
Fig. 1.
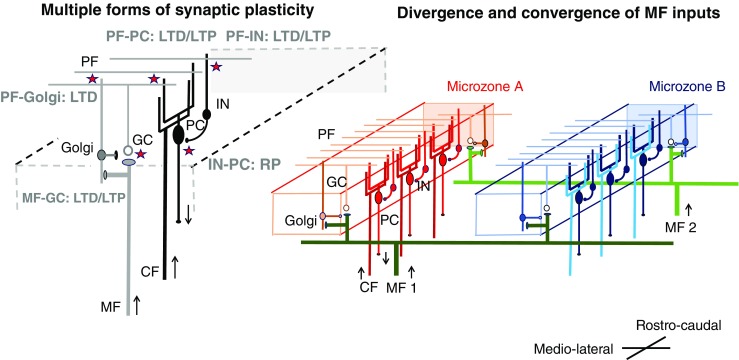
A schematic diagram of circuits in the cerebellar cortex. PC, Purkinje cell; GC, granule cell; IN, molecular layer interneurons; Golgi, Golgi cell; MF, mossy fiber; PF, parallel fiber; CF, climbing fiber. Two cellular mechanisms underlying cerebellar reserve are illustrated; (1) Multiple forms of synaptic plasticity and (2) convergence and divergence of mossy fiber inputs. Multiple forms of synaptic plasticity. Multiple forms of synaptic plasticity (illustrated by stars) co-exist in the cerebellar cortex [6, 7]. For example, conjunctive inputs from PF and CF on PC lead to depression of parallel fiber synapses [8]. LTD, long-term depression; LTP, long-term potentiation; RP, rebound potentiation. Excitatory neurons are shown with a white soma, whereas inhibitory neurons are shown with a black soma. Convergence of inputs from different MF in one microzone and divergence of inputs from one MF to several microzones. A group of CFs from a restricted area of the inferior olive nucleus innervates PCs which are located in a rostro-caudal sagittal area, forming a functional unit (microzone) [9]. MFs extend widely with a medio-lateral direction, terminating on GCs in different microzones [10]. For example, MF1 innervates both microzones A and B. Different MFs converge simultaneously to multiple microzones. For example, microzone A receives inputs from both MF1 and MF2. Thus, one microzone receives abundant central and peripheral inputs through MFs, which results in a redundancy of information. Most of them might be unutilized by a mechanism of silent synapse [11]. Possible mechanisms underlying cerebellar reserve. Damage to a single microzone can be compensated by other microzones reconstructing the lost internal model using synaptic plasticity and redundant central and peripheral information