

Abstract
Several bacterial etiological agents of streptococcal disease have been associated with fish mortality and serious global economic loss. Bacterial identification based on biochemical, molecular, and phenotypic methods has been routinely used, along with assessment of morphological analyses. Among these, the molecular method of 16S rRNA sequencing is reliable, but presently, advanced genomics are preferred over other traditional identification methodologies. This review highlights the geographical variation in strains, their relatedness, as well as the complexity of diagnosis, pathogenesis, and various control methods of streptococcal infections. Several limitations, from diagnosis to control, have been reported, which make prevention and containment of streptococcal disease difficult. In this review, we discuss the challenges in diagnosis, pathogenesis, and control methods and suggest appropriate molecular (comparative genomics), cellular, and environmental solutions from among the best available possibilities.
Keywords: antimicrobial, aquaculture, geography, sequencing, Streptococcus
INTRODUCTION
Aquaculture is among the fastest growing businesses in the food production industry (Bondad-Reantaso et al., 2005), and streptococcal infections have caused significant economic losses in the aquaculture industry (Austin and Austin, 2007; Toranzo et al., 2005).
Various bacterial agents cause streptococcosis; Streptococcus parauberis, Streptococcus iniae, Streptococcus agalactiae, and Streptococcus dysgalactiae are the prominent species regardless of geographical region (Agnew and Barnes, 2007; Nho et al., 2009; 2013; Toranzo et al., 2005; Vendrell et al., 2006). Fish are an important food source, but have always been at risk of acquiring Streptococcus infections owing to continuous exposure and ubiquitous global presence of various bacterial strains and species. Currently, there are several methods for detecting and identifying Streptococcus spp., depending upon the phenotypic characteristics, species, and strains of bacteria.
Control of streptococcus infection mainly relies on the use of antimicrobial compounds, vaccinations, and environmental strategies (Cheng et al., 2010; Darwish and Hobbs, 2005; Hastein et al., 2005; Sommerset et al., 2005; Woo and Park, 2014), of which vaccines and antimicrobial compounds have been ineffective for various reasons (Agnew and Barnes, 2007; Park et al., 2009; Shoemaker et al., 2001; Toranzo et al., 2005). Environmental strategies have been used to control fish infections in their natural and artificial habitats by several methods (Holmer, 2010). In this review, we discuss the current status and challenges in diagnosis, pathogenesis, and control of streptococcal disease in fish and we suggest effective control strategies.
WORLDWIDE DISTRIBUTION AND RISK FACTORS FOR STREPTOCOCCAL DISEASE
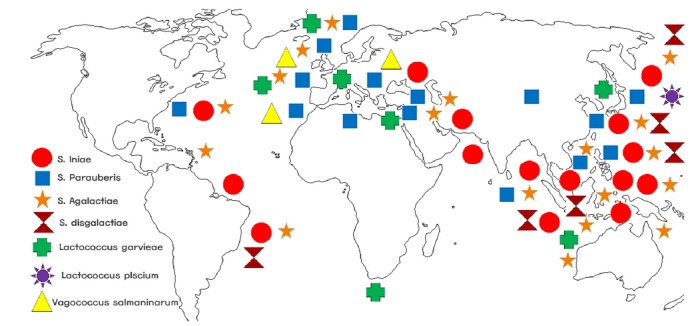
Streptococcal disease occurs in all continents (Americas, Asia, Europe, Africa, and Australia) (Table 1 and Fig. 1). Thousands of Streptococcus species (S. parauberis, S. iniae, S. agalactiae, Lactococcus garvieae, S. dysgalactiae, and Vagococcus salmoninarum) have been reported in different parts of the world (Abdelsalam et al., 2013; Agnew and Barnes, 2007; Baeck et al., 2006; Carson et al., 1993; Chang and Plumb, 1996; Diler et al., 2002; Li et al., 2015; Nho et al., 2013; Pereira et al., 2010; Ruiz-Zarzuela et al., 2005) (Fig. 1). Streptococcosis is a multifactorial disease in fish, depending on host variety, age, immune status, type of pathogen (species and strain), and environmental conditions (Ghittino et al., 1999; Ravelo et al., 2001; Vendrell et al., 2006).
Table 1.
Streptococcus bacterial agents and detailed information of affected fish species, locations, hosts, and clinical criteria
| Species | Host | Fish species | Clinical Criteria | Geographical Location |
|---|---|---|---|---|
| Streptococcus iniae | Fish, Human | Hybrid striped bass, Nile tilapia, Hybrid tilapia, Rainbow trout, Red drum, Rabbitfish, Sea bass, Olive flounder, Barramundi, Wild fish | Hemorrhage, exophthalmia, abdominal distension, ascites, lesions (liver, kidney, spleen, and intestine) | Canada, Americas, Bahrain, Israel, Thailand, China, Japan, Singapore, Taiwan, Korea |
| Streptococcus parauberis | Fish, Cow | Olive flounder, Rainbow trout, Cultured turbot, Hybrid striped bass | Chronic wasting syndrome, hemorrhagic septicemia, exophthalmia, meningitis with abnormal swimming | Israel, Italy, Japan, Spain, USA, China, Iran, Korea, Malaysia, India |
| Streptococcus agalactiae | Fish, Cow, Human, Chickens, Camels, Dogs, Horses, Cats, Frogs, Hamsters, Monkeys | Nile tilapia, Barcoo grunter, Golden pompano, Giant Queensland grouper, Ya-fish, Silver pomfret | Erratic swimming, appetite, lethargy, uncoordinated movements, exophthalmia (uni- or bi-lateral), intraocular hemorrhage, opaqueness of cornea, ascites | Europe, Turkey, China, Indonesia, Malaysia, Japan, Korea, Vietnam, Philippines, Americas |
| Lactococcus garvieae | Fish, Cow, Human, Cat, Dog, Water buffalo | Rainbow trout, Yellowtail, Tilapia, Japanese eel, Grey mullet, Black rockfish, Catfish, Wild wrasse, Giant fresh water prawn, Olive flounder, Amberjack, kingfish | Melanosis, lethargy, erratic swimming, disorientation, fins, exophthalmia (uni- or bi-lateral), swollen abdomens, anal prolapses, hemorrhages (periorbital, perianal, buccal regions) | Turkey, Australia, South Africa, England, Portugal France, Balkans, Israel, Korea |
| Streptococcus dysgalactiae | Fish, Calves, Lamb, Human, Sheep, Dogs, Pig, Lamb, Cats | White spotted snapper, Kingfish, Grey mullet, Cobia, Hybrid red tilapia, Pompano, Basket mullet, Pompano, Golden pomfret, Amur sturgeon, Nile tilapia, Yellow tail, Amber-jack | Abnormal swimming, loss of orientation, exophthalmia | Brazil, Indonesia, Malaysia, Taiwan, China, Japan |
| Vagococcus salmoninarum | Fish | Rainbow trout, Atlantic salmon, Brown trout | Loss of equilibrium, exophthalmia, melanosis, bleeding (jaw, eye, mouth, abdomen, fins, and anus), necropsy, transparent fluid accumulation, fibrinous deposits (heart, liver, spleen) | France, Italy, Spain |
Fig. 1. Phylogeography of major fish pathogens belonging to Streptococcus species, S. iniae (red circle), S. parauberis (blue square), S. dysgalactiae (brown inverted box), S. agalactiae (orange star), Lactococcus garvieae (green plus sign), Vagococcus salmoninarum (yellow triangle), and Lactococcus piscium (violet rising sun). Distribution pattern shows the presence of these bacterial isolates over the continents.
