

Abstract
Background
We have previously shown that 8 weeks’ treatment with phenylbutyrate (PBA) (500mgx2/day) with or without vitamin D3 (vitD3) (5000 IU/day) as host-directed therapy (HDT) accelerated clinical recovery, sputum culture conversion and increased expression of cathelicidin LL-37 by immune cells in a randomized, placebo-controlled trial in adults with pulmonary tuberculosis (TB). In this study we further aimed to examine whether HDT with PBA and vitD3 promoted clinically beneficial immunomodulation to improve treatment outcomes in TB patients.
Methods
Cytokine concentration was measured in supernatants of peripheral blood mononuclear cells (PBMC) from patients (n = 31/group). Endoplasmic reticulum stress-related genes (GADD34 and XBP1spl) and human beta-defensin-1 (HBD1) gene expression were studied in monocyte-derived-macrophages (MDM) (n = 18/group) from PBMC of patients. Autophagy in MDM (n = 6/group) was evaluated using LC3 expression by confocal microscopy.
Results
A significant decline in the concentration of cytokines/chemokines was noted from week 0 to 8 in the PBA-group [TNF-α (β = − 0.34, 95% CI = − 0.68, − 0.003; p = 0.04), CCL11 (β = − 0.19, 95% CI = − 0.36, − 0.03; p = 0.02) and CCL5 (β = − 0.08, 95% CI = − 0.16, 0.002; p = 0.05)] and vitD3-group [(CCL11 (β = − 0.17, 95% CI = − 0.34, − 0.001; p = 0.04), CXCL10 (β = − 0.38, 95% CI = − 0.77, 0.003; p = 0.05) and PDGF-β (β = − 0.16, 95% CI = − 0.31, 0.002; p = 0.05)] compared to placebo. Both PBA- and vitD3-groups showed a decline in XBP1spl mRNA on week 8 (p < 0.03). All treatment groups demonstrated increased LC3 expression in MDM compared to placebo over time (p < 0.037).
Conclusion
The use of PBA and vitD3 as adjunct therapy to standard TB treatment promoted favorable immunomodulation to improve treatment outcomes.
Trials registration
This trial was retrospectively registered in clinicaltrials.gov, under identifier NCT01580007.
Electronic supplementary material
The online version of this article (10.1186/s12879-018-3203-9) contains supplementary material, which is available to authorized users.
Keywords: Mycobacterium tuberculosis, Cytokines, Chemokines, Endoplasmic reticulum stress, Human beta-defensin-1 (HBD1)
Background
Tuberculosis remains among the top 10 leading causes of death with global estimates of 10.4 million new cases and 1.4 million deaths in 2015 [1]. Control or exacerbation of TB is dependent on host immune responses generated to combat Mycobacterium tuberculosis (Mtb) infection [2–4]. Several studies have unravelled immunological pathways that influence the outcome of Mtb infection which include cytokine-mediated signalling among T cells, macrophages and neutrophils, and phagocytes-mediated antimicrobial processes [4–9]. Studying cytokine profiles in TB patients has demonstrated its potential for use in diagnostic purposes, monitoring of treatment efficacy and development of novel treatment strategies [10–14]. Autophagy in macrophages and intracellular lysosomal degradation are important for killing of pathogens although Mtb has evolved to escape elimination by blocking phagosomal acidification and phagosome-lysosomal fusion [15–17]. Endoplasmic-reticulum (ER) stress response is triggered by mycobacterial infection and plays a critical role in the pathogenesis of TB [18].
The rise in antibiotic resistance among Mtb in the last decade rekindled attention towards alternative chemotherapies. Host-directed therapies (HDT) have emerged as a promising avenue for adjunctive treatment with the aim to modulate immune responses against the pathogen by targeting clinically relevant host biological pathways. This strategy would be beneficial in reducing the course of antibiotic treatment, preventing the spread of drug-resistant Mtb, and reducing inflammation in the lung (reviewed in [19, 20]). Phenylbutyrate (PBA) is a licensed drug indicated for the management of urea cycle disorders [21]. Vitamin D3 (vitD3), a dietary supplement, has diverse immune-modulatory properties. Our group has shown that PBA and vitD3 have a strong synergistic effect on induction of antimicrobial peptides (AMPs) in lung epithelial cell lines, macrophages, and in healthy human immune cells [22–24]. In a randomized clinical trial, we have shown that PBA alone or combined with vitD3 is a promising candidate for HDT in the treatment of drug sensitive pulmonary TB by speeding up clinical recovery [25]. Treatment with vitD3 or the combination with PBA accelerated sputum culture conversion and enhanced expression of LL-37, the human AMP of the cathelicidin family, by immune cells. Moreover, PBA adjunctive therapy increased macrophage-mediated killing of Mtb ex vivo compared to placebo. Our in vitro study further showed that PBA can induce autophagy in a LL-37 dependent pathway and promotes intracellular killing of Mtb in human macrophages [23]. PBA is known to reduce ER stress in cells and thereby reduce inflammatory responses [26, 27]. We hypothesized that the beneficial effects imparted by HDT of PBA and vitD3 in TB patients may be mediated through regulating expression of cytokines, chemokines and AMPs by immune cells, augmentation of autophagic responses of macrophages and reduction of chronic ER stress. Thus, in a sub-group of TB patients, we evaluated the effect of HDT on dynamics of cytokines and chemokines in culture supernatants of PBMC, HBD1 and ER stress genes and expression of LC3, an autophagy marker, in macrophages from TB patients in response to the disease and clinical improvement.
Methods
Patients, study design and interventions
For this study, we used materials collected during our previously published clinical trial [25]. Briefly, 288 adult patients with newly diagnosed sputum smear-positive TB (18–55 years of age) were recruited from the National Institute of the Diseases of the Chest and Hospital (NIDCH) in Dhaka, Bangladesh. The study was approved by the Research and Ethical Review Committees at icddr,b, an international health research institute based in Bangladesh. The study was a double-blind, placebo-controlled trial in which patients were randomized to the following adjunct therapy arms (72 × 4) receiving oral doses of either: (1) placebo PBA and placebo vitD3; or (2) 500 mg twice daily of PBA and placebo vitD3; or (3) placebo PBA and 5000 IU of vitD3 (Cholecalciferol) once daily; or (4) PBA combined with vitD3 (PBA + vitD3). In parallel, standard care of directly observed therapy short-course (DOTS) regimen consisting of isoniazid (INH), rifampicin (RIF), pyrazinamide, and ethambutol was given to all patients for 2 months followed by INH and RIF for 4 months. Clinical evaluation, sputum microscopy, sputum culture and chest radiography were performed [25]. A total of 249 patients (modified intention-to-treat) completed the week 12 follow-up and 219 completed week 24 follow-up visit.
In the published TB trial, clinical assessments were performed by the study doctor and were used to calculate numerical clinical scores which was defined as a TB score [25]. The TB score is an evaluation tool developed by clinicians to quantify/determine changes in clinical symptoms of the TB patients in an impartial and objective manner. The TB score allocated points for self-reported symptoms (cough, shortness of breath/dyspnea, chest pain, haemoptysis, anorexia), and clinical signs assessed by study doctors (fever, anemia (< 11 g/dl), tachycardia, auscultatory findings). The TB score was determined at week 0–4, 6, 8, 10, 12 and 24.
For the present study, a sub-sample of 127 patients (32/group) was selected for studying cytokine and chemokine responses of PBMC for which adequate volume and complete set of PBMC culture supernatants at 2 time points (week 0 and 8) were available. In the vitD3 group, a complete set of 31 patients were only available. ER stress (18/group) and HBD1 (18/group) genes were studied using cDNA synthesized from monocyte-derived-macrophages (MDM) of the TB patients at week 0, 4 and 8. For studying autophagy marker, MDM from 24 patients (6/group) were selected with adequate number of cells available at 4 time points (week 0, 4, 8 and 12).
