

Abstract
Antipsychotic drugs were incidentally discovered in the 1950s, but their mechanisms of action are still not understood. Better understanding of schizophrenia pathogenesis could shed light on actions of current drugs and reveal novel druggable pathways for unmet therapeutic needs. Recent genome-wide association studies offer unprecedented opportunities to characterize disease gene networks and uncover drug-disease relationships. Polygenic overlap between schizophrenia risk genes and antipsychotic drug targets has been demonstrated. However, the specific genes and pathways constituting this overlap are undetermined. Risk genes of polygenic disorders do not operate in isolation, but in combination with other genes. Thus, we utilized protein-protein interaction networks (interactome) to map antipsychotic drug targets (n=88) to networks of schizophrenia risk genes (n=328). Our results showed that schizophrenia risk genes were significantly localized in the interactome (p=0.0015), forming a distinct disease module. Core genes of the module were enriched for genes involved in developmental biology and cognition, which may have a central role in schizophrenia etiology. Intriguingly, antipsychotic drug targets overlapped with the core disease module and comprised multiple pathways beyond dopamine. Some important risk genes like CHRN, PCDH and HCN families were not connected to existing antipsychotics, but may be suitable targets for novel drugs or drug repurposing opportunities to treat other aspects of schizophrenia such as cognitive dysfunction and negative symptoms. This network medicine approach provides a platform to collate information of disease genetics and drug-gene interactions to shift focus from development of antipsychotics to multi-target anti-schizophrenia drugs. This approach is transferable to other diseases.
Introduction
Antipsychotics were incidentally discovered over 60 years ago, but their mechanisms of actions have still not been fully revealed. Antipsychotics are the main medication available to schizophrenia patients, but have little effect on negative and cognitive symptoms of the disease. Current antipsychotics are also limited by serious side effects1,2 that reduce treatment compliance3, and about one-third of psychotic patients are classified as treatment resistant4. Thus, there is great need for improved medications for these patients, but drug development has been hampered by poor knowledge of disease etiology and underlying genetics. There is a growing interest in harnessing knowledge of risk genes for developing better treatments for common diseases5–7. For example, Nelson et al. found that drugs with genetically-supported mechanisms of action succeeded from phase 1 to approval twice as often as drugs without genetic support8. Schizophrenia is highly heritable9 and a well-powered genome wide association study (GWAS) recently identified as many as 108 independent loci related to the disease, containing over 300 genes10. Improved knowledge of these risk genes may be used to inform drug development by revealing potential mechanisms of action of current drugs and by identifying new drug targets. Antipsychotics bind to numerous proteins11, of which dopamine and serotonin receptors are the only ones with known biological links to schizophrenia. However, a recent study found genetic overlap between schizophrenia risk genes and antipsychotic target genes, which suggests the pharmacological mechanisms to be polygenic, also involving pathways that are not yet identified12.
Most genotype-phenotype relationships arise from complexity of cellular interactions13. Risk genes of polygenic disorders do not operate in isolation, but in combination and interaction with other risk genes. The effect of a perturbation in one gene can propagate to impact other nearby proteins in the protein-protein interaction (PPI) network, referred to here as the interactome. Therefore, protein products of genes that are associated with a particular disease tend to interact with each other and converge on related biological and functional networks, the so-called disease module, rather than being randomly spread throughout the interactome14,15. Thus network biology16 or network medicine15 provides an important framework where knowledge of PPI’s can be used to gain more comprehensive insight into the molecular mechanisms of complex diseases14,17.
The network approach also provides a unique opportunity to study drug effects by integrating the human interactome with knowledge of drug targets18. The Drug-Gene interaction database (DGIdb)11,19 contains information on genes whose products are known to interact with drugs in humans (drug target genes), as well as genes that belong to the ‘druggable‘ genome11,20. Okada et al.21 examined how targets of rheumatoid arthritis (RA) drugs map on to the interactome neighborhood of identified RA risk genes and found overlap between drug target genes and risk genes, as well as interactome neighbors of risk genes21. In addition, a recent study examined how drug target genes overlap with GWAS hits across a variety of diseases, and found very little direct overlap22. However, drug target genes showed threefold enrichment among the closest interactome neighbors, and enrichment was also significant among the second neighbors, suggesting that neighboring genes in the interactome should be included when searching for suitable candidates for drug repurposing22.
Here, adopting a novel network biology approach14, we study the schizophrenia disease module and its intersection with current antipsychotic drug target genes integrating information from the human interactome14 with data from DGIdb. Our objective was to inform development of new medications20 by improving understanding of disease etiology and the function of current medications, as well as by identifying genes worth further examination as new drug targets or drug repurposing opportunities. We first identified and characterized a schizophrenia disease module in the interactome, then examined the interactome link between antipsychotic drug target and schizophrenia risk genes through protein interactions. We revealed the specific risk genes and pathways that are involved in this link, and the risk genes that are not linked to current antipsychotics. This information may be useful for identifying targets of future drugs that may also treat other symptoms than psychosis.
Methods
Interactome database
The PPI network used in our main analyses is the human interactome created by Menche et al.14, consisting of 13,460 proteins interconnected by 141,296 high-confident physical interactions with experimental support (see Supplementary Information). For comparison, we also used the STRING database23 (Supplementary Information).
Network terminology and illustrations
In the network terminology used throughout this paper, nodes refer to genes/their corresponding gene products (proteins), and edges refer to connections between two genes through identified PPIs between their products. The distance (ds) between two nodes in the network is defined as the smallest number of edges, or steps, connecting them (shortest path length). Network figures were created using Cytoscape24, with nodes illustrated as a shape (octagons for drug target genes, circles for disease genes and neighbors of disease genes or drug targets) and edges as lines.
