

12.1 Background
The central nervous system (CNS) has many unique anatomic and physiologic properties that considerably affect manifestation of diseases within this complex organ.
12.1.1 Blood-brain-CSF barriers and the brain “privilege”
A question is posed whether our CNS, as the main controlling complex, is a particularly protected or “privileged” place?
A series of regulatory interfaces between the blood stream and the CNS parenchyma or CSF compartment, determine both the entry of needed metabolites and other compounds and the removal or exclusion of toxic or unnecessary metabolites and pathogens. They differ significantly by their anatomical, physiological, biochemical or even immunological features. However, each of them is characterized by the presence of tight junctions with extremely high electrical resistence between certain cell types; epithelial cells of choroid plexuses (“blood-CSF barrier”), or endothelial cells of brain capillaries (“blood-brain barrier”) (Fig. 1).
Figures 1a-c.
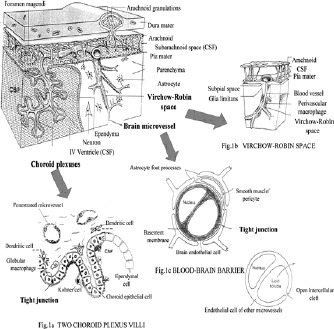
The major components of the blood-brain-CSF barriers and their relationship to brain parenchyma, CSF compartments (ventricle and subarachnoid space) and Virchow-Robin space.
From an immunological point of view, the most important consequences of the barriers are the restricted entry of immunocompetent cells and the low concentration of proteins, particularly antibodies and complement factors within the CNS. Thus, in health, the cells of the CNS, such as neurons, macroglia (astrocytes and oligodendrocytes) and microglia, function in an immunosuppressive environment which differs from that of other organs. The absence of organized lymphoid tissue reflects the fact that brain is not normally exposed to significant levels of antigenic stimulation.
Therefore, for many years the brain was regarded as a well-protected organ shielded from attack by invading organisms, but especially in normal condition, immunologically inert. Over the years this view has had to be modified.
12.1.2 The role of T lymphocytes in immune surveillance in health
The immune system is a surveillance mechanism that operates via cellular immunity and humoral immunity. The duality of these overlapping systems arise from cells called lymphocytes.
Although the intact blood-brain barrier constitutes a major barrier to humoral effector molecules such as autoantibodies and complement, it is less of a barrier to activated cells. It has been recently demonstrated in animal studies that the CNS tissue (like any non-lymphoid organ which could be inflamed) is routinelly patrolled by a subset of activated CD4+Th1 lymphocytes (“pioneer cells”) in the absence of an inflammatory focus to perform immune surveillance in normal condition. Such cells quickly disappeared from the tissue unless they encounter appropriate antigen within the CNS compartment. Thus, activated, but not naïve, lymphocytes can enter the CNS to perform immune surveillance under normal condition.
The findings that the lymphocyte subsets in normal (human) CSF differ from that in venous blood has been confirmed by experimental studies. The majority of T lymphocytes in CSF are memory cells (Table 1).
Table 1.
Lymphocyte subsets determined by flow cytometry in lumbar cerebrospinal fluid (CSF) and venous blood from control individuals (average values of reported results).
| Lymphocyte subsets (mean percentages) | Cerebrospinal fluid | Blood |
|---|---|---|
| CD3+ (T cells, total) | 90 | 70 |
| CD3+ HLA-DR+ (activated T cells) | 10 | 10 |
| CD4+ (helper inducer) | 65 | 45 |
| CD8+ (cytotoxic suppressor) | 25 | 30 |
| CD4+ CD8+ ratio | 2.5 | 1.5 |
| CD45RA (naive or virgin cells) | 35 | 65 |
| CD46RO (naive or virgin cells) | 65 | 35 |
| CD29 (memory cells) | 80 | 50 |
| CD16+ 56+ (NK cells) | 5 | 20 |
| CD19+ (B lymphocytes) | 2 | 15 |
Note that the majority of T lymphocytes in the cerebrospinal fluid are memory cells. Conversion of naive to memory T cells alters their surface molecule phenotype such as the change of CD45 molecule isoform RA to RO and the increased expression of numerous adhesion and activation molecules, e.g., CD29 (common β-subunit of the VLA integrin family).
12.1.3 Inflammation and blood-brain-CSF barriers
It has become apparent that the limited capacity of the brain to react depends upon the “integrity” of the barriers. Various inflammatory mediators increase vascular permeability of the barriers and allow effector immune cells, as well as humoral effector molecules (antibodies, complement) to enter the CNS compartment. Thus, in such conditions the effect of inflammation is to abrogate, if only temporarily, the CNS isolation from immune processes of the body.
In most cases the protein leak out of the small vessels is accompanied by an accomulation of inflammatory cells (e.g. bacterial infections). In many viral infections the vascular permeability changes are often transitory and disappear, but cells continue to enter from the circulation. However, the “break-down” in the barrier may be quite insufficient, with the result that immune response in the CNS may be restricted, delayed and ineffectual (e.g. in herpes simplex encephalitis). Further, serum proteins may be observed outside the vessel walls in the absence of inflammatory cells (e.g., Guillain-Barré syndrome, tumours) and conversely, vessels may be surrounded by cells and yet show no evidence of a protein leak (e.g. in same cases of encephalitis, early stage of multiple sclerosis).