Samples and cell culture supernatant
Peripheral blood mononuclear cells (PBMC) and plasma were separated from whole blood by Ficoll-PaqueTM PLUS (GE Healthcare, Uppsala, Sweden) density gradient centrifugation. PBMC were used directly following isolation and were not stored in liquid nitrogen for future use. After washing, PBMCs were suspended in culture medium and cultured for 2 days. Intracellular fluid was collected after adding 0.1% saponin (Sigma-Aldrich, Steinheim, Germany) and stored in ultra-low freezer for analyses of cytokines (Fig. 1). Forty-eight hours of incubation of PBMC without any stimulation allows partial maturation of monocytes in the mixed cell culture (T and B lymphocytes, NK cells, plasma cells and monocytes). There is spontaneous and active release of cytokines or antibodies directly from the immune cells into the culture fluid (extracellular cytokines); reversible permeabilization of PBMC by saponin additionally allows excretion of intra-cellular cytokines into the supernatant.
Fig. 1.

The flow chart illustrates collection, processing and storage of different cell types from freshly isolated peripheral blood mononuclear cells (PBMC), the time periods of cell culture and the various signature immune markers evaluated in these patients
Five million PBMC were plated in 4-well culture plates (NUNC, Roskilde, Denmark) for three days, after removing nonadherant cells the adherent MDMs were harvested using a cell scraper, treated with RNAlater (Qiagen GmbH, Hilden, Germany). Total RNA was isolated from MDM using the RNeasy Mini Kit (Qiagen GmbH). mRNA was reverse-transcribed and corresponding cDNA was synthesized using the SuperScript III First-Strand Synthesis System (Invitrogen Life Technologies, CA) with CFX96 real-time PCR detection System (Bio-Rad, Hercules, CA, USA). The cDNA was used for analysis of ER stress-related genes and HBD1 (Fig. 1).
Cytokines and chemokines
Multiplex kits (Bio-Rad) were used to analyze cytokines and chemokines in the cell supernatants of PBMC following manufacturer’s instructions in the Bioplex 200 system (Bio-Rad). Two types of kits were used, (i) the 27-plex kit contained a panel of 27 analytes: IL-1β, IL-1ra, IL-2, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-12(p70), IL-13, IL-15, IL-17, CCL11 (eotaxin), FGF basic, G-CSF, GM-CSF, IFN-γ, CXCL10 (IP-10), CCL2 (MCP-1), CCL3 (MIP-1α), CCL4 (MIP-1β), PDGF-BB, CCL5 (RANTES), TNF-α, and VEGF. (ii) The 17-plex kit contained: IL-1β, IL-2, IL-4, IL-5, IL-6, IL-7, IL-8, IL-10, IL-12(p70), IL-13, IL-17, G-CSF, GM-CSF, IFN-γ, CCL2, CCL3 and TNF-α. For 75 patients 27-plex kit was used (19/group; 18/vitD3 group), while for the rest of 52 patients (13/group) the 17-plex kit was utilized, therefore results for 8 analytes were not available for 52 patients.
Autophagy marker in macrophages
Autophagy is a physiological process; it is applied by macrophages to control growth or elimination of intracellular pathogens. LC3 is a classical marker and most widely used for assessing the autophagy process LC3-I is the cytosolic form of this protein. Upon activation of autophagy, LC3-I is subsequently conjugated with phosphatidylethanolamine (PE) to generate LC3-II by a ubiquitination-like enzymatic reaction. In contrast to the cytoplasmic localization of LC3-I, LC3-II associates with both the outer and inner membranes of the autophagosome. This LC3-II protein looks like dots and is known as ‘Puncta’. In a previous study, we have shown that treatment of MDM with PBA increased the expression of LC3 with simultaneous decrease in p62 protein expression compared to untreated MDM [23]. Due to the availability of limited number of cells from patients at each time point, simultaneous staining of both LC3 and p62 was not feasible.
Freshly isolated PBMCs from TB patients [25] (above) were cultured in culture medium for 3 days in 8-well chamber slides (Nunc® Lab-Tek® Chamber Slide™ system). After removal of non-adherent cells, adherent MDM were infected with virulent Mtb strain, H37Rv (Tuberculosis Research Center, Chennai, India) at a multiplicity of infection (MOI) of 1:5 (1 macrophage to 5 bacilli (Mtb)) in culture media for 2 h without antibiotics in the BSL-3 facility. After washing to remove the extracellular bacteria, the infected cells were fixed with 4% paraformaldehyde (Sigma-Aldrich) in phosphate buffered saline and stored in − 80 °C until shipped to the Laboratory in Karolinska Institute. The frozen slides were brought to room temperature, and staining was performed as mentioned earlier [23]. Quantification of autophagy was performed based on the percentage of the cells with LC3 punctate dots by ImageJ software (National Institutes of Health, US).
Endoplasmic reticulum stress in MDM
As markers of ER stress, growth arrest and DNA damage inducible protein 34 (GADD34) and spliced X-box binding protein-1 (XBP1spl) mRNA were assessed in MDM from TB patients (Fig. 1). The qPCR method was performed as described previously [28] using the primer pairs as follows: GADD34 forward primer 5’-ATGTATGGTGAGCGAGAGGC-3′, reverse primer 5’-GCAGTGTCCTTATCAGAAGGC-3′; XBP1spl forward primer 5’-TGCTGAGTCCGCA-GCAGGTG-3′, reverse primer 5’-GCTGGCAGGCTCTGGGGAAG-3′. Relative mRNA concentration of the reference gene ATP5B, (GeNorm, PrimerDesign Ltd., Southampton, UK) was used to calculate normalized expression. Each assay was run on a CFX96 (Bio-Rad) in triplicates and arbitrary mRNA concentrations were calculated by the Bio-Rad software, using the relative standard curve method.
Human beta-defensin-1 (HBD1) in MDM
Expression of the gene encoding human β-defensin-1 was measured by qPCR using CFX96 (Bio-Rad) (Fig. 1). The following primer sequences were used for quantification of HBD1. Sense primer: 5′-ATGGCCTCA-GGTGGTAACTTTC-3′; antisense primer: 5′-CACTTGGCCTTCCCTCTGT-AAC-3′. Relative quantification method was used to the housekeeping RNA18S/18S rRNA (18S rRNA–housekeeping gene kit, Applied Biosystems, Foster. City, USA). Results were expressed as relative gene expression and the 2–ΔΔCT method or fold changes. ΔCt values were calculated by subtracting 18 s rRNA Ct values from HBD1 Ct values of the same sample. Thereafter, ΔΔCt was obtained by subtracting average placebo week 0 ΔCt value from all ΔCt values. Finally, HBD1 relative gene expression was calculated by the formula, 2-ΔΔCt [29].
Statistical analysis
Distribution of different adjunct treatments (Placebo, PBA, vitD3 and PBA + vitD3) were calculated based on demographic characteristics (age, gender, history of contact, body weight, vitamin D status). Cytokine and chemokine data was log transformed due to non-normal distribution, and a simple interpretation of the beta-coefficient (average changes in outcome (concentration of cytokines, chemokines, number (%) of autophagy marker LC3 etc.) associated with change in treatment group with respect to placebo). Cytokine and chemokine responses of each of the 127 patients were studied at 2 time points, at week 0 and week 8. Wilcoxon Signed Ranks test was performed to evaluate the changes in cytokine and chemokine concentrations within each treatment group. To assess the treatment effects on changes in outcome variables (cytokine/chemokine concentrations and autophagy marker LC3+ cell counts) over time, generalized estimating equation (GEE) model was performed considering exchangeable correlation matrix. Each cytokine, chemokine and LC3+ cell counts (autophagy marker) was individually analyzed using the GEE model allowing repeated measures per patient to reduce longitudinal multicollinearity. The treatment effects were adjusted by covariates that influence the model by at least 5% (age, sex, duration of symptoms, history of contact with active TB cases and time at week 0 and week 8 (week 4 and 12 were considered where applicable)). The interaction between the treatment groups and time were also minimized. An ANCOVA model was applied to see the mean difference of gadd34, xbp1spl and HBD1 among different treatment groups. In both cases of GEE and ANCOVA, a p-value of < 0.05 was considered significant. All the data were analyzed by using the Statistical package for the Social Science (SPSS) for Windows (version 20; Armonk, NY: IBM SPSS corp.; 2011) and Stata/IC, version13 (Stata Corp, Texas, USA).
