku323. [DOI] [PubMed] [Google Scholar]
- 20.The, H. C. et al. A high-resolution genomic analysis of multidrug- resistant hospital outbreaks of Klebsiella pneumoniae. EMBO Mol Med. 7, 227–239 (2015). [DOI] [PMC free article] [PubMed]
- 21.Zowawi HM, et al. Stepwise evolution of pandrug-resistance in Klebsiella pneumoniae. Sci. Rep. 2015;5:15082. doi: 10.1038/srep15082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Wright MS, et al. Genomic and Transcriptomic Analyses of Colistin-Resistant Clinical Isolates of Klebsiella pneumoniae Reveal Multiple Pathways of Resistance. Antimicrob. Agents Chemother. 2015;59:536–543. doi: 10.1128/AAC.04037-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Jayol, A., Nordmann, P. & Brink, A. High level resistance to colistin mediated by various mutations in the crrB gene among carbapenemase-producing Klebsiella pneumoniae. Antimicrob. Agents Chemother61 (2017). [DOI] [PMC free article] [PubMed]
- 24.Cheng, Y.-H., Lin, T.-L., Lin, Y.-T., Wang, J.-T. Amino acid substitutions of CrrB responsible for resistance to colistin through CrrC in Klebsiella pneumoniae, 10.1128/AAC.00009-16 (2016). [DOI] [PMC free article] [PubMed]
- 25.Cheng, Y.-H., Lin, T.-L., Lin, Y.-T. & Wang, J.-T. A putative RND-type efflux pump, H239_3064, contributes to colistin resistance through CrrB in Klebsiella pneumoniae. J. Antimicrob. Chemother. dky054–dky054 (2018). [DOI] [PMC free article] [PubMed]
- 26.El-Halfawy OM, Valvano Ma. Antimicrobial Heteroresistance: an Emerging Field in Need of Clarity. Clin. Microbiol. Rev. 2015;28:191–207. doi: 10.1128/CMR.00058-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Falagas ME, Makris GC, Dimopoulos G, Matthaiou DK. Heteroresistance: A concern of increasing clinical significance? Clin. Microbiol. Infect. 2008;14:101–104. doi: 10.1111/j.1469-0691.2007.01912.x. [DOI] [PubMed] [Google Scholar]
- 28.Poudyal A, et al. In vitro pharmacodynamics of colistin against multidrug-resistant Klebsiella pneumoniae. J. Antimicrob. Chemother. 2008;62:1311–1318. doi: 10.1093/jac/dkn425. [DOI] [PubMed] [Google Scholar]
- 29.Jayol A, Nordmann P, Brink A, Poirel L. Heteroresistance to colistin in Klebsiella pneumoniae associated with alterations in the PhoPQ regulatory system. Antimicrob. Agents Chemother. 2015;59(5):2780–4. doi: 10.1128/AAC.05055-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lee, J.-Y., Choi, M.-J., Choi, H. J. & Ko, K. S. Preservation of Acquired Colistin Resistance in Gram-Negative Bacteria, 10.1128/AAC.01574-15 [DOI] [PMC free article] [PubMed]
- 31.Fookes M, Yu J, De Majumdar S, Thomson N, Schneiders T. Genome Sequence of Klebsiella pneumoniae Ecl8, a Reference Strain for Targeted Genetic Manipulation. Genome Announc. 2013;1:4–5. doi: 10.1128/genomeA.00027-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sanowar S, Martel A, Herve LM. Mutational Analysis of the Residue at Position 48 in the Salmonella enterica Serovar Typhimurium PhoQ Sensor Kinase. J. Bacteriol. 2003;185:1935–1941. doi: 10.1128/JB.185.6.1935-1941.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Gunn JS, Miller SI. PhoP-PhoQ activates transcription of pmrAB, encoding a two-component regulatory system involved in Salmonella typhimurium antimicrobial peptide resistance. J. Bacteriol. 1996;178:6857–6864. doi: 10.1128/jb.178.23.6857-6864.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Harris SR, et al. Evolution of MRSA During Hospital Transmission and Intercontinental Spread. Science (80-.) 2010;327:469–474. doi: 10.1126/science.1182395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hall BG, Acar H, Nandipati A, Barlow M. Growth Rates Made Easy. Mol. Biol. Evol. 2014;31:232–238. doi: 10.1093/molbev/mst187. [DOI] [PubMed] [Google Scholar]
- 36.Srinivasan, V. B., Singh, B. B., Priyadarshi, N., Chauhan, N. K. & Rajamohan, G. Role of novel multidrug efflux pump involved in drug resistance in Klebsiella pneumoniae. PLoS One9 (2014). [DOI] [PMC free article] [PubMed]
- 37.Hernández-Allés S, et al. Porin expression in clinical isolates of Klebsiella pneumoniae. Microbiology. 1999;145(Pt 3):673–9. doi: 10.1099/13500872-145-3-673. [DOI] [PubMed] [Google Scholar]