The essential contributions of proinflamatory cytokines, chemokines, adhesion molecules, and proteases in the recruitment of inflammatory cells into the nervous tissue
The leukocyte attachment to the lamina surface of cerebral post capillary venula is now known to precede a whole cascade of molecular and morphologic events that culminate in the migration of immune cells (diapedesis) across endothelium into perivascular (Virchow-Robin) spaces were they accumulate, and some of them may move into CNS parenchyma and (or) CSF space. The entire process from arrival of “pioneer” cells to the appearence of overt clinical illnes is complex, with distinct adhesion molecules participing at different phases. All of them act via ligand-receptor binding and their expression is under regulatory control by various inflammatory mediators, primarily cytokines and leuko-attractant cytokines –chemokines and their receptors.
There is ample evidence that the classic concept of leukocyte extravasation, which is also termed the three (or more) signal paradigms, has relevance in CNS inflammation. It involves: immune cells tethering and rolling, chemo-attractant-triggered activation, firm arrest and diapedesis.
The interactions between cell-type selective leukoattractants (chemokines) and their receptors, are proposed to define the cellular composition of inflammatory infiltrate and may represent the key molecules that control the leukocyte invasion in the CNS.
However, some aspects of this regulation are not completely understood, for example, the restricted recruitment of neutrophils into the CNS or selective accumulation of monocytes and macrophages and their prevalence in numerous different neuropathological processes. Unlike other tissue, the CNS inflammatory reactions are often restricted to mononuclear cells, blood-born monocytes, lymphocytes and perivascular resident macrophages and the CNS resident cells (microglia).
12.1.4 The immunological significance of the Virchow-Robin space
The Virchow-Robin space (VR) separates cerebral blood vessels from the surrounding brain parenchyma (see Fig. 1).
In inflammatory disorders such as encephalitis or multiple sclerosis (MS), VR spaces typically expand and become a prominent site of accumulation of tightly packed cuffs of inflammatory cells. Intimate contact between cells in the RV space allows more effective presentation of antigens by resident macrophages to incoming immune cells and the effective utilization of various immunomodulatory factors. Ordered structures have been described that resemble lymphatic capillaries in close anatomical relation with chronically inflamed Virchow-Robin spaces. There, plasma cells and reticular cells surrounding T cells and macrophages within the perivascular channels are in close contact. It is supposed that these lymph node-like structures are the sites of B cell/plasma cell persistence, continuous antigen presentation and intrathecal antibody synthesis. Thus, within VR space immunological reaction could be orchestrated towards the most desirable response. Such reaction may be terminated in this space, sparing the CNS parenchyma. However, if the main source of antigen was in the brain itself, effector immune cells then migrate from the VR spaces into the brain parenchyma, causing encephalitis, myelitis or brain abscess, or in CSF space, causing meningoencephalitis.
12.1.5 Self-tolerance
It is well known that immunological tolerance to some self-components from nervous tissue, particularly the components of myelin (e.g. myelin basic protein – MBP, myelin-associated oligodendrocyte glycoprotein – MOG, ganglioside GM1 etc.) is poorly developed and autoimmune reactions are relatively easily induced, especially in experimental conditions (e.g., experimental allergic encephalomylitis ). The “opening” of the BBB, regardless of mechanisms (infections, inflammation, trauma) is a two-way street, permitting both the entry of immune cells and various inflammatory mediators into brain parenchyma, but also egress of e.g. myelin products to the blood stream, for example, during CNS infections, trauma etc.
It is well known that the infections with envelope viruses (e.g., measles, mumps, rubella, herpes simplex/zoster etc.) are more often associated with CNS damage and demyelination when compared to other infective agents. The cross-reactivity between a glycolipid in the envelope of a “budding” viruses and glycolipid in CNS myelin has been demonstrated. Thus, a special role is attributed to diverse infecting agents as trigger mechanisms in the pathogenesis of various (auto)immune-mediated diseases affecting nervous system (Guillain-Barré syndeome, acute disseminated encephalomyelitis-ADEM, multiple sclerosis – MS, etc.). Another mechanism by which nervous tissue might be involved in the pathogenetic mechanism is based on molecular mimicry (post-infection syndromes, paraneoplastic syndromes etc.). Several reports have shown that MBP and several viruses share amino acid sequences in the immunodominant region.
Interestingly, after cerebrovascular accidents or brain trauma, “expected” vigorous autoimmune reaction(s) to realeased components of destroyed nervous tissue is generally absent. Such finding support the concept of “neuroprotective (auto)immunity”.
However, a strong expansion of both T- and B- autoreactive lymphocytes which respond to myelin, or other self-antigens may have immunopathological consequences in genetically susceptible individuals (e.g., in subjects predisposed to develop multiple sclerosis -MS).
12.1.6 The humoral immunity and the clonal selection theory
The conventional defensive role of immune responses and much of their regulation are satisfactorily explained by clonal selection theory. The key feature of a specialized immune system is the fact that organisms contain specific antibodies before encountering antigen. The clonal selection theory postulated that each antibody-forming cell is genetically committed to express an antibody distinct from that of its companions. Each cell expresses its antibody as a surface receptor and can thus be selected by antigen.
Later, on encountering external antigens, the cells respond by clonal expansion and differentiation into antibody-secreting cells.
12.1.7 Peripheral B-cell pool
The peripheral B-cell pool consists mostly of naive B cells, the cells involved in primary immune responses that express the IgM+ IgD+ B-cell receptor (BCR) type, and several smaller subsets, including memory cells and the so-called B-1 cells.
Memory B cells form a minor subset of B cells. They are mostly resting, long-lived and, in part, recirculating cells. Naive B cells develop in memory B cells after antigen challenger in the secondary lymphoid organ with the help of CD4+ T-helper lymphocytes (see below).
The population of B-1 cells (CD5+) seems to be generated early in ontogeny, but are later largely confined to the peritoneal and pleural cavities. Their important role in natural defence is proposed (see later).