Disease risk genes
Schizophrenia risk genes were derived from the largest multi-center GWAS of schizophrenia to date10 (36,989 cases and 113,075 controls), performed by the Psychiatric Genomic Consortia (PGC). In this GWAS, 108 independent loci were identified with a p-value threshold of 5×10−8, which contained 326 potential causal protein coding genes (see Supplementary Table 3 in Ripke et al., 201410). In addition, one of the loci spans a large region on chromosome 6 containing the major histocompatibility complex (MHC). As this locus contains hundreds of genes, all could not be included in the analyses as the majority might not be functionally related to schizophrenia. A recent paper showed that the MHC- schizophrenia association is partly driven by the genes C4A and C4B25. We therefore included these two genes in the analyses to represent the signal from the MHC region, resulting in a total of 328 risk genes included in the analyses.
Network localization
To estimate network localization of the schizophrenia risk genes, we first calculated the shortest distance, ds, between each disease gene to the next closest disease gene and the corresponding frequency distribution, P(ds). To estimate if the disease genes were more localized than that expected by chance, 1000 random sets of genes with the same number of genes as the disease set were generated to yield the distribution Prand(ds) from which test statistics could be calculated for the observed value of ds (see Menche et al.14 for details). We also calculated the largest connected component of disease genes in the interactome space, containing S disease genes. S was compared with 1,000 random gene sets with the same number of genes to yield the distribution Prand(S). Following Menche et al., we define the schizophrenia disease module based on both ds and S. A statistically significant ds is driven by risk genes with the shortest distance to another risk gene, i.e. interconnected risk genes (d=1), and genes in the largest connected component, S, is a subset of interconnected risk genes. Thus, we refer to interconnected risk genes as core genes of the disease module or core disease module. Control analyses with calculation of ds that takes node degree into account for the selection of random gene sets was also performed26. Node degree was also incorporated to an alternative network localization method based on network neighborhood overlap27 (Supplementary Information).
Antipsychotic drug targets
We included 64 drugs listed as antipsychotics in the Anatomical Therapeutic Chemical (ATC) classification system of the world health organization (WHO) collaborating center for drug statistics methodology. To identify drug-gene interactions in the interactome, we used data from the drug-gene interaction database (DGIdb v2.22, http://dgidb.genome.wustl.edu, downloaded on 10/17/2016) a freely available database for identifying known and potential drug-gene interactions11,19. We identified 88 genes targeted by at least one of the 64 antipsychotics. Eighty of these were included in the Menche et al. interactome with at least one interactor.
Gene set enrichment analysis
We used MAGENTA (Meta-Analysis Gene-set Enrichment of variaNT Associations)28 to test if antipsychotic drug targets, or their first interactome neighbors, were enriched for associations with schizophrenia compared to a large number of randomly selected genes (see supplementary information for details). As control analyses, we also examined potential enrichment of antipsychotic drug target genes among GWAS identified risk genes for three control conditions, depressive symptoms29, Alzheimer’s disease30 and type 2 diabetes31.
Network graph of genetic overlap between antipsychotics targets and disease genes
We calculated the number of interaction steps (edges) between each specific schizophrenia risk gene and its closest antipsychotic drug target in the interactome, with 0, 1, 2, or >2 steps.
Gene ontology enrichment
ToppGene32 was used to examine enrichment in gene ontology (GO) annotations and pathways for the following subsets of risk gene sets related to the schizophrenia disease module: 1) all risk genes, 2) interconnected risk genes, 3) risk genes belonging to the largest connected component of risk genes (Supplementary Table 1). We also examined risk gene sets according to their shortest path length (distance) to antipsychotic drug targets in the interactome (0 or 1, 2, or >2 steps to an antipsychotic drug target, Supplementary Table 2). ToppGene uses hypergeometric distribution with Bonferroni correction for determining statistical significance. We included the GO annotation categories molecular function and biological process, as well as pathways, with a Bonferroni corrected p-value threshold of 0.05.
Results
The schizophrenia disease module
Using the Menche et al. interactome 251 schizophrenia risk genes were included. Schizophrenia risk genes were significantly localized in the interactome compared to random gene sets (p=0.0015), with an average of 1.7 edges between two risk genes, forming a disease module14. Eighty-two of 251 risk genes (33%) directly connected with at least one other risk gene (i.e., interconnected risk genes) as indicated by the outer circle in Figure 1. Three interconnected risk genes, CHRNA3, CHRNA5 and CHRNB4, were derived from the same loci in the GWAS, leaving 79 interconnected risk genes for GO enrichment analyses. The largest connected component consisted of 32 risk genes (z=1.1, p=0.27) as indicated by the inner circle in Figure 1. Using the STRING23 interactome 254 schizophrenia risk genes were included. The shortest average distance between gene pairs was 1.59 edges, again significantly shorter than for the random gene sets (p=5.2e-4). The largest connected component consisted of 57 genes (z-score=2.2, p=0.028). Control analyses taking node degree into account26 confirmed the main results of schizophrenia risk genes being significantly localized, both using the Menche et al., interactome (p=0.033) and the STRING database (p=0.006), as well as using the network neighborhood overlap method (Supplementary Information).
Figure 1. Connection between schizophrenia (SCZ) risk genes and antipsychotic drug targets.
