

Abstract
Human mast cells (MCs) express a novel G protein-coupled receptor (GPCR), known as Mas-related G protein coupled receptor X2 (MRGPRX2). Activation of this receptor by a diverse group of cationic ligands such as neuropeptides (NPs), host defense peptides (HDPs) and FDA-approved drugs contributes to chronic inflammatory diseases and pseudo-allergic drug reactions. For most GPCRs, the extracellular domains and their associated transmembrane domains display the greatest structural diversity and are responsible for binding different ligands. The goal of the present study was to determine if naturally occurring missense variants within MRGPRX2’s extracellular/transmembrane domains contribute to gain or loss of function phenotype for MC degranulation in response to NPs (substance P; SP and hemokinin-1, HK-1), an HDP (human β-defensin-3, hBD3) and an FDA-approved cationic drug (bradykinin B2 receptor antagonist, icatibant). We have identified eight missense variants within MRGPRX2’s extracellular/transmembrane domains from publically available exome-sequencing databases. We investigated the ability of MRGPRX2 ligands to induce degranulation in RBL-2H3 (RBL) cells individually expressing these naturally occurring MRGPRX2 missense variants. Using stable and transient transfections, we found that all variants express in RBL cells. However, four natural MRGPRX2 variants, G165E (rs141744602), D184H (rs372988289), W243R (rs150365137) and H259Y (rs140862085) failed to respond to any of the ligands tested. Thus, diverse MRGPRX2 ligands utilize common sites on the receptor to induce MC degranulation. These findings have important clinical implications for MRGPRX2 and MC-mediated pseudo-allergy and chronic inflammatory diseases.
Keywords: Mast cell, MRGPRX2, missense variants, Substance P, Hemokinin-1, Human β-defensin-3, Icatibant
Introduction
In addition to FcεRI, mast cells (MCs) express a novel seven transmembrane (7TM) domain receptor known as MAS-related G protein-coupled receptor X2 (MRGPRX2) (1, 2). Emerging evidence suggests that MRGPRX2 contributes to the pathogenesis of a number of chronic inflammatory diseases and is responsible for injection site reactions to opioids and other FDA-approved cationic drugs (3–9). For example, expression of MRGPRX2 is upregulated in skin MCs of patients with chronic urticaria when compared to MCs of normal subjects (7). Furthermore, MRGPRX2-mediated MC activation by the neuropeptide, substance P (SP) contributes to the pathogenesis of chronic urticaria. Hemokinin-1 (HK-1) is a novel neuropeptide, which is released from antigen/IgE activated MCs and human bronchial cells and causes contraction of human bronchi ex vivo (10, 11). We recently reported that MRGPRX2 is expressed at low level in non-asthmatic lung MCs but its expression is significantly upregulated in asthmatic lung MCs and demonstrated that HK-1 causes MC degranulation via this receptor (12).
Human β-defensin-3 (hBD3) and MRGPRX2-expressing MCs are present in healthy gingiva and their levels are elevated in patients with chronic periodontitis (6, 13). Furthermore, hBD3 causes MC degranulation via MRPGRX2 (14). Icatibant, a bradykinin B2 receptor antagonist used for the treatment of hereditary angioedema, promotes MC degranulation via MRPGRX2 and causes injection-site erythema and swelling in nearly every patient (15, 16). These findings suggest that activation of MRGPRX2 by a diverse group of ligands such as SP, HK-1, hBD3 and icatibant contribute to the pathogenesis of chronic urticaria, asthma, periodontitis and pseudo-allergic reactions. Modeling and mutagenesis studies with SP and various opioid ligands led to the suggestion that different agonists interact with different amino acid residues in MRGPRX2’s predicted ligand-binding pocket to induce MC degranulation (17, 18). However, the sites of interaction of HK-1, hBD3 and icatibant on MRGPRX2 for MC degranulation have not been determined.
Recent crystallography data obtained for a number of GPCRs and comparison of sequence homology have provided prediction regarding the regions of GPCRs that are involved in ligand binding and G protein coupling. The 7TM bundles are connected by three extracellular loops (ECL1, ECL2 and ECL3) and three intracellular loops (ICL1, ICL-2 and ICL-3). The extracellular part also includes the N-terminus (N-term) and the intracellular (IC) part includes the helix VIII and a C-terminal sequence. GPCRs can be divided into modules. The EC and their closest TM regions (see Fig. 1A) have the greatest structural diversity and are responsible for the binding of diverse ligands. By contrast, the IC and its closest TM regions are responsible for G protein coupling and downstream signaling (19).
Figure 1. Naturally occurrence MRGPRX2 variants identified from the NHLBI GO ESP database.

(A): Serpentine diagram of the secondary structure of human MRGPRX2. Each circle represents amino acid residue with one letter code. Solid background denotes the eight naturally occurring missense variants used in the present study. Extracellular (ECL), intracellular (ILC) and transmembrane (TM) domains are shown. (B): Amino acid change for each MRGPRX2 variant, PolyPhen-2 score, Minor allele frequency (MAF) and CADD score are shown.
