

Abstract
The luciferase isolated from the firefly Photinus pyralis (Ppy) catalyzes a two-step reaction that results in the oxidation of d-luciferin accompanied by emission of yellow–green light with a peak at 560 nm. Among many applications, Ppy luciferase has been used extensively as a reporter gene in living cells and organisms. However, some biological applications are limited by the low stability of the luciferase and limited intracellular luciferin concentration. To address these challenges, efforts to protein engineer Ppy luciferase have resulted in a number of mutants with improved properties such as thermostability, pH tolerance, and catalytic turn over. In this work, we combined amino acid mutations that were shown to enhance the enzyme’s thermostability (Mutant E) with those reported to enhance catalytic activity (LGR). The resulting mutant (YY5) contained eight amino acid changes from the wild-type luciferase and exhibited both improved thermostability and brighter luminescence at low luciferin concentrations. Therefore, YY5 may be useful for reporter gene applications.
Introduction
Bioluminescence is widely observed among organisms from different branches of life, including bacteria, fungi, and various metazoans. The luciferase from the North American firefly Photinus pyralis (Ppy) is one of the most extensively studied enzymes involved in bioluminescence. The Ppy luciferase catalyzes a two-step reaction that uses d-luciferin, adenosine triphosphate (ATP), and oxygen as substrates to yield oxyluciferin in an electronically excited state. Yellow–green light (560 nm) is emitted when the excited oxyluciferin relaxes to the ground state.1−5 Because of the low luminescence background of cells and tissues, luciferases have been used as reporter genes to sensitively monitor gene expression in living cells and animals.6−8
While the Ppy luciferase has proved to be highly useful for numerous applications, improvements are still needed. There are several desirable characteristics of the luciferase for biological applications in living cells and animals. For example, emission of longer-wavelength light improves tissue penetration in animals. Lower Km’s for the substrate ATP and d-luciferin would also likely enhance brightness of the light produced by intracellular luciferase. Similarly, increased thermostability of the firefly luciferase can enhance bioluminescence in vivo by increasing the effective half-life of the enzyme.9 Protein engineering efforts have resulted in a number of luciferase variants with altered or improved properties such as shifted luminescence spectra, thermostability, pH tolerance, and catalytic activity.5,10,11 For example, a recent study reported a chimeric luciferase that emits red light with a lower Km value for ATP.12 Alternatively, instead of engineering the enzyme, synthetic luciferin substrates have been developed that improve cell permeability of the substrate to improve emission and alter the luminescence spectra of the emitted light.13−15 Recombinant luciferases have also been used for applications in vitro, for example, as biosensors16 and as the light source in bioluminescence resonance energy transfer reactions.17,18 Such applications will also benefit from engineered luciferases with improved sensitivity and thermostability.
Further engineering of luciferases that exhibit combinations of the desirable characteristics mentioned above can lead to improved mutants for biological applications. Fujii et al. and Noda et al. discovered and characterized a triple mutant LGR (I423L, D436G, L530R) via random mutagenesis and screening that displays 20-fold lower Km values for ATP and d-luciferin compared to the wild-type (WT).19,20 The kcat values of the LGR mutant were also reported to be 4-fold higher than those of the WT.19 However, this LGR mutant was expected to show low stability comparable to that of the WT. Baggett et al. reported a combination of five amino acid substitutions (Mutant E: T214A, A215L, I232A, F295L, and E345K) that conferred significant thermostability, which led to improved bioluminescence in mammalian cells.9 The five point mutations in Mutant E have been successfully combined with those that shift the emission peak to green and red wavelengths to confer thermostability to the color mutants.21 Here, we combined the mutations reported in LGR and Mutant E with the expectation of obtaining a mutant (YY5) with improved catalytic activity at low d-luciferin concentration and thermostability.
Results and Discussion
Protein Expression and Purification
The genes encoding the WT and mutant luciferases were cloned in a plasmid and recombinantly expressed in Escherichia coli BL21(DE3) in terrific broth (50 mL). The enzymes were tagged with 6× histidine at the C-terminus, which was used for purification by immobilized metal ion affinity chromatography. All purified luciferases were analyzed by SDS polyacrylamide gel electrophoresis (SDS-PAGE) and were found to be highly homogenous (Figure S1).