CURRENT CHALLENGES IN STREPTOCOCCOSIS DIAGNOSIS
Currently, the immediate and inexpensive diagnosis of streptococcosis in infected fish is difficult as fish exhibit similar clinical symptoms regardless of the etiological agent (Table 1 and Fig. 1) (Baeck et al., 2006; Muzquiz et al., 1999). Various issues of diagnosis are discussed below.
Clinical phenotype is the primary signature of bacterial infections that depends on various factors, so it is difficult to understand the precise cause of infection. A study of tilapia fish showed that clinical phenotypes and degree of lesions depend on several factors such as S. agalactiae strain variations, their infectious dose, water conditions, temperature, and handling procedures (Chang and Plumb, 1996). Due to the complexity of bacteria and their interrelations, microbiologists occasionally misidentify or cannot identify isolates (Lau et al., 2006). Asymptomatic fish serve as a pathogen reservoir and pose challenges for correct identification due to the absence of clinical signs (Bromage et al., 1999). Presently, diagnosis of subclinical infections in fish is a major concern.
Bacterial identification methods based on culture, morphology, or biochemical reactions are time- and resource-consuming. Some pathogen databases (RAPID Strep strip, VITEX systems, API 20E STREP, Rapid Strep 32 and ATB Expression System) are incomplete or incorrect, and result in improper identification of bacteria (Dodson et al., 1999; Facklam et al., 2005; Lau et al., 2006). Additionally, other challenges for accurate identification include the mixed nature of the aquaculture environment, low numbers of biological samples, or unknown tissue location in carriers (Klesius et al., 2006). Identification of S. agalactiae based on biochemical features (i.e. capacity to hydrolyze hippurate) or phenotypic characteristics (acidification of tagatose, ribose, and sucrose) are not effective due to high levels of biochemical heterogeneity among strains (Ravelo et al., 2001).
Molecular methods are based on several candidate genes (Table 2), that have been well characterized for diversity, including 16S rRNA, heat-shock genes (groESL), and tRNA gene intergenic spacer regions (ITSs) (Clarridge et al., 2001; Teng et al., 2002). Comparative studies have discussed various methodologies and found molecular methods to be most effective for bacterial identification (Bosshard et al., 2006). S. dysgalactiae (GCSD) was recently identified as a fish-specific pathogen based on 16S rRNA, sodA, and tuf gene sequence analysis (Abdelsalam et al., 2013). Phenotypic criteria failed to differentiate between genealogically distinct L. garvieae and Lactococcus lactis strains, so correct identification was determined by molecular methods (Klijn et al., 1991; Vendrell et al., 2006). However, some limitations are associated with diversity experiments. For example, closely related species could not be distinguished by 16S rRNA in a study by the Mitis group at the NHS (Nielsen et al., 2009). Moreover, the taxonomic position of several aquatic isolates is still unclear (Austin and Austin, 2007).
Table 2.
Candidate genes used for differentiation and diagnosis of various Streptococcus bacterial agents
| Candidate gene | References |
|---|---|
| Manganese-dependent superoxide dismutase gene (sodA) | Kitten et al., 2012; Poyart et al., 2000 |
| Heat shock protein (groESL) | Hung et al., 2013; Teng et al., 2014 |
| Ribosomal protein (rpoB) | Drancourt et al., 2013 |
| Recombination and repair protein (recN) | Hung et al., 2013 |
| Repair protein recN | Glazunova et al., 2013 |
| Lactate oxidase gene (lctO) | Zlotkin et al., 1998 |
| rRNA | Clarridge et al., 2002 |
| RNA polymerase | Drancourt et al., 2004 |
| D-alanine-D-alanine ligase | Garnier et al., 1997 |
| b-subunit of the elongation factor | Picard et al., 2004 |
| Polysaccharide capsules gene (cps) | Lowe et al., 2007 |
| Invasion associated gene (iag) | Rajagopal, 2009 |
| Surface immunogenic protein (sip) | Springman et al., 2009 |
| C5a peptidase (scp) | Springman et al., 2009 |
| Serine protease (csp) | Springman et al., 2009 |
| tRNA gene intergenic spacer region (ITS) | Tung et al., 2007 |
Another molecular method, multilocus sequence typing (MLST; analyses of multiple genetic loci or housekeeping genes) is considered the “gold standard” of typing for many bacterial species (Maiden, 2006; Jolly et al., 2012). However, insufficient resolution among very closely related bacteria can be a problem (Achtman, 2008).
CURRENT CHALLENGES IN ELUCIDATING STREPTOCOCCOSIS PATHOGENESIS
The pathogenesis of streptococcosis depends upon several factors that vary with fish species and bacterial species and isolates. Further details of virulence and pathogenicity of streptococcosis are given below.
Genetic virulence depends on several factors; for example, S. iniae virulence is associated with a unique genetic profile (Fuller et al., 2001). Comparison of 17 geographically different strains of L. garvieae based on genetic homogeneity vs. serological data showed that pathogen diversity is related to virulence factors (Barnes and Ellis, 2004). Another study showed that the lactococcal bacterial population presented a clonal structure in endemic regions, while in sporadic regions, it displayed a high genetic heterogeneity (Eyngor et al., 2004). Virulence experiments have shown that capsulated L. garvieae strains are more virulent than non-capsulated strains in rainbow trout (Barnes et al., 2002). Virulence varies with bacterial isolates within the same species in S. dysgalactiae (Abdelsalam et al., 2010). Currently, the potential for Streptococcus species to cross interspecies barriers and cause disease in other hosts is poorly understood.
Many streptococcal species are multi-host pathogens. Humans constantly face the risk of infection due to close interactions with the fish industry (Abdelsalam et al., 2010). S. iniae, S. agalactiae, L. garvieae, and S. dysgalactiae are human pathogens, and thus are a major threat to public health. S. iniae can cause bacteremic cellulitis, septic arthritis, meningitis, and endocarditis (Agnew and Barnes, 2007; Al-Harbi, 2011; Facklam et al., 2005; Lau et al., 2006; Weinstein et al., 1997), while S. agalactiae can cause meningitis and pneumonia in humans (Brimil et al., 2006; Johri et al., 2006).