Results
Patients and clinical scores
Of total 249 patients, samples from 127 patients were included in the present study where mean age of the patients was 27.25 ± 8.37 years and female male ratio was 1:1.86 (35:65). The patient groups in the present study did not differ significantly in demographic and baseline characteristics amongst themselves (Table 1) or from the primary patient cohort who were not included in the present study (Additional file 1: Table S1).
Table 1.
Demographic characteristics of the study participants
| Characteristics | Placebo (n = 32) | PBA (n = 32) | vitD3 (n = 31) | PBA + vitD3 (n = 32) |
|---|---|---|---|---|
| Age, years | 27.38 ± 8.65 | 27.72 ± 7.25 | 25.42 ± 8.15 | 27.16 ± 8.38 |
| Males | 20 (62.50%) | 22 (25.90%) | 21 (67.70%) | 22 (68.80%) |
| History of contacts | 10 (31.30%) | 11 (34.40%) | 6 (19.40%) | 8 (25.00%) |
| BCG given | 22 (68.8%) | 19 (59.4%) | 26 (83.90%) | 23 (71.90%) |
| Weight, kg | 44.75 ± 8.20 | 45.74 ± 8.91 | 42.91 ± 8.70 | 44.00 ± 7.89 |
| Duration of illness, days | 48.91 ± 25.42 | 50.63 ± 21.80 | 58.39 ± 26.81 | 52.47 ± 27.58 |
| Vitamin D status | ||||
| Deficient, < 30 nmol/L | 18 (56%) | 23 (71%) | 21 (67%) | 16 (50%) |
| Insufficient, 30–50 nmol/L | 10 (31.30%) | 6 (18.8%) | 5 (16.10%) | 12 (37.50%) |
| Sufficient, > 50 nmol/L | 4 (12.50%) | 3 (9.4%) | 5 (16.10%) | 4 (12.50%) |
| Baseline clinical score (25) | 6.47 ± 0.86 | 6.45 ± 1.09 | 6.27 ± 1.01 | 6.39 ± 0.96 |
| Baseline sputum smear | ||||
| < 3 acid-fast bacilli | 17 (43.10%) | 17 (43.10%) | 16 (51.60%) | 18 (56.20%) |
| > 3 acid-fast bacilli | 15 (46.90%) | 15 (46.90%) | 15 (48.40%) | 14 (43.80%) |
Data is presented as mean ± standard deviation or number with percentage in parentheses
Abbreviations: BCG Bacillus Calmette–Guérin
Longitudinal change in TB scores as analyzed by GEE showed a significant decline over time (week 0, 4, 8 and 12) in the PBA group compared to placebo (Additional file 1: Table S2) in the present cohort (n = 127). A similar reduction in TB scores was obtained in the PBA group compared to placebo in the parent cohort of 249 patients [25]. Significantly higher percentages of patients in the PBA + vitD3 and vitD3 groups became sputum culture negative at week 4 compared to placebo though no difference was obtained with sputum smear conversion.
Cytokines and chemokines
Concentrations of cytokines IL-2, IL-5, IL-13 and IL-15 in PBMC supernatants were low or below detection limit in most patients. The other cytokines and chemokines showed a wide range of concentrations in all the supernatants measured. Concentrations of TNF-α, IL-17 (p = 0.05) and CCL11 (eotaxin) (p = 0.01) declined significantly from week 0 to week 8 in the PBA group. In the PBA + vitD3 group, FGF-basic and PDGF-β reduced in concentrations after 8 weeks from week 0 (p = 0.01) (Additional file 1: Table S3). Other groups did not show any significant reduction in cytokine/chemokine concentrations with time.
Analysis by GEE method exhibited a significant decline in TNF-α (β = − 0.34, 95% CI = − 0.68, − 0.003, p = 0.04) concentrations from week 0 to week-8 in the PBA group compared to placebo (Table 2). A significant reduction in the concentrations of the chemokines CCL11 (β = − 0.19, 95% CI = − 0.36, − 0.03, p = 0.02) and CCL5 (RANTES) (β = − 0.08, 95% CI = − 0.16, 0.002, p = 0.05) was observed in the PBA group, while CCL11 (β = − 0.17, 95% CI = − 0.34, − 0.001, p = 0.04), CXCL10 (IP-10) (β = − 0.38, 95% CI = − 0.77, 0.003, p = 0.05) and PDGF-β (β = − 0.16, 95% CI = − 0.31, 0.002, p = 0.05) declined significantly in the vitD3 group (Table 2). However, no differences in chemokine concentrations were noted in the PBA + vitD3 group compared to placebo. The results demonstrated that 8 weeks’ adjunct therapy with oral PBA or vitD3 effectively decreased expression of mononuclear cell-derived inflammatory cytokines and chemokines thereby reflecting reduced inflammatory responses in TB patients.
Table 2.
Longitudinal change (week 0 and 8) in inflammatory cytokines in treatment groups compared to placebo
| Crude | Adjusteda | |||
|---|---|---|---|---|
| β(95% CI) | p-value | β(95% CI) | p-value | |
| TNF-α | ||||
| PBA (n = 32) | − 0.17 (− 0.42, 0.09) | 0.20 | − 0.34 (− 0.68, − 0.003) | 0.04 |
| vitD3 (n = 31) | − 0.16 (− 0.41, 0.10) | 0.23 | − 0.22 (− 0.57, 0.13) | 0.22 |
| PBA + vitD3 (n = 32) | −0.13 (− 0.38, 0.13) | 0.32 | − 0.03 (− 0.37, 0.31) | 0.85 |
| IL-8 | ||||
| PBA (n = 32) | −0.29 (− 0.68, 0.09) | 0.13 | − 0.33 (− 0.71, 0.05) | 0.08 |
| vitD3 (n = 31) | − 0.11 (− 0.50, 0.28) | 0.57 | −0.09 (− 0.49, 0.30) | 0.63 |
| PBA + vitD3 (n = 32) | −0.17 (− 0.55, 0.22) | 0.39 | −0.18 (− 0.55, 0.20) | 0.36 |
| GM-CSF | ||||
| PBA (n = 32) | −0.13 (− 0.38, 0.11) | 0.28 | − 0.27 (− 0.56, 0.01) | 0.06 |
| vitD3 (n = 31) | − 0.19 (− 0.44, 0.05) | 0.12 | −0.19 (− 0.49, 0.10) | 0.20 |
| PBA + vitD3 (n = 32) | 0.08 (−0.17, 0.32) | 0.54 | 0.06 (−0.23, 0.35) | 0.69 |
| IL-4 | ||||
| PBA (n = 32) | −0.10 (− 0.23, 0.03) | 0.14 | − 0.14 (− 0.31, 0.01) | 0.08 |
| vitD3 (n = 31) | − 0.08 (− 0.21, 0.06) | 0.26 | −0.11 (− 0.27, 0.06) | 0.20 |
| PBA + vitD3 (n = 32) | −0.06 (− 0.20, 0.07) | 0.35 | −0.05 (− 0.22, 0.11) | 0.52 |
| CCL11 (eotaxin) | ||||
| PBA (n = 20) | −0.03 (− 0.17, 0.10) | 0.63 | − 0.19 (− 0.36, − 0.03) | 0.02 |
| vitD3 (n = 17) | −0.11 (− 0.26, 0.03) | 0.11 | −0.17 (− 0.34, − 0.001) | 0.04 |
| PBA + vitD3 (n = 19) | −0.04 (− 0.18, 0.10) | 0.59 | −0.07 (− 0.23, 0.10) | 0.44 |
| CCL5 (RANTES) | ||||
| PBA (n = 20) | −0.07 (− 0.13, − 0.001) | 0.04 | −0.08 (− 0.16, 0.002) | 0.05 |
| vitD3 (n = 17) | −0.05 (− 0.12, 0.01) | 0.11 | −0.06 (− 0.15, 0.02) | 0.14 |
| PBA + vitD3 (n = 19) | −0.04 (− 0.10, 0.03) | 0.27 | −0.06 (− 0.14, 0.03) | 0.18 |
| CXCL10 (IP-10) | ||||
| PBA (n = 20) | −0.11 (− 0.41, 0.18) | 0.45 | − 0.13 (− 0.51, 0.24) | 0.47 |
| vitD3 (n = 17) | − 0.25 (− 0.57, 0.06) | 0.10 | −0.38 (− 0.77, 0.003) | 0.05 |
| PBA + vitD3 (n = 19) | −0.15 (− 0.46, 0.15) | 0.32 | −0.16 (− 0.54, 0.21) | 0.39 |
| PDGF-BB | ||||
| PBA (n = 20) | −0.03 (− 0.18, 0.12) | 0.71 | 0.01 (− 0.15, 0.16) | 0.92 |
| vitD3 (n = 17) | −0.16 (− 0.32, − 0.003) | 0.04 | −0.16 (− 0.31, 0.002) | 0.05 |
| PBA + vitD3 (n = 19) | −0.12 (− 0.28, 0.03) | 0.10 | −0.12 (− 0.27, 0.03) | 0.12 |
Data is presented as beta coefficient with 95% confidence intervals in parentheses
Abbreviations: CI confidence interval, PBA phenylbutyrate, vitD3 vitamin D3, TNF-α tumor necrosis factor-alpha, IL interleukin, GM-CSF granulocyte-macrophage-colony stimulating factor, PDGF-BB platelet-derived growth factor-BB
aAdjusted for age, sex, duration of treatment, history of contact with active TB cases.