12.1.8 B-cells development
In adult mammals, B cells develop in the bone marrow and are produce throughout life. The stages of B cell development are marked by a series of changes that are determined by sequential rearragement of the Ig gene through the process of VDJ recombination. Successful completion of both heavy chain (IgH) and light chain (IgL) gene rearrangements result in an immature B cell expressing IgM. Once surface IgM is expressed, it can further act as an antigen receptor, and immature B cells that bind Ag in the bone marrow or in the periphery will die, change their receptor or become anergic. Only a small percentage of the immature B cells generated in the bone marrow appear to reach the periphery and thus become mature B-cells, some of them as long-lived cells with life spans of perheps even decades.
12.1.9 Affinity maturation of antibodies, immunological memory and germinal centres
Unlike T cells, B cells are subject to an antigen-dependent somatic mutation and a selection process that is designed to increase the affinity and functional efficiency of the memory Ig repertoire. The mechanism responsible for mutation is targetted to rearrange V-region genes and their immediate vicinity, introducing point mutations at a high rate. Affinity maturation and memory generation take place in the special microenvironment of germinal centres. Germinal centres (e.g., in human tonsil) arise inside “follicles” composed of naive B cells on immunisation with T-cell-dependent antigen.
Naive B cells develop in memory B cells with the help of CD4+ T-helper lymphocytes and follicular dendritic cells (FDC). The FDC carry antigen complexed to antibodies and components of the complement system on their surface. This form of antigen presentation, together with signals delivered by the T helper cells, is thought to be critical for the selection and maturation of high affinity memory cells. Thus, the germinal centre reactions are characterized by the rapid expansion of an oligoclonal population of antigen-activated B cells that express rapidly accumulating somatic mutations. Mutants accumulated rapidly over the first few weeks, and cells expressing high-affinity antibodies were strongly selected in the population. Furthermore, the Ig isotype switch that takes place in the germinal centre may be also an important contributor to efficient selection.
Finally, after a few weeks, the germinal centres shrink and largely disappear. Survival B-cells, after several runs of positive selection upregulate bcl-2 and enter a resting state. These cells express a novel repertoire of hypermutated, high-affinity antibodies.
Memory B cells present antigen very efficiently to T cells, so that renewed antigenic contact rapidly draws both T and B memory cells into the secondary response. In this response, proliferative expansion can take place withouth further somatic hypermutation (since most cells proliferate outside the germinal centre), and the cells finally undergo terminal differentiation into antibody-secreting nondividing plasma cells.
In the peripheral B-cell pool, memory cells can be distinquished by their somatically mutated antibodies, lack of IgD, characteristic surface markers such as CD27, and many of them have the switched Ig isotype class from IgM to IgG, A or E. Both, naive and memory B-cells express MHC class II molecules, which enables them to function as antigen presenters to T cells.
12.1.10 B-cell response within the T-cell network
The need of naive B cells to receive T-cell help for their activation is antigen dependent. Thymus-independent B-cell antigens are mitogens that induce a polyclonal B-cell activation. Immunoglobulin molecules on the surface of B-cells recognize antigen in its native form. Multimeric antigens, particularly on cell surfaces such as coating viral glycoproteins, can often directly stimulate B-cell proliferation and differentiation into plasma cells by efficient crosslinking the B cell receptors (BCR).
By contrast, most protein antigens cannot trigger an antibody response by themselves. In these cases induction of antibody response requires presentation of antigen in an “immunogenic form” (T-cell-dependent-response). It is in these responses that such B cells generate immunological memory and a new antibody repertoire by somatic hypermutation. Lastly, (auto)antibodies may be generated accidentally by B-cell superantigens and molecular mimicry.
12.1.11 Th1/Th2 paradigm
The concept that B-cells response within the T-cell network has been proposed by the discovery of functional T-helper cell subsets with characteristic cytokine release profiles and distinct regulatory functions on humoral immune responses (Th1/Th2 paradigm).
Activated Th1 lymphocytes typically secrete proinflammatory cytokines such as IL-2, INFy and lymphotoxin, which support cell-mediated and cytotoxic immune reaction, important in resistence to infection with intracellular pathogens. Activated Th2 lymphocytes typically secrete cytokines IL4-6, IL-10 and IL-13 which favor the activation of B-cells and eosinophils and thus support the production of antibodies (IgG1, IgA, IgE), important in resistence to infection with extracellular pathogens. However, Th1 cytokine IFNy can also promote the generation of antibodies, IgG2a and IgG3 classes. Th1 and Th2 cells crossregulate with each other through their cytokines, thus, for example, increase production of IgG2a, downregulate the production of IgG1 class and vice versa.
12.1.12 Second generation immune networks model
Although clonal selection has provided the rational basis for anti-infectious protection, many questions have been left unsolved such as: - internal lymphocyte activities and natural antibody production in unimmunized animals, pre-immune repertoire selection, tolerance and self-nonself discrimination, memory and the evolution of immune system. Recently, some theoretical advances, observations in unimmunized mice and humans, and the success of novel therapeutics in autoimmune diseases (e.g., with high doses of human immunoglobulins vs classic immunosuppresion) have supported the new ideas, that can be called second generation immune networks, proposing that some global properties of the immune system (such as natural tolerance) emerge from its network organisation expressed early in development and cannot be understood from the analysis of component parts only. “Immune networks” represent self-organizing activity of the molecular and cellular components via molecular affinities (complementarities), such as antibody networks.