We have identified eight naturally occurring missense variants within MRGPRX2’s extracellular and transmembrane domains from publically available databases; NHLBI-GO-ESP, 1000 Genomes and ExAC. The goal of the present study was to determine if any of these variants display gain or loss of function phenotype for MC degranulation in response to NPs (substance P; SP and hemokinin-1, HK-1), an HDP (hBD3) and a cationic drug (bradykinin B2 receptor antagonist, icatibant). The data presented herein provide novel insights on the impact of single naturally occurring mutation on MRGPRX2 activation by a diverse group of cationic ligands and have important implications for MC-mediated health and disease.
Materials and Methods
Materials
All cell culture reagents and Indo-1 were purchased from Invitrogen (Gaithersburg, MD). Substance P, Fluorescent-labeled substance P (FAM-SP), hBD3 and icatibant were purchased from Anaspec (Fremont, CA). Human hemokinin-1 was purchased from Alpha Diagnostic (San Antonio, TX). MRGPRX2 plasmid encoding hemagglutinin (HA)-tagged human MRGPRX2 in pReceiver-MO6 vector was obtained from GeneCopoeia (Rockville, MD). Amaxa transfection kit (Kit V) was purchased from Lonza (Gaithersburg, MD). PE-Anti human MRGPRX2 was obtained from Biolegend (San Diego, CA). QuikChange II Site-directed mutagenesis kit was purchased from Agilent Genomics (Santa Clara, CA).
Cell culture
Rat basophilic leukemia (RBL-2H3; RBL) cells were maintained as monolayer cultures in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% FBS, L-glutamine (2 mM), penicillin (100 IU/ml) and streptomycin (100 μg/ml) (20).
Site-directed mutagenesis
Quick II change site-directed mutagenesis kit (Agilent) was used to generate MRGPRX2 variants. DNA sequencing was performed to confirm the nucleotide sequences for each construct. The forward and reverse primers were used for each variant are listed below.
N16H: Forward: 5′-GGGCTTGGTCATTTCCATGCACTGTTGTACTTTCTGT-3′ Reverse: 5′-ACAGAAAGTACAACAGTGCATGGAAATGACCAAGCCC-3′ L31V: Forward: 5′-GAAGACCGGGATCACGGTCTCCTTGCCAC-3′ Reverse: 5′-GTGGCAAGGAGACCGTGATCCCGGTCTTC-3′ V43I: Forward: 5′-TTTCCTACCAGCCCGATCAGGGCAATGAAAAGG-3′Reverse: 5′-CCTTTTCATTGCCCTGATCGGGCTGGTAGGAAA-3′ F78L: Forward: 5′-GGAAGCAGAGTAAGAGGAAGTCGGCCCCG-3′ Reverse: 5′-CGGGGCCGACTTCCTCTTACTCTGCTTCC-3′ G165E: Forward: 5′-GCTGAGCATCTTGGAAGAGAAGTTCTGTGGCTTCT-3′ Reverse: 5′-AGAAGCCACAGAACTTCTCTTCCAAGATGCTCAGC-3′ D184H: Forward: 5′-CTCTGGTTGGTGTCAGACATTTCATTTCATCACTGC-3′ Reverse: 5′-GCAGTGATGAAATGAAATGTCTGACACCAACCAGAG-3′ W243R: Forward: 5′-CTGCCCTTTGGCATTCAGCGGTTCCTAATATTATGGAT-3′ Reverse: 5′-ATCCATAATATTAGGAACCGCTGAATGCCAAAGGGCAG-3′ H259Y: Forward: 5′-AAGGATTCTGATGTCTTATTTTGTTATATTCATCCAGTTTCAGTTGTC-3′ Reverse: 5′GACAACTGAAACTGGATGAATATAACAAAATAAGACATCAGAATCCTT-3′
Transfection of RBL cells and Flow cytometry
Cells (1. 5 x106) were transfected with plasmids (1.5 μg) encoding MRGPRX2 or its missense variants using the Amaxa Nucleofector device and Amaxa kit V according to the manufacturer’s protocol. For stable transfection, cells were cultured in the presence of G-418 (1 mg/ml) and used within one month of transfection (21). For transient transfection, cells were used within 16–20 hours after transfection. To detect MRGPRX2 expression, cells (0.5 x 106) were incubated with PE-conjugated anti-MRGPRX2 antibody, washed in FACS buffer, fixed and analyzed on a BD LSR II flow cytometer. For FAM-SP binding RBL-2H3 cells stably overexpressing MRGPRX2 (RBL-MRGPRX2) were used (12). Untransfected RBL cells and RBL-MRGPRX2 cells (0.5 x 106) were incubated for with FAM-SP (1 μM, 60 min, 4°C). The cells were washed twice with FACS buffer and analyzed by flow cytometry.
Calcium mobilization
Ca2+ mobilization was determined as described previously (21). Briefly, cells (2 x 106) were loaded with 1 μM indo-1 AM for 30 min at room temperature. Cells were then washed and resuspended in 1.5 ml of HEPES-buffered saline. The cells were then stimulated with SP and Ca2+ mobilization was determined using a Hitachi F-2500 spectrophotometer with an excitation wavelength of 355 nm and an emission wavelength of 410 nm.