CURRENT CHALLENGES FOR CONTROLLING STREPTOCOCCOSIS
Disease control using antimicrobials, vaccination, and environmental strategies are used extensively; however, some are associated with various downstream challenges that are mentioned in Fig. 2 and discussed in detail below.
Fig. 2. Schematic diagrams of diagnosis, control, and present challenges of Streptococcus disease. Bottom of the diagram shows the effect of these challenges.
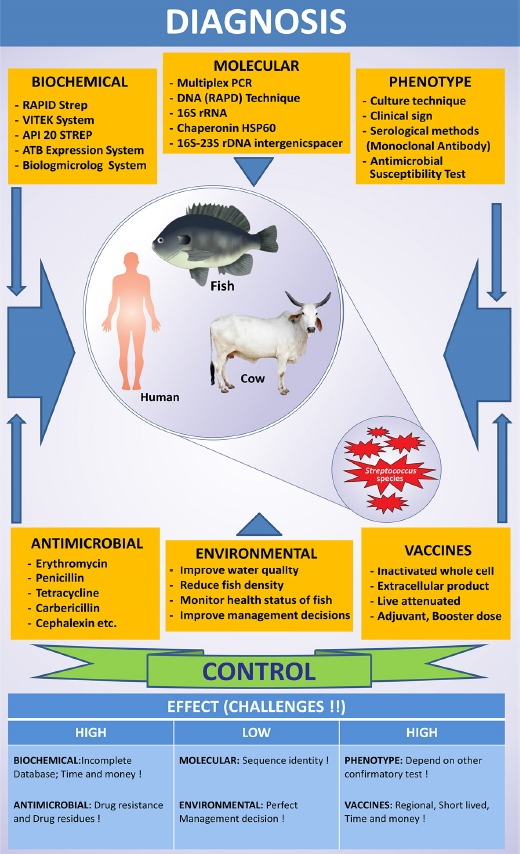
Streptococcal diseases in fish initially affect the skin, fins, gills, and external organs. Thus, controlling infections externally through liquid disinfecting agents that can dissolve in water very easily (copper sulfate and formalin) are a good option, however, these agents cause hazardous environmental side effects.
Antibiotic resistance genes are frequently transferred among bacterial species, leading to resistant Streptococcus isolates and imposing a lasting risk to public health (Park et al., 2009). The selection for resistance among dense populations and drug residues in farmed fish are major concerns associated with human health; moreover, elimination of therapeutic compounds depends on several factors that can affect the fish metabolic rates such as dose rate, route of administration, water temperature, and type of isolate (Agnew and Barnes, 2007). V. salmoninarum is resistant to most antibiotics registered for aquaculture use in the European Union (Ruiz-Zarzuela et al., 2005). Studies show that the long-term use of antibiotics in olive flounder fish may have generated a higher degree of antibiotic resistance in S. parauberis than in S. iniae (Park et al., 2009). Antibiotic sensitivity and resistance vary by geographical region in L. garvieae (Diler et al., 2002; Ravelo et al., 2001; Vendrell et al., 2006).
The capacity of bacteria to bypass any phagocyte activity and oxidative killing of host cells is a very important step utilized for vaccination strategies (Buchanan et al., 2008). Pathogenicity of streptococcal bacteria depends on their capability to survive in host immune cells. The bacteria induce internal apoptosis while avoiding killing host cells to establish infections (Woo and Park, 2014; Zlotkin et al., 2003). The pathogenesis of S. iniae infection is still not fully understood as it depends upon various virulence factors and multistep processes (Zlotkin et al., 2003). Many wide spread vaccination programs have been unsuccessful for various reasons such as regional or short-lived effects due to limited knowledge of immunity and virulence factors in fish (Agnew and Barnes, 2007). One of the recent challenges in streptococcal disease is the histological infection of various organs (liver, spleen, kidney, brain, etc.). Thus, vaccination methods will not be effective unless they provoke a systemic immune response (Li et al., 2015). Additionally, complete knowledge of molecular and cellular processes involved in disease progression is crucial to understand various downstream complications. Vaccine preparation and optimizations solely depend on an understanding of many biological processes such as bacterial morphology, biochemical assays (serotype), pathogenicity (virulence), and immunogenicity. However, understanding molecular and cellular processes for long-term protection and cost effectiveness will be crucial to promote the widespread use of vaccines.
The environment can modulate the innate immune system in fish, so any intensive culture systems immediately make fish susceptible to infection and provide a further source for the spread of infection (Magnadottir, 2006). Studies show that increased fish density and other stress factors can elicit harmful effects in fish (Eldar et al., 1995; Shoemaker et al., 2000; 2001). A system for improving water quality and monitoring fish health is another parameter that can decrease the chances of infection, as deteriorating water quality promotes the rapid spread of bacteria and mortality (Eldar et al., 1995). Various additional strategies, such as reducing fish density by using effective physical barriers (netting), and removing moribund fish are also considered to be effective steps (Shoemaker et al., 2000; 2001).
EFFECTIVE CONTROL OF STREPTOCOCCUS
Effective control systems can be established through coordination and complete knowledge of the fishery industry, fish molecular and cellular biology, ecological conditions, bacterial molecular and cellular biology, and appropriate management. However, based on various studies, it is also clear that due to the unavailability of any effective and universal vaccines or antibiotics for fish diseases, environmental protective measures may be the best strategies for controlling streptococcal disease (Fig. 3).
Fig. 3. Schematic diagram of effective control strategy of Streptococcus infection in fish.
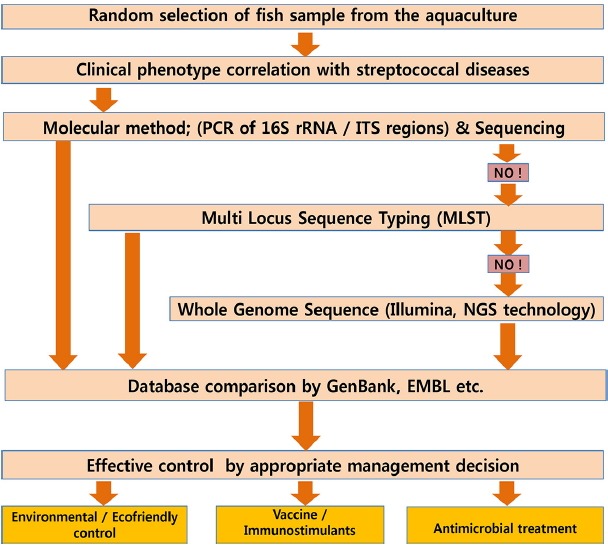
Environmental control is most ideal as it is inexpensive, easily monitored, and is not associated with any side effects (Figs. 2 and 3). Here, we briefly discuss the important parameters that should be addressed for effective environmental control.