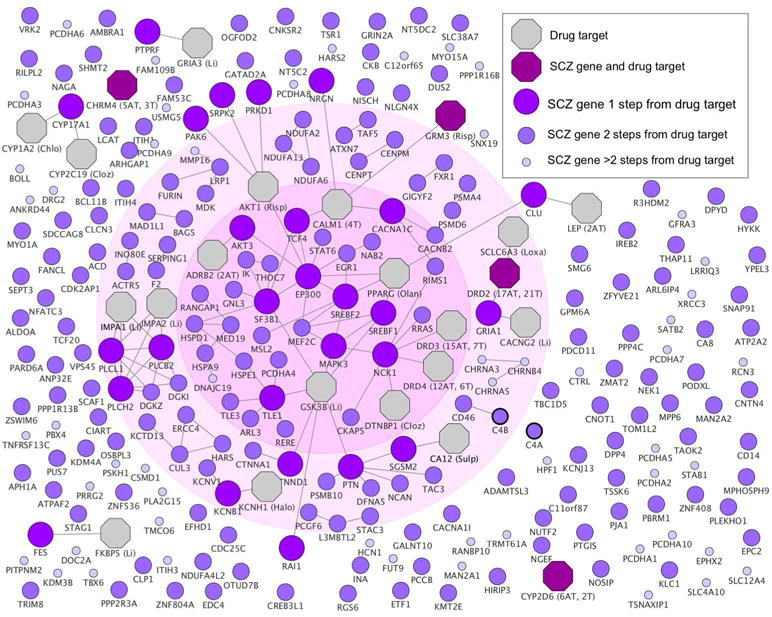
Schizophrenia risk genes are color labeled and sized according to their shortest path lengths to antipsychotic drug targets. Antipsychotic drug targets are shown as octagons, with number of atypical (AT) and typical (T) antipsychotics in parenthesis. Drug targets that do not interact with any risk gene are not shown. Abbreviated drug names are given for proteins that are targeted by a single antipsychotic: Chlo=Chlorpromazine, Cloz=Clozapine, Li=Lithium, Loxa=Loxapine, Olan=Olanzapine, Risp=Risperidone, Sulp=Sulpiride. The inner circle indicates the genes belonging to the largest connected component and the outer circle indicates the interconnected risk genes. We refer to these two gene set components as the core disease module, given that they are used as network properties to quantify the degree to which disease risk genes cluster in specific interactome neighborhoods. Risk genes close to antipsychotic drug targets were significantly overrepresented among interconnected risk genes (p=0.00023) (χ2 test) (counts shown in Supplementary Table 4). For full details of specific antipsychotics targets, see Supplementary Table 3. Further details on C4A and C4B (marked with thicker borders) are shown in Supplementary Figure 2.
To gain further insight into the biological function of risk genes we performed GO and pathway enrichment analyses for a) all schizophrenia risk genes (n=328), b) interconnected risk genes (n=79), and c) risk genes belonging to the largest connected component in the interactome (n=32). All significant results from analyses a-c are presented in full detail in Supplementary Table 1 (p <0.05, corrected). Importantly, by restricting the analyses to genes in b) and c), the core genes of the disease module only, we were able to identify enrichment for several biological processes and pathways that could not be identified when treating all risk genes equally33, including developmental biology, learning or memory, and cognition (Figure 2 and Supplementary Table 1).
Figure 2. Significant biological processes and pathways for schizophrenia disease module.
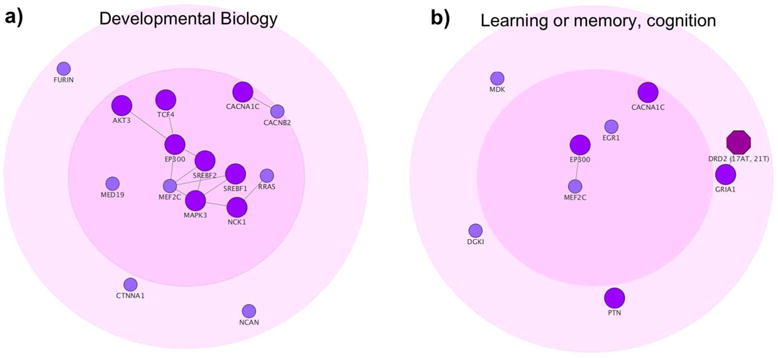
The figure shows two of the top ranked biological processes and pathways derived from Gene ontology and pathway enrichment analysis for the interconnected risk genes (n=79). a) Developmental biology pathway (477129, REACTOME, p=6.3e-9). b) Learning or memory (GO:0007611, p=1.87e-6) and Cognition (GO: 0050890, p=4.64e-6), which consisted of the same set of risk genes. See Table 1 and Supplementary Table 1.
Examining the link between antipsychotic drug targets and schizophrenia
First, using gene-set enrichment analysis (MAGENTA28), we found that antipsychotic drug target genes (n=88 from DGIdb) were significantly enriched for associations with schizophrenia (p=0.0099) compared to random gene sets, confirming a previous report of polygenic overlap between schizophrenia risk genes and antipsychotic drug targets12. Beyond replicating the previous finding, we incorporated network information as it has been shown that drug targets were substantially enriched in nearest neighbors of GWAS genes22. We identified 1,133 first neighbors of an antipsychotic drug target (proteins in direct PPI with drug targets, excluding the 88 direct drug target genes) through the Menche et al. interactome, and found that these were also significantly enriched for association with schizophrenia (p=0.0057). Twenty-seven of the 251 schizophrenia risk genes were first neighbors of drug target genes (Table 2a). No antipsychotic gene-set enrichment was seen for the control conditions depressive symptoms29, Alzheimer’s disease30 and type 2 diabetes31 (all p’s > 0.1).
