

Abstract
Background:
Dandelion is commonly used in traditional Chinese medicine with several active compounds found in extracts. It has a variety of pharmacological effects, such as a reduction in swelling and inflammation, and detoxification. The mechanism by which dandelion extract inhibits the inflammatory response in skeletal muscle cells remains unknown; therefore, the aim of this study was to investigate the effects of dandelion extract root on the proliferation of skeletal muscle cells and the alleviation of lipopolysaccharide (LPS)-induced inflammatory response in vitro.
Methods:
Rat skeletal muscle cells were isolated from Sprague-Dawley rat and cultured in vitro which were cultured in basal medium, or medium containing LPS or dandelion extract. Cell counting kit-8 (CCK-8) was employed to measure cell proliferation; meanwhile, the optimal concentration of dandelion extract and treatment time were selected. Crystal violet staining was used to detect the proliferation of muscle cells. Western blotting analysis was used to detect the levels of inflammatory factors, myogenic factor, and p-AKT protein expression.
Results:
The optimal concentration and treatment time of dandelion extract for the following study were 5 mg/ml and 4 days, respectively. Dandelion extract was found to increase proliferation of rat skeletal muscle cells (t = 3.145, P < 0.05), with the highest effect observed at 5 mg/ml. LPS was found to decrease proliferation of skeletal muscle cells (t = −131.959, P < 0.001), and dandelion extract could against this affection (t = 19.466, P < 0.01). LPS could induce expression of inflammatory factors, including interleukin (IL)-1β, IL-6 and tumor necrosis factor (TNF)-α (IL-1β: t = 9.118, P < 0.01; IL-6: t = 4.346, P < 0.05; TNF-α: t = 15.806, P < 0.05), and dandelion extract was shown to reduce LPS-induced expression of IL-1β, IL-6 and TNF-α (IL-1β: t = −2.823, P < 0.05; IL-6: t = −3.348, P < 0.01; and TNF-α: t = −3.710, P < 0.01). Furthermore, LPS was also shown to decrease expression of myogenic factor, including myod1 and myogenin (MyoD1: t = 4.039, P < 0.05 and myogenin: t = 3.300, P < 0.01), but dandelion extract was shown to against this effect of LPS (MyoD1: t = −3.160, P < 0.05 and myogenin: t = −3.207, P < 0.01). And then, LPS was found to increase expression of p-AKT protein (p-AKT/AKT: t = 4.432, P < 0.05). Moreover, expression of p-AKT protein was found to decrease, with 5 mg/ml of dandelion extract (p-AKT/AKT: t = −3.618, P < 0.05).
Conclusions:
The findings indicate that dandelion extract plays an important role in skeletal muscle cells viability regulation, promote cells proliferation by increasing level of p-AKT protein expression, and reduce LPS-induced expression of inflammatory factors, inhibiting the inflammatory response of rat skeletal muscle cells.
Keywords: Anti-Inflammation Mechanism, Dandelion Extract, Herbal Plant, Inflammation, Skeletal Muscle Cells
摘要
背景:
公英通常作为传统中医药,其提取物中含有几种活性化合物。蒲公英具有多种药理作用,如缓解肿胀、炎症,以及 抗毒性作用等。但蒲公英提取物抑制骨骼肌细胞炎症反应的机制仍不清楚。因此,本研究的目的是探讨蒲公英根部提取物对 骨骼肌细胞增殖的影响以及体外抑制LPS诱导的炎症反应。
方法:
从SD大鼠中分离大鼠骨骼肌细胞,并在体外进行普通培养基、含有脂多糖(LPS)或蒲公英提取物的培养基中培养。采 用细胞计数试剂盒-8(CCK-8)对细胞增殖情况进行检测,同时找出蒲公英提取物的最佳浓度和最佳处理时间。结晶紫染色用 于检测肌细胞的增殖情况。 Western blot检测并分析骨骼肌细胞中炎症因子,生肌因子和p-AKT蛋白的表达水平。
结果:
研究发现蒲公英提取物促进骨骼肌细胞增殖的最佳浓度和最佳处理时间分别为5mg/ml和4d。我们发现蒲公英提取物能 够促进大鼠骨骼肌细胞增殖(t = 3.145,p <0.05),在5mg/ml时效果最佳。并且LPS可以抑制骨骼肌细胞增殖(t = -131.959,p <0.001),蒲公英提取物可以抵抗LPS的这种作用(t = 19.466,p <0.01)。 LPS可以诱导炎症因子IL-1β,IL-6和TNF-α的表达 (IL-1β:t = 9.118,p <0.01; IL-6:t = 4.346,p <0.05; TNF-α: t=15.806, p<0.05),蒲公英提取物减少LPS诱导的IL-1β,IL-6 和TNF-α的表达(IL-1β:t = -2.823,p <0.05; IL-6:t = -3.348,p <0.01; TNF -α:t = -3.710,p <0.01)。此外,LPS还显示降低 生肌因子的表达,包括myod1和肌细胞生成素(myod1:t = 4.039,p <0.05;肌细胞生成素:t = 3.3,p <0.01),但蒲公英提取 物显示出对抗LPS的这种作用 (myod1:t = -3.16,p <0.05;肌细胞生成素:t = -3.207,p <0.01)。我们进一步发现,LPS能够 增加骨骼肌细胞p-AKT蛋白的表达(p-AKT/AKT:t = 4.432,p <0.05)。 此外,在LPS处理细胞后,用5mg/ml的蒲公英提 取物发现p-AKT蛋白的表达减少(p-AKT/AKT:t = -3.618,p <0.05)。
结论:
研究结果表明,蒲公英提取物在骨骼肌细胞增殖调节中起重要作用,通过提高p-AKT蛋白表达水平促进细胞增殖,并 减少LPS诱导的炎性因子表达,抑制大鼠骨骼肌细胞的炎症反应。
INTRODUCTION
Inflammation is a complex pathophysiological process mediated by various signaling molecules produced by leukocytes, macrophages, and mast cells. Damage to the connective tissue of muscle fibers resulting from eccentric exercise can lead to the accumulation of bradykinin, histamine, and prostaglandins, which attract monocytes and neutrophils to the injury site from small blood vessels. Monocytes, neutrophils and the products they release can stimulate type III and IV nerve endings, resulting in delayed-onset muscle soreness (DOMS).
Inflammation can induce the release of various cytokines such as interferon-gamma, interleukin (IL)-1 beta, IL-8, IL-6, and tumor necrosis factor alpha (TNF-α).[1] The purpose of the inflammatory response is to promote the healing of injured tissue.[2,3] Inflammation-triggered phagocytosis can clean the necrotic muscle tissue, which plays an important role in the process of muscle repair and ultimately promotes the recovery of the tissue. However, excessive inflammation can cause the secondary damage to skeletal muscle. Thus, the inflammatory response in muscle recovery is a “double-edged sword” and controlling the inflammatory responses has become a potential therapeutic target for the alleviation of DOMS.
At present, therapeutic options for DOMS include Western anti-inflammatory medicines, application of either hot or old compresses, physiotherapy, and acupuncture. However, their reputed effects remain controversial. The inflammatory response is essential for the recovery process and cannot be inhibited completely. Effective treatment for DOMS must, therefore, ensure that an appropriate inflammatory response is maintained while limiting excessive inflammation to avoid secondary damage. Most anti-inflammatory drugs are relatively strong and could result in an over-suppression of inflammation. Traditional Chinese medicine (TCM) is known to be mild and to have less side effects. Currently, there are few studies that have focused on the effects of Chinese medicine on DOMS.
Dandelion contains plant-based cholesterol, has long been used clinically because of its diuretic and anti-inflammatory properties.[4] The study has shown that dandelion has acute anti-inflammatory activities and protects cholecystokinin-induced acute pancreatitis in rats.[5] Pharmaceutical compounds that have been identified in dandelion extract include phenolic acids and flavonoids.[6,7,8,9] Studies have shown that some of these compounds have potential therapeutic effects in inflammation.[10,11,12,13] Dandelion also contains triterpenes and sterols.[14,15] Triterpenes are widely distributed in plants and are commonly identified as the major components responsible for anti-inflammatory effects.[16,17,18] Sterols have also been shown as good inhibitors of inflammation.[19,20]
However, the role of dandelion extract on skeletal muscle DOMS remains unknown. In this study, we investigated the effects of dandelion extract on skeletal muscle cell proliferation, as well as its ability to inhibit the inflammatory response to prevent secondary inflammation.