Luminescence Spectra
Luminescence spectra of the luciferase variants were measured in the presence of the enzyme (1.6 μg/mL), d-luciferin (50 μM), and ATP (2 mM) at pH 7.8 (Figure 1). The WT and Mutant E showed essentially identical spectra with a peak at approximately 560 nm, as previously reported.21 The LGR and YY5 mutants both display a noticeable shoulder at the 600–650 nm region. Although the spectrum of the LGR mutant has not been reported, the double mutant (D436G, L530R) characterized by Fujii et al., shows a similar spectrum.20
Figure 1.

Normalized luminescence spectra of the luciferase variants. All luminescence spectra were measured at 25 °C. (A) WT, (B) LGR, (C) Mutant E, (D) YY5.
Thermostability Measurement
To measure the thermostability of each luciferase variant, the enzymes were incubated at different temperatures (25, 30, 35, 40, and 45 °C) and sampled over time. Activity of the luciferase samples were then measured at 25 °C using a microplate reader in the presence of d-luciferin (40 μM) and ATP (2 mM) (Figure 2). The WT was stable for the duration of the experiment (360 min) at 25 and 30 °C. However, the WT luciferase lost approximately 70% activity after 120 min at 35 °C and was completely inactivated within 30 min at 40 and 45 °C (Figure 2A). Qualitatively, a similar trend was observed with the LGR mutant (Figure 2B). As expected, Mutant E displayed high thermostability (Figure 2C), retaining ∼60% activity after 120 min at 45 °C. Interestingly, activity of Mutant E abruptly increased when incubated at 35 and 40 °C by >60% within the first measurement time (30 or 60 min). This observation was reproducible, and we speculate that the purified enzyme may be partially misfolded, which refolds into the active structure at the elevated temperature. Nevertheless, the decay of activity following the initial jump parallels that at 45 °C, further confirming the enhanced thermostability at physiological temperature. The YY5 mutant showed essentially the same characteristics, but the rates of the decay at 35–45 °C appear to be slower than those of Mutant E.
Figure 2.

Thermostability assay of the luciferase variants (A) WT, (B) LGR, (C) Mutant E, and (D) YY5. The purified luciferases were incubated at various temperatures (25–45 °C) and sampled over time. Then, the samples were assayed for enzyme activity at 25 °C. The experiments were performed in triplicate, and the error bars indicate standard error.
Measurement of Kinetic Parameters
Luciferase activity in the presence of varying concentrations of d-luciferin was measured in the presence of the enzyme (0.44 μg/mL) and excess ATP (2 mM) at 25 °C. The Ppy luciferase reaction kinetics is characterized by initial burst of light (flash) upon substrate addition followed by the steady “glow” phase because of product inhibition (Figure S2).22 In this study, light intensity measured 5 min after the start of the reaction was used as the reference point for activity because the glow phase of the luciferase light output would be more representative of the pseudo-steady state of the luciferase reactions relevant to bioimaging applications. The activity profile over different d-luciferin concentrations was fitted to the standard Michaelis–Menten equation to calculate the apparent Km and kcat values for the substrate (Figure 3, Table 1). It should be noted that because luminescence intensity is measured instead of the product concentration, the calculated Km and kcat values are apparent or relative values based on the luminescence measurement.
Figure 3.

Determination of kinetic parameters of the luciferase variants. (A) WT, (B) LGR, (C) Mutant E, (D) YY5, and (E) all variants at low d-luciferin concentrations.
Table 1. Kinetic Parameters of Firefly Luciferase Variantsa.
| enzyme | Km (μM) | kcat (cps/nM) | kcat/Km |
|---|---|---|---|
| WT | 1.21 ± 0.04 | 9.70 × 104 | 8.00 × 104 |
| LGR | 0.76 ± 0.06 | 3.87 × 105 | 5.09 × 105 |
| mutant E | 8.81 ± 0.27 | 1.05 × 106 | 1.20 × 105 |
| YY5 | 4.20 ± 0.18 | 1.37 × 106 | 3.26 × 105 |
Apparent kcat values were obtained by dividing the calculated apparent Vmax(cps) by the luciferase concentration (7 nM).
The LGR mutant showed a lower apparent Km value of 0.76 μM than that of the WT (1.21 μM). Although Noda et al. reported a more dramatic decrease in Km of the LGR mutant (>20-fold reduction relative to the WT), it should be noted that they used different measurement conditions, for example, lower ATP concentration (10 μM) and the timing of luminescence measurement (5 s). Consistent with the previous reports, we observed approximately 4-fold increase in the apparent kcat relative to the WT (Table 1).