12.2 Assessment of the humoral immune response within the CNS compartment
Human B cells function as effector cells through their secreted antibodies, the activation-dependent release of cytokines, and the mutual activation of T cells. Activated T and B cells easily cross the blood-brain barrier. If appropriate (auto) antigen is presented in the context of a MHC class II, B cells are activated in a T-cell-dependent manner, which results in B-cell differentiation into plasma cells, release of cytokines, and local antibody synthesis, which can induce diverse antibody-dependent effector mhanisms.
Many stages of lymphocyte transformation within the CNS compartment in different inflammatory diseases (infections, postinfection syndromes, autoimmune disease, neoplastic disease etc.) could be recapitulated in-vivo,
in the analysis of cerebrospinal fluid cells (morphologic study, immunocytochemistry, flow-cytometry, analysis of the IgG heavy chain variable (VH) region repertoire) and
B cells soluble products, primarily immunoglobulins (quantitative analysis, detection of “oligoclonal “ IgGs, detection of antibody specificity - target antigen(s).
According to such investigations, some characteristic features of intrathecal antibody responses have been confirmed, such as disease-related Ig class immune response, long-term Ig production within the CNS compartment, or polispecific and oligoclonal immune response.
In normal condition the antibodies within the CNS compartment is blood-derived and penetrate the CNS through the blood-brain barrieres (primarily blood-CSF barrier). Possible causes underlying the appearance of intrathecal antibodies are: a) acute CNS infections with a very specific immune response to a target microorganism, b) infections in the distant past with a persisting immune respons (e.g. TPHA Abs in neurosyphilis, or herpes simplex type 1 virus Abs in herpes simplex encephalitis), and c) a polyspecific intrathecal immune response associated with chronic inflammatory CNS diseases without the presence of the corresponding antigens (most frequently MS and systemic autoimmune diseases involving CNS). Although the target Ags for these polispecific Abs are generally unidentified, some of these Abs are produceed characteristically to measles, rubella and zoster viruses (“MRZ reaction”). Thus, the positive MRZ reaction in the CSF at time of first clinical symptom(s) strongly support the presence of chronic inflammatory processes like MS (vs an acute infections or post-infection syndromes such as ADEM).
In contrast to systemic immunity, in a various subacute and chronic inflammatory diseases, a variable amounts of Igs, up to 90%, could be synthesized intrathecally, commonly may exhibit oligoclonal restriction (see later), and can persist for months to years, e.g., in response to particular microorganisms such as paramyxoviruses (mumps, measles), herpes viruses, coxsackievirus or treponema palidum, or can be produced for lifetime (MS, SSPE).
How is this long-term antibody production maintained in CNS tissue?
Possible mechanisms include: a) reexposure to the (auto)antigen, b) persistent low-grade chronic, primarily viral infections, c) structural homologies of common viral antigens and self- antigens (eg, myelin) generating cross-reactive immune response (“molecular mimicry”), d) persistence of antigen on local reticular cells which form specialized contactas with plasma cells in lymph-node like structures, and e) the existence of B-cell-supporting microenvironment within the CNS compartment.
12.2.1 Long-term B-cell surveillance and memory B cells activation
The factors governing B-cells recruitment within CNS compartment (chemokines) and B- cells surveillance in the inflamed lesions remains to be elucidated. Interestingly, recent study has demonstrated that the normal brain microenvironment constitutively supports the differentiation of B cells into antibody-secreting plasma cells (i.e., Th2-type hierarchy of CNS immune regulation. Further, similar lymph-node like structures have been observed within the target tissue of a variety of autoimmune and inflamatory disorders characterized by antigen-driven clonal proliferation of B cells. Finally, a variety of cellular and soluble factors that are favorable for survival of memory B cells (e.g., nerve growth factors) and their activation and differentiation in plasma cells could be demonstrated, for example, in MS plaques or CSF samples. Memory B cells can be activated with specific T-cell help, similarly to naive B cells. However, in addition, it has been demonstrated that the orchestrated action of specific cytokines such as IL2, IL4 and IL10, and provision of the essential signal from any activated T cell (CD40L) could be sufficient to induce bystander memory B-cell activation, differentiation and Ig secretion in vitro. If TNFa is present, there is no need for T cells.
12.2.2 Intrathecal antibody response: disease-related immunoglobulin class response
The antibody response within the CNS is commonly characterized by the lack of switch from IgM class response to IgG class response, it can exhibit relatively constant relation over months of disease and can be related to a particular disease with particular cause (Igs “pattern”). For example, typical pattern of theree-class immune response with predominant IgM class could be seen in neuroborreliosis, or IgG and IgM in some cases of MS, or IgG in herpes simplex encephalitis and neurosyphilis. This common absence of classic immune regulation in brain might reflect the low level of regulatory cells and immunomodulatory molecules.
12.2.3 High- and low-affinity antigen-specific antibodies (IgG)
In a various pathologic conditions intrathecally produced Igs could be confirmed. However, the pathogenetic role of locally produced antibodies (autoantibodies) is less well defined (key players or bystanders ?). In many cases is unknown wheather B cells synthesize beneficial, deleterious, or irrelevant antibodies, because, among others, the antigen-specificity of such antibodies is unknown, or diverse antibody specificities could be demonstrated (polyspecific immune response).
In MS, but also in patients with chronic CNS inflammation (eg, systemic autoimmune diseases involving CNS, cronic infections), CSF analysis can demonstrate the presence of antigen-specific IgGs to one or more viral (measles, rubela, varicella/zoster) antigens («MRZ»-reaction) or other antigens (herpes simplex, toxoplasma, a number of self-antigens, heat-shock proteins, Alu reapet etc. Thus, the finding of antigen-specific IgG per se does not help to differentiate between patients with chronic inflammatory autoimmune disease, such as MS and chronic CNS infections.