Studies have suggested that virulence factors play an important role in pathogenesis and disease (Barnett et al., 2015; Rajagopal, 2009). Thus, basic and advanced research at the cellular level can provide more knowledge of biological processes involved in fish resistance against streptococcal diseases. These studies should focus on understanding innate and adaptive responses through cellular (macrophages, T cell and B cell markers) and humoral (various immunoglobulin classes, complement factors, cytokines) pathways. Studies have shown that several factors are involved in the regulation of Group B streptococci (GBS) disease pathogenesis, including pore-forming toxins and several adherence and immune evasion factors, which are reviewed in detail in Table 1 of Rajagopal, 2009. Additionally, signal transduction systems (STSs) are also important drug targets for effective disease control (Barrett et al., 1998; Rajagopal, 2009). Several S. agalactiae factors, such as polysaccharide capsule, hemolysin, superoxide dismutase, and D-alanylated lipoteichoic acid, hold major importance for virulence (Lindahl et al., 2005). Various surface proteins of S. agalactiae participate in important functions during disease progression, thus they are also considered important vaccine candidates (Carey-Ann et al., 2003; Lindahl et al., 2005).
Many immunostimulants can stimulate pathogen-associated molecular patterns (PAMPS), which are part of fish immune systems as pathogen recognition receptors (PRRs) and participate in maintaining innate immune protection for fish (Chettri et al., 2011).
Recently, dietary intake of bacterial components, polysaccharides, animal-derived nutrients, plant extracts, nutritional factors, and cytokines has been reported to be an effective method for immunostimulation in fish (Sakai, 1999; Villegas et al., 2006). Further advanced cellular studies of immunostimulants are required for understanding various downstream cellular activities such as hemotaxis, respiratory burst, phagocytosis, and lysozymes to establish the most effective stimulants for fish. Additionally, the development of a range of assays such as immunohistochemistry, immunocytochemistry, flow-cytometry, and gene expression technology combined with in vivo challenge studies can improve our understanding in a more advanced way and help design effective vaccines or therapeutic agents for fish diseases.
Knowledge of genetic virulence factors is important for understanding disease mechanisms and progression. Unfortunately, studies have been limited to rat and mouse models for various streptococci so far. The molecular basis of virulence was described in detail by Bennedsen et al. in 2011 (Bennedsen et al., 2011; Springman, 2009). Virulence factors vary with the streptococci strain and host since host/external environments are also responsible for variable expression of virulent genes.
Some important genetic virulence factors like polysaccharide capsules gene (cps), invasion associated gene (iag), surface immunogenic protein (sip), C5a peptidase (scp), serine protease (csp), and many surface proteins have been studied extensively in various streptococcus species (Table 2) (Lowe, 2007; Rajagopal, 2009; Springman, 2009). Additional factors regulating expression of virulence genes, like STSs, membrane associated sensor histidine kinase (HK), and toxins can also be targeted for disease pathogenesis (Lowe, 2007; Rajagopal, 2009; Springman, 2009). Genomic comparison of virulent and non-virulent S. agalactiae showed differences among isolates that infect fish, humans, cattle, and sea mammals; moreover, several host (fish) genes translate proteins that are responsible for adaptation in the aquatic environment (Delannoy et al., 2016). Isolates from one host are not necessarily pathogenic to other hosts or to different strains of the same fish (Nishiki et al., 2010).
The recent introduction of ‘next-generation’ sequencing technology has brought a revolution in bacterial research as many bacterial genomes and antimicrobial resistance and virulence genes are available for analysis (Chain et al., 2009; Kwong et al., 2015; Medini et al., 2008). Currently, the use of whole-genome sequencing (WGS) for typing any bacterial agent is possible in a cost effective and timely manner (Kwong et al., 2015). The Genus Streptococcus comprises diverse bacterial species that emerged through the course of evolution with various known and unknown evolutionarily important factors distributed worldwide. These evolutionary genetic, environmental, biochemical, and physiological factors shape streptococcus at the phenotype and genotype levels. Among them, genetic factors such as genomic variation through addition, reduction, and gene transfer play a crucial role for appropriately measuring exponential growth of diversified streptococcus genomes. Therefore, a future study should examine geographically distributed strains as they develop specific variations under different environmental conditions, which can affect their functional properties, and ultimately their virulence.
Choosing conserved genes (e.g., 16S rRNA, ITS) over other genes relies on the fact that any variation observed can be directly correlated to the physiology and virulence properties of an organism (Nho et al., 2013) and for understanding evolutionary relationships and disease epidemiology (Dobrindt and Hacker, 2001; Sakala et al., 2002; Wren, 2000). Additionally, a well-defined 16S rRNA and ITS database for sequence comparisons suggested that molecular techniques are appropriate for both taxonomic and identification purposes (Mishra et al., 2017; Mora et al., 2003; Yoon et al., 2017). Advanced genomics refers to high-throughput genetic technologies (based on DNA or RNA nucleotides) and their evaluation through comparative, functional, or environmental parameters (Gao et al., 2014). Comparative genomics through chemotaxonomic, DNA-DNA hybridization, and 16S rRNA sequencing approaches should be further evaluated occasionally through advanced computational technologies for establishing the evolutionary relationship among strains.
In this review, based on the NCBI database, we selected and analyzed the 16S rRNA gene in 51 representative worldwide strains of streptococcus species (S. parauberis, S. iniae, S. agalactiae, L. garvieae, S. dysgalactiae, and V. salmoninarum) (Table 3 and Supplementary Table 1). Our phylogenetic analysis demonstrates the interrelation of various strains and suggests that the 16S rRNA gene can easily segregate various strains based on genetic variations (Fig. 4 and Supplementary Fig. 1). The evolutionary history was inferred using the Neighbor-Joining method and MEGA7. Earlier studies of streptococci microevolution have transferred to knowledge about the therapeutic developments (Gao et al., 2014). Although our phylogenetic dendrograms clarify strain diversity according to 16S rRNA, evaluating other genetic factors related to virulence or adaptation will further elucidate pathogenic and evolutionary mechanisms.
Table 3.