Degranulation
Cells (5 x 104) were seeded into 96-well plates in a total volume of 50 μl HEPES buffer containing 0.1% bovine serum albumin (BSA) and exposed to ligands for 30 min. Cells without treatment were designated as controls. For total β-hexosaminidase release, unstimulated cells were lysed in 50 μl of 0.1% Triton X-100. Aliquots (20 μl) of supernatants or cell lysates were incubated with 20 μl of 1 mM p-nitrophenyl-N-acetyl-β-D-glucosamine for 1 hr at 37°C. The reaction was stopped by adding 250 μl of a 0.1 M Na2CO3/0.1 M NaHCO3 buffer and absorbance was measured at 405 nm (21).
Results
Identification of D184H and G165E as loss of function MRGPRX2 variants for SP, hemokinin-1, hBD3 and icatibant in stably transfected RBL-2H3 cells
To identify naturally occurring missense variants in the extracellular and transmembrane domains of MRGPRX2, we searched publically available databases, NHLBI-GO-ESP, 1000 Genomes and ExAC. We found eight targets in the MRGPRX2 gene with missense mutations (Fig. 1A). Amino acid change for each mutation is shown in Fig. 1B. Data from a computational protein prediction program, Polymorphism Phenotyping v2 (PolyPhen-2), under the HumDiv model on a scale of 0 (benign) to 1 (damaging) are shown in are shown in Fig. 1B (22). The G165E and D184H variants have the lowest minor allele frequency (average MAF, <0.01%) whereas the N16H variant has the highest MAF (8.95%). Combined annotation-dependent depletion (CADD) is an in silico tool used for scoring the deleteriousness of single nucleotide variants as well as insertion/deletion variants in the human genome (23). A CADD score of >10 indicates that a variant may be deleterious. As shown in Fig. 1B, all eight MRGPRX2 variants targeted for the present study have CADD scores of <10. Thus, the combined in silico analyses suggest that although some of the MRGPRX2 variants may be damaging (Polyphen2 score >0.5), these mutations are rare (MAF <1%) and may not be associated with pathogenicity (CADD <10) (Fig. 1).
Studies on the effects of missense or other mutations on GPCR functions are routinely conducted with transiently transfected HEK293 cells (17, 18, 24–26). However, our previous studies on the regulation of MRGPRX2 have been performed with stably transfected rodent mast cell line (RBL-2H3 cell) that does not endogenously express the receptor (6, 12). To determine if any of the MRGPRX2 variants shown in Fig. 1 display gain or loss of function for MC activation, we first generated stable transfectants in RBL cells. The stable transfection procedure involves cDNA nucleofection followed by culturing cells in the presence of selection marker G418. We were quite surprised to find cells expressing three mutants (N16H, W243R or H259Y) did not survive the G418 selection procedure. We found that 5 days after the start of G418 selection, cells expressing the wild-type (WT) receptor were almost fully covering the surface of the tissue culture dish but only few cells could be detected in N16H, W243R or H259Y transfectants. Triton X-lysed cells were used to measure total β-hexosaminidase content as an assay to quantitate cell number. As shown in Fig. 2A, unlike the situation with cells expressing the WT receptor, we could not detect any β-hexosaminidase in transfectants with cDNA encoding N16H, W243R or H259Y (Fig. 2A). We therefore focused our initial studies on variants L31V, V43I, F78L, G165E and D184H, which stably express on the cell surface similar to the WT receptor (Fig 2B). Although MRGPRX2 is activated by multiple ligands, SP is probably the most well characterized (1, 2, 5, 7, 16, 18). We therefore tested the effects of SP on Ca2+ mobilization and degranulation in cells expressing these variants. We found that the variants L31V, V43I and F78L responded normally to SP for Ca2+ mobilization and degranulation (Fig. 2C and D). By contrast, when compared to the WT receptor, Ca2+ mobilization and degranulation to SP were substantially inhibited in cells expressing G165E and D184H variants (Fig. 2C and 2D), despite normal cell surface expression (Fig. 2B). These findings are consistent with higher polyPhen2 score for G165E and D184H when compared to L31A, V43I and F78L variants (Fig. 1B).
Figure 2. Effects of naturally occurring MRGPRX2 mutations (L31I, V43I, F78L, G165E and D184H) on cell surface expression, SP-induced Ca2+ mobilization and degranulation in stably transfected RBL cells.

