

Abstract
Carcinoembryonic antigen-related cell adhesion molecule 1 (CEACAM-1) is the major antigen of the CD66 cluster of granulocyte differentiation antigens. The present study aimed to assess the biological function of CEACAM-1 on the growth of human colorectal cancer (CRC) cells in vitro. Treatment of cultured CRC HCT-8 cells with CEACAM-1-specific siRNA successfully downregulated CEACAM-1 expression by 61% compared with control cells. The effects of CEACAM-1 downregulation on HCT-8 cell proliferation and apoptosis were then assessed via Cell Counting kit-8 assay and flow cytometry, respectively. The results demonstrated that siRNA-induced CEACAM-1 downregulation significantly inhibited proliferation and increased apoptosis, but had no significant effect on cell cycle progression in HCT-8 cells. Together, these results suggest that CEACAM-1 activity is critical to CRC growth, and thus, CEACAM-1 may be a promising therapeutic target for the treatment of CRC.
Keywords: colon cancer, carcinoembryonic antigen-related cell adhesion molecule 1, HCT-8 cell line, small interfering RNA
Introduction
Malignant tumor growth is a common and serious disease condition that may affect any number of different organs and tissues throughout the body, and incur varied and complex symptoms that are often life threatening. Colorectal cancer (CRC) is one of the most common gastrointestinal malignancies, and localizes to the rectum or the junction of the rectum and the sigmoid colon. In fact, CRC is the fourth most common type of cancer internationally, following gastric, esophageal, and lung cancer (1,2).
Malignant tumors are often characterized by both tissue architecture disruption, and differentiation derangement. Cell adhesion dysregulation contributes to tumor invasion and metastasis (3), such that the abnormal expression of various cell adhesion molecules induces a loss of cell-cell binding, thereby promoting tumor differentiation and malignant invasion (4). Carcinoembryonic antigen-related cell adhesion molecules (CEACAMs) are members of the glycosylphosphatidylinositol (GPI)-linked immunoglobulin (Ig) superfamily (5). CEACAM-1 is a CEACAM subtype that is also known as biliary glycoprotein I or CD66a (6,7). Previous studies have demonstrated that CEACAM-1 expression is reduced in several tumor types, such as melanoma, lung, colon and ovarian cancer, compared with the corresponding normal tissues (8–14). This suggests that CEACAM-1 may function to inhibit carcinogenesis. In addition, CEACAM-1 has been reported to promote the apoptosis of various cells, including pulmonary and mammary epithelial cells, oral keratinocytes, cancer cells, Jurkat T cells, and cardiomyocytes (13,15,16).
To date, the expression of CEACAM-1 in CRC has not been investigated. In the present study, CEACAM-1 expression was silenced in a CRC cell line, and its effects on cell growth and apoptosis were examined. The findings provide the first evidence that decreased CEACAM-1 expression promotes CRC progression.
Materials and methods
Cell culture
HCT-8 cells (Type Culture Collection of the Chinese Academy of Sciences, Shanghai, China) were maintained (37°C, 5% CO2) in Dulbecco's modified Eagle's medium supplemented with 10% fetal calf serum (all from Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 100 µg/ml penicillin, and 100 µg/ml streptomycin. The cells were passaged every 2–3 days, using 0.02% EDTA and 0.1% trypsin.
Small-interfering RNA (siRNA) design and cell transfection
Vectors carrying either siRNA specific to CEACAM-1 or a negative control siRNA were generated by GeneChem Co., Ltd. (Shanghai, China). Three CEACAM-1-specific siRNAs were designed, as follows: siRNA1 sense, 5′-CAGCCACAGAAAUAAUUUATT-3′ and antisense, 5′-UAAAUUAUUUCUGUGGCUGTT-3′; siRNA2 sense, 5′-CCGUCAAAUUGUAGGAUAUTT-3′ and antisense, 5′-AUAUCCUACAAUUUGACGGTT-3′; and siRNA3 sense, 5′-GAGCUCUUUAUCCCUAACATT-3′ and antisense, 5′-UGUUAGGGAUAAAGAGCUCTT-3′. A siRNA encoding a nonsense sequence (sense, 5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense, 5′-ACGUGACACGUUCGGAGAATT-3′), was designed and used as a negative control (GenePharma Co., Ltd., Shanghai, China). HCT-8 cells were cultured until they reached 50% confluence, washed with PBS, and transfected with 50 nM CEACAM-1-specific or control siRNA, according to the manufacturer's instructions. Cells were collected 48 h post-transfection, and maintained in fresh medium for 24 h prior to further experimentation and/or analysis.