To identify the specific risk genes and pathways that constitute the observed drug-disease link, we calculated the number of interactions between each risk gene and its closest antipsychotic drug target (Figure 1, Supplementary Table 2a). Four genes were both a risk gene and a drug target gene; glutamate metabotropic receptor 3 (GRM3), dopamine receptor D2 (DRD2), cholinergic receptor muscarinic 4 (CHRM4) and cytochrome P450 family 2 subfamily D member 6 (CYP2D6). Risk genes that overlap or connect with antipsychotic drug targets (≤ 1 steps from drug targets) were overrepresented among interconnected risk genes as well as genes in the largest connected component (p=9.0e-8 and p=0.00036 respectively, Supplementary Table 4). For Alzheimer’s disease, used as a control condition, interconnected risk genes were not overrepresented among risk genes ≤ 1 steps from antipsychotic drug targets (p=0.35). GO and pathway enrichment results of schizophrenia risk genes ≤ 1 steps from drug targets revealed partly overlapping results to that of interconnected risk genes, such as learning or memory, and developmental biology (Table 1). In addition, enrichment was also seen for cholinergic, glutamatergic, and dopaminergic pathways (Supplementary Table 2b). None of the schizophrenia risk genes that were not included in the interactome (n=77) were directly targeted by an antipsychotic drug, which is not extreme in comparison to all genes (p=0.89, see Supplementary Information for methods). For comparisons with the STRING database, see Supplementary Information and Supplementary Table 3.
Table 1. Biological processes and pathways related to schizophrenia risk genes overlapping with current drug targets.
The table lists the top five most significant biological processes and pathways from GO enrichment analysis of risk genes ≤1 step from an antipsychotic drug target gene (n=31). For GO terms or pathways with the same risk gene set (complete overlap or one gene set being a subset of the other), we only include the most significant here. Full details are shown in Supplementary Table 2.
| ID | Name | P value |
|---|---|---|
| Biological processes | ||
| GO:0098916 | Anterograde trans-synaptic signaling | 8.05E-08 |
| GO:0007611 | Learning or memory | 2.41E-07 |
| GO:0051130 | Positive regulation of cellular component organization | 3.54E-07 |
| GO:0010628 | Positive regulation of gene expression | 5.66E-07 |
| GO:0050769 | Positive regulation of neurogenesis | 6.56E-07 |
| Pathways | ||
| 477129 (REACTOME) | Developmental Biology | 4.40E-08 |
| 83085 (KEGG) | Long-term potentiation | 6.60E-07 |
| 217716 (KEGG) | Cholinergic synapse | 8.84E-06 |
| 213818 (KEGG) | Glutamatergic synapse | 1.01E-05 |
| 868086 (KEGG) | Rap1 signaling pathway | 1.35E-05 |
Identification of schizophrenia risk genes untargeted by antipsychotic drugs
We next sought to map risk genes that were not connected to current antipsychotics (>2 steps in Figure 1 and Supplementary Table 2, n=53) together with their first interactome neighbors (Supplementary Figure 1, n=122). The purpose of this analysis is to identify druggable genes among molecular pathways involved in other aspects of schizophrenia than those targeted by antipsychotics as potential candidates for future drug development or repurposing. These genes were enriched for calcium ion binding and acetylcholine receptor related molecular functions and pathways (Supplementary Table 2d). Three genetic clusters from this map with potential as targets for cognitive enhancers in schizophrenia are exemplified in Figure 3. Risk genes with intermediate link to drug targets (2 steps) showed few GO enrichments (Supplementary Table 2c). Finally, we examined if any existing drugs listed in the DrugBank mapped onto schizophrenia risk genes >2 steps from an antipsychotic drug target, and identified eleven approved and eight experimental drugs, as listed in the Supplementary Table 5.
Figure 3.
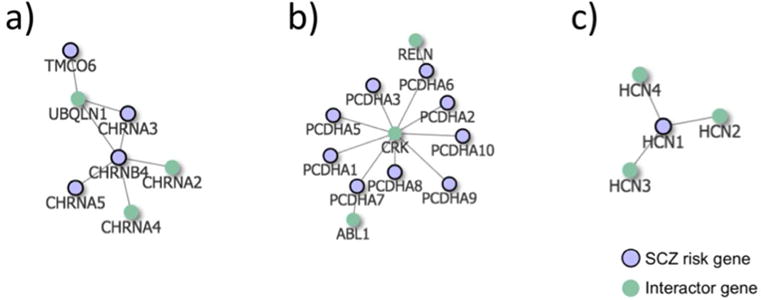
Schizophrenia risk gene networks untargeted by current antipsychotics. Highlighted schizophrenia risk genes with >2 steps/edges to an antipsychotic drug target genes, with their first interactome neighbors. a) The nicotinic acetylcholine receptor (nAChRs) genes CHRNA3, CHRNA5 and CHRNB4, b) Protocadherin alpha cluster genes, PCDHA1 - PCDHA10, and c) HCN1. We show three examples here, see Supplementary Figure 1 for the full network.
Discussion
We examined the interactome link between antipsychotics drug targets and schizophrenia risk genes identified from the latest schizophrenia GWAS10. Firstly, we identified a schizophrenia disease module, characterized by core genes involved in developmental biology and cognition. Secondly, we found antipsychotic drug target genes, as well as their first interactome neighbors, to be enriched for association with schizophrenia. Through network graphs, we found the observed interactome link between existing drugs and the disease to be located among core genes in the disease module and to involve multiple pathways. Important risk genes that were not linked to current drug targets were also identified.