(A): Cells were transfected with cDNA encoding wild-type (WT), N16H, W243R, or H253Y variant, transferred to 24 well plate and G418 was added to the culture medium 16 h after transfection. After 5 days, non-adherent cells were removed, adherent cells were lysed and total β-hexosaminidase content was determined. (B): Flow cytometry was performed with PE-anti-MRGPRX2 antibody to determine cell surface expression of WT and variants in stably transfected RBL cells. Representative histograms for WT/Variant (thick line) and control untransfected cells (thin line) are shown. (C): Cells expressing WT and MRGPRX2 variants were loaded with Indo-1 and intracellular Ca2+ mobilization in response to SP (1 μM) was determined. Data shown are representative of three independent experiments. (D): Cells were exposed to buffer (control) or SP (0. 3μM and 1 μM) for 30 min and β-hexosaminidase release was determined. All data points are expressed as mean ± SEM of three experiments performed in triplicate. Statistical significance was determined by two-tailed unpaired t-Test. *** indicates P value<.001, and ** indicates P value <.01.
Reddy et al., (17) recently showed that a point mutant of MRGPRX2 (E164R) was resistant to SP for Ca2+ mobilization but responded normally to a host defense peptide, LL-37. Based on this finding, it was proposed that different amino acid residues on MRGPRX2 are responsible for binding to different ligands. We therefore tested the effects of naturally occurring missense mutations (L31A, V43I, F78L, G165E and D184H) on degranulation in response to HK-1, hBD3 and icatibant. As shown in Fig. 3, A–C, cells expressing L31V, V43I or F78L variant responded to all ligands tested for degranulation similar in extent to the WT receptor. By contrast, cells expressing G165E or D184H variant were resistant to HK-1, hBD3 and icatibant for degranulation. These findings suggest that SP, HK-1, hBD3 and icatibant all interact with the same amino acids on MRGPRX2’s 4th and 5th TM domains (Fig. 1) to induce MC degranulation.
Figure 3. Effects of naturally occurring MRGPRX2 mutations (L31I, V43I, F78L, G165E and D184H) on HK-1, hBD3 and icatibant-induced degranulation in stably transfected RBL cells.
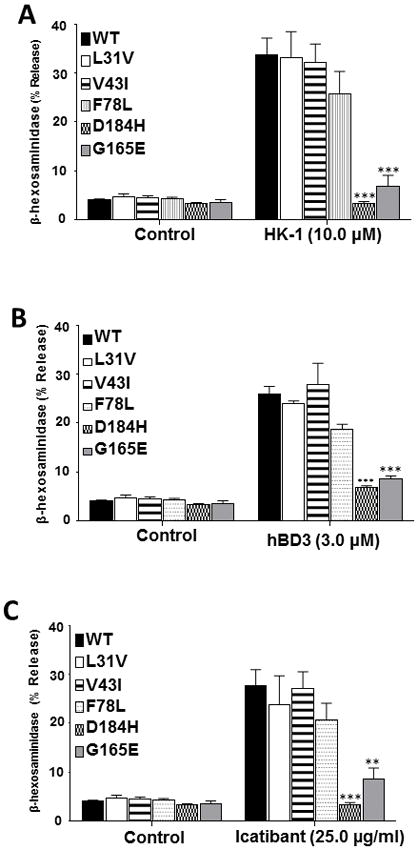
Cells stably expressing WT and MRGPRX2 variants were exposed to buffer (control) or stimulated with (A) hemokinin-1 (HK-1, 10 μM), (B) hBD3 (3 μM) or (C) icatibant (25 μg/ml) for 30 min and β-hexosaminidase release was determined. All the points expressed as a mean ± SEM of three experiments in triplicate. Statistical significance was determined by two tailed unpaired t-Test. *** indicates P value<.001, ** indicates and P value <.01.
Our next goal was to perform binding studies to determine if reduced MC activation in missense variants correlates with correspondingly reduced ligand binding to MRGPRX2. Of the agonists used in this study, only SP is available as a fluorescent-labeled conjugate (FAM-SP). Our initial goal was to perform flow cytometry analysis to determine if FAM-SP binds to RBL cells stably expressing MRGPRX2 and to test if this binding could be blocked by unlabeled HK-1, hBD3 or icatibant. For these studies, we used RBL cells stably overexpressing MRGPRX2 (RBL-MRGPRX2). We confirmed cell surface MRGPRX2 expression by flow cytometry using PE-conjugated anti-MRGPRX2 antibody (Fig. 4A). To validate the functional activity of FAM-SP for MRGPRX2, we compared its ability to induce degranulation in untransfected RBL and RBL-MRGPRX2 cells. As shown in Fig. 4B, both unlabeled SP and FAM-SP induced degranulation in RBL-MRGPRX2 cells but not in untransfected cells, validating the specificity of FAM-SP for the receptor. However, flow cytometry experiment demonstrated that FAM-SP interact equally well with untransfected RBL and RBL-MRGPRX2 cells (Fig. 4C). These findings suggest that the amphipathic nature of SP facilitates a strong interaction with the plasma membrane even in the absence of MRGPRX2, thus making it difficult to perform the proposed binding studies.
Figure 4. FAM-SP-induced degranulation and binding in RBL cells overexpressing MRGPRX2.