Statistical analysis was performed using generalized estimating equation (GEE) model. P-value of < 0.05 is significant
Autophagy marker in ex vivo infected macrophages
Confocal microscopic analysis revealed the activation of autophagy process in macrophages measured by the presence of LC3 positive puncta in MDM. LC3 puncta structures in ex vivo infected MDM showed highest percentage of cells with LC3-positive puncta in PBA + vitD3 group at week 8 (Fig. 2a).
Fig. 2.

a Expression of LC3+ macrophages in TB patients after adjunct therapy with PBA and/or vitD3 and simultaneously under treatment with anti-TB drugs. MDMs derived from the patients were infected with the virulent strain of Mycobacterium tuberculosis H37Rv for 2 h. The cells were fixed and stained with DAPI to visualize the nuclei (blue), and with anti-LC3, followed by the addition of Alexafluor 488-conjugated goat anti-mouse IgG (green color). One representative immunofluorescence image out of 6 independent replicates are shown; scale bars: 10 μm. b Percentage of LC3 expressing cells among uninfected and Mtb-infected monocyte-derived-macrophages (MDM) in the four treatment arms
There was a basal number of LC3 expressing cells (4.3 ± 2.4) among the uninfected MDM in all the treatment arms of patients that did not change with different therapies or over the duration of the study period (Fig. 2b). In all groups a gradual increase in percentage of LC3-expressing H37Rv-infected MDM from week 0 up to week 12 was noted, indicating that the capacity of MDM to undergo autophagy increased with time during clinical recovery (Fig. 2a and b). Analysis by GEE model showed that mean changes of LC3-expressing macrophage percentage in PBA (β = 8.26, 95% CI = 3.61, 12.9, p < 0.001), vitD3 (β = 4.73, 95% CI = 0.28, 9.19, p = 0.037) and PBA+ vitD3 (β = 10.47, 95% CI = 5.33, 15.6, p < 0.001) were significantly higher compared to placebo (Fig. 3). Hence, oral adjunct therapy with PBA, vitD3 and the combined dose had a positive impact on priming for induction of autophagy in ex vivo-infected macrophages from TB patients that persisted up to additional 4 weeks after the adjunct therapy was completed.
Fig. 3.
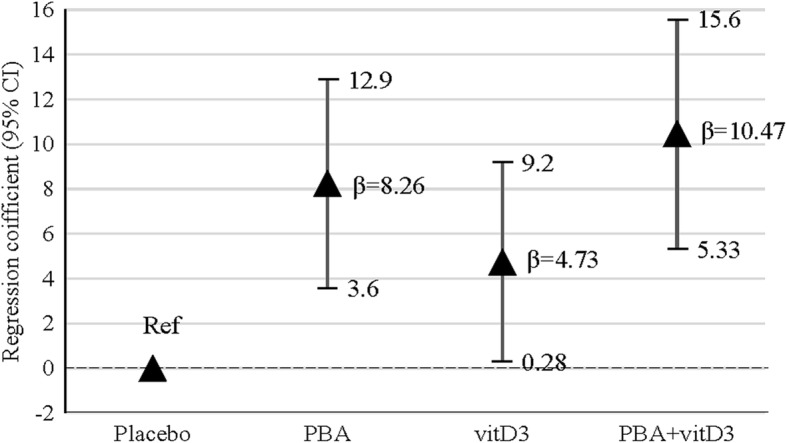
Generalized estimating equation (GEE) model was used to estimate the effect of adjunct therapy on LC3-expressing macrophages over time (week 0, 4, 8 and 12). β coefficient values are shown for three intervention groups (PBA, vitD3 and PBA + vitD3) compared to placebo. Arrowhead points represent the adjusted beta (β) coefficient values and vertical lines define 95% confidence intervals (CI). The analysis showed that mean % changes of LC3-expressing macrophage in PBA (β = 8.26; p < 0.001), vitD3 (β = 4.73; p = 0.037) and PBA+ vitD3 (β = 10.47; p < 0.001) were significantly higher compared to the placebo group
Endoplasmic reticulum stress markers and HBD1 in macrophages
When MDM from TB patients were examined for ER stress genes, there were few time points where gene expression was below detection limit for GADD34 or XBP1spl. These were considered as missing values in ANCOVA. All groups revealed a decline in expression of GADD34 transcripts from week 0 to week 8. However, sex and age-adjusted ANCOVA analysis did not reveal any significant difference in the expression of GADD34 transcripts between the treatment groups and placebo (Fig. 4a). Interestingly, a significant reduction in the expression of XBP1spl transcripts was found at week 8 in PBA (p = 0.036) and vitD3 (p = 0.008) groups compared to placebo but not in PBA + vitD3 (p = 0.12) group (Fig. 4b). Both PBA and vitamin D3 are known for their role in reducing ER stress in cells (26 or 27) [30]. Host directed therapy with PBA or vitamin D3 could individually dampen the ER stress in MDM, although the combined intervention did not reach a statistically significant level.
Fig. 4.