However, in MS most of these antibodies bind weakly to target antigens, and a central pathogenic role of these low-affinity IgGs of highly diverse antigenic specificity is less likely. They are probably synthesized as a secondary phenomenon (eg, inbalance of cytokines). However, it is important to notice that the target antigen(s) for the most intratecally sinthesized antibodies (oligoclonal IgGs) seen in MS, is yet unindentified.
In contrast, the patients with CNS infections (e.g., herpes simplex encephalitis, varicella zoster encephalitis, SSPE), produce locally high-affinity antibodies, most of which are directed against the causative antigen. For example, in SSPE anti-measles IgGs bind firmly to viral antigen(s) vs anti-measles IgGs in MS.
However, a number of other (auto)antigen-specific Igs which are not directed against the causative organism, could be demonstrated in such chronic CNS infections. The majority of these antibodies were probablly generated by antigen-independent mechanism(s), and they are of low-affinity.
12.3 “Oligoclonal” immune response within the CNS compartment (OIg)
One of the most characteristic features of intrathecally synthesized antibodies (most frequentlly of IgG class) in many subacute and primarily chronic CNS inflammatory diseases is their restricted heterogeneity according to their mobility in electrical field, that could be demonstrated by different electrophoretic techniques. From an immunological point of wiev, Abs of restricted heterogeneity indicates clonally restricted B cell activity. Since the “oligoclonal” IgGs (OIgGs) in CSF generally express diverse antibody specificities (polyspecific immune response), so-called «oligoclonal IgG» reflect primarily the polyspecific and only secondarily the oligoclonal nature of immune response. The term «oligoclonal» Ig persists so far, because it was introduce in CSF analysis long before a clonal selection theory of the immune system was considered.
12.3.1 Elecrophoretic metods for investigation of OIg bands (methodological improvements)
After the introduction of electrophoresis on solid matrix (paper, agar-gel) it was possible to show that proteins accumulate as individual bands in the gamma globulin region in patients with MS, neurosyphilis and a variety of other chronic inflammatory neurological diseases. These bands of restricted heterogeneity were designated as ”oligoclonal” in contrast to the normal “polyclonal” Ig pattern of normal serum and CSF. Polyclonal Igs pattern of normal serum and CSF reflect the practically infinite heterogeneity of the individual antibodies. “Oligoclonal” Ig bands can be seen in CSF more easily because the background concentration of the polyclonal gamma globulin in CSF is normally very low, due to the presence of the blood-brain barriers and restricted entrance of blood proteins in CNS compartment.
During the years, many procedures for the analysis of oligoclonal Igs have required a large volume of CSF specimens, which must be previously concentrated by ultrafiltration. Also, some highly alkaline proteins which comigrate with gamaglobulin fraction (e.g., gama- trace protein) could be misinterpreted as IgG band. Further, the resolving power of conventional zonal electrophoreses on agarose or cellulose acetate is limited. Therefore, the trends in methodological improvment for the detection of oligoclonal Igs (generally IgG) have been focused to improve resolution power of the protein separation method, to increase the specificity of Ig detection and to enhance the sensitivity of detection system (Fig. 2).
Figure 2.
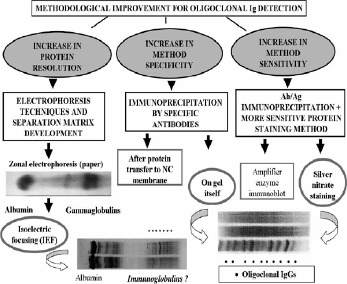
Trends in methodological improvement for oligoclonal Ig detectiom
Several new procedures have been gradually introduced:
the method for protein separation in a buffering pH gradient gel (isoelectric focusing -IEF) greatly improved the resolution power of the separation method (proteins are separated according to their different isoelectric points - pI values, therefore IEF allows concentrating small amounts of proteins into tight bands ),
immunofixation step with Abs to ã-chain of human Ig, or ê/ë free Ig chains increases the specificity of the method (immunoglobulin class characterization), and also the sensitivity of the method, because there is Ag+At complex,
development of higly sensitive detection method (silver staining, immunoenzyme staining after protein transfer to nitrocellulose membrane and ultrasensitive procedure of immunoenzyme staining +avidin-biotin amplification step) makes possible to detect oligoclonal Igs in unconcentrated (native) CSF sample. By including avidin-biotin amplification step, the sensitivity of detection system is increased five to ten times,
lastly, the method called “affinity-mediate immunoblot” has been introduced to identify the antigenic specificity of oligoclonal IgGs bands (this procedure includes protein transfer to antigen-coated nitrocellulose membrane after IEF). Accompanied with avidin-biotin amplification procedure, the method is very suitable for searching of minor fractions of antigen-specific oligoclonal Igs (covered with polyclonal Igs, or with other more prominent oligoclonal bands), such as antigen-specific oligoclonal IgG in AIDS-related cytomegalovirus and toxoplasma encephalitis, (Figs. 3a, b).
Figure 3a.

Demonstration of oligoclonal IgG by IEF and two different detection systems
In our Laboratory we use a very simple, highly sensitive and inexpensive method for detection of oligoclonal IgG bands, which we developed twenty years ago. Our method include IEF on ultrathinlayer-polyacrylamide gel (IEF/PAGE), followed by direct immunofixation on gel and double silverstaining of precipitated At+Ag complexes. Omiting the step of protein transfer to nitrocellulose membrane, the loss of weak bands is almost neglected (Figs. 3a, 4, 5).
Figure 3b.