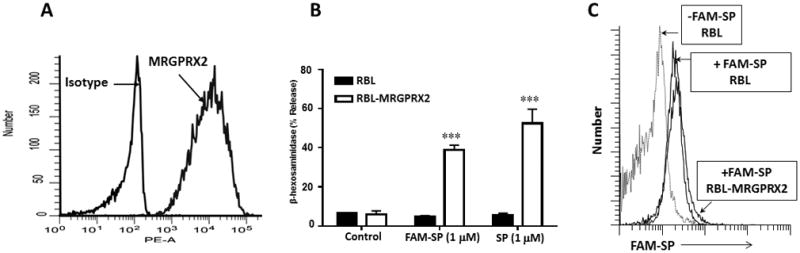
(A): RBL cells stably overexpressing MRGPRX2 (RBL-MRGPRX2) were exposed to PE-conjugated anti-MRGPRX2 or PE-conjugated isotype matched antibody and cell surface receptor expression was determined by flow cytometry. (B): RBL and RBL-MRGPRX2 cells were exposed to buffer (control), SP or FAM-SP for 30 min and β-hexosaminidase release was determined. (C): RBL or RBL-MRGPRX2 cells were exposed to FAM-SP (1 μM, 60 min, 4°C), washed and fluorescence was determined by flow cytometry. Data presented for flow are representative of 3 similar experiments and degranulation data are expressed as mean ± SEM of three experiments in triplicate. Statistical significance was determined by two tailed unpaired t-Test. *** indicates P value<.001
Identification of W243R and H259Y as loss of function MRGPRX2 variants for SP, HK-1, hBD3 and icatibant in transiently transfected RBL cells
The low PolyPhen-2 score for the MRGPRX2 variant H259Y (Fig. 1B) indicates that this mutation is benign. Thus, our inability to generate a stable transfectant expressing this variant in RBL cells was surprising. We therefore, generated transient transfectant in RBL cells expressing H259Y. We also performed similar studies with two other variants with high PolyPhen2 score (N16H; 0.937 and W243R; 0.760) that did not survive the stable transfection procedure (Fig. 1A). We found that all three variants expressed on the surface of RBL cells (Fig. 5A). The N16H variant responded to SP for Ca2+ mobilization and degranulation similar to the WT receptor (Fig. 5B and C). However, W243R and H259Y variants were resistant to SP-induced Ca2+ mobilization and degranulation (Fig. 5B and C). HK-1, hBD3 and icatibant induced normal degranulation in cells transiently expressing N16H but they failed to activate this response in cells expressing W243R or H259Y variant (Fig. 6).
Figure 5. Effects of naturally occurring MRGPRX2 mutations (N16H, W243R and H259Y) on cell surface expression, Ca2+ mobilization and SP-induced degranulation in transiently transfected RBL cells.

(A): Flow cytometry was performed using PE-anti-MRGPRX2 antibody to determine cell surface expression of wild-type (WT) and variants in transiently transfected in RBL cells. Representative histograms for WT/Variant (thick line) and control untransfected cells (thin line) are shown. (B): Cells expressing WT and MRGPRX2 variants were loaded with Indo-1 and intracellular calcium mobilization in response to SP (1 μM) was determined. Data shown are representative of three independent experiments. (C): Cells were exposed to buffer (control) or SP (1 μM) for 30 min and β-hexosaminidase release was determined. All data points are expressed as mean ± SEM of three experiments performed in triplicate. Statistical significance was determined by two tailed unpaired t-Test. ** indicates P value <.01.
Figure 6. Effects of MRGPRX2 mutations (N16H, W243R and H259Y) on degranulation in response to HK-1, hBD3 and icatibant.

RBL cells transiently expressing WT, N16H, W243R and H259Y variants were stimulated with (A) HK-1 (10 μM), (B) hBD3 (3 μM) and (C) icatibant (25 μg/ml) and β-hexosaminidase release was determined. All the points expressed as a mean ± SEM of three experiments in triplicate. Statistical significance was determined by two tailed unpaired t-Test. *** indicates P value<.001, ** indicates P value <.01 and * indicates P value <.05.
Discussion
The unique features of MRGPRX2 that distinguish it from other GPCRs are its predominant expression on human MCs and its activation by a diverse group of cationic ligands (6, 7, 12, 16). Emerging evidence suggests that MRGPRX2 contributes to pseudo-allergic drug reactions and a number of chronic inflammatory diseases (16, 27, 28). Although activation of MRGPRX2 or FcεRI on MCs elicits intracellular Ca2+ mobilization and comparable MC degranulation, MRGPRX2 stimulation triggers little to no cytokine or prostaglandin E2 generation (16, 29, 30). Thus, the effects on MRGPRX2 on MC-mediated disorders likely reflects Ca2+ mobilization and degranulation. In the present study, we have utilized eight naturally occurring missense MRGPRX2 variants and identified rare mutations that display loss of function phenotype for MC degranulation in response to ligands that participate in pseudo-allergy (icatibant) (16), itch/chronic urticaria (SP) (5, 7), asthma (HK-1) (10–12) and periodontitis (hBD3) (6).