Epidemiological specification (accession number, strain, and geographical location) of Streptococcus bacterial agents based on 16S rRNA gene sequences from the NCBI database
| Species | Accession number | Strain | Geographical region/country |
|---|---|---|---|
| Streptococcus iniae | DQ985468.1 | CGX | China |
| KY781829.1 | HNM-1 | China | |
| KJ162337.1 | Ab130920 | China | |
| KF815728.1 | WZMH110819 | China | |
| KF555592.1 | NS1-2011 | Thailand | |
| KC748467.1 | FC0924 | China | |
| KM209199.1 | SK10-S | Indonesia | |
| AB593340.1 | Feb-45 | Japan | |
| Streptococcus parauberis | AY942573.1 | LMG 14376 | Finland |
| FJ009631.1 | JJI51 | Korea | |
| JQ780604.1 | partial sequence | Israel | |
| KC836715.1 | RU37-6 | China | |
| AF284579.2 | SAP 99 | Italy | |
| KP137361.1 | F21 | Turkey | |
| KP137342.1 | F57 | Turkey | |
| KP240952.1 | CIFT MFB 10119(2) | India | |
| KC699192.1 | CNM465_12 | Spain | |
| Streptococcus agalactiae | LC071815.1 | JCM 5671 | Japan |
| AB596948.1 | JCM 5671 | Japan | |
| DQ303183.1 | ATCC 13813 | Canada | |
| AB002479.1 | ATCC 13813-NCTC 8181 | Japan | |
| NR_117503.1 | ATCC 13813 | USA | |
| NR_115728.1 | ATCC 13813 | USA | |
| GU360730.1 | ATCC 13813 | Netherlands | |
| KT869025.1 | SAG | Malaysia | |
| KY635952.1 | S29 | Brazil | |
| KY635949.1 | S73 | Brazil | |
| Streptococcus dysgalactiae | AB002485.1 | ATCC 43078 | Japan |
| AB002500.1 | isolate L32 | Japan | |
| AB002509.1 | isolate L9 | Japan | |
| NR_027517.1 | ATCC 43078 | USA | |
| DQ232540.1 | CIP 105120 | France | |
| JN639380.1 | CCUG 7977A | Denmark | |
| JN639434.1 | SK1333 | Denmark | |
| JN639432.1 | CCUG 36637 | Denmark | |
| JN639410.1 | CCUG 48101 | Denmark | |
| AB002484.1 | ATCC 27957 | Japan | |
| AY121361.1 | ATCC 12394 | China | |
| AF015928.1 | ATCC 27957 | USA | |
| AJ314611.1 | AC-2074 | Germany | |
| AJ314609.1 | AC-2713 | Germany | |
| AJ314610.1 | AC-2832 | Germany | |
| Lactococcus garvieae | LC145570.1 | JCM 12256 | Japan |
| KF111340.1 | TRF1 | USA | |
| HM536980.1 | PW1537 | China | |
| KX671996.1 | FJ6 | Iran | |
| KF849271.1 | SI-IRI | Iran | |
| AF352164.1 | FLG4 | China | |
| KM659863.1 | Fish 10/10 LKF | South Africa | |
| Vagococcus salmoninarum | AM490375.1 | JIP 20-00 | France |
| AM490374.1 | JIP 27-01(2) | France |
Fig. 4. Phylogenetic analysis of total 51 bacterial strains of Streptococcus species (S. parauberis, S. iniae, S. agalactiae, Lactococcus garvieae, S. dysgalactiae, and Vagococcus salmoninarum) causing streptococcal diseases based on 16S rDNA sequences from the NCBI database.

The evolutionary history was inferred using the Neighbor-Joining method. The optimal tree with the sum of branch length = 0.54759876 is shown. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (100 replicates) is shown next to the branches.
Fully developed fishery management systems according to current challenges (diagnosis, pathogenesis, and control mechanism) are required. Understanding fish management at the local level is useful for the aquaculture industry (Aquilera et al., 2015). We therefore suggest an effective aquaculture management module that utilizes a scientific approach to reconcile complex data of various cellular, molecular, and environmental approaches, and monitoring programs generated through research surveys.
Our study has important implications for the epidemiology of streptococcosis in fish, provides important information about the current scenario and challenges in the fish industry, and suggests joint molecular (for diagnosis) and cellular (for control) strategies along with environmental control methods as appropriate (Tables 1, 2 and Figs. 1–4). The above integrated strategies should focus on a worldwide sampling network for appropriate evolutionary monitoring through comparative genomics, then combine this information with ongoing aquaculture management systems.
CONCLUSION AND FUTURE PROSPECTS
Streptococcal infectious diseases, along with complicated control mechanisms, have contributed to a considerable decrease in fish production. This review highlights the current status of Streptococcus bacterial diagnosis, pathogenesis, hazardous effects of the host fish, and the overall negative economic impacts on the fish industry. In the absence of extensive knowledge about virulence, new emerging strains, treatments, and control of streptococcal disease, streptococcosis is becoming difficult to control. Based on the information presented in this review, we recommend early diagnosis using molecular methods and improved cellular research along with environmental control through appropriate managerial decisions considering the present scenario.
Supplementary data
ACKNOWLEDGMENTS
This research was a part of the project titled “Omics based on fishery disease control technology development and industrialization (20150242)”, funded by the Ministry of Oceans and Fisheries, Korea. Anshuman Mishra would like to acknowledge PRF in Varanasi, India.
Footnotes
Note: Supplementary information is available on the Molecules and Cells website (www.molcells.org).
REFERENCES
- Abdelsalam M., Chen S.-C., Yoshida T. Dissemination of streptococcal pyrogenic exotoxin G (spegg) with an IS-like element in fish isolates of Streptococcus dysgalactiae. FEMS Microbiol Lett. 2010;309:105–113. doi: 10.1111/j.1574-6968.2010.02024.x. [DOI] [PubMed] [Google Scholar]
- Abdelsalam M., Asheg A., Eissa A.E. Streptococcus dysgalactiae: An emerging pathogen of fishes and mammals. Int J Vet Sci Med. 2013;1:1–6. [Google Scholar]
- Achtman M. Evolution, population structure, and phylogeography of genetically monomorphic bacterial pathogens. Annu Rev Microbiol. 2008;62:53–70. doi: 10.1146/annurev.micro.62.081307.162832. [DOI] [PubMed] [Google Scholar]
- Agnew W., Barnes A.C. Streptococcus iniae: an aquatic pathogen of global veterinary significance and a challenging candidate for reliable vaccination. Vet Microbiol. 2007;122:1–15. doi: 10.1016/j.vetmic.2007.03.002. [DOI] [PubMed] [Google Scholar]
- Al-Harbi A.H. Molecular characterization of Streptococcus iniae isolated from hybrid tilapia (Oreochromis niloticus× Oreochromis aureus) Aquaculture. 2011;312:15–18. [Google Scholar]