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR)
CEACAM-1 expression was analyzed via RT-qPCR, as previously described (17). Briefly, total RNA was extracted from CRC cells, using an RNeasy Mini kit (Invitrogen; Thermo Fisher Scientific, Inc.). Genomic DNA was removed from the extracted total RNA via DNase I digestion, and then the total RNA was reverse transcribed to generate cDNA, according to the manufacturer's instructions (PrimeScript™ 1st strand cDNA Synthesis kit; Takara Biotechnology Co., Ltd., Dalian, China). qPCR was performed in triplicate in a 10 µl reaction mix consisting of 4 µl template DNA (0.05 µg/µl), 5 µl SYBR-Green (Takara Biotechnology Co., Ltd.), 0.2 µl each forward and reverse oligonucleotide (10 µM each) and 0.6 µl deionized water. The thermocycling conditions were as follows: 95°C for 10 min, followed by 40 cycles of 95°C for 10 sec, 60°C for 30 sec, and 72°C for 30 sec. The primers used for qPCR were: CEACAM-1 forward, 5′-CAGGGGCTTCTGCTCACAGC-3′ and reverse, 5′-AGTTGCTTCTTCACAAGAT-3′; β-actin forward, 5′-GGCTGTGGAGACAAAAATGACCTC-3′ and reverse, 5′-AGGCTTGGGCTTGAATGGAGTC-3′. The expression level was estimated with the 2−ΔΔCq method (18).
Western blot analysis
CEACAM-1 expression levels were determined via western blot analysis. CRC cells were washed twice with PBS, and collected via centrifugation. Proteins were then extracted with cell lysis buffer (CST Biological Reagents Company, Ltd., Shanghai, China) containing 1 mM phenylmethylsulfonyl fluoride, and the protein concentration of each sample was determined using the BCA protein assay reagent kit (Beyotime Institute of Biotechnology, Lianyungang, China), and bovine serum albumin was used as a standard. The samples were denatured, and 50–80 µg were separated via polyacrylamide gel electrophoresis and transferred onto nitrocellulose membranes. The membranes were incubated first (4°C, overnight) with anti-CEACAM-1 (cat. no. AF1857; Novus Biologicals, Ltd., Cambridge, UK; dilution, 1:200) and anti-β-actin (cat. no. ab8227; Abcam, Cambridge, UK; dilution, 1:4,000) antibodies. The membranes were then incubated with horseradish peroxidase-conjugated rabbit anti-human secondary antibodies (cat. no. ab6759; Abcam; dilution, 1:3,000) for 12 h at 4°C, and finally with enhanced chemiluminescence substrate (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The resulting blots were analyzed using ImageJ software (NIH, Bethesda, MA, USA).
Cell proliferation
Cell proliferation was assessed by a CCK-8 assay (Roche Diagnostics GmbH, Mannheim, Germany), according to the manufacturer's instructions. Typically, cells were plated at a density of 1.5–2×103 cells/well in 96-well plates. After 48 h (to allow cell adherence to occur), cells were incubated with colorimetric substrates. Colorimetric changes were then measured in a multi-well spectrophotometer (MR5000 Multiplate Reader; Dynatech, Denkendorf, Germany), and cell survival following treatment was expressed as a % of viable cells relative to control cell values. All experiments were independently conducted three times, and the results of the three experiments were then averaged.
Cell cycle assay
Cells were fixed, washed with PBS, treated with RNaseA, and stained (37°C, 30 min) with 25 µg/ml propidium iodide (PI). The samples were then analyzed via flow cytometry, and the cell cycle phase distribution was quantified using Modfit Software (BD Biosciences, Franklin Lakes, NJ, USA). The proliferative index was calculated to represent the % of cells identified to occur in the S/G2/M phase.
Apoptosis assay
Cells were collected, stained with Annexin V-fluorescein isothiocyanate (FITC) and 7-Aminoactinomycin D using an Annexin V-FITC Apoptosis Detection kit (KeyGen Biotech Co., Ltd., Nanjing, China) according to the manufacturer's instructions, and analyzed via flow cytometry (BD Biosciences).
Statistical analysis
All results are presented as the mean ± standard error of the mean. Differences between two groups were evaluated by the unpaired Student's t-test. One-way analysis of variance with post-hoc analysis by Bonferroni's test was performed to evaluate the data generated by the cellular viability and apoptosis assays. Additional statistical analyses were performed by Student's t-tests. P<0.05 was considered to indicate a statistically significant difference.
Results
Knockdown of CEACAM-1 in HCT-8 cells
To investigate the function of CEACAM-1 expression in CRC cells, three separate siRNA sequences, comprising CEACAM-1-siRNA1, CEACAM-1-siRNA2, and CEACAM-1-siRNA3, were designed. When testing their efficacy, the three sequences induced a 61.3, 32.4 and 47.3% decrease in CEACAM-1 mRNA expression, respectively, in the CEACAM-1-knockdown cells compared with the negative control HCT-8 cells (Fig. 1A). The results from western blot analysis revealed a concordant reduction in CEACAM-1 protein expression levels compared with the control group (Fig. 1B). The CEACAM-1-siRNA1 sequence was selected for use in subsequent experiments, since it induced the greatest reduction in CEACAM-1 expression.
Figure 1.
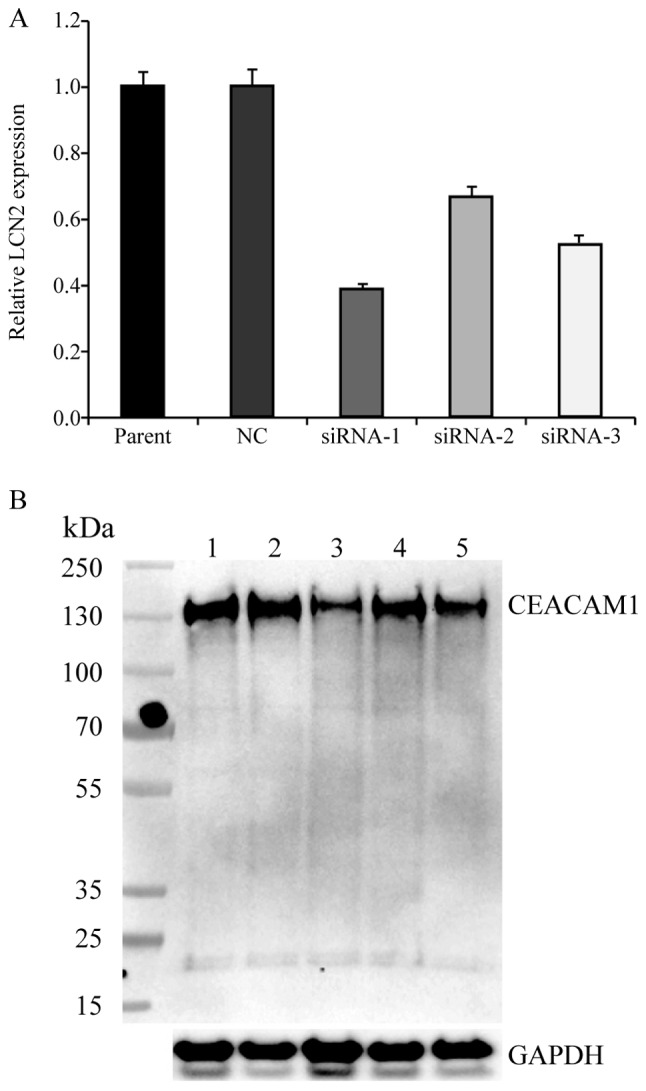
Knockdown of CEACAM-1 expression in HCT-8 cells. (A) CEACAM-1 mRNA expression was suppressed using three different siRNA constructs in HCT-8 cells. Parental untreated cells and cells transfected with negative control (NC) siRNA were used as controls. The experiment was performed in triplicate. (B) Representative immunoblot of CEACAM-1 protein expression in HCT-8 cells, where GAPDH was used as a loading control. Lane 1, untreated group; lane 2, NC siRNA; lane 3, siRNA-1; lane 4, siRNA-2; lane 5, siRNA-3. CEACAM-1, carcinoembryonic antigen-related cell adhesion molecule 1; si, small interfering; NC, negative control.
CEACAM-1 downregulation inhibits cell proliferation
CEACAM-1 knockdown significantly decreased the numbers of viable HCT-8 cells compared with the control group (Fig. 2), suggesting that CEACAM-1 silencing inhibited CRC cell proliferation.
Figure 2.

Downregulation of CEACAM-1 inhibits colorectal cancer cell proliferation. HCT-8 cells transfected with either CEACAM-1-siRNA or a negative control siRNA were assessed for cell proliferation using the CCK-8 assay. Results are presented relative to the control group. CEACAM-1, carcinoembryonic antigen-related cell adhesion molecule 1; si, small interfering.
CEACAM-1 downregulation increases cell apoptosis
Next, the effect of CEACAM-1 silencing on apoptosis was determined in CRC cells. The results from Annexin V cytometric analysis revealed that CEACAM-1 downregulation resulted in a significant increase in the apoptotic rate of HCT-8 cells (1.23% in the CEACAM-1-knockdown group compared with 0.197% in the control group; P<0.01; Fig. 3). The experiment was performed in triplicate.
Figure 3.

Downregulation of CEACAM-1 promotes colorectal cancer cell apoptosis. Representative plots are shown from flow cytometry analysis following Annexin V staining. (A) Untreated group. (B) Negative control siRNA group. (C) CEACAM-1-specific siRNA group. (D) Quantification of the results. *P<0.01. CEACAM-1, carcinoembryonic antigen-related cell adhesion molecule 1; si, small interfering.
CEACAM-1 might function to inhibit carcinogenesis, which is often associated with both cell cycle arrest and activation of the cell death pathway. Therefore, the cell cycle phase distribution was examined in the CEACAM-1-knockdown cells to determine whether CEACAM-1 knockdown has an effect in cell cycle arrest. The results revealed no significant difference in the % of G2/M-phase cells that occurred in the CEACAM-1-knockdown compared with the control group (P>0.05; Fig. 4). The experiment was performed in triplicate.
Figure 4.

Effect of CEACAM-1 downregulation on the cell cycle distribution of colorectal cancer cells. Representative plots are shown from flow cytometry analysis following propidium iodide staining. (A) Untreated group. (B) Negative control siRNA group. (C) CEACAM-1-specific siRNA group. (D) Quantification of the results. CEACAM-1, carcinoembryonic antigen-related cell adhesion molecule 1; si, small interfering.
Discussion
To date, the function of CEACAM-1 expression in malignant tumors remains unclear. Previous studies have investigated CEACAM expression via either immunohistochemical or serum expression analyses (19,20). To elucidate its functional role in CRC, the present study assessed the effect of silencing CEACAM-1 expression on the viability and proliferation of a CRC cell line CEACAM-1 belongs to a diverse family of GPI-linked Igs that combine the structural features of the Ig superfamily with the functional properties of cadherins (21).
In the present study, CEACAM-1 expression was inhibited in the HCT-8 cell line via three custom-designed siRNA sequences, achieving a maximal reduction in CEACAM-1 mRNA expression by 61.3% compared with the control cells, and a similar reduction in CEACAM-1 protein production. Subsequently, it was determined that CEACAM-1 downregulation significantly inhibited cell proliferation and promoted cell apoptosis, suggesting that CEACAM-1 may have a potential clinical use in the treatment of CRC. Consistent with these results, a previous study has demonstrated that CEACAM-1 knockdown results in the decreased proliferation and migration of human pancreatic adenocarcinoma Pac 5061 cells (17). Thus, it can be concluded that CEACAM-1 is likely an important modulator of CRC.
Collectively, the results of the present study demonstrated that CEACAM-1 downregulation in CRC cells significantly inhibited cell proliferation and promoted apoptosis. Thus, CEACAM-1 may be a critical mediator of CRC cell growth and progression, and as a result, a promising potential target for novel CRC treatment strategies.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Key Project of Universities and Colleges in Henan, China (grant no. 15A310033).
Availability of data and materials
The analyzed datasets generated during the study are available from the corresponding author on reasonable request.
Authors' contributions
Z-MH designed, analyzed the experiments and wrote the manuscript. H-MH and Y-WS performed the experiments and co-wrote the manuscript.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
- 1.Chen W, Zheng R, Zeng H, Zhang S. The incidence and mortality of major cancers in China, 2012. Chin J Cancer. 2016;35:73. doi: 10.1186/s40880-016-0137-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Rosso T, Malvezzi M, Bosetti C, Bertuccio P, Negri E, La Vecchia C. Cancer mortality in Europe, 1970–2009: An age, period, and cohort analysis. Eur J Cancer Prev. 2016;27:88–102. doi: 10.1097/CEJ.0000000000000282. [DOI] [PubMed] [Google Scholar]
- 3.Cavallaro U, Christofori G. Cell adhesion in tumor invasion and metastasis: Loss of the glue is not enough. Biochim Biophys Acta. 2001;1552:39–45. doi: 10.1016/s0304-419x(01)00038-5. [DOI] [PubMed] [Google Scholar]
- 4.Pignatelli M, Vessey CJ. Adhesion molecules: Novel molecular tools in tumor pathology. Hum Pathol. 1994;25:849–856. doi: 10.1016/0046-8177(94)90002-7. [DOI] [PubMed] [Google Scholar]
- 5.Oettle H, Post S, Neuhaus P, Gellert K, Langrehr J, Ridwelski K, Schramm H, Fahlke J, Zuelke C, Burkart C, et al. Adjuvant chemotherapy with gemcitabine vs observation in patients undergoing curative-intent resection of pancreatic cancer: A randomized controlled trial. JAMA. 2007;297:267–277. doi: 10.1001/jama.297.23.2582-a. [DOI] [PubMed] [Google Scholar]
- 6.Rojas M, Fuks A, Stanners CP. Biliary glycoprotein, a member of the immunoglobulin supergene family, functions in vitro as a Ca2(+)-dependent intercellular adhesion molecule. Cell Growth Differ. 1990;1:527–533. [PubMed] [Google Scholar]
- 7.Beauchemin N, Draber P, Dveksler G, Gold P, Gray-Owen S, Grunert F, Hammarström S, Holmes KV, Karlsson A, Kuroki M, et al. Redefined nomenclature for members of the carcinoembryonic antigen family. Exp Cell Res. 1999;252:243–249. doi: 10.1006/excr.1999.4610. [DOI] [PubMed] [Google Scholar]
- 8.Arabzadeh A, Chan C, Nouvion AL, Breton V, Benlolo S, DeMarte L, Turbide C, Brodt P, Ferri L, Beauchemin N. Host-related carcinoembryonic antigen cell adhesion molecule 1 promotes metastasis of colorectal cancer. Oncogene. 2013;32:849–860. doi: 10.1038/onc.2012.112. [DOI] [PubMed] [Google Scholar]
- 9.Thöm I, Schult-Kronefeld O, Burkholder I, Schuch G, Andritzky B, Kastendieck H, Edler L, Wagener C, Bokemeyer C, Schumacher U, Laack E. Expression of CEACAM-1 in pulmonary adenocarcinomas and their metastases. Anticancer Res. 2009;29:249–254. [PubMed] [Google Scholar]
- 10.Thies A, Moll I, Berger J, Wagener C, Brümmer J, Schulze HJ, Brunner G, Schumacher U. CEACAM1 expression in cutaneous malignant melanoma predicts the development of metastatic disease. J Clin Oncol. 2002;20:2530–2536. doi: 10.1200/JCO.2002.05.033. [DOI] [PubMed] [Google Scholar]
- 11.Dango S, Sienel W, Schreiber M, Stremmel C, Kirschbaum A, Pantel K, Passlick B. Elevated expression of carcinoembryonic antigen-related cell adhesion molecule 1 (CEACAM-1) is associated with increased angiogenic potential in non-small-cell lung cancer. Lung Cancer. 2008;60:426–433. doi: 10.1016/j.lungcan.2007.11.015. [DOI] [PubMed] [Google Scholar]
- 12.Yoshikawa M, Morine Y, Ikemoto T, Imura S, Higashijima J, Iwahashi S, Saito YU, Takasu C, Yamada S, Ishikawa D, et al. Elevated preoperative serum CEA level is associated with poor prognosis in patients with hepatocellular carcinoma through the epithelial-mesenchymal transition. Anticancer Res. 2017;37:1169–1175. doi: 10.21873/anticanres.11430. [DOI] [PubMed] [Google Scholar]
- 13.Li N, Yang JY, Wang XY, Wang HT, Guan BX, Zhou CJ. Carcinoembryonic antigen-related cell adhesion molecule 1 is expressed and as a function histotype in ovarian tumors. Ann Diagn Pathol. 2016;20:7–12. doi: 10.1016/j.anndiagpath.2015.10.012. [DOI] [PubMed] [Google Scholar]
- 14.Gebauer F, Wicklein D, Horst J, Sundermann P, Maar H, Streichert T, Tachezy M, Izbicki JR, Bockhorn M, Schumacher U. Carcinoembryonic antigen-related cell adhesion molecules (CEACAM) 1, 5 and 6 as biomarkers in pancreatic cancer. PLoS One. 2014;9:e113023. doi: 10.1371/journal.pone.0113023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Li Y, Shively JE. CEACAM1 regulates Fas-mediated apoptosis in Jurkat T-cells via its interaction with β-catenin. Exp Cell Res. 2013;319:1061–1072. doi: 10.1016/j.yexcr.2013.02.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Liu GX, Xie Q, Zhou CJ, Zhang XY, Ma BL, Wang CQ, Wei FC, Qu X, Sun SZ. The possible roles of OPN-regulated CEACAM1 expression in promoting the survival of activated T cells and the apoptosis of oral keratinocytes in oral lichen planus patients. J Clin Immunol. 2011;31:827–839. doi: 10.1007/s10875-011-9552-4. [DOI] [PubMed] [Google Scholar]
- 17.Beauchemin N, Arabzadeh A. Carcinoembryonic antigen-related cell adhesion molecules (CEACAMs) in cancer progression and metastasis. Cancer Metastasis Rev. 2013;32:643–671. doi: 10.1007/s10555-013-9444-6. [DOI] [PubMed] [Google Scholar]
- 18.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 19.Simeone DM, Ji B, Banerjee M, Arumugam T, Li D, Anderson MA, Bamberger AM, Greenson J, Brand RE, Ramachandran V, Logsdon CD. CEACAM1, a novel serum biomarker for pancreatic cancer. Pancreas. 2007;34:436–443. doi: 10.1097/MPA.0b013e3180333ae3. [DOI] [PubMed] [Google Scholar]
- 20.Blumenthal RD, Leon E, Hansen HJ, Goldenberg DM. Expression patterns of CEACAM5 and CEACAM6 in primary and metastatic cancers. BMC Cancer. 2007;7:2. doi: 10.1186/1471-2407-7-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hauck W, Nédellec P, Turbide C, Stanners CP, Barnett TR, Beauchemin N. Transcriptional control of the human biliary glycoprotein gene, a CEA gene family member down-regulated in colorectal carcinomas. Eur J Biochem. 1994;223:529–541. doi: 10.1111/j.1432-1033.1994.tb19022.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The analyzed datasets generated during the study are available from the corresponding author on reasonable request.