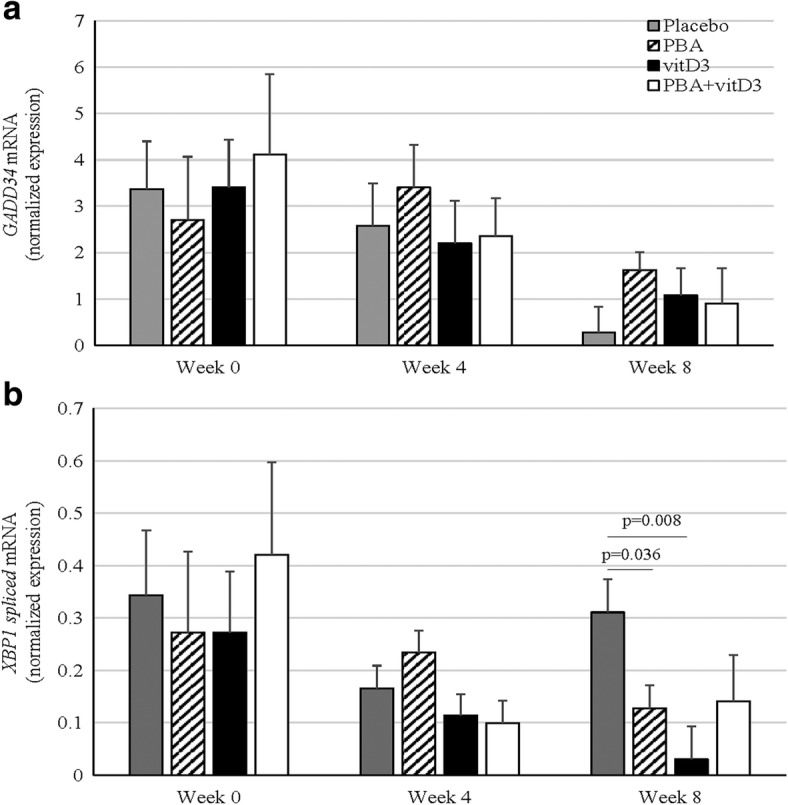
Real-time RT-PCR assays of GADD34 and XBP1spl mRNAs from monocyte-derived-macrophages from TB patients receiving adjunct therapy with PBA and/or vitD3 or placebo (18/group) at week 0, week 4 and 8 after therapy. Data are normalized expression of GAD34 (a) and XBP1spl (b) mRNAs presented as means ± standard deviation. The results showed that PBA and vitD3 groups showed a significant reduction in expression of ER stress gene spliced XBP1 mRNA at week 8 compared to placebo
In addition to studying expression of cathelicidin LL-37 in MDM in the original study [25], we further assessed effect of PBA and/or vitD3 on induction of HBD1 in MDMs. The expression of HBD1 transcripts (2–ΔΔCT) in different intervention groups did not change significantly with time. Furthermore, the fold change in HBD1 transcripts did not exhibit any difference over time in any of the intervention groups compared to placebo (Table 3).
Table 3.
Longitudinal change (week 0, 4, 8) in HBD1 in the treatment groups compared to Placebo (n = 18)
| Crude | Adjusteda | |||
|---|---|---|---|---|
| HBD1 | β(95% CI) | p-value | β(95% CI) | p-value |
| PBA (n = 18) | 0.26 (−0.10, 0.62) | 0.15 | 0.31 (−0.06, 0.67) | 0.10 |
| vitD3 (n = 18) | −0.05 (− 0.40, 0.29) | 0.76 | 0.04 (− 0.31, 0.40) | 0.81 |
| PBA + vitD3 (n = 18) | 0.21 (−0.15, 0.56) | 0.25 | 0.28 (−0.09, 0.66) | 0.14 |
Data is presented as beta coefficient with 95% confidence intervals in parentheses
Abbreviations: CI confidence interval, HBD1 human beta-defenisn-1
aAdjusted for age, sex, duration of treatment, history of contact with active TB cases, time and the interaction between the treatment groups and time.
Statistical analysis was performed using generalized estimating equation (GEE) model. P-value of < 0.05 is significant
Discussion
It is important to evaluate the role of immune-modulatory agents as host directed therapy in the treatment of TB that can improve efficacy of therapy, reduce or prevent detrimental outcomes of toxic drugs and improve patient compliance. Our interest in using PBA and/or vitD3 as HDT in the treatment of TB was based on their diverse roles in modifying the host defense system. Our findings showed that adjunctive treatment with PBA or vitD3 over 8 weeks’ time reduced concentration of cytokines/chemokines and endoplasmic reticulum stress in parallel to clinical recovery. All three adjunctive therapies increased frequency of autophagy in macrophages compared to placebo.
TB is a chronic infectious disease with a spectrum of symptoms and presentations, wherein cytokines and chemokines play important roles in manipulation of the immune responses, containment of Mtb, pathogenesis and disease manifestations. Various studies have compared serum cytokine/chemokine profiles of patients with different clinical presentations of TB, to establish the expression levels as early surrogate biomarkers of therapeutic responses, bacteriological confirmation of TB and sputum culture conversion [31–33]. We measured cytokines and chemokines in culture supernatants of PBMC instead of serum or plasma, and the findings were similar to previous studies, showing marked decline of serum cytokine/chemokine concentrations at different treatment stages [14, 33–37]. Particularly we found that compared to conventional anti-TB treatment in the placebo group, cytokine/chemokine reduction was greater in the PBA or vitD3 groups that occurred in parallel to clinical recovery [25] (reduced TB score), suggesting a role of PBA and vitD3 in dampening of inflammatory responses.
Mycobacterial infection has been shown to induce ER stress and damage ER. Cells deal with ER stress by activation of the unfolded protein response (UPR), but chronic activation of these pathways can eventually result in apoptosis and lung injury [18, 38–40]. Several biomarkers of ER stress have been used in various studies by assessing activation of the UPR; spliced X-box binding protein-1 (XBP1spl) mRNA has been shown to be a reliable marker in revealing dose-dependent changes in the transcript level of this gene [28]. Mycobacterial secreted antigens such as ESAT-6 has been shown to increase XBP1spl as well as other ER stress genes in epithelial cell lines [38]. Toll-like receptor activation was also shown to increase splicing of the transcription factor XBP1 in the absence of other signs of activation of the UPR, and this was found to regulate innate immune responses in macrophages [41]. PBA is a classic ER stress inhibitor; it is a chemical chaperone that improves ER folding capacity and trafficking of mutant proteins out of the ER, stabilization of protein conformation and thereby reduces accumulation of misfolded proteins in ER lumen. In cell lines, experimental models and tissue samples, PBA has been shown to reduce expression of GRP-78, CHOP and other UPR-related markers such as ATF6, XBP1 and phosphorylation of eIF2α [27, 42–44]. Vitamin D is also considered a natural ER stress reliever [30, 45]. Down-regulation of GADD34 transcripts after 8 weeks of therapy was evident in all 4 arms, however down-regulation of spliced XBP1 was evident only in PBA and vitD3 arms compared to placebo. Thus, in vivo effect of PBA and vitD3 adjunct treatments was demonstrated in diminishing ER stress in MDM during TB infection that may have a role in improving clinical outcome. Moreover, diminished inflammatory cytokine responses as we have shown here may be linked to suppression of ER stress [46, 47].
Autophagy is an important physiological process that is applied by macrophages to control and eliminate intracellular pathogens. We have earlier demonstrated in a mechanistic study that PBA, active vitD3 separately and in combination induced autophagy in macrophages from healthy donors when infected in vitro with Mtb [23], increased the intracellular killing of Mtb compared to untreated MDM and induced LL-37 in cell supernatants of macrophages and lymphocytes after oral ingestion by healthy adults [22]. The novel activity of PBA as an inducer of autophagy was LL-37-dependent. We further showed in the present study that, all three adjunctive therapies (PBA, vitD3 and PBA + vitD3) in the clinical trial [25] promoted autophagy in ex vivo infected macrophages from the patients, indicating that the inducers can prime the macrophages for autophagy response. Moreover, the autophagy process occurred in parallel to induction of LL-37 in macrophage/PBMC [25], continuing up to an additional week after the adjunct therapy was completed. MDM-mediated elimination of Mtb in the vitamin D and PBA + vitamin D3 arms in the trial thus strongly suggest a role of the autophagy process in the eradication. Autophagy also plays a housekeeping role in removing misfolded, unfolded or aggregated proteins and clearing damaged organelles, such as ER and mitochondria. In view of the involvement of the UPR in activating autophagy, it is interesting to note that whereas treatment increased autophagy, it appeared to decrease activation of the UPR to ER stress. This suggests that mechanisms distinct from the UPR are involved in the observed treatment-induced autophagy.