Using network methods and a high-confident human interactome14, we showed that schizophrenia risk genes are significantly localized rather than randomly scattered in the interactome, forming a distinct disease module14. Previous pathway analyses performed on schizophrenia risk genes from the latest large-scale GWAS10 treated all risk genes equally and mainly identified enrichment for synapse- and dendrite-related pathways33. One problem with the previous approach is that many GWAS identified genetic loci contain several genes, of which all may not be causal to the disease. Importantly, it has been shown that when several genes are present in the same loci of a GWAS, risk genes that interact with other risk genes are more likely to be causally linked to the disease34,35. Restricting analyses to interconnected risk genes enabled the identification of novel and highly disease-relevant biological processes and pathways (Supplementary Table 1b-c). These core genes were most strongly related to developmental biology, which may constitute a central role in disease etiology (Figure 2a). Interestingly, core genes were also enriched for cognition-related processes and pathways (Figure 2b). Although schizophrenia is diagnosed based on positive and negative symptoms, cognitive impairment is a core clinical feature of the disease, with evidence of premorbid cognitive impairment from as young age as 736.
We confirmed previously reported enrichment of antipsychotic drug target genes for association with schizophrenia12 and, importantly, also found enrichment for neighbors of antipsychotic drug target genes. These results indicate that the pharmacological mechanisms of current antipsychotics overlap with the pathogenesis of schizophrenia, and that the drug-disease link extends beyond dopamine to also involve other neurotransmitter systems, such as glutamatergic and cholinergic pathways (Table 1, Supplementary Table 2b). These pathways include risk genes directly targeted by antipsychotics (the glutamatergic and cholinergic receptor genes GRM3 and CHRM4), but also risk genes interacting with drug targets, such as CACNA1C, coding for a calcium channel subunit, and cell signaling kinases such as MAP kinase 3 (MAPK3) and AKT serine/threonine kinase 3 (AKT3). Indeed, previous efforts have identified potential drug targets within the cholinergic and glutamatergic systems37, and pharmacogenetic studies have linked antipsychotic response to variants in GRM338. Genetic variations in CYP2D6, a gene linked to drug metabolism and one of the schizophrenia risk genes directly targeted by antipsychotics, have been linked to side effects of antipsychotics39. Some studies also linked variants in CYP2D6 to antipsychotics response, but results have been mixed40.” Although we did not observe significant overlap between antipsychotic targets and control conditions in our enrichment analyses, it remains possible for partial overlap at the individual gene level (e.g., pleiotropic variants) which warrants future investigation.
Although genes close to current antipsychotics were related to learning and cognition, antipsychotics show only weak improvements of cognition in randomized controlled studies41. Most antipsychotics reduce dopamine release, although differential direction of altered dopamine levels in different brain areas has been linked to the different aspects of the schizophrenia42–44. Dopamine dysfunction cannot explain all aspects of schizophrenia, and, to develop improved medication, it is important to focus also on other potential targets. We identified the risk genes that are least connected to current antipsychotics together with their first neighbors (Supplementary Figure 1). This information may be used together with other sources of knowledge, such as the druggable genome11, to inform future studies aiming to identify candidate targets for new drugs or drug repurposing to treat symptoms that are little affected by current antipsychotic medication, such as cognitive enhancing drugs. Among these genes, one important example is the nicotinic acetylcholine receptor (nAChRs) genes (Figure 3a). The nAChRs are thought to be important for the cognitive symptoms of schizophrenia45 and are also implicated in Alzheimer’s disease46,47. The nAChRs are targeted by several existing drugs, including the Alzheimer’s disease drug galantamine (Supplementary Information). Galantamine has been studied in humans with schizophrenia as well as in animal models, with indications of improved effect on cognitive symptoms and enhanced efficacy of antipsychotics in rats48,49. NAChRs are also currently being examined as potential drug targets to enhance cognition in schizophrenia patients50, further justifying our hypothesis that suitable drug targets may be found among these genes. Other interesting genes that did not overlap with antipsychotic drug genes are the Protocadherin gene cluster (PCDHA1-10), and hyperpolarization activated cyclic nucleotide gated potassium channel 1 (HCN1) (Figure 3 b and c). The PCDHA’s are neural cell adhesion proteins involved in neural differentiation during development51, processes thought to be important for the development of schizophrenia. Protocadherine genes have also been linked to cognition, personality and mood disorders52. The PCDHA genes are not considered druggable11, but two neighbor genes are (RELN and ABL1). HCN1 is druggable11 and may have a link to cognition in schizophrenia through involvement in mechanisms of synaptic plasticity and memory53. Further, these ion channels have been suggested as a potential new targets for depression54 and cognitive dysfunction in Neurofibromatosis type 155.
Limitations
A major limitation of the human interactome is potential bias towards well-studied proteins. To minimize this bias, the Menche et al. interactome also includes PPI’s derived from unbiased high-throughput data-sets56–58, and we found that schizophrenia risk gene were significantly localized also when using node degree-preserved methods. Also, the known human PPI network covers only an estimated 20% of all potential PPIs14. However, the current level of network completeness was shown to successfully identify the disease module of 226 complex diseases14. The available drug target data is also incomplete. Among the identified risk genes with more than two steps to a drug gene target, some may have undiscovered connections with drug target genes. To further address this limitation, we repeated our analyses using the less conservative STRING interactome, with consistent results.