In contrast, Mutant E displayed an apparent Km value that is 7.3-fold higher than that of the WT (Figure 3, Table 1). However, the apparent kcat was higher than that of the WT or the LGR mutant. Therefore, although Mutant E shows higher maximum activity (Vmax) at high d-luciferin concentrations, its activity at low d-luciferin concentrations (below 4 μM) lags behind that of the LGR mutant (Figure 3E).
The YY5 mutant shows an apparent Km that is lower than Mutant E (4.2 μM) and the highest apparent kcat among all variants. Although the Km value is higher than those of the WT and the LGR mutant, the improved activity makes YY5 the most active luciferase at 2 μM d-luciferin concentration (Figure 3E).
Structural Analysis
The crystal structure of the WT Ppy luciferase23 (PDB ID: 1BA3) was used to visualize the relative positions of the mutations that were incorporated into YY5 (Figure 4). The luciferase structure is divided into the large N-terminal and the small C-terminal domains, which are connected by a short linker. The three substitutions (I423L, D436G, and L530R) that comprise the LGR mutant are localized in the vicinity of the interface of the two domains where the substrates are bound, which is consistent with their effects on Km and kcat values.19,20 The mutations that confer thermostability in Mutant E (T214A, A215L, I232A, F295L, and E345K), however, are all located in the N-terminal domain distant from the active site.24 Consequently, it is reasonable that the two sets of mutations do not interfere with the respective phenotypes when combined in YY5.
Figure 4.
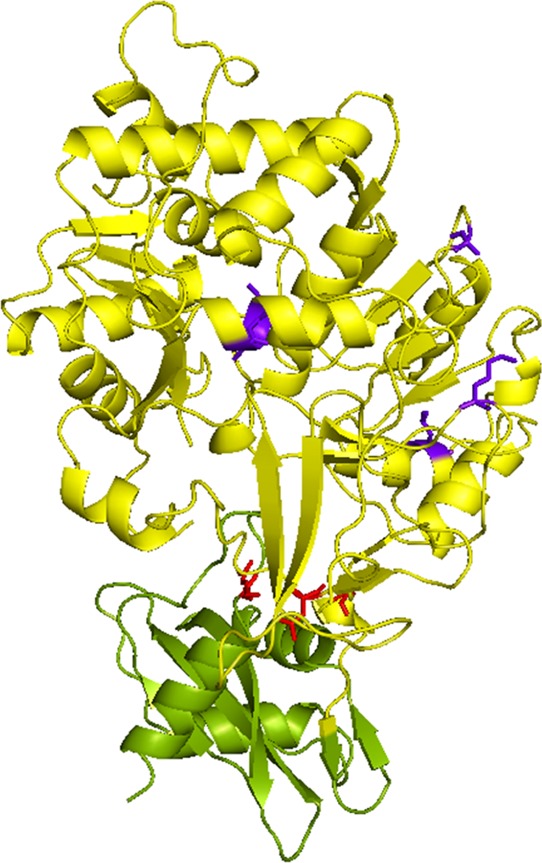
Locations of the introduced mutations depicted in the reported crystal structure of the Ppy luciferase. Protein Data Bank file 1BA3 was obtained and analyzed by PyMOL. The N-terminal domain is shown in yellow, and the C-terminal domain is shown in green. The amino acid mutations derived from LGR and Mutant E are indicated in red and purple, respectively.
Conclusions
Over the years, luciferases have been engineered for various applications in chemistry and biology.11 In particular, the Ppy luciferase remains the most popular bioluminescence reporter for bioimaging applications. However, most protein engineering efforts on the Ppy luciferase have focused on improving or altering a specific property, for example, thermostability, emission spectrum, or catalytic activity. Combinations of the mutations that confer distinct characteristics could further enhance the practical utility of the luciferase for reporter gene applications. For example, Branchini et al. incorporated a set of thermostabilizing mutations into color mutants of Ppy luciferase.21
Our objective was to construct a firefly luciferase mutant with higher activity (lower Km and higher kcat values) and higher thermostability by combining the characteristics of the previously reported mutations for each property. Through in vitro biochemical assays of the WT and the variants (LGR, Mutant E, and YY5), we demonstrated that the new mutant YY5 displays improved luminescence activity at low d-luciferin concentrations. As it has been suggested that the low cellular permeability of d-luciferin is a limiting factor in bioimaging applications of the firefly luciferase, YY5 may be useful for such applications.