Human beta defensin-1 (HBD1) is produced constitutively by all human epithelia and some immune cells. Monocyte-derived-macrophages and -dendritic cells both express HBD1 mRNA, showing increased expression after activation with IFN-γ and/or lipopolysaccharide in a dose- and time-dependent fashion [48]. When expression of HBD1 transcripts was studied in MDMs from the patients we did not observe any increase or down-regulation of the peptide in the treatment groups compared to placebo during the course of TB disease. Thus, PBA or vitD3 did not seem to have any modulating effect on HBD1 expression in peripheral MDM as seen with LL-37 [25].
One of the limitations of this study was the small size used per treatment group for evaluation of ER stress genes and HBD1 genes. The expression of transcripts was studied only in macrophages, not other cells in PBMC. This was done because of unavailability of adequate number of matched/paired samples from different time points and suboptimum concentrations of specimens remaining from the clinical trial. Our analysis of activation of the UPR was restricted to measurement of GADD34 and XBP1spl because of limitations in the availability of RNA, and therefore other UPR markers (including protein markers) should be assessed in future studies. Only one autophagy marker LC3 was used instead of a combination of markers such as ATG 5 and Beclin-1; however, our earlier studies showed the LC3 marker as reliable for assessment of autophagy in macrophages [23]. The combined adjunct therapy of PBA and vitD3 did not exhibit any effect on cytokines/chemokines or ER stress reduction in TB patients and require further investigations. Notably, the combined treatment with PBA and vitD3 of the patients exhibited the fastest sputum culture conversion [25]. Another limitation was that we measured cytokines and chemokines in saponin-treated cell culture fluid instead of serum or plasma as traditionally done [10–14, 31–33]. Measurement of cytokines in the serum does not accurately reflect active secretion of cytokines due to an ongoing infection; moreover it is affected by clearance from the circulation. Our hypothesis is that the PBMC from TB patients are in vivo activated and they do not require ex-vivo stimulation; release of cytokines/chemokines from unstimulated cells into the culture supernatant indicate active secretion and thus are more relevant to the ongoing disease process. However, use of saponin allowed excretion of intra-cellular cytokines into the culture supernatant which was not a spontaneous release of cytokines.
Conclusions
demonstration of clinically beneficial immunomodulation that occur in parallel to improved treatment outcomes suggest that use of repurposed agents such as Phenylbutyrate and vitamin D3 can be a valuable strategy in HDT against drug-sensitive TB. The toxicities and poor treatment outcomes of current therapies against MDR-TB necessitate newer approaches to improve the management and control of MDR-TB. Development of host-directed therapies integrated with Mtb-targeted chemotherapy provides a complicated challenge because temporal events of infection and host immunity may play a critical role in determining HDT efficacy. Thus, the therapeutic potential of PBA and vitD3 HDT against multidrug resistant TB warrants urgent clinical evaluation in well-designed multi-center clinical trials in endemic settings.
Additional file
Table S1. Descriptive statistics of the current studied patients and the patients who were not studied. Table S2. Longitudinal change (week 0, 4, 8 and 12) in TB score in the intervention groups compared to placebo. Table S3. Concentrations of inflammatory cytokines and chemokines in treatment groups at different time intervals. (DOCX 24 kb)
Acknowledgements
We thank Dr. Mami Taniuchi, Dr. Rashidul Haque and Dr. Mustafizur Rahman for accommodating multiplex cytokine analysis.
Funding
This study was supported by the International Centre for Diarrheal Disease Research, Bangladesh (icddr,b), Sida (Sida-icddrb Agreement support; Grant 384, SWE-2008-065) and Swedish Strategic Foundation (SSF, Grant No. RBd08–0014), the Swedish Heart-Lung Foundation (Grant No. 2013–0366) and Swedish Research Council (Grant No. 2016–01496). No funding bodies had any role in the design of the study, collection, analysis, and interpretation of data and in writing the manuscript.
Availability of data and materials
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request. For additional information please refer to http://www.icddrb.org/policies.
Abbreviations
- 95% CI
95% confidence interval
- AMPs
Antimicrobial peptides
- DOTS
Directly observed therapy short-course
- ER
Endoplasmic-reticulum
- GADD34
Growth arrest and DNA damage inducible protein 34
- GEE
Generalized estimating equation
- HBD1
Human β-defensin-1
- HDT
Host-directed therapies
- MDM
Monocyte-derived-macrophages
- Mtb
Mycobacterium tuberculosis
- PBA
Phenylbutyrate
- PBMC
Peripheral blood mononuclear cells
- TB
Tuberculosis
- vitD3
Vitamin D3
- XBP1spl
Spliced X-box binding protein-1
Authors’ contributions
Conceived and designed the experiments: RR, BA and GHG. Patient sample collection, microbiology and acquisition of data: SMMK. Performed the experiment: RSR, AM, TS, SA and AVS. Data entry, statistical analysis of the data, prepared figures and tables: AH. Data interpretation: RR, BA, GHG, PSH and AH. Contributed reagents/materials/analysis tools: RR, BA, GHG and PSH. Drafting the article for important content: RR, BA, RSR, AM, SMMK and PSH. Approval of final version: RR, RSR, AM, BA, GHG, AVS, PSH, AH, SA, TS and SMMK. All authors have read and approved the final version of the manuscript.
Ethics approval and consent to participate
The study was approved by the Research and Ethical Review Committees at the icddr,b Dhaka, Bangladesh. Written informed consent was obtained from patients before enrollment into the study that included permission to store specimens for future use.
Consent for publication
Not applicable.
Competing interests
All authors: No reported conflicts of interest. All authors have submitted the ICMJE Form for Disclosure of Potential Conflicts of Interest. Conflicts that the editors consider relevant to the content of the manuscript have been disclosed.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Footnotes
Rokeya Sultana Rekha and Akhirunnesa Mily contributed equally to this work.
Birgitta Agerberth and Rubhana Raqib contributed equally to this work as senior authors.
Electronic supplementary material
The online version of this article (10.1186/s12879-018-3203-9) contains supplementary material, which is available to authorized users.
Contributor Information
Rokeya Sultana Rekha, Email: rokeya.sultana.rekha@ki.se.
Akhirunnesa Mily, Email: mily@icddrb.org.
Tajnin Sultana, Email: tajnin.sultana@icddrb.org.
Ahsanul Haq, Email: shohag@icddrb.org.
Sultan Ahmed, Email: sultanahmed@icddrb.org.
S. M. Mostafa Kamal, Email: ntrlnidchbd@yahoo.com
Annemarie van Schadewijk, Email: A.van_Schadewijk@lumc.nl.
Pieter S. Hiemstra, Email: P.S.Hiemstra@lumc.nl
Gudmundur H. Gudmundsson, Email: ghrafn@hi.is
Birgitta Agerberth, Email: Birgitta.Agerberth@ki.se.
Rubhana Raqib, Phone: +8802-9827068, Email: rubhana@icddrb.org.