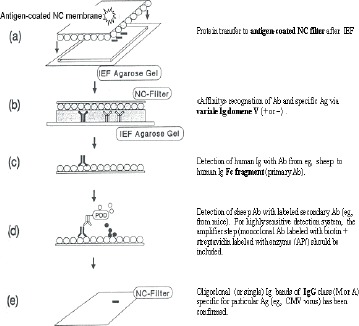
Detection of antigen-specific oligoclonal IgG by «affinity mediated immunoblot» method.
Figure 4.
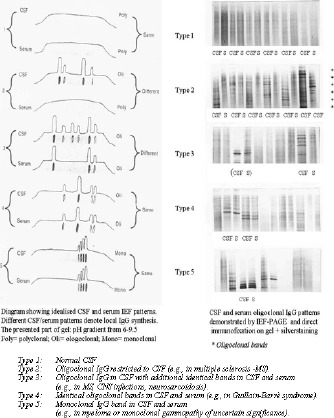
International Consensus according to interpretation of oligoclonal IgG detection by IEF/immunoblot: five types of results for paired analyses of CSF and serum are recomended.
Lastly, it is importane to notice that all variants of high-resolution zonal electrophoresis (e.g., high-resolution electrophoresis on “Cellogel” strips + immunofixation) are inferior to IEF with immunofixation, especially when the CSF-restricted oligoclonal IgGs were present in low concentrations, or in many cases of faint so-called “mirror pattern”.
12.3.2 Classification of isoelectric focussing OIgGs patterns (International consensus)
The strongest consensus is that isoelectric focusing, followed by IgG specific antobody staining (immunofixation) is the most sensitive test for the detection of oligoclonal bands when using the same amounts of IgG in parallel CSF and serum samples. This method (variants) should be used as the standard method in the reference laboratory. It is important to exclude artefactual bands that are caused by non-linearity of the isoelectric focusing pH gradient. Therefore, the choice of commercial source of ampholytes (synthetic basis and acids) is more important than the choice of support media (for example, agarose vs polyacrylamide). Since many disorders cause oligoclonal IgG bands in serum, which may also be present in the CSF due to disruption of the blood-brain barriers, the parallel analysis of CSF and serum is imperative.
The definition of oligoclonal Igs in CSF using IEF is that is necesary to demonstrate the products of at least two clones of lymphocytes within the CSF (i.e., at least two IgG bands in CSF pattern which are absent from serum pattern). The International Consensus for detection of oligoclonal IgG proposed five types of results for paired analysis of CSF and serum (Fig. 4):
Type 1: Normal CSF
Type 2: Oligoclonal IgG restricted to CSF
Type 3: Oligoclonal IgG in CSF with additional identical bands in CSF and serum (combination of types 2 and 4)
Type 4: Identical oligoclonal bands in CSF and serum
Type 5: Monoclonal IgG bands in CSF and serum (rnyeloma or monoclonal gammopathy of uncertain significance).
In addition, in a small subset of patients only a single IgG band restricted to CSF could be demonstrated (~ 0.5%).
One of the major problem which may occures in the interpretation of IEF pattern is that of “monoclonal“ IgG. It is important to know that classic “monoclonal” IgG band seen in zonal electrophoresis in patients with monoclonal gammopathy, on IEF produces a very specific ladder pattern of four to five bands equally spaced from each other with descending intensity toward the anodic part of the blot. Such pattern has been shown to be due to postsynthetic modification of the antobody and does not represent multiple unique antibodies. Further, the finding of a single IgG band on IEF does not necessarily mean also a monoclonal IgG (i.e., the product of a single plasma cell clone), because two-dimensional gel electrophoresis studies have shown that an individual IgG band detected by IEF actually contains a few well resolved individual spots (i.e., they are products of several plasma cell clones).
12.4 The clinical significance of oligoclonal IgGs in neurological diseases
It is well known that subacute and primarily chronic inflammatory disorders of the CNS are frequently not recognized until a CSF examination is performed when both cellular and humoral reactions could be observed. Perheps the greates value of oligoclonal IgGs detection is that it can more fequently show an inflammatory origin (key, or bystander?) of neurological disfunction. Of most interest for neurological diagnosis are types 1, 2 and 3, because they reject (type 1) or confirm (types 2 and 3) the intrathecal IgG production. The most patients showing OIgGs could be interpreted as type 2 (also see tab. 3).
Table 3.
Laboratory findings in MS and diseases mimicking MS
| CSF oligoclonal IgG | Magnetic resonance imaging | Visual evoked potentials | Peripheral nerve and/or muscle involvment | |
|---|---|---|---|---|
| Multiple sclerosis | + | + | + | - |
| Disseminated | ||||
| encephalomyelitis | + | + | + | + |
| Devic's disease | + | + | + | + |
| HTLV-I assosiated | ||||
| paraparesis | + | + | + | + |
| Nervous system AIDS | + | + | + | + |
| Neurolupus | + | + | + | + |
| Neurobrucellosis | + | + | + | + |
| Neuroborreliosis | + | + | + | + |
| Neurosarcoidosis | + | + | + | + |
| Chronic fatigue | ||||
| postviral syndrome | + | + | + | + |
The frequencies of oligoclonal IgG positive findings in neurological patients greatly depend on diagnosis (Tab. 2), but also on method applied. All patients with SSPE and almost all patients with definite MS (~95%) show OIgG. Thus, the presence, or particularly the absence of oligoclonal IgG in CSF, may strongly support or reject the possible clinical diagnosis of MS and SSPE.
Table 2.