- Aguilera S.E., Cole J., Finkbeiner E.M., Cornu E.L., Ban N.C., Carr M.H., Cinner J.E., Crowder L.B., Gelcich S., Hicks C.C., et al. Managing small-scale commercial fisheries for adaptive capacity: insights from dynamic social-ecological drivers of change in monterey bay. PLoS One. 2015;10:e0118992. doi: 10.1371/journal.pone.0118992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Austin B., Austin D.A. Bacterial fish pathogens: disease of farmed and wild fish. Springer Science & Business Media; 2007. [Google Scholar]
- Baeck G.W., Kim J.H., Gomez D.K., Park S.C. Isolation and characterization of Streptococcus sp. from diseased flounder (Paralichthys olivaceus) in Jeju Island. J Vet Sci. 2006;7:53–58. doi: 10.4142/jvs.2006.7.1.53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barnes A.C., Ellis A.E. Role of capsule in serotypic differences and complement fixation by Lactococcus garvieae. Fish Shellfish Immunol. 2004;16:207–214. doi: 10.1016/S1050-4648(03)00079-2. [DOI] [PubMed] [Google Scholar]
- Barnes A.C., Guyot C., Hansen B.G., Mackenzie K., Horne M.T., Ellis A.E. Resistance to serum killing may contribute to differences in the abilities of capsulate and non-capsulated isolates ofLactococcus garvieae to cause disease in rainbow trout (Oncorhynchus mykiss L.) Fish Shellfish Immunol. 2002;12:155–168. doi: 10.1006/fsim.2001.0361. [DOI] [PubMed] [Google Scholar]
- Barnett T.C., Cole J.N., Rivera-Hernandez T., Henningham A., Paton J.C., Nizet V., Walker M.J. Streptococcal toxins: role in pathogenesis and disease. Cell Microbiol. 2015;17:1721–1741. doi: 10.1111/cmi.12531. [DOI] [PubMed] [Google Scholar]
- Bennedsen M., Stuer-Lauridsen B., Danielsen M., Johansen E. Screening for antimicrobial resistance genes and virulence factors via genome sequencing. Appl Environ Microbiol. 2011;77:2785–2787. doi: 10.1128/AEM.02493-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bondad-Reantaso M.G., Subasinghe R.P., Arthur J.R., Ogawa K., Chinabut S., Adlard R., Tan Z., Shariff M. Disease and health management in Asian aquaculture. Vet Parasitol. 2005;132:249–272. doi: 10.1016/j.vetpar.2005.07.005. [DOI] [PubMed] [Google Scholar]
- Bosshard P.P., Zbinden R., Abels S., Boddinghaus B., Altwegg M., Bottger E.C. 16S rRNA gene sequencing versus the API 20 NE system and the VITEK 2 ID-GNB card for identification of nonfermenting Gram-negative bacteria in the clinical laboratory. J Clin Microbiol. 2006;44:1359–1366. doi: 10.1128/JCM.44.4.1359-1366.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brimil N., Barthell E., Heindrichs U., Kuhn M., Lütticken R., Spellerberg B. Epidemiology of Streptococcus agalactiae colonization in Germany. Int J Med Microbiol. 2006;296:39–44. doi: 10.1016/j.ijmm.2005.11.001. [DOI] [PubMed] [Google Scholar]
- Bromage E.S., Thomas A., Owens L. Streptococcus iniae, a bacterial infection in barramundi Lates calcarifer. Dis Aquat Organ. 1999;36:177–181. doi: 10.3354/dao036177. [DOI] [PubMed] [Google Scholar]
- Buchanan J.T., Colvin K.M., Vicknair M.R., Patel S.K., Timmer A.M., Nizet V. Strain-associated virulence factors of Streptococcus iniae in hybrid-striped bass. Vet Microbiol. 2008;131:145–153. doi: 10.1016/j.vetmic.2008.02.027. [DOI] [PubMed] [Google Scholar]
- Carson J., Gudkovs N., Austin B. Characteristics of an Enterococcus-like bacterium from Australia and South Africa, pathogenic for rainbow trout, Oncorhynchus mykiss (Walbaum) J Fish Dis. 1993;16:381–388. [Google Scholar]
- Carey-Ann D., Burnhama, Gregory J.T. Virulence factors of group B streptococci. Rev Med Microbiol. 2003;14:109–118. [Google Scholar]
- Chain P.S., Grafham D.V., Fulton R.S., Fitzgerald M.G., Hostetler J., Muzny D., Ali J., Birren B., Bruce D.C., Buhay C., et al. Genomics. Genome project standards in a new era of sequencing. Science. 2009;326:236–237. doi: 10.1126/science.1180614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang P., Plumb J. Effects of salinity on Streptococcus infection of Nile tilapia, Oreochromis niloticus. J Appl Aquaculture. 1996;6:39–45. [Google Scholar]
- Cheng S., Hu Y.H., Jiao X.D., Sun L. Identification and immunoprotective analysis of a Streptococcus iniae subunit vaccine candidate. Vaccine. 2010;28:2636–2641. doi: 10.1016/j.vaccine.2010.01.016. [DOI] [PubMed] [Google Scholar]
- Chettri J.K., Raida M.K., Holten-Andersen L., Kania P.W., Buchmnann K. PAMP induced expression of immune relevant genes in head kidney leukocytes of rainbow trout (Oncorhynchus mykiss) Dev. Comp Immunol. 2011;35:476–482. doi: 10.1016/j.dci.2010.12.001. [DOI] [PubMed] [Google Scholar]
- Clarridge J.E., Attorri S.M., Zhang Q., Bartell J. 16S ribosomal DNA sequence analysis distinguishes biotypes of Streptococcus bovis: Streptococcus bovis Biotype II/2 is a separate genospecies and the predominant clinical isolate in adult males. J Clin Microbiol. 2001;39:1549–1552. doi: 10.1128/JCM.39.4.1549-1552.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darwish A.M., Hobbs M.S. Laboratory efficacy of amoxicillin for the control of Streptococcus iniae infection in blue tilapia. J Aquat Anim Health. 2005;17:197–202. [Google Scholar]
- Delannoy C., Zadoks R., Crumlish M., Rodgers D., Lainson F., Ferguson H., Turnbull J., Fontaine M. Genomic comparison of virulent and non-virulent Streptococcus agalactiae in fish. J Fish Dis. 2016;39:13–29. doi: 10.1111/jfd.12319. [DOI] [PubMed] [Google Scholar]
- Diler O., Altun S., Adiloglu A., Kubilay A., Istklt B. First occurrence of Streptococcosis affecting farmed rainbow trout (Oncorhynchus mykiss) in Turkey. Bull Eur Ass Fish Pathol. 2002;22:21–26. [Google Scholar]
- Dobrindt U., Hacker J. Whole genome plasticity in pathogenic bacteria. Curr Opin Microbiol. 2001;4:550–557. doi: 10.1016/s1369-5274(00)00250-2. [DOI] [PubMed] [Google Scholar]
- Dodson S., Maurer J., Shotts E. Biochemical and molecular typing of Streptococcus iniae isolated from fish and human cases. J Fish Dis. 1999;22:331–336. [Google Scholar]