References
- 1.Organization WH . Report. 2016. Global Tuberculosis Report 2016. [Google Scholar]
- 2.Casarini M, Ameglio F, Alemanno L, Zangrilli P, Mattia P, Paone G, Bisetti A, Giosue S. Cytokine levels correlate with a radiologic score in active pulmonary tuberculosis. Am J Respir Crit Care Med. 1999;159(1):143–148. doi: 10.1164/ajrccm.159.1.9803066. [DOI] [PubMed] [Google Scholar]
- 3.Chowdhury IH, Ahmed AM, Choudhuri S, Sen A, Hazra A, Pal NK, Bhattacharya B, Bahar B. Alteration of serum inflammatory cytokines in active pulmonary tuberculosis following anti-tuberculosis drug therapy. Mol Immunol. 2014;62(1):159–168. doi: 10.1016/j.molimm.2014.06.002. [DOI] [PubMed] [Google Scholar]
- 4.Ravimohan S, Tamuhla N, Steenhoff AP, Letlhogile R, Nfanyana K, Bellamy SL, MacGregor RR, Gross R, Weissman D, Bisson GP. Immunological profiling of tuberculosis-associated immune reconstitution inflammatory syndrome and non-immune reconstitution inflammatory syndrome death in HIV-infected adults with pulmonary tuberculosis starting antiretroviral therapy: a prospective observational cohort study. Lancet Infect Dis. 2015;15(4):429–438. doi: 10.1016/S1473-3099(15)70008-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ahmed RK, Rohava Z, Balaji KN, Hoffner SE, Gaines H, Magalhaes I, Zumla A, Skrahina A, Maeurer MJ. Pattern recognition and cellular immune responses to novel Mycobacterium tuberculosis-antigens in individuals from Belarus. BMC Infect Dis. 2012;12:41. doi: 10.1186/1471-2334-12-41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dorhoi A, Reece ST, Kaufmann SH. For better or for worse: the immune response against Mycobacterium tuberculosis balances pathology and protection. Immunol Rev. 2011;240(1):235–251. doi: 10.1111/j.1600-065X.2010.00994.x. [DOI] [PubMed] [Google Scholar]
- 7.Kaufmann SH, Lange C, Rao M, Balaji KN, Lotze M, Schito M, Zumla AI, Maeurer M. Progress in tuberculosis vaccine development and host-directed therapies--a state of the art review. Lancet Respir Med. 2014;2(4):301–320. doi: 10.1016/S2213-2600(14)70033-5. [DOI] [PubMed] [Google Scholar]
- 8.Korbel DS, Schneider BE, Schaible UE. Innate immunity in tuberculosis: myths and truth. Microbes Infect. 2008;10(9):995–1004. doi: 10.1016/j.micinf.2008.07.039. [DOI] [PubMed] [Google Scholar]
- 9.O'Garra A, Redford PS, McNab FW, Bloom CI, Wilkinson RJ, Berry MP. The immune response in tuberculosis. Annu Rev Immunol. 2013;31:475–527. doi: 10.1146/annurev-immunol-032712-095939. [DOI] [PubMed] [Google Scholar]
- 10.Dhanasekaran S, Jenum S, Stavrum R, Ritz C, Faurholt-Jepsen D, Kenneth J, Vaz M, Grewal HM, Doherty TM, Group TBTS Identification of biomarkers for Mycobacterium tuberculosis infection and disease in BCG-vaccinated young children in southern India. Genes Immun. 2013;14(6):356–364. doi: 10.1038/gene.2013.26. [DOI] [PubMed] [Google Scholar]
- 11.John SH, Kenneth J, Gandhe AS. Host biomarkers of clinical relevance in tuberculosis: review of gene and protein expression studies. Biomarkers. 2012;17(1):1–8. doi: 10.3109/1354750X.2011.628048. [DOI] [PubMed] [Google Scholar]
- 12.Lalor MK, Floyd S, Gorak-Stolinska P, Ben-Smith A, Weir RE, Smith SG, Newport MJ, Blitz R, Mvula H, Branson K, et al. BCG vaccination induces different cytokine profiles following infant BCG vaccination in the UK and Malawi. J Infect Dis. 2011;204(7):1075–1085. doi: 10.1093/infdis/jir515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mihret A, Bekele Y, Bobosha K, Kidd M, Aseffa A, Howe R, Walzl G. Plasma cytokines and chemokines differentiate between active disease and non-active tuberculosis infection. J Inf Secur. 2013;66(4):357–365. doi: 10.1016/j.jinf.2012.11.005. [DOI] [PubMed] [Google Scholar]
- 14.Xiong W, Dong H, Wang J, Zou X, Wen Q, Luo W, Liu S, He J, Cai S, Ma L. Analysis of plasma cytokine and chemokine profiles in patients with and without tuberculosis by liquid Array-based multiplexed immunoassays. PLoS One. 2016;11(2):e0148885. doi: 10.1371/journal.pone.0148885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gutierrez MG, Master SS, Singh SB, Taylor GA, Colombo MI, Deretic V. Autophagy is a defense mechanism inhibiting BCG and Mycobacterium tuberculosis survival in infected macrophages. Cell. 2004;119(6):753–766. doi: 10.1016/j.cell.2004.11.038. [DOI] [PubMed] [Google Scholar]
- 16.Deretic V, Singh S, Master S, Harris J, Roberts E, Kyei G, Davis A, de Haro S, Naylor J, Lee HH, et al. Mycobacterium tuberculosis inhibition of phagolysosome biogenesis and autophagy as a host defence mechanism. Cell Microbiol. 2006;8(5):719–727. doi: 10.1111/j.1462-5822.2006.00705.x. [DOI] [PubMed] [Google Scholar]
- 17.Kumar D, Rao KV. Regulation between survival, persistence, and elimination of intracellular mycobacteria: a nested equilibrium of delicate balances. Microbes Infect. 2011;13(2):121–133. doi: 10.1016/j.micinf.2010.10.009. [DOI] [PubMed] [Google Scholar]
- 18.Cui Y, Zhao D, Barrow PA, Zhou X. The endoplasmic reticulum stress response: a link with tuberculosis? Tuberculosis (Edinb) 2016;97:52–56. doi: 10.1016/j.tube.2015.12.009. [DOI] [PubMed] [Google Scholar]
- 19.O'Connor G, Gleeson LE, Fagan-Murphy A, Cryan SA, O'Sullivan MP, Keane J. Sharpening nature's tools for efficient tuberculosis control: a review of the potential role and development of host-directed therapies and strategies for targeted respiratory delivery. Adv Drug Deliv Rev. 2016;102:33–54. doi: 10.1016/j.addr.2016.04.024. [DOI] [PubMed] [Google Scholar]
- 20.Zumla A, Rao M, Dodoo E, Maeurer M. Potential of immunomodulatory agents as adjunct host-directed therapies for multidrug-resistant tuberculosis. BMC Med. 2016;14:89. doi: 10.1186/s12916-016-0635-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Batshaw ML, MacArthur RB, Tuchman M. Alternative pathway therapy for urea cycle disorders: twenty years later. J Pediatr. 2001;138(1 Suppl):S46–S54. doi: 10.1067/mpd.2001.111836. [DOI] [PubMed] [Google Scholar]
- 22.Mily A, Rekha RS, Kamal SM, Akhtar E, Sarker P, Rahim Z, Gudmundsson GH, Agerberth B, Raqib R. Oral intake of phenylbutyrate with or without vitamin D3 upregulates the cathelicidin LL-37 in human macrophages: a dose finding study for treatment of tuberculosis. BMC Pulm Med. 2013;13:23. doi: 10.1186/1471-2466-13-23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Rekha RS, Rao Muvva SS, Wan M, Raqib R, Bergman P, Brighenti S, Gudmundsson GH, Agerberth B. Phenylbutyrate induces LL-37-dependent autophagy and intracellular killing of Mycobacterium tuberculosis in human macrophages. Autophagy. 2015;11(9):1688–1699. doi: 10.1080/15548627.2015.1075110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Steinmann J, Halldorsson S, Agerberth B, Gudmundsson GH. Phenylbutyrate induces antimicrobial peptide expression. Antimicrob Agents Chemother. 2009;53(12):5127–5133. doi: 10.1128/AAC.00818-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Mily A, Rekha RS, Kamal SM, Arifuzzaman AS, Rahim Z, Khan L, Haq MA, Zaman K, Bergman P, Brighenti S, et al. Significant effects of oral Phenylbutyrate and vitamin D3 adjunctive therapy in pulmonary tuberculosis: a randomized controlled trial. PLoS One. 2015;10(9):e0138340. doi: 10.1371/journal.pone.0138340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Jellbauer S, Perez Lopez A, Behnsen J, Gao N, Nguyen T, Murphy C, Edwards RA, Raffatellu M. Beneficial effects of sodium Phenylbutyrate administration during infection with Salmonella enterica Serovar typhimurium. Infect Immun. 