METHODS
Preparation of dandelion extract, and primary rat skeletal muscle cell isolation and culture
Dandelion root was purchased from Guangxi Wuzhou Pharmaceutical (Group) Co., Ltd. (Guangxi, China). The dandelion was ground to a powder and soaked in different concentrations and volumes of solvents (water, ethanol, methanol, and chloroform), mixed and left to stand. The resulting supernatant was filtered, concentrated under reduced pressure at low temperature, and lyophilized to obtain dandelion extract powder. The extract was dissolved in cell culture medium to different concentrations (1.25, 2.5, 5, 10, 20, and 40 mg/ml).
Four-week-old SPF grade male Sprague-Dawley rats, average weight 200 ± 3 g, were purchased from Beijing Huafukang Bioscience Co., Ltd. (Huafukang Bioscience Co., Ltd., Beijing, China). Quadriceps were dissected out from both hindlimbs under sterile conditions. All adipose and connective tissue were removed and minced into small pieces, then digested in 0.25% trypsin with shaking and titration every 3–5 min. After 1 h, Dulbecco's Modified Eagle Medium (DMEM) containing 15% fetal bovine serum was added to stop the reaction. The digestion mix was then centrifuged at 800 rpm for 5 min. The supernatant was collected and filtered through a 100-μm nylon mesh. After filtration, the suspension was centrifuged again at 800 rpm for 5 min. The supernatant was discarded, and the pellet was resuspended in culture medium. Cells were seeded and cultured at 37°C with 5% CO2. All procedures were carried out following the Laboratory Animal Management Regulations published by People's Republic of China Science and Technology Commission. All protocols were approved by the Animal Care Committee at Shengjing Hospital of China Medical University (ID: 2015PS321K).
Cell counting kit-8
Cell activity was determined using Cell counting kit-8 (CCK-8; Solarbio Science and Technology Co., Ltd., Beijing, China). Skeletal muscle cells were seeded into 96-well plates at a density of 2 × 103 per well, with three replicate wells per group. After cells attached to wells, they were treated with different concentrations of dandelion extract (1.25, 2.5, 5, 10, 20, and 40 mg/ml) for various time points (1, 2, 3, 4, and 5 days). At the end of each time point, CCK-8 (10 μl/100 μl) was added into the desired wells and incubated for another 4 h. Absorbance values were assessed using a microplate reader at an absorbance of 450 nm. And then, according to the results of the previous experimental results obtained, the cells were treated with 0, lipopolysaccharide (LPS) with 5 mg/ml dandelion extract and LPS, CCK-8 (10 μl/100 μl) was added into the desired wells and incubated for another 4 h at 4 days. Absorbance values were assessed using a microplate reader at an absorbance of 450 nm.
Crystal violet staining
The effect of dandelion extract on the proliferation of rat skeletal muscle cells was assessed by crystal violet staining. Primary muscle cells were seeded into 6-well plates at a density of 2 × 105 per well. Cells were divided into three groups: control group, experimental group (LPS + dandelion extract), and positive control group (LPS alone). After cells attached to the wells, LPS (Solarbio Science and Technology Co., Ltd.) was added to the positive control, and experimental groups and cells were incubated overnight. The following day, media from control and experimental groups was replaced with media containing 5 mg/ml dandelion extract (based on CCK-8 results). Four days after the dandelion extract treatment, the media was aspirated, and cells were washed three times with phosphate-buffered saline (PBS; Hyclone, Utah, USA). Triton solution (0.2%) was added to each well and incubated at room temperature for 15 min. After incubation, Triton solution was replaced with crystal violet solution (Solarbio Science and Technology Co., Ltd.). Thirty minutes later, crystal violet solution was discarded, and cells were washed three times with PBS, before recording cell images.