Conclusions
By applying a network biology approach, we first identified a schizophrenia disease module, consisting of genes involving in developmental biology and cognition. Second, we found an overlap between the pathological mechanisms of schizophrenia and the pharmacological mechanisms of antipsychotics. This overlap involved protein interactions among drug targets and risk genes, which strengthens the proposal that network biology can be used to inform identification of new drug targets59. Results might be used to further advance the field of network pharmacology59 in schizophrenia, with the potential development of more efficient multi-target drugs targeting not only psychosis but also other, poorly treated aspects of schizophrenia.
Supplementary Material
Acknowledgments
This project was funded by National Institute of Mental Health R01MH100351. K.K. is supported by the Swedish Research council.
Footnotes
Previous presentation:
Winter q-bio meeting, Hawaii, 02/24/2017.
Disclosure:
The authors declare no conflict of interest.
References
- 1.Wood AJJ, Freedman R. Schizophrenia. N Engl J Med. 2003;349:1738–1749. doi: 10.1056/NEJMra035458. [DOI] [PubMed] [Google Scholar]
- 2.Tschoner A, Engl J, Laimer M, Kaser S, Rettenbacher M, Fleischhacker WW, et al. Metabolic side effects of antipsychotic medication. Int J Clin Pract. 2007;61:1356–70. doi: 10.1111/j.1742-1241.2007.01416.x. [DOI] [PubMed] [Google Scholar]
- 3.Divac N, Prostran M, Jakovcevski I, Cerovac N. Second-generation antipsychotics and extrapyramidal adverse effects. Biomed Res Int. 2014;2014:656370. doi: 10.1155/2014/656370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Meltzer HY. Treatment-Resistant Schizophrenia - The Role of Clozapine. Curr Med Res Opin. 1997;14:1–20. doi: 10.1185/03007999709113338. [DOI] [PubMed] [Google Scholar]
- 5.Sansei P, Agarwal P, Barnes M, Pastinen T, Richards JB, Cardon LR, et al. Use of genome-wide association studies for drug repositioning. Nat Biotechnol. 2012;30:317–320. doi: 10.1038/nbt.2151. [DOI] [PubMed] [Google Scholar]
- 6.Wang Z-Y, Zhang HY. Rational drug repositioning by medical genetics. Nat Publ Gr. 2013;31 doi: 10.1038/nbt.2758. [DOI] [PubMed] [Google Scholar]
- 7.Breen G, Li Q, Roth BL, Donnell PO, Didriksen M, Dolmetsch R, et al. Translating genome-wide association findings into new therapeutics for psychiatry. Nat Neurosci. 2016;19:1392–1396. doi: 10.1038/nn.4411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Nelson MR, Tipney H, Painter JL, Shen J, Nicoletti P, Shen Y, et al. The support of human genetic evidence for approved drug indications. Nat Genet. 2015;47:856–860. doi: 10.1038/ng.3314. [DOI] [PubMed] [Google Scholar]
- 9.Cardno AG, Marshall EJ, Coid B, Macdonald AM, Ribchester TR, Davies NJ, et al. Heritability Estimates for Psychotic Disorders. Arch Gen Psychiatry. 1999;56:162–168. doi: 10.1001/archpsyc.56.2.162. [DOI] [PubMed] [Google Scholar]
- 10.Ripke S, Neale BM, Corvin A, Walters JTR, Farh K-H, Holmans PA, et al. Biological insights from 108 schizophrenia-associated genetic loci. Nature. 2014;511:421–427. doi: 10.1038/nature13595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Griffith M, Griffith OL, Coffman AC, Weible JV, McMichael JF, Spies NC, et al. DGIdb: mining the druggable genome. Nat Methods. 2013;10:1209–10. doi: 10.1038/nmeth.2689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ruderfer DM, Charney AW, Readhead B, Kidd BA, Kähler AK, Kenny PJ, et al. Polygenic overlap between schizophrenia risk and antipsychotic response: A genomic medicine approach. The Lancet Psychiatry. 2016;3:350–357. doi: 10.1016/S2215-0366(15)00553-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Vidal M, Cusick ME, Barabási A-L. Interactome Networks and Human Disease. Cell. 2011;144:986–998. doi: 10.1016/j.cell.2011.02.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Menche J, Sharma A, Kitak M, Ghiassian SD, Vidal M, Loscalzo J, et al. Disease networks. Uncovering disease-disease relationships through the incomplete interactome. Science (80-) 2015;347:1257601–8. doi: 10.1126/science.1257601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Barabasi A-L, Gulbahce N, Loscalzo J. Network medicine: a network-based approach to human disease. Nat Rev Genet. 2011;12:56–68. doi: 10.1038/nrg2918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ryan CJ, Cimermančič P, Szpiech ZA, Sali A, Hernandez RD, Krogan NJ. High-resolution network biology: connecting sequence with function. Nat Rev Genet. 2013;14:865–79. doi: 10.1038/nrg3574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Parikshak NN, Gandal MJ, Geschwind DH. Systems biology and gene networks in neurodevelopmental and neurodegenerative disorders. Nat Rev Genet. 2015;16:441–458. doi: 10.1038/nrg3934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Yıldırım MA, Goh K-I, Cusick ME, Barabási A-L, Vidal M. Drug—target network. Nat Biotechnol. 2007;25:1119–1126. doi: 10.1038/nbt1338. [DOI] [PubMed] [Google Scholar]