Methods
Plasmid Construction
The WT luciferase and its variants were cloned into pTrcHis2 vector (Thermo Fisher) by standard molecular biology techniques and sequence verified by Sanger sequencing. The plasmid map and the DNA sequence of the expression cassette are provided in the Supporting Information (Figures S3, S4, and S5).
Expression and Purification of Luciferase Variants
The luciferase expression plasmids were transformed into E. coli BL21(DE3) competent cells (Novagen). The plasmid-containing cells were cultured in 50 mL of terrific broth supplemented with ampicillin (50 μg/mL) at 200 rpm and 37 °C. Protein expression was induced by the addition of isopropyl β-d-1-thiogalactopyranoside (0.1 mM for WT and LGR, 0.4 mM for Mutant E and YY5) when the OD600 value reached 0.6. Then, the cells were cultured for 16 h at 200 rpm and 28 °C.
The cells were centrifuged (Beckman Coulter, Allegra 6KR) at 10 000 rpm for 20 min at 4 °C, and the cell pellet was resuspended in 1 mL of the lysis buffer (50 mM Tris–HCl pH 7.5, 5% glycerol, 1 mM dithiothreitol (DTT), and 1 mM phenylmethylsulfonyl fluoride). The cells were lysed by adding a stock solution of lysozyme to the final concentration of 300 μg/mL and incubated for 16 hours. The lysate was treated with 0.5 U/μL Benzonase Nuclease (Sigma-Aldrich) for 30 min at 4 °C. Then, the cell lysate was centrifuged (Eppendorf 5415D) at 13 000 rpm for 30 min at 4 °C, and the supernatant was applied to the His-Spin Protein Miniprep kit (Zymo Research) following the manufacturer’s instructions.
The purified enzyme solution was dialyzed against a 50 mM Tris–HCl buffer (pH 7.8) using concentrator tubes with a molecular weight cut-off value of 50 kDa (Amicon Ultra-15 EMD Millipore). The enzymes were analyzed by SDS-PAGE (Figure S1), and protein concentrations were determined using the Micro BCA Protein Assay Kit (Thermo Fisher Scientific). The purified enzymes were stored in 50% glycerol (v/v) solution at −20 °C until use.
Measurement of Luminescence Spectra
To characterize the luminescence spectra of the luciferase variants, individual buffered solutions containing DTT, bovine serum albumin (BSA), MgCl2, Tris–HCl, and each enzyme were prepared at pH 7.8. The reaction solutions were prepared in 1 mL quartz cuvettes by combining 400 μL of the buffered enzyme solution with 100 μL of a d-luciferin stock solution (500 μM) and 500 μL of an ATP stock solution (4 mM). The final concentrations of each component in the reactions were 0.5 mM DTT, 0.1 mg/mL BSA, 2.5 mM MgCl2, 50 mM Tris–HCl, 1.6 μg/mL enzyme, 50 μM d-luciferin, and 2 mM ATP. Luminescence spectra were acquired by scanning from 400 to 800 nm at room temperature using the Fluorolog-3 spectrofluorometer (HORIBA).
Thermostability Assay
The thermostability assays were set up by first preparing an 8 mL buffered enzyme solution at pH 7.8 that contained 1.3 mM DTT, 0.3 mg/mL BSA, 6.3 mM of MgCl2, and 125 mM Tris–HCl. Aliquots of 50 μL of buffered enzyme solutions were incubated at temperatures ranging from 25 to 45 °C in triplicate using a thermal cycler (T-100, Bio-Rad). The enzyme solutions were removed from the thermal cycler at appropriate time intervals and were kept on ice until luminescence measurement. Luciferase activity was measured using the Tecan Infinite M1000 PRO microplate reader equipped with an autoinjector module. The incubated enzyme solutions (40 μL) were mixed with 400 μM d-luciferin stock solution (10 μL) in 96-well microplate wells. Luciferase reaction was started by injecting 50 μL of ATP stock solution (4 mM), and the luminescence intensity 5 min after injection was recorded. The final concentrations in the reaction were 0.5 mM DTT, 0.1 mg/mL BSA, 2.5 mM MgCl2, 50 mM Tris–HCl, 0.44 μg/mL (7 nM) enzyme, 2 mM ATP, and 40 μM d-luciferin.