Frequencies of CSF oligoclonal IgG bands and the intrathecally synthesized IgG in subjects affected with various neurologic diseases
| Diagnosis | CSF oligoclonal IgG | Intrathecally sintesized IgG fraction |
|---|---|---|
| Patients (%) with positive results | Patients (%) with positive results (IgGIF>0) | |
| 1. Subacute sclerosing panencephalomyelitis (SSPE) | 100 | 100 |
| 2. Multiple sclerosis (MS): | ||
| ♢ Definite | 97 | 75 |
| ♢ Probable | 73 | 63 |
| ♢ Primary progresive MS | 79 | |
| ♢ MS in childhood | 64 | |
| 3. Optic neuritis | 63 | 33 |
| 4. Chronic focal myelopathy of unknown cause | 67 | 33 |
| 5. Postinfective and postvaccinal complications | 58 | 33 |
| ♢ Acute disseminated encephalomyelitis (ADEM) | 29 | 16 |
| 6. CNS infections: | ||
| ♢ Neurosyphilis | 80 | 50 |
| ♢ Neuroborreliosis | 63 | 38 |
| ♢ Neurotuberculosis | 20 | 15 |
| ♢ HIV encephalytisst. III | 45 | 20 |
| ♢ Opportunistic infections | 50 | 50 |
| ♢ VZV ganglionitis | 30 | 15 |
| ♢ VZV meningitis | 15 | 15 |
| ♢ Acute mumps meningitis | 12 | 19 |
| 7. Connective tissue diseases involving CNS and vasculitides | ||
| ♢ Neurosarcoidosis | 36 | |
| ♢ Neurolupus | 28 | |
| ♢ Behcet's disease | 16 | 78 |
| 8. Nervous system inflammations | ||
| ♢ Inflammatory polyneuropathy | 33 | 11 |
| 8 | 16 | |
| ♢ Guillain-Barrè syndrome | ||
| 9. Other neurological diseases |
By using a high sensitive IEF/immunoblot method (variants), oligoclonal IgGs bands can be demonstrated in a greater number of non-MS patients, such as in patients with chonic CNS infections (30-70%), post-infectious CNS diseasese such as ADEM (up to 30%), systemic autoimmune diseases involving the CNS (up to 50%), paraneoplasic syndromes, cerebrovascula diseases etc. Thus, from the aspect of MS diagnosis, there is a loss in specificity because IEF methods reveal more bands (vs the variants of high-resolution zonal elecropforesis) and more non-MS patients become positive.
Further, blood-derived oligoclonal IgGs in CSF (types 3 and 4) can give additional information because provide evidence of systemic immune activation only (type 4), or both, systemic and intrathecal (type 3). Diagnosis in group of patients showing type 3 generally resemble those among patients with type 2 (MS patient and patients with CNS infections). Lastly, the “mirror-pattern” (type 4) is in most frequently associated with various peripheral inflammatory neuropathies (e.g., Guillain-Barre sy.), neoplastic disorders, and less frequently with systemic autoimmune diseases or systmic infections. The patients with paraneoplastic syndromes and systemic autoimmune diseases involving CNS, synthesize IgG intrathecally more common (see tab. 2.).
12.5 Multiple sclerosis
MS is the most common chronic neurological diseases in young adults of unknow cause yet, characterize by inflammation, demyelination, axonal degeneration and gliosis. It is extremely variable disease in terms of its clinical course, prognosis and particularly clinical manifestations. Regardless of the clinical course and pathogenetic pathway, the “MS brain” is genetically programed to produce unique pathological changes, so-called “MS plaques”.
However, many changes often found in MS by magnetic resonance imaging techniques (MRI) and CSF analysis (including oligoclonal IgG bands) are non-specific, and can be found in a number of other unrelated conditions (Tab. 3) which could be confused with MS.
12.5 1 The early stage of multiple sclerosis (MS)
During the early stage of MS pathology, primarily lymphocytic infiltration of Robin-Virchow spaces could be demonstrated in normal appearing white matter. These minor vascular changes (mild vasculitis) produce very minor alteration of blood-brain barrier (demonstrated by gadolinium-enhanced MRI), but may be the pathway for activated B-cells to penetrate into the CNS, where they produce a broad range of polyspecific antibodies, most of them of restricted homogeneity, that could be demonstrated in CSF as «oligoclonal IgG bands». Such intrathecal production of oligoclonal IgG has been demonstrated in almost all MS patients, but also in the CSF of many unaffected siblings and twins of MS patients. This implies that at this early stage of «disease» the parenchyma of the CNS remains generally intact. The true disease with its unique «MS plaques» may never develop in such an individual. Furthermore, MS patients as well as their unaffected siblings and tweens have the enhanced ability to produce antibodies against many antigens, including neurotropic viruses (measles, rubella, varicella/zoster). From an immunological point of view such persons are characterized as «strong responders». Such, unique response is even more prominent within the CNS compartment and could be demonstrated as positive «MRZ» reaction, i.e., increased antibody specificity index (ASI) for such pathogens. All these findings imply the immunogenetic background as a risk of acquiring MS.
12.5.2 Oligoclonal IgGs in MS patients
It has been proposed that OIgGs appear early in the course of MS. The numbers, intensity and the position of OIgG bands on IEF gel differ from patient to patient but remain fairly constant in a given patient during the course of diseases (vs other conditions such as ADEM) (Fig. 5). ADEM and postinfectious encephalomyelitis (PEM) resemble MS and constitute a major category of differential diagnosis with MS (especially in childrens).The target antigen(s) for the major OIgs are unknown so far. The treatment by corticosteroids failes to modify the oligoclonal IgG band pattern (vs other conditions such as systemic autoimmune disease. The negative results for OIgGs by IEF/immunoblot should address the investigators to another diagnosis, because only about 3% of patients with definite MS are IgG band negative MS. Young MS patients show less frequently OIgGs, than adults (~ 75% vs ~ 95%). However, in rare cases of young MS patients, oligoclonal IgG bands could be absent in early stage of disease, or be presented as a weak single band. However, during the dissemination of pathological process, in time and space, the new bands could be appeared.