- 19.Wagner AH, Coffman AC, Ainscough BJ, Spies NC, Skidmore ZL, Campbell KM, et al. DGIdb 2.0: mining clinically relevant drug-gene interactions. Nucleic Acids Res. 2016;44:D1036–44. doi: 10.1093/nar/gkv1165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Santos R, Ursu O, Gaulton A, Bento AP, Donadi RS, Bologa CG, et al. A comprehensive map of molecular drug targets. Nat Rev Drug Discov. 2016;16:2016. doi: 10.1038/nrd.2016.230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Okada Y, Wu D, Trynka G, Raj T, Terao C, Ikari K, et al. Genetics of rheumatoid arthritis contributes to biology and drug discovery. Nature. 2014;506:376–81. doi: 10.1038/nature12873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Cao C, Moult J, Corvin A, Craddock N, Sullivan P, Johnson A, et al. GWAS and drug targets. BMC Genomics. 2014;15:S5. doi: 10.1186/1471-2164-15-S4-S5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.von Mering C, Jensen LJ, Snel B, Hooper SD, Krupp M, Foglierini M, et al. STRING: Known and predicted protein-protein associations, integrated and transferred across organisms. Nucleic Acids Res. 2005;33:433–437. doi: 10.1093/nar/gki005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kohl M, Wiese S, Warscheid B. Cytoscape: software for visualization and analysis of biological networks. Methods Mol Biol. 2011;696:291–303. doi: 10.1007/978-1-60761-987-1_18. [DOI] [PubMed] [Google Scholar]
- 25.Sekar A, Bialas AR, de Rivera H, Davis A, Hammond TR, Kamitaki N, et al. Schizophrenia risk from complex variation of complement component 4. Nature. 2016;530:177–183. doi: 10.1038/nature16549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Guney E, Menche J, Vidal M, Barábasi A-L. Network-based in silico drug efficacy screening. Nat Commun. 2016;7:10331. doi: 10.1038/ncomms10331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Novarino G, Fenstermaker AG, Zaki MS, Hofree M, Silhavy JL, Heiberg AD, et al. Exome sequencing links corticospinal motor neuron disease to common neurodegenerative disorders. Science. 2014;343:506–11. doi: 10.1126/science.1247363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Segre AV, Groop L, Mootha VK, Daly MJ, Altshuler D. Common inherited variation in mitochondrial genes is not enriched for associations with type 2 diabetes or related glycemic traits. PLoS Genet. 2010;6:1–19. doi: 10.1371/journal.pgen.1001058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Okbay A, Baselmans BML, De Neve J-E, Turley P, Nivard MG, Fontana MA, et al. Genetic variants associated with subjective well-being, depressive symptoms, and neuroticism identified through genome-wide analyses. Nat Genet. 2016;48:624–633. doi: 10.1038/ng.3552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lambert J-C, Ibrahim-Verbaas CA, Harold D, Naj AC, Sims R, Bellenguez C, et al. Meta-analysis of 74,046 individuals identifies 11 new susceptibility loci for Alzheimer’s disease. Nat Genet. 2013;45:1452–1458. doi: 10.1038/ng.2802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Barrett JC, Clayton DG, Concannon P, Akolkar B, Cooper JD, Erlich HA, et al. Genome-wide association study and meta-analysis find that over 40 loci affect risk of type 1 diabetes. Nat Genet. 2009;41:703–7. doi: 10.1038/ng.381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Chen J, Bardes EE, Aronow BJ, Jegga AG. ToppGene Suite for gene list enrichment analysis and candidate gene prioritization. Nucleic Acids Res. 2009;37:W305–W311. doi: 10.1093/nar/gkp427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.O’Dushlaine C, Rossin L, Lee PH, Duncan L, Parikshak NN, Newhouse S, et al. Psychiatric genome-wide association study analyses implicate neuronal, immune and histone pathways. Nat Neurosci. 2015;18:199–209. doi: 10.1038/nn.3922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Franke L, van Bakel H, Fokkens L, de Jong ED, Egmont-Petersen M, Wijmenga C. Reconstruction of a functional human gene network, with an application for prioritizing positional candidate genes. Am J Hum Genet. 2006;78:1011–25. doi: 10.1086/504300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Walhout AJM, Vidal M, Dekker J. Hanbook of systems biology. Elsevier; 2013. Interactome Networks; pp. 45–63. [Google Scholar]
- 36.Meier MH, Caspi A, Reichenberg A, Keefe RSE, Fisher HL, Harrington H, et al. Neuropsychological Decline in Schizophrenia From the Premorbid to the Postonset Period: Evidence From a Population-Representative Longitudinal Study. Am J Psychiatry. 2014;171:91–101. doi: 10.1176/appi.ajp.2013.12111438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Marder SR. The NIMH-MATRICS project for developing cognition-enhancing agents for schizophrenia. Dialogues Clin Neurosci. 2006;8:109–13. doi: 10.31887/DCNS.2006.8.1/smarder. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bishop JR, Ellingrod VL, Moline J, Miller D. Association between the polymorphic GRM3 gene and negative symptom improvement during olanzapine treatment. Schizophr Res. 2005;77:253–260. doi: 10.1016/j.schres.2005.04.001. [DOI] [PubMed] [Google Scholar]