2016;84(9):2639–2652. doi: 10.1128/IAI.00132-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wang Z, Huang Y, Cheng Y, Tan Y, Wu F, Wu J, Shi H, Zhang H, Yu X, Gao H, et al. Endoplasmic reticulum stress-induced neuronal inflammatory response and apoptosis likely plays a key role in the development of diabetic encephalopathy. Oncotarget. 2016;7(48):78455–78472. doi: 10.18632/oncotarget.12925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.van Schadewijk A, van't Wout EF, Stolk J, Hiemstra PS. A quantitative method for detection of spliced X-box binding protein-1 (XBP1) mRNA as a measure of endoplasmic reticulum (ER) stress. Cell Stress Chaperones. 2012;17(2):275–279. doi: 10.1007/s12192-011-0306-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods. 2001;25(4):402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 30.Haas MJ, Jafri M, Wehmeier KR, Onstead-Haas LM, Mooradian AD. Inhibition of endoplasmic reticulum stress and oxidative stress by vitamin D in endothelial cells. Free Radic Biol Med. 2016;99:1–10. doi: 10.1016/j.freeradbiomed.2016.07.020. [DOI] [PubMed] [Google Scholar]
- 31.den Hertog AL, Montero-Martin M, Saunders RL, Blakiston M, Menting S, Sherchand JB, Lawson L, Oladimeji O, Abdurrahman ST, Cuevas LE, et al. Cytokine kinetics in the first week of tuberculosis therapy as a tool to confirm a clinical diagnosis and guide therapy. PLoS One. 2015;10(6):e0129552. doi: 10.1371/journal.pone.0129552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Jayakumar A, Vittinghoff E, Segal MR, MacKenzie WR, Johnson JL, Gitta P, Saukkonen J, Anderson J, Weiner M, Engle M, et al. Serum biomarkers of treatment response within a randomized clinical trial for pulmonary tuberculosis. Tuberculosis (Edinb) 2015;95(4):415–420. doi: 10.1016/j.tube.2015.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lui G, Wong CK, Ip M, Chu YJ, Yung IM, Cheung CS, Zheng L, Lam JS, Wong KT, Sin WW, et al. HMGB1/RAGE signaling and pro-inflammatory cytokine responses in non-HIV adults with active pulmonary tuberculosis. PLoS One. 2016;11(7):e0159132. doi: 10.1371/journal.pone.0159132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Chavez K, Ravindran R, Dehnad A, Khan IH. Gender biased immune-biomarkers in active tuberculosis and correlation of their profiles to efficacy of therapy. Tuberculosis (Edinb) 2016;99:17–24. doi: 10.1016/j.tube.2016.03.009. [DOI] [PubMed] [Google Scholar]
- 35.Iqbal NT, Hussain R, Shahid F, Dawood G. Association of plasma cytokines with radiological recovery in pulmonary tuberculosis patients. Int J Mycobacteriol. 2016;5(2):111–119. doi: 10.1016/j.ijmyco.2015.12.003. [DOI] [PubMed] [Google Scholar]
- 36.Riou C, Perez Peixoto B, Roberts L, Ronacher K, Walzl G, Manca C, Rustomjee R, Mthiyane T, Fallows D, Gray CM, et al. Effect of standard tuberculosis treatment on plasma cytokine levels in patients with active pulmonary tuberculosis. PLoS One. 2012;7(5):e36886. doi: 10.1371/journal.pone.0036886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Suzukawa M, Akashi S, Nagai H, Nagase H, Nakamura H, Matsui H, Hebisawa A, Ohta K. Combined analysis of IFN-gamma, IL-2, IL-5, IL-10, IL-1RA and MCP-1 in QFT supernatant is useful for distinguishing active tuberculosis from latent infection. PLoS One. 2016;11(4):e0152483. doi: 10.1371/journal.pone.0152483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Choi HH, Shin DM, Kang G, Kim KH, Park JB, Hur GM, Lee HM, Lim YJ, Park JK, Jo EK, et al. Endoplasmic reticulum stress response is involved in Mycobacterium tuberculosis protein ESAT-6-mediated apoptosis. FEBS Lett. 2010;584(11):2445–2454. doi: 10.1016/j.febslet.2010.04.050. [DOI] [PubMed] [Google Scholar]
- 39.Deng W, Yang W, Zeng J, Abdalla AE, Xie J. Mycobacterium tuberculosis PPE32 promotes cytokines production and host cell apoptosis through caspase cascade accompanying with enhanced ER stress response. Oncotarget. 2016;7(41):67347–67359. doi: 10.18632/oncotarget.12030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Lim YJ, Choi JA, Choi HH, Cho SN, Kim HJ, Jo EK, Park JK, Song CH. Endoplasmic reticulum stress pathway-mediated apoptosis in macrophages contributes to the survival of Mycobacterium tuberculosis. PLoS One. 2011;6(12):e28531. doi: 10.1371/journal.pone.0028531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Martinon F, Chen X, Lee AH, Glimcher LH. TLR activation of the transcription factor XBP1 regulates innate immune responses in macrophages. Nat Immunol. 2010;11(5):411–418. doi: 10.1038/ni.1857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kim HJ, Jeong JS, Kim SR, Park SY, Chae HJ, Lee YC. Inhibition of endoplasmic reticulum stress alleviates lipopolysaccharide-induced lung inflammation through modulation of NF-kappaB/HIF-1alpha signaling pathway. Sci Rep. 2013;3:1142. doi: 10.1038/srep01142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Placido AI, Pereira CM, Duarte AI, Candeias E, Correia SC, Santos RX, Carvalho C, Cardoso S, Oliveira CR, Moreira PI. The role of endoplasmic reticulum in amyloid precursor protein processing and trafficking: implications for Alzheimer's disease. Biochim Biophys Acta. 2014;1842(9):1444–1453. doi: 10.1016/j.bbadis.2014.05.003. [DOI] [PubMed] [Google Scholar]
- 44.Zeng M, Sang W, Chen S, Chen R, Zhang H, Xue F, Li Z, Liu Y, Gong Y, Zhang H, et al. 4-PBA inhibits LPS-induced inflammation through regulating ER stress and autophagy in acute lung injury models. Toxicol Lett. 2017;271:26–37. doi: 10.1016/j.toxlet.2017.02.023. [DOI] [PubMed] [Google Scholar]
- 45.Riek AE, Oh J, Sprague JE, Timpson A, de las Fuentes L, Bernal-Mizrachi L, Schechtman KB, Bernal-Mizrachi C. Vitamin D suppression of endoplasmic reticulum stress promotes an antiatherogenic monocyte/macrophage phenotype in type 2 diabetic patients. J Biol Chem. 2012;287(46):38482–38494. doi: 10.1074/jbc.M112.386912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Lawrence T. The nuclear factor NF-kB pathway in inflammation. Cold Spring Harb Perspect Biol. 2009;1(6):a001651. doi: 10.1101/cshperspect.a001651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Zhang K, Kaufman RJ. From endoplasmic-reticulum stress to the inflammatory response. Nature. 2008;454(7203):455–462. doi: 10.1038/nature07203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Duits LA, Ravensbergen B, Rademaker M, Hiemstra PS, Nibbering PH. Expression of beta-defensin 1 and 2 mRNA by human monocytes, macrophages and dendritic cells. Immunology. 2002;106(4):517–525. doi: 10.1046/j.1365-2567.2002.01430.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1. Descriptive statistics of the current studied patients and the patients who were not studied. Table S2. Longitudinal change (week 0, 4, 8 and 12) in TB score in the intervention groups compared to placebo. Table S3. Concentrations of inflammatory cytokines and chemokines in treatment groups at different time intervals. (DOCX 24 kb)
Data Availability Statement
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request. For additional information please refer to http://www.icddrb.org/policies.