- 38.Subramanian A, et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA. 2005;102:15545–50. doi: 10.1073/pnas.0506580102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Deris, Z. Z. et al. ORIGINAL ARTICLE A secondary mode of action of polymyxins against Gram-negative bacteria involves the inhibition of NADH-quinone oxidoreductase activity. 67, 147–151 (2014). [DOI] [PMC free article] [PubMed]
- 40.Shively, J. M. Inclusion Bodies of Prokaryotes. Annu. Rev. Microbiol. 167–187 (1974). [DOI] [PubMed]
- 41.Moreno Switt, A. I. et al. Salmonella phages and prophages: genomics, taxonomy, and applied aspects. Salmonella Methods and Protocols1225 (2015). [DOI] [PubMed]
- 42.Kropinski AM, Sulakvelidze A, Konczy P, Poppe C. Salmonella phages and prophages–genomics and practical aspects. Methods Mol. Biol. 2007;394:133–175. doi: 10.1007/978-1-59745-512-1_9. [DOI] [PubMed] [Google Scholar]
- 43.Thomson N, et al. The role of prophage-like elements in the diversity of Salmonella enterica serovars. J. Mol. Biol. 2004;339:279–300. doi: 10.1016/j.jmb.2004.03.058. [DOI] [PubMed] [Google Scholar]
- 44.Zhou Y, Liang Y, Lynch KH, Dennis JJ, Wishart DS. PHAST: A Fast Phage Search Tool. Nucleic Acids Res. 2011;39:1–6. doi: 10.1093/nar/gkr595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Plesa, M., Hernalsteens, J., Vandenbussche, G. & Ruysschaert, J. The SlyB outer membrane lipoprotein of Burkholderia multivorans contributes to membrane integrity 157, 582–592 (2006). [DOI] [PubMed]
- 46.Baumler AJ, Hantke K. A lipoprotein of Yersinia enterocolitica facilitates ferrioxamine uptake in Escherichia coli. J. Bacteriol. 1992;174:1029–1035. doi: 10.1128/jb.174.3.1029-1035.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.López-Rojas, R., García-Quintanilla, M., Labrador-Herrera, G., Pachón, J. & McConnell, M. J. Impaired growth under iron-limiting conditions associated with the acquisition of colistin resistance in Acinetobacter baumannii. Int. J. Antimicrob. Agents, 10.1016/j.ijantimicag.2016.03.010 (2016). [DOI] [PubMed]
- 48.Kadner RJ, H. KJ. Mutual Inhibition of Cobalamin and Siderophore Uptake Systems Suggests Their Competition for TonB Function. J. Bacteriol. 1995;177:4829–4835. doi: 10.1128/jb.177.17.4829-4835.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Raetz CRH, Reynolds CM, Trent MS, Bishop RE. Lipid A modification systems in gram-negative bacteria. Annu. Rev. Biochem. 2007;76:295–329. doi: 10.1146/annurev.biochem.76.010307.145803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Holt KE, et al. Genomic analysis of diversity, population structure, virulence, and antimicrobial resistance in Klebsiella pneumoniae, an urgent threat to public health. Proc. Natl. Acad. Sci. USA. 2015;112:E3574–81. doi: 10.1073/pnas.1501049112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Beceiro A, et al. Biological cost of different mechanisms of colistin resistance and their impact on virulence in acinetobacter baumannii. Antimicrob. Agents Chemother. 2014;58:518–526. doi: 10.1128/AAC.01597-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Snitkin ES, et al. Genomic insights into the fate of colistin resistance and Acinetobacter baumannii during patient treatment. Genome Res. 2013;23:1155–1162. doi: 10.1101/gr.154328.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Falagas ME, Rafailidis PI, Matthaiou DK. Resistance to polymyxins: Mechanisms, frequency and treatment options. Drug Resist. Updat. 2010;13:132–138. doi: 10.1016/j.drup.2010.05.002. [DOI] [PubMed] [Google Scholar]
- 54.Hjort, K., Nicoloff, H. E. & Andersson, D. I. Unstable tandem gene amplification generates heteroresistance (variation in resistance within a population) to colistin in Salmonella enterica, 10.1111/mmi.13459. [DOI] [PubMed]
- 55.Band VI, et al. Antibiotic failure mediated by a resistant subpopulation in Enterobacter cloacae. Nat. Microbiol. 2016;1:16053. doi: 10.1038/nmicrobiol.2016.53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Andrews, J. M., Working, B. & Testing, S. JAC BSAC standardized disc susceptibility testing method. J. Antimicrob. Chemother. 48, Suppl. S1, 43–57 (2001). [DOI] [PubMed]