- 39.Nussbaum LA, Dumitraşcu V, Tudor A, Grădinaru R, Andreescu N, Puiu M. Molecular study of weight gain related to atypical antipsychotics: clinical implications of the CYP2D6 genotype. Rom J Morphol Embryol. 2014;55:877–84. [PubMed] [Google Scholar]
- 40.Puangpetch A, Vanwong N, Nuntamool N, Hongkaew Y, Chamnanphon M, Sukasem C. CYP2D6 polymorphisms and their influence on risperidone treatment. Pharmgenomics Pers Med. 2016;9:131–147. doi: 10.2147/PGPM.S107772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Hill SK, Bishop JR, Palumbo D, Sweeney JA. Effect of second-generation antipsychotics on cognition: current issues and future challenges. Expert Rev Neurother. 2010;10:43–57. doi: 10.1586/ern.09.143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Gopalakrishna G, Ithman MH, Lauriello J. Update on New and Emerging Treatments for Schizophrenia. Psychiatr Clin North Am. 2016;39:217–238. doi: 10.1016/j.psc.2016.01.005. [DOI] [PubMed] [Google Scholar]
- 43.Takahashi H, Kato M, Takano H, Arakawa R, Okumura M, Otsuka T, et al. Differential contributions of prefrontal and hippocampal dopamine D(1) and D(2) receptors in human cognitive functions. J Neurosci. 2008;28:12032–12038. doi: 10.1523/JNEUROSCI.3446-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Condray R, Yao JK. Cognition, dopamine and bioactive lipids in schizophrenia. Front Biosci (Schol Ed) 2011;3:298–330. doi: 10.2741/s153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Friedman JI. Cholinergic targets for cognitive enhancement in schizophrenia: focus on cholinesterase inhibitors and muscarinic agonists. Psychopharmacology (Berl) 2004;174:45–53. doi: 10.1007/s00213-004-1794-x. [DOI] [PubMed] [Google Scholar]
- 46.Lombardo S. Role of the nicotinic acetylcholine receptor in Alzheimer’s disease pathology and treatment. Neuropharmacology. 2015;96:255–262. doi: 10.1016/j.neuropharm.2014.11.018. [DOI] [PubMed] [Google Scholar]
- 47.Burghaus L, Schütz U, Krempel U, de Vos RA, Jansen Steur EN, Wevers A, et al. Quantitative assessment of nicotinic acetylcholine receptor proteins in the cerebral cortex of Alzheimer patients. Mol Brain Res. 2000;76:385–388. doi: 10.1016/s0169-328x(00)00031-0. [DOI] [PubMed] [Google Scholar]
- 48.Wadenberg M-LG, Manetti D, Romanelli MN, Arias HR. Significance of the nicotinic alpha7 receptor in cognition and antipsychotic-like behavior in the rat. Behav Brain Res. 2017;333:129–134. doi: 10.1016/j.bbr.2017.07.001. [DOI] [PubMed] [Google Scholar]
- 49.Koola MM, Buchanan RW, Pillai A, Aitchison KJ, Weinberger DR, Aaronson ST, et al. Potential role of the combination of galantamine and memantine to improve cognition in schizophrenia. Schizophr Res. 2014;157:84–89. doi: 10.1016/j.schres.2014.04.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.McLean SL, Grayson B, Marsh S, Zarroug SHO, Harte MK, Neill JC. Nicotinic α7 and α4β2 agonists enhance the formation and retrieval of recognition memory: Potential mechanisms for cognitive performance enhancement in neurological and psychiatric disorders. Behav Brain Res. 2016;302:73–80. doi: 10.1016/j.bbr.2015.08.037. [DOI] [PubMed] [Google Scholar]
- 51.Yagi T, Takeichi M. Cadherin superfamily genes: functions, genomic organization, and neurologic diversity. Genes Dev. 2000;14:1169–1180. [PubMed] [Google Scholar]
- 52.Chang H, Hoshina N, Zhang C, Ma Y, Cao H, Wang Y, et al. The protocadherin 17 gene affects cognition, personality, amygdala structure and function, synapse development and risk of major mood disorders. Mol Psychiatry. 2017:1–13. doi: 10.1038/mp.2016.231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Herrmann S, Stieber J, Ludwig A. Pathophysiology of HCN channels. Pflügers Arch - Eur J Physiol. 2007;454:517–522. doi: 10.1007/s00424-007-0224-4. [DOI] [PubMed] [Google Scholar]
- 54.Shah MM. HCN1 channels: a new therapeutic target for depressive disorders? Sci Signal. 2012;5:1–3. doi: 10.1126/scisignal.2003593. [DOI] [PubMed] [Google Scholar]
- 55.Omrani A, van der Vaart T, Mientjes E, van Woerden GM, Hojjati MR, Li KW, et al. HCN channels are a novel therapeutic target for cognitive dysfunction in Neurofibromatosis type 1. Mol Psychiatry. 2015;20:1311–1321. doi: 10.1038/mp.2015.48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Rolland T, Taşan M, Charloteaux B, Pevzner SJ, Zhong Q, Sahni N, et al. A proteome-scale map of the human interactome network. Cell. 2014;159:1212–26. doi: 10.1016/j.cell.2014.10.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Venkatesan K, Rual J-F, Vazquez A, Stelzl U, Lemmens I, Hirozane-Kishikawa T, et al. An empirical framework for binary interactome mapping. Nat Methods. 2009;6:83–90. doi: 10.1038/nmeth.1280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Yu H, Tardivo L, Tam S, Weiner E, Gebreab F, Fan C, et al. Next-generation sequencing to generate interactome datasets. Nat Methods. 2011;8:478–480. doi: 10.1038/nmeth.1597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Hopkins AL. Network pharmacology: the next paradigm in drug discovery. Nat Chem Biol. 2008;4:682–90. doi: 10.1038/nchembio.118. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.