Determination of Kinetic Parameters
Preliminary assays were performed using different ATP concentrations ranging from 10 μM to 6 mM, and 2 mM ATP was found to be sufficient to saturate the luciferase activity under our assay conditions (data not shown). Luciferase assay was performed as described above for thermostability assay except that different concentrations (0–60 μM) of d-luciferin were used. Each reaction was performed in quadruplicate. Luminescence intensity values at the 5 min time point following ATP injection were recorded as the apparent activity, which was used to plot the graphs shown in Figure 3. The data were fitted to the Michaelis–Menten equation using the software GraphPad Prism 6 (GraphPad Software) to calculate apparent Km and kcat values.
Acknowledgments
This project was financially supported by W.M. Keck Foundation and OIST.
Supporting Information Available
The Supporting Information is available free of charge on the ACS Publications website at DOI: 10.1021/acsomega.7b02068.
Characterization of the purified enzymes by SDS-PAGE; Plasmid map; Amino acid; time course of luciferase variant light emissions; and DNA sequences of the luciferase variants (PDF)
Author Contributions
T.P. performed all the experiments and modeling and drafted the manuscript. F.A. performed preliminary characterization of the luciferase variants and optimization of assay conditions. Y.N. constructed the expression plasmids and performed initial characterization of the luciferase variants. A.L. planned experimental work and reviewed the manuscript. Y.Y. conceived and supervised the project and reviewed the manuscript.
The authors declare no competing financial interest.
Supplementary Material
References
- Green A. A.; McElroy W. D. Crystalline firefly luciferase. Biochim. Biophys. Acta 1956, 20, 170–176. 10.1016/0006-3002(56)90275-x. [DOI] [PubMed] [Google Scholar]
- Deluca M.; McElroy W. D. [1] Purification and properties of firefly luciferase. Methods Enzymol. 1978, 57, 3–15. 10.1016/0076-6879(78)57003-1. [DOI] [Google Scholar]
- de Wet J. R.; Wood K. V.; Helinski D. R.; DeLuca M. Cloning of firefly luciferase cDNA and the expression of active luciferase in Escherichia coli. Proc. Natl. Acad. Sci. U.S.A. 1985, 82, 7870–7873. 10.1073/pnas.82.23.7870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conti E.; Franks N. P.; Brick P. Crystal structure of firefly luciferase throws light on a superfamily of adenylate-forming enzymes. Structure 1996, 4, 287–298. 10.1016/s0969-2126(96)00033-0. [DOI] [PubMed] [Google Scholar]
- Kaskova Z. M.; Tsarkova A. S.; Yampolsky I. V. 1001 lights: luciferins, luciferases, their mechanisms of action and applications in chemical analysis, biology and medicine. Chem. Soc. Rev. 2016, 45, 6048–6077. 10.1039/c6cs00296j. [DOI] [PubMed] [Google Scholar]
- Naylor L. H. Reporter gene technology: the future looks bright. Biochem. Pharmacol. 1999, 58, 749–757. 10.1016/s0006-2952(99)00096-9. [DOI] [PubMed] [Google Scholar]
- Liu A. M. F.; New D. C.; Lo R. K. H.; Wong Y. H. Reporter gene assays. Methods Mol. Biol. 2009, 486, 109–123. 10.1007/978-1-60327-545-3_8. [DOI] [PubMed] [Google Scholar]
- Niu G.; Chen X. Molecular imaging with activatable reporter systems. Theranostics 2012, 2, 413–423. 10.7150/thno.3940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baggett B.; Roy R.; Momen S.; Morgan S.; Tisi L.; Morse D.; Gillies R. J. Thermostability of firefly luciferases affects efficiency of detection by in vivo bioluminescence. Mol. Imaging 2004, 3, 324–332. 10.1162/1535350042973553. [DOI] [PubMed] [Google Scholar]