- 57.Lo-Ten-Foe JR, De Smet AMGA, Diederen BMW, Kluytmans JAJW, Van Keulen PHJ. Comparative evaluation of the VITEK 2, disk diffusion, etest, broth microdilution, and agar dilution susceptibility testing methods for colistin in clinical isolates, including heteroresistant Enterobacter cloacae and Acinetobacter baumannii strains. Antimicrob. Agents Chemother. 2007;51:3726–3730. doi: 10.1128/AAC.01406-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Dordel J, et al. Novel determinants of antibiotic resistance: Identification of mutated Loci in Highly methicillin-resistant subpopulations of methicillin-resistant Staphylococcus aureus. MBio. 2014;5:1–9. doi: 10.1128/mBio.01000-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Simpson, J. T. et al. Efficient de novo assembly of large genomes using compressed data structures sequence data. 549–556, 10.1101/gr.126953.111 (2012). [DOI] [PMC free article] [PubMed]
- 60.Holt, K. E. et al. Diversity, population structure, virulence and antimicrobial resistance of Klebsiella pneumoniae, an urgent threat to public health. PNAS (2015). [DOI] [PMC free article] [PubMed]
- 61.Zerbino DR, Birney E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008;18:821–829. doi: 10.1101/gr.074492.107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Boetzer M, Henkel CV, Jansen HJ, Butler D, Pirovano W. Scaffolding pre-assembled contigs using SSPACE. Bioinformatics. 2011;27:578–579. doi: 10.1093/bioinformatics/btq683. [DOI] [PubMed] [Google Scholar]
- 63.Boetzer M, Pirovano W. Toward almost closed genomes with GapFiller. Genome Biol. 2012;13:R56. doi: 10.1186/gb-2012-13-6-r56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Bankevich A, et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012;19:455–477. doi: 10.1089/cmb.2012.0021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Camacho C, et al. BLAST+: architecture and applications. BMC Bioinformatics. 2009;10:421. doi: 10.1186/1471-2105-10-421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Kurtz S, et al. Versatile and open software for comparing large genomes. Genome Biol. 2004;5:R12. doi: 10.1186/gb-2004-5-2-r12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Carver TJ, et al. ACT: The Artemis comparison tool. Bioinformatics. 2005;21:3422–3423. doi: 10.1093/bioinformatics/bti553. [DOI] [PubMed] [Google Scholar]
- 68.Wick RR, Schultz MB, Zobel J, Holt KE. Bandage: Interactive visualization of de novo genome assemblies. Bioinformatics. 2015;31:3350–3352. doi: 10.1093/bioinformatics/btv383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Gouy M, Guindon S, Gascuel O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010;27:221–4. doi: 10.1093/molbev/msp259. [DOI] [PubMed] [Google Scholar]
- 70.Stamatakis A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 2014;30:1312–1313. doi: 10.1093/bioinformatics/btu033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Vergara-Irigaray M, Fookes MC, Thomson NR, Tang CM. RNA-seq analysis of the influence of anaerobiosis and FNR on Shigella flexneri. BMC Genomics. 2014;15:438. doi: 10.1186/1471-2164-15-438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.De Majumdar S, et al. Elucidation of the RamA Regulon in Klebsiella pneumoniae Reveals a Role in LPS Regulation. PLOS Pathog. 2015;11:e1004627. doi: 10.1371/journal.ppat.1004627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Robinson MD, McCarthy DJ, Smyth GK. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics. 2009;26:139–140. doi: 10.1093/bioinformatics/btp616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Law CW, Chen Y, Shi W, Smyth GK. Voom: precision weights unlock linear model analysis tools for RNA-seq read counts. Genome Biol. 2014;15:R29. doi: 10.1186/gb-2014-15-2-r29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Smyth, G. K. In Bioinformatics and Computational Biology Solutions Using R and Bioconductor SE - 23 (eds Gentleman, R., Carey, V., Huber, W., Irizarry, R. & Dudoit, S.) 397–420, 10.1007/0-387-29362-0_23 (Springer New York, 2005).
- 76.Binns D, et al. The Gene Ontology - Providing a Functional Role in Proteomic. Studies. 2009;25:3045–3046. [Google Scholar]
- 77.Caspi R, et al. The MetaCyc database of metabolic pathways and enzymes and the BioCyc collection of Pathway/Genome Databases. Nucleic Acids Res. 2014;42:459–471. doi: 10.1093/nar/gkt1103. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All sequence data is available at the European Nucleotide Archive (ENA) (http://www.ebi.ac.uk/ena) under the study accession number ERP004913. Specific sample accession numbers are given in Table S2. Other data generated or analysed during this study are included in this published article and its supplementary information files.