Molecular modeling and docking approaches have recently been used to identify Glu164 (E164) and Asp184 (D184) in MRPGRX2’s 4th and 5th TM domains as the negatively charged residues that make ionic contact with cationic opioid ligands (18). Accordingly, E164Q or D184N substitution that retains the steric property of the wild-type residue but removes the negative charge resulted in loss of receptor activation by dextromethorphan, morphine and related opioids ligands. These findings suggest that both Glu164 and Asp184 are important for the binding of opioids to MRGPRX2 and that removal of one negatively charged residue results in loss of ligand-receptor interaction. However, MRGPRX2 activation by a metabolite of an endogenous opioid peptide, dynorphin A (1–13), is lost in the D184N but not the E164Q mutation (18). Modeling studies predict that Arg7 and Phe4 of dynorphin interact with Asp184 but not Glu164. These findings suggest that one or both anionic Glu164 and Asp184 participate in the binding of cationic opioids and endogenous opioid peptide metabolites. Using publically available web portals, Reddy et al., (17) also predicted that SP-binding pocket in MRGPRX2 consists of a number of structurally conserved hydrophilic residues along with a buried glutamic acid residue (Glu164, E164). Accordingly, replacement of Glu164 with a positively charged Arg (E164R) results in loss MRGPRX2 activation by SP. However, this mutant respond normally to the host defense peptide, LL-37 (17). These findings suggest that different ligands interact with different amino acid residues on MRGPRX2’s predicted ligand binding pocket to induce MC degranulation (17, 18).
The goal of the present study was to use publically available databases to identify naturally occurring missense MRGPRX2 variants that display gain or loss of function phenotype for MC degranulation by a diverse group of cationic ligands. There are a number of interesting features regarding the rare MRGPRX2 variant G165E (MAF; <0.01%) that warrant discussion. First, it is present within the predicted binding pocket for opioids and SP, as determined from modeling studies (17, 18). Second, although we were unsuccessful in demonstrating specific binding of FAM-SP to RBL cells stably overexpressing MRGPRX2 due to high background binding, it is likely that this mutation interferes with SP’s binding to MRGPRX2. This contention was supported by the finding that cells expressing G165E variant did not respond to SP for Ca2+ mobilization despite normal cell surface expression. Third, and most importantly, none of the other MRGPRX2 ligands tested caused degranulation in cells expressing this variant. It is noteworthy that G165E mutation results in the replacement of an aliphatic side chain with a positively charged residue next to a negatively charged amino (Glu164, E164) required for ionic interaction with SP and opioids (17, 18). It is therefore likely that the presence two adjacent negatively charged side chains (E164, E165) in the variant G165E interferes with the integrity of the binding pocket and thus preventing MC activation by SP and all other cationic ligands used in this study.
Studies by Reddy et al., (17) did not consider Asp184 as a possible site for SP interaction with MRGPRX2 despite the fact this amino acid is critical for receptor activation of by all opioid ligands tested, including dynorphin A (18). It is interesting to note that the naturally occurring D184H variant results in the switching of a negatively charged side chain to a positively charged one. An important finding of the present study was that none of the MRGPRX2 agonists tested stimulated degranulation in RBL cells stably expressing either G165E or D184H variant. These results suggest that SP and opioid binding pocket identified within MRGPRX2’s 4th and 5th TM domains (17, 18) is shared by all MRGPRX2 agonists that induce MC degranulation and that G165E or D184H mutation disrupts the integrity of the binding pocket preventing receptor activation.
We found that two additional mutations, W243R and H259Y, in the 6th and 7th TM domains outside MRGPRX2’s predicted ligand binding pocket also rendered the receptor unresponsive to all ligands tested. The mechanism by which these mutations lead to loss of degranulation is unknown. Interestingly, we found that while cells expressing these variants do not survive the stable transfection procedure they express normally in transiently transfected cells. It is therefore possible that replacement of the bulky Trp with a positively charged Arg in variant W243R and His with Tyr in variant H269Y influences both the receptor’s expression status and its ability to interact with diverse ligands. In future studies, it will be interesting to determine the MC status of individuals harboring these mutations and their responsiveness to MGRPRX2 ligands.
In summary, we have shown that naturally occurring rare MRGPRX2 variants with single amino acid substitution in the receptor’s predicted ligand binding pocket (G165E and D184H) renders it unresponsive to SP, HK-1, hBD3 and icatibant for MC degranulation. Although the naturally occurring variants W243R and H259Y in the receptor’s 6th and 7th TM domain also rendered the receptor unresponsive to all ligands tested for degranulation, the mechanisms of their actions are unknown and remain to be determined. An important clinical implication of the present study is that individuals harboring any of the missense MRGPRX2 mutation (G165E, D184H, W243R or H259Y) may be protected from MC-mediated drug-induced pseudo-allergy and chronic inflammatory diseases such as itch, chronic urticaria, asthma and periodontitis. A goal of our future studies is to determine the responsiveness of MCs isolated from these individuals to MRGPRX2 ligands and to test whether these individuals develop skin reactions to pseudo-allergic drugs.
Acknowledgments
This work was supported by NIH grant R01-AI124182 to HA.