Western blotting analysis
Western blotting analysis was employed to detect the level of expression of inflammatory factors in skeletal muscle cells. Cultured cells were passed into three flasks and as assigned to either control group, experimental group (LPS + dandelion extract), or positive control group (LPS alone). After cells were attached to flasks, LPS (0.1 μg/ml) was added to the positive control, and experimental groups and cells were incubated overnight. The next day, media from the control and experimental groups were replaced with media containing 5 mg/ml dandelion extract (based on CCK-8 results). Four days later, cells were harvested and incubated with protein lysis buffer on ice for 30 min. Cells then were centrifuged at 12,000 rpm for 30 min (4°C), and the supernatants were collected. A BCA assay kit (Beyotime Biotechnology, Shanghai, China) was used to measure the total protein concentration. After boiling at 100°C for 5 min, protein extracts were aliquoted and stored at −20°C until used for detection. Twenty μg of protein was separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and electrophoretically transferred to a polyvinylidene fluoride membrane at 100 V for various times, depending on the size of the proteins of interest. Membranes were blocked in Tris-buffered saline (TBS) containing 5% non-fat milk at room temperature for 2 h and then probed with either anti-IL-1β antibody (Proteintech, Chicago, USA; 1:2000 dilution), anti-IL-6 antibody (NOVUS, Colorado, USA; 1:1000 dilution), anti-TNF-α antibody (NOVUS, 1:1000 dilution), anti-Myogenin antibody (NOVUS, 1:1000 dilution), anti-MyoD1 antibody (Proteintech, Chicago, USA; 1:2000 dilution), anti-AKT antibody (Cell Signaling Technology, Boston, USA; 1:2000 dilution), anti-p-AKT antibody (Cell Signaling Technology, 1:1000 dilution), or anti-glyceraldehyde phosphate dehydrogenase antibody (Proteintech, 1:10000 dilution) at 4°C. After overnight incubation, membranes were washed three times with TBS and Tween 20 (TBST) and then incubated with an anti-rabbit secondary antibody (Proteintech, 1:5000 dilution) or an anti-mouse secondary antibody (Proteintech, 1:5000 dilution) at room temperature with gentle shaking for 2 h. After washing the membranes with TBST, ECL Western blotting substrate (Thermo Fisher Scientific, Waltham, MA, USA) was added to the membranes and images were collected using a C300 Chemiluminescent Blot Imaging System (Azure, USA). Absorbance values for each band were quantified.
Statistical analysis
All the data analyses were done using SPSS 19.0 (SPSS Inc., Chicago, IL, USA). Each experiment was repeated three times and data were presented as mean ± standard deviation (SD). The difference between each group was determined using a two-tailed Student's t-test and statistical significance was assessed at P < 0.05.
RESULTS
Dandelion extract enhanced the proliferation of rat skeletal muscle cells
Exponentially growing muscle cells (2 × 103 cells/100 μl) were seeded into 96-well plates and incubated with dandelion extract at different concentrations (0, 1.25, 2.5, 5, 10, 15, 20, 40 mg/ml) for different time periods (1, 2, 3, 4, and 5 days). Variability in cell numbers was detected using CCK-8 after treatment with various concentrations of the dandelion extract [Figure 1]. A significant increase was shown in rat skeletal muscle cell number and activity 4 days after treatment with 5 mg/ml dandelion extract (t = 3.145, P < 0.05).
Figure 1.
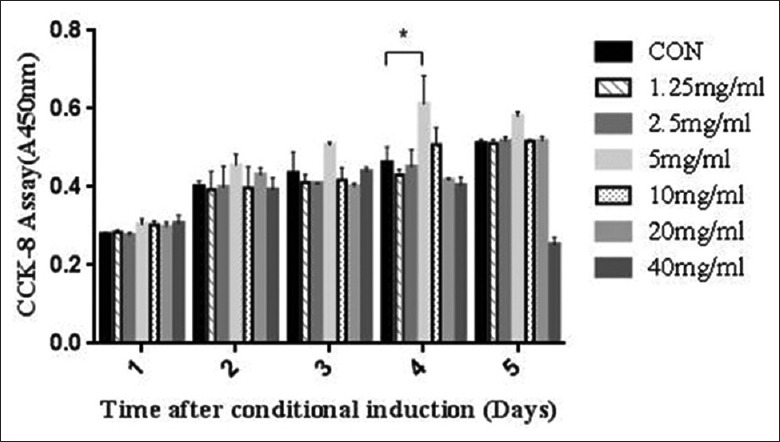
The effect of various concentrations of dandelion extract on the proliferation of rat skeletal muscle cells. The group treated with 5 mg/ml