- Koksharov M. I.; Ugarova N. N. Approaches to engineer stability of beetle luciferases. Comput. Struct. Biotechnol. J. 2012, 2, e201204004 10.5936/csbj.201209004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loening A. M.; Walls Z. F.. Engineering Luciferases for Assays and Imaging. In Engineering in Translational Medicine; Cai W., Eds., 1st ed.; Springer: London, 2014, pp 203–231. [Google Scholar]
- Branchini B. R.; Southworth T. L.; Fontaine D. M.; Kohrt D.; Welcome F. S.; Florentine C. M.; Henricks E. R.; DeBartolo D. B.; Michelini E.; Cevenini L.; Roda A.; Grossel M. J. Red-emitting chimeric firefly luciferase for in vivo imaging in low ATP cellular environments. Anal. Biochem. 2017, 534, 36–39. 10.1016/j.ab.2017.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reddy G. R.; Thompson W. C.; Miller S. C. Robust light emission from cyclic alkylaminoluciferin substrates for firefly luciferase. J. Am. Chem. Soc. 2010, 132, 13586–13587. 10.1021/ja104525m. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans M. S.; Chaurette J. P.; Adams S. T. Jr.; Reddy G. R.; Paley M. A.; Aronin N.; Prescher J. A.; Miller S. C. A synthetic luciferin improves bioluminescence imaging in live mice. Nat. Methods 2014, 11, 393–395. 10.1038/nmeth.2839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mofford D. M.; Reddy G. R.; Miller S. C. Aminoluciferins extend firefly luciferase bioluminescence into the near-infrared and can be preferred substrates over D-luciferin. J. Am. Chem. Soc. 2014, 136, 13277–13282. 10.1021/ja505795s. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang W.; Zhao Q.; Luo M.; Li M.; Wang D.; Wang Y.; Liu Q. Immobilization of Firefly Luciferase on PVA-co-PE Nanofibers Membrane as Biosensor for Bioluminescent Detection of ATP. ACS Appl. Mater. Interfaces 2015, 7, 20046–20052. 10.1021/acsami.5b07339. [DOI] [PubMed] [Google Scholar]
- Alam R.; Zylstra J.; Fontaine D. M.; Branchini B. R.; Maye M. M. Novel multistep BRET-FRET energy transfer using nanoconjugates of firefly proteins, quantum dots, and red fluorescent proteins. Nanoscale 2013, 5, 5303–5306. 10.1039/c3nr01842c. [DOI] [PubMed] [Google Scholar]
- Alam R.; Karam L. M.; Doane T. L.; Coopersmith K.; Fontaine D. M.; Branchini B. R.; Maye M. M. Probing Bioluminescence Resonance Energy Transfer in Quantum Rod–Luciferase Nanoconjugates. ACS Nano 2016, 10, 1969–1977. 10.1021/acsnano.5b05966. [DOI] [PubMed] [Google Scholar]
- Noda K.; Matsuno T.; Fujii H.; Kogure T.; Urata M.; Asami Y.; Kuroda A. Single bacterial cell detection using a mutant luciferase. Biotechnol. Lett. 2008, 30, 1051–1054. 10.1007/s10529-008-9643-3. [DOI] [PubMed] [Google Scholar]
- Fujii H.; Noda K.; Asami Y.; Kuroda A.; Sakata M.; Tokida A. Increase in bioluminescence intensity of firefly luciferase using genetic modification. Anal. Biochem. 2007, 366, 131–136. 10.1016/j.ab.2007.04.018. [DOI] [PubMed] [Google Scholar]
- Branchini B. R.; Ablamsky D. M.; Murtiashaw M. H.; Uzasci L.; Fraga H.; Southworth T. L. Thermostable red and green light-producing firefly luciferase mutants for bioluminescent reporter applications. Anal. Biochem. 2007, 361, 253–262. 10.1016/j.ab.2006.10.043. [DOI] [PubMed] [Google Scholar]
- DeLuca M.; McElroy W. D. Kinetics of the firefly luciferase catalyzed reactions. Biochemistry 1974, 13, 921–925. 10.1021/bi00702a015. [DOI] [PubMed] [Google Scholar]
- Franks N. P.; Jenkins A.; Conti E.; Lieb W. R.; Brick P. Structural basis for the inhibition of firefly luciferase by a general anesthetic. Biophys. J. 1998, 75, 2205–2211. 10.1016/s0006-3495(98)77664-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Branchini B. R.; Southworth T. L.; Khattak N. F.; Michelini E.; Roda A. Red- and green-emitting firefly luciferase mutants for bioluminescent reporter applications. Anal. Biochem. 2005, 345, 140–148. 10.1016/j.ab.2005.07.015. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.