We thank the FACS core facility of the School of Dental Medicine, University of Pennsylvania for flow cytometry data acquisition and analysis. We are also grateful to the Institute for Biomedical Informatics (IBI) Bioinformatics Core, Perelman School of Medicine University of Pennsylvania for providing CADD scores for all MRGPRX2 variants utilized in this study. We thank Dr. Saptarshi Roy for critical review of this manuscript.
Abbreviations
- MC
mast cell
- GPCR
G protein coupled receptor
- MRGPRX2
Mas-related G protein coupled receptor X2
- SP
substance P
- HK-1
hemokinin-1
- hBD3
human β-defensin 3
References
- 1.Tatemoto K, Nozaki Y, Tsuda R, Konno S, Tomura K, Furuno M, Ogasawara H, Edamura K, Takagi H, Iwamura H, Noguchi M, Naito T. Immunoglobulin E-independent activation of mast cell is mediated by Mrg receptors. Biochem Biophys Res Commun. 2006;349:1322–1328. doi: 10.1016/j.bbrc.2006.08.177. [DOI] [PubMed] [Google Scholar]
- 2.Subramanian H, Kashem SW, Collington SJ, Qu H, Lambris JD, Ali H. PMX-53 as a dual CD88 antagonist and an agonist for Mas-related gene 2 (MrgX2) in human mast cells. Mol Pharmacol. 2011;79:1005–1013. doi: 10.1124/mol.111.071472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Azimi E, Lerner EA. Implications of MRGPRX2 in human and experimental cardiometabolic diseases. Nat Rev Cardiol. 2017;14:124. doi: 10.1038/nrcardio.2016.212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Azimi E, Reddy VB, Lerner EA. Brief communication: MRGPRX2, atopic dermatitis and red man syndrome. Itch (Phila) 2017:2. doi: 10.1097/itx.0000000000000005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Azimi E, V, Reddy B, Pereira PJS, Talbot S, Woolf CJ, Lerner EA. Substance P activates Mas-related G protein-coupled receptors to induce itch. J Allergy Clin Immunol. 2017;140:447–453. doi: 10.1016/j.jaci.2016.12.980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gupta K, Idahosa C, Roy S, Lee D, Subramanian H, Dhingra A, Boesze-Battaglia K, Korostoff J, Ali H. Differential Regulation of Mas-Related G Protein-Coupled Receptor X2-Mediated Mast Cell Degranulation by Antimicrobial Host Defense Peptides and Porphyromonas gingivalis Lipopolysaccharide. Infect Immun. 2017;85:1–13. doi: 10.1128/IAI.00246-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Fujisawa D, Kashiwakura J, Kita H, Kikukawa Y, Fujitani Y, Sasaki-Sakamoto T, Kuroda K, Nunomura S, Hayama K, Terui T, Ra C, Okayama Y. Expression of Mas-related gene X2 on mast cells is upregulated in the skin of patients with severe chronic urticaria. J Allergy Clin Immunol. 2014;134:622–633. doi: 10.1016/j.jaci.2014.05.004. [DOI] [PubMed] [Google Scholar]
- 8.Okamura Y, Mishima S, Kashiwakura JI, Sasaki-Sakamoto T, Toyoshima S, Kuroda K, Saito S, Tokuhashi Y, Okayama Y. The dual regulation of substance P-mediated inflammation via human synovial mast cells in rheumatoid arthritis. Allergol Int. 2017;66S:S9–S20. doi: 10.1016/j.alit.2017.03.002. [DOI] [PubMed] [Google Scholar]
- 9.Reddy VB, Azimi E, Chu L, Lerner EA. Mas-Related G-Protein Coupled Receptors and Cowhage-Induced Itch. J Invest Dermatol. 2018;138:461–464. doi: 10.1016/j.jid.2017.05.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Grassin-Delyle S, Naline E, Buenestado A, Risse PA, Sage E, Advenier C, Devillier P. Expression and function of human hemokinin-1 in human and guinea pig airways. Resp Res. 2010;11:1–12. doi: 10.1186/1465-9921-11-139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sumpter TL, Ho CH, Pleet AR, Tkacheva OA, Shufesky WJ, Rojas-Canales DM, Morelli AE, Larregina AT. Autocrine hemokinin-1 functions as an endogenous adjuvant for IgE-mediated mast cell inflammatory responses. J Allergy Clin Immunol. 2015;135:1019–1030. doi: 10.1016/j.jaci.2014.07.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Manorak W, Idahosa C, Gupta K, Roy S, Panettieri R, Jr, Ali H. Upregulation of Mas-related G Protein coupled receptor X2 in asthmatic lung mast cells and its activation by the novel neuropeptide hemokinin-1. Resp Res. 2018;19:1–5. doi: 10.1186/s12931-017-0698-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Jin L. An update on innate defense molecules of human gingiva. Periodontol 2000. 2011;56:125–142. doi: 10.1111/j.1600-0757.2010.00364.x. [DOI] [PubMed] [Google Scholar]