- Eldar A., Bejerano Y., Livoff A., Horovitcz A., Bercovier H. Experimental streptococcal meningo-encephalitis in cultured fish. Vet Microbiol. 1995;43:33–40. doi: 10.1016/0378-1135(94)00052-x. [DOI] [PubMed] [Google Scholar]
- Eyngor M., Zlotkin A., Ghittino C., Prearo M., Douet D.-G., Chilmonczyk S., Eldar A. Clonality and diversity of the fish pathogen Lactococcus garvieae in Mediterranean countries. Appl Environ Microbiol. 2004;70:5132–5137. doi: 10.1128/AEM.70.9.5132-5137.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Facklam R., Elliott J., Shewmaker L., Reingold A. Identification and characterization of sporadic isolates of Streptococcus iniae isolated from humans. J Clin Microbiol. 2005;43:933–937. doi: 10.1128/JCM.43.2.933-937.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuller J.D., Bast D.J., Nizet V., Low D.E., de Azavedo J.C. Streptococcus iniae virulence is associated with a distinct genetic profile. Infect Immun. 2001;69:1994–2000. doi: 10.1128/IAI.69.4.1994-2000.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao X.Y., Zhi X.Y., Li H.W., Klenk H.P., Li W.J. Comparative genomics of the bacterial genus Streptococcus illuminates evolutionary implications of species groups. PLoS One. 2014;9:e101229. doi: 10.1371/journal.pone.0101229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghittino C., Accornero P., Prearo M., Rogato F., Zlotkin A., Eldar A. Coldwater streptococcoses in salmonids, with particular reference to Vagococcus salmoninarum infection. Proceedings of Workshop in Fish Streptococcoses, IZS–State Veterinary Institute; Turin, Italy. 1999. [Google Scholar]
- Hastein T., Gudding R., Evensen O. Bacterial vaccines for fish--an update of the current situation worldwide. Dev Biol (Basel) 2005;121:55–74. [PubMed] [Google Scholar]
- Holmer M. Environmental issues of fish farming in offshore waters: perspectives, concerns and research needs. Aquac Environ Interact. 2010;1:57–70. [Google Scholar]
- Johri A.K., Paoletti L.C., Glaser P., Dua M., Sharma P.K., Grandi G., Rappuoli R. Group B Streptococcus: global incidence and vaccine development. Nat Rev Microbiol. 2006;4:932–942. doi: 10.1038/nrmicro1552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jolley K.A., Bliss C.M., Bennett J.S., Bratcher H.B., Brehony C., Colles F.M., Wimalarathna H., Harrison O.B., Sheppard S.K., Cody A.J., et al. Ribosomal multilocus sequence typing: universal characterization of bacteria from domain to strain. Microbiology. 2012;158:1005–1015. doi: 10.1099/mic.0.055459-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klesius P., Evans J., Shoemaker C., Yeh H., Goodwin A., Adams A., Thompson K. Rapid detection and identification of Streptococcus iniae using a monoclonal antibody-based indirect fluorescent antibody technique. Aquaculture. 2006;258:180–186. [Google Scholar]
- Klijn N., Weerkamp A.H., de Vos W.M. Identification of mesophilic lactic acid bacteria by using polymerase chain reaction-amplified variable regions of 16S rRNA and specific DNA probes. Appl Environ Microbiol. 1991;57:3390–3393. doi: 10.1128/aem.57.11.3390-3393.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lau S.K., Woo P.C., Luk W.K., Fung A.M., Hui W.T., Fong A.H., Chow C.W., Wong S.S., Yuen K.Y. Clinical isolates of Streptococcus iniae from Asia are more mucoid and beta-hemolytic than those from North America. Diagn Microbiol Infect Dis. 2006;54:177–181. doi: 10.1016/j.diagmicrobio.2005.09.012. [DOI] [PubMed] [Google Scholar]
- Li L., Wang R., Liang W., Huang T., Huang Y., Luo F., Lei A., Chen M., Gan X. Development of live attenuated Streptococcus agalactiae vaccine for tilapia via continuous passage in vitro. Fish Shellfish Immunol. 2015;45:955–963. doi: 10.1016/j.fsi.2015.06.014. [DOI] [PubMed] [Google Scholar]
- Lowe B.A., Miller J.D., Neely M.N. Analysis of the polysaccharide capsule of the systemic pathogen Streptococcus iniae and its implications in virulence. Infect Immun. 2007;75:1255–1264. doi: 10.1128/IAI.01484-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magnadottir B. Innate immunity of fish (overview) Fish Shellfish Immunol. 2006;20:137–151. doi: 10.1016/j.fsi.2004.09.006. [DOI] [PubMed] [Google Scholar]
- Maiden M.C. Multilocus sequence typing of bacteria. Annu Rev Microbiol. 2006;60:561–588. doi: 10.1146/annurev.micro.59.030804.121325. [DOI] [PubMed] [Google Scholar]
- Medini D., Serruto D., Parkhill J., Relman D.A., Donati C., Moxon R., Falkow S., Rappuoli R. Microbiology in the post-genomic era. Nat Rev Microbiol. 2008;6:419–430. doi: 10.1038/nrmicro1901. [DOI] [PubMed] [Google Scholar]
- Mishra A., Nam G.H., Gim J.A., Seong M., Choe Y., Lee H.E., Jo A., Kim S., Kim D.H., Cha H.J., et al. Comparative evaluation of 16S rRNA gene in world-wide strains of Streptococcus iniae and Streptococcus parauberis for early diagnostic marker. Genes Genom. 2017;39:779–791. [Google Scholar]
- Mora D., Ricci G., Guglielmetti S., Daffonchio D., Fortina M.G. 16S–23S rRNA intergenic spacer region sequence variation in Streptococcus thermophilus and related dairy streptococci and development of a multiplex ITS-SSCP analysis for their identification. Microbiology. 2003;149:807–813. doi: 10.1099/mic.0.25925-0. [DOI] [PubMed] [Google Scholar]
- Muzquiz J., Royo F., Ortega C., De Blas I., Ruiz I., Alonso J. Pathogenicity of streptococcosis in rainbow trout (Oncorhynchus mykiss): dependence on age of diseased fish. Bull Eur Ass Fish Pathol. 1999;19:114–119. [Google Scholar]
- Kwong J.C., McCallum N., Sintchenko V., Howden B.P. Whole genome sequencing in clinical and public health microbiology. Pathology. 2015;47:199–210. doi: 10.1097/PAT.0000000000000235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nho S.W., Shin G.W., Park S.B., Jang H.B., Cha I.S., Ha M.A., Kim Y.R., Park Y.K., Dalvi R.S., Kang B.J., et al. Phenotypic characteristics of Streptococcus iniae and Streptococcus parauberis isolated from olive flounder (Paralichthys olivaceus) FEMS Microbiol Lett. 2009;293:20–27. doi: 10.1111/j.1574-6968.2009.01491.x. [DOI] [PubMed] [Google Scholar]