Finally, the presence of oligoclonal IgGs also provides some prognostic information. In patients with isolated neurological syndromes which might represent a first attack of MS, such as optic neuritis, brainsteam or spinal cord syndromes (eg, transverse myelitis), evidence of OIgGs in CSF indicates an increased risk factor for the future development of clinically definite MS.
12.6 In summary
Many studies have reported that detection of the oligoclonal IgG in the CSF is the most sensitive test to confirme intrathecal IgG synthesis in various subacute and chronic neuroinflammatory diseases, but the presence of oligoclonal IgGs per se is not a specific marker for any diseases, and the location and number of bands generally have no importance for interpretation and diagnosis. Perheps the greates value of oligoclonal IgGs detection is that it can more fequently show an inflammatory origin (key, or bystander?) of neurological disfunction. Of most interest for neurological diagnosis are types 1, 2 and 3, because they reject (type 1) or confirm (types 2 and 3) the intrathecal IgG production.
Figure 5.
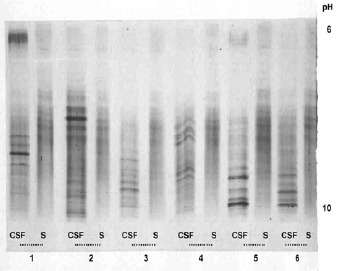
CSF and serum oligoclonal IgG patterns demonstrated by IEF-PAGE and direct immunofixation on gel + silver staining in patients with MS (2-6) and cryptococcal meningitis (1).
References
- 1.Rajewsky K. Clonal selection and learning in the antibody system. Nature 1996; 381: 751-388. [DOI] [PubMed] [Google Scholar]
- 2.Xiao BG, Link H. Immune regulation within the central nervous sytem. J Neurol Sci 1998;157: 1-212. [DOI] [PubMed] [Google Scholar]
- 3.Esiri MM, Gay D. Immunological and neuropathological significance of the Virchow-Robin space. J Neurol Sci 1990; 100: 33-108. [DOI] [PubMed] [Google Scholar]
- 4.Kerschensteiner M, Stadelmann C, Dechant G, Wekerle H, Hohlfeld R. Neurotrophic cross-talk between the nervous and immune system: Implications for neurological diseases. Ann Neurol 2003; 53: 292-304. [DOI] [PubMed] [Google Scholar]
- 5.Reiber H. Cerebrospinal fluid-physiology, analysis and interpretation of protein patterns for diagnosis of neurological diseases. Multiple Sclerosis 1998; 4: 99-107. [DOI] [PubMed] [Google Scholar]
- 6.Reiber H, Ungefehr S, Jacobi Chr. The intrathecal, polyspecific and oligoclonal immune response in multiple sclerosis. Multiple Sclerosis 1998; 4: 111-117. [DOI] [PubMed] [Google Scholar]
- 7.Archelos JJ, Storch MK, Hartung H-P. The role of B cells and autoantibodies in Multiple sclerosis. Ann Neurol 2000; 47: 694-706. [PubMed] [Google Scholar]
- 8.Correale J, Molinas M. Oligoclonal bands and antibody responses in multiple sclerosis. J Neurol 2002; 249; 375-389. [DOI] [PubMed] [Google Scholar]
- 9.Paolino E, Fainardi E, Tola MR., Govoni V, Casetta I, Monetti VC, Granieri E, Carreras M. A prospective study on the predictive value of CSF oligoclonal bands and MRI in acute isolated neurological syndromes for subsequent progression to multiple sclerosis. J Neurol Neurosurg Psychiatry 1996; 60: 572-575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.McLean BN, Miller D, Thompson EJ. Oligoclonal banding of IgG in CSF, blood-brain barrier function and MRI findings in patients with sarcoidosis, systemic lupus erithematosus. and Behcet's disease involving the nervous system. J Neurol Neurosurg Psychiatry 1995; 58: 548-554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kostulas VK, Link H, Lefvert AK. Oligoclonal IgG bands in cerebrospinal fluid. Principles for demonstration and interpretation based on findings in 1114 neurological patients. Arch Neurol 1987; 44: 1041-1044. [DOI] [PubMed] [Google Scholar]
- 12.McLean BN, Luxton RW, Thompson EJ. A study of immunoglobulin G in the cerebrospinal fluid of 1007 patients with suspected neurological disease using isoelectric focusing and the Log IgG-index. A comparison and diagnostic application. Brain 1990;113: 1269-1289. [DOI] [PubMed] [Google Scholar]
- 13.Dörries R, Ter , Meulen V. Detection and identification of virus-specific, oligoclonal IgG in unconcentrated cerebrospinal fluid by immunoblot technique. J Neuroimmunol 1984; 7: 77-89. [DOI] [PubMed] [Google Scholar]
- 14.Zeman A, McLean B, Luxton R, Sharief M, Thompson E. The significance of serum oligoclonal bands in neurological diseases. J Neurol Neurosurg Psychiatry 1993; 56; 32-35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Andersson M, Alvarez-Cermeno J, Bernardi G, Cogato I, Fredman P, et al. Cerebrospinal fluid in diagnosis of multiple sclerosis: a consensus report. J Neurol Neurosurg Psychiatry 1994; 57: 897-902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Verbeek MM, de Reus HPM, Weykamp. Comparison of methods for the detection of oligoclonal IgG bands in cerebrospinal fluid and serum: Results of the Dutc Quality Control Survey. Clin Chem 2002; 9: 1578-1580. [PubMed] [Google Scholar]