- 14.Subramanian H, Gupta K, Lee D, Bayir AK, Ahn H, Ali H. beta-Defensins activate human mast cells via Mas-related gene X2. J Immunol. 2013;191:345–352. doi: 10.4049/jimmunol.1300023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lumry WR, Li HH, Levy RJ, Potter PC, Farkas H, Moldovan D, Riedl M, Li H, Craig T, Bloom BJ, Reshef A. Randomized placebo-controlled trial of the bradykinin B(2) receptor antagonist icatibant for the treatment of acute attacks of hereditary angioedema: the FAST-3 trial. Ann Allergy Asthma Immunol. 2011;107:529–537. doi: 10.1016/j.anai.2011.08.015. [DOI] [PubMed] [Google Scholar]
- 16.McNeil BD, Pundir P, Meeker S, Han L, Undem BJ, Kulka M, Dong X. Identification of a mast-cell-specific receptor crucial for pseudo-allergic drug reactions. Nature. 2015;519:237–241. doi: 10.1038/nature14022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Reddy VB, Graham TA, Azimi E, Lerner EA. A single amino acid in MRGPRX2 necessary for binding and activation by pruritogens. J Allergy Clin Immunol. 2017;140:1726–1728. doi: 10.1016/j.jaci.2017.05.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lansu K, Karpiak J, Liu J, Huang XP, McCorvy JD, Kroeze WK, Che T, Nagase H, Carroll FI, Jin J, Shoichet BK, Roth BL. In silico design of novel probes for the atypical opioid receptor MRGPRX2. Nat Chem Biol. 2017;13:529–536. doi: 10.1038/nchembio.2334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Katritch V, Cherezov V, Stevens RC. Diversity and modularity of G protein-coupled receptor structures. Trends Pharmacol Sci. 2012;33:17–27. doi: 10.1016/j.tips.2011.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ali H, Richardson RM, Tomhave ED, DuBose RA, Haribabu B, Snyderman R. Regulation of stably transfected platelet activating factor receptor in RBL-2H3 cells. Role of multiple G proteins and receptor phosphorylation. J Biol Chem. 1994;269:24557–24563. [PubMed] [Google Scholar]
- 21.Subramanian H, Gupta K, Guo Q, Price R, Ali H. Mas-related gene X2 (MrgX2) is a novel G protein-coupled receptor for the antimicrobial peptide LL-37 in human mast cells: resistance to receptor phosphorylation, desensitization, and internalization. J Biol Chem. 2011;286:44739–44749. doi: 10.1074/jbc.M111.277152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Adzhubei I, Jordan DM, Sunyaev SR. Predicting functional effect of human missense mutations using PolyPhen-2. Curr Protoc Hum Genet. 2013;Chapter 7(Unit7):20. doi: 10.1002/0471142905.hg0720s76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kircher M, Witten DM, Jain P, O’Roak BJ, Cooper GM, Shendure J. A general framework for estimating the relative pathogenicity of human genetic variants. Nat Genet. 2014;46:310–315. doi: 10.1038/ng.2892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hauser AS, Chavali S, Masuho I, Jahn LJ, Martemyanov KA, Gloriam DE, Babu MM. Pharmacogenomics of GPCR Drug Targets. Cell. 2018;172:41–54. doi: 10.1016/j.cell.2017.11.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Doyle JR, Lane JM, Beinborn M, Kopin AS. Naturally occurring HCA1 missense mutations result in loss of function: potential impact on lipid deposition. J Lipid Res. 2013;54:823–830. doi: 10.1194/jlr.M034660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Fischer L, Wilde C, Schoneberg T, Liebscher I. Functional relevance of naturally occurring mutations in adhesion G protein-coupled receptor ADGRD1 (GPR133) BMC Genomics. 2016;17:609. doi: 10.1186/s12864-016-2937-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Subramanian H, Gupta K, Ali H. Roles of Mas-related G protein-coupled receptor X2 on mast cell-mediated host defense, pseudoallergic drug reactions, and chronic inflammatory diseases. J Allergy Clin Immunol. 2016;138:700–710. doi: 10.1016/j.jaci.2016.04.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ali H. Emerging Roles for MAS-Related G Protein-Coupled Receptor-X2 in Host Defense Peptide, Opioid, and Neuropeptide-Mediated Inflammatory Reactions. Adv Immunol. 2017;136:123–162. doi: 10.1016/bs.ai.2017.06.002. [DOI] [PubMed] [Google Scholar]
- 29.Gaudenzio N, Sibilano R, Marichal T, Starkl P, Reber LL, Cenac N, McNeil BD, Dong X, Hernandez JD, Sagi-Eisenberg R, Hammel I, Roers A, Valitutti S, Tsai M, Espinosa E, Galli SJ. Different activation signals induce distinct mast cell degranulation strategies. J Clin Invest. 2016;126:3981–3998. doi: 10.1172/JCI85538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Karhausen J, Abraham SN. How mast cells make decisions. J Clin Invest. 2016;126:3735–3738. doi: 10.1172/JCI90361. [DOI] [PMC free article] [PubMed] [Google Scholar]