- Nho S.W., Hikima J., Park S.B., Jang H.B., Cha I.S., Yasuike M., Nakamura Y., Fujiwara A., Sano M., Kanai K., et al. Comparative genomic characterization of three Streptococcus parauberis strains in fish pathogen, as assessed by wide-genome analyses. PLoS One. 2013;8:e80395. doi: 10.1371/journal.pone.0080395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nielsen X.C., Justesen U.S., Dargis R., Kemp M., Christensen J.J. Identification of clinically relevant nonhemolytic Streptococci on the basis of sequence analysis of 16S–23S intergenic spacer region and partial gdh gene. J Clin Microbiol. 2009;47:932–939. doi: 10.1128/JCM.01449-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishiki I., Noda M., Itami T., Yoshida T. Homogeneity of Streptococcus dysgalactiae from farmed amberjack Seriola dumerili in Japan. Fish Sci. 2010;76:661–668. [Google Scholar]
- Park Y.K., Nho S.W., Shin G.W., Park S.B., Jang H.B., Cha I.S., Ha M.A., Kim Y.R., Dalvi R.S., Kang B.J., et al. Antibiotic susceptibility and resistance of Streptococcus iniae and Streptococcus parauberis isolated from olive flounder (Paralichthys olivaceus) Vet Microbiol. 2009;136:76–81. doi: 10.1016/j.vetmic.2008.10.002. [DOI] [PubMed] [Google Scholar]
- Pereira U., Mian G., Oliveira I., Benchetrit L., Costa G., Figueiredo H. Genotyping of Streptococcus agalactiae strains isolated from fish, human and cattle and their virulence potential in Nile tilapia. Vet Microbiol. 2010;140:186–192. doi: 10.1016/j.vetmic.2009.07.025. [DOI] [PubMed] [Google Scholar]
- Rajagopal L. Understanding the regulation of Group B Streptococcal virulence factors. Future Microbiol. 2009;4:201–221. doi: 10.2217/17460913.4.2.201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ravelo C., Magariños B., Romalde J.L., Toranzo A.E. Conventional versus miniaturized systems for the phenotypic characterization of Lactococcus garvieae strains. Bull Eur Ass Fish Pathol. 2001;21:136–144. [Google Scholar]
- Ruiz-Zarzuela I., de Bias I., Gironés O., Ghittino C., Múazquiz J. Isolation of Vagococcus salmoninarum in rainbow trout, Oncorhynchus mykiss (Walbaum), broodstocks: characterization of the pathogen. Vet Res Commun. 2005;29:553–562. doi: 10.1007/s11259-005-2493-8. [DOI] [PubMed] [Google Scholar]
- Sakai M. Current research status of fish immunostimulants. Aquaculture. 1999;172:63–92. [Google Scholar]
- Sakala R., Hayashidani H., Kato Y., Kaneuchi C., Ogawa M. Isolation and characterization of Lactococcus piscium strains from vacuum-packaged refrigerated beef. J Appl Microbiol. 2002;92:173–179. doi: 10.1046/j.1365-2672.2002.01513.x. [DOI] [PubMed] [Google Scholar]
- Shoemaker C.A., Evans J.J., Klesius P.H. Density and dose: factors affecting mortality of Streptococcusiniae infected tilapia (Oreochromisniloticus) Aquaculture. 2000;188:229–235. [Google Scholar]
- Shoemaker C.A., Klesius P.H., Evans J.J. Prevalence of Streptococcus iniae in tilapia, hybrid striped bass, and channel catfish on commercial fish farms in the United States. Am J Vet Res. 2001;62:174–177. doi: 10.2460/ajvr.2001.62.174. [DOI] [PubMed] [Google Scholar]
- Sommerset I., Krossoy B., Biering E., Frost P. Vaccines for fish in aquaculture. Exp Rev Vaccines. 2005;4:89–101. doi: 10.1586/14760584.4.1.89. [DOI] [PubMed] [Google Scholar]
- Springman A.C., Lacher D.W., Wu G., Milton N., Whittam T.S., Davies H.D., Manning S.D. Selection, Recombination, and Virulence gene diversity among Group b Streptococcal Genotypes. J Bacteriol. 2009;17:5419–5427. doi: 10.1128/JB.00369-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teng L.J., Hsueh P.R., Tsai J.C., Chen P.W., Hsu J.C., Lai H.C., Lee C.N., Ho S.W. groESL sequence determination, phylogenetic analysis, and species differentiation for viridans group streptococci. J Clin Microbiol. 2002;40:3172–3178. doi: 10.1128/JCM.40.9.3172-3178.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toranzo A.E., Magariños B., Romalde J.L. A review of the main bacterial fish diseases in mariculture systems. Aquaculture. 2005;246:37–61. [Google Scholar]
- Tung S.K., Teng L.J., Vaneechoutte M., Chen H.M., Chang T.C. Identification of species of Abiotrophia, Enterococcus, Granulicatella and Streptococcus by sequence analysis of the ribosomal 16S–23S intergenic spacer region. J Med Microbiol. 2007;56:504–513. doi: 10.1099/jmm.0.47027-0. [DOI] [PubMed] [Google Scholar]
- Vendrell D., Balcázar J.L., Ruiz-Zarzuela I., De Blas I., Gironés O., Múzquiz J.L. Lactococcus garvieae in fish: a review. Comp Immunol Microbiol Infect Dis. 2006;29:177–198. doi: 10.1016/j.cimid.2006.06.003. [DOI] [PubMed] [Google Scholar]
- Villegas J.G., Fukada H., Masumoto T., Hosokawa H. Effect of Dietary Immunostimulants on Some Innate Immune Responses and Disease Resistance against Edwardsiella tarda Infection in Japanese Flounder (Paralichthys olivaceus) Aquaculture Science. 2006;2:153–162. [Google Scholar]
- Weinstein M.R., Litt M., Kertesz D.A., Wyper P., Rose D., Coulter M., McGeer A., Facklam R., Ostach C., Willey B.M., et al. Invasive infections due to a fish pathogen, Streptococcus iniae. S. iniae Study Group. N Engl J Med. 1997;337:589–594. doi: 10.1056/NEJM199708283370902. [DOI] [PubMed] [Google Scholar]
- Woo S.H., Park S.I. Effects of phosphoglucomutase gene (PGM) in Streptococcus parauberis on innate immune response and pathogenicity of olive flounder (Paralichthys olivaceus) Fish Shellfish Immunol. 2014;41:317–325. doi: 10.1016/j.fsi.2014.09.012. [DOI] [PubMed] [Google Scholar]
- Wren B.W. Microbial genome analysis: insights into virulence, host adaptation and evolution. Nat Rev Genet. 2000;1:30–39. doi: 10.1038/35049551. [DOI] [PubMed] [Google Scholar]
- Yoon S.H., Ha S.M., Kwon S., Lim J., Kim Y., Seo H., Chun J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA and whole genome assemblies. Int J Syst Evol Microbiol. 2017;67:1613–1617. doi: 10.1099/ijsem.0.001755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zlotkin A., Chilmonczyk S., Eyngor M., Hurvitz A., Ghittino C., Eldar A. Trojan horse effect: phagocyte-mediated Streptococcus iniae infection of fish. Infect Immun. 2003;71:2318–2325. doi: 10.1128/IAI.71.5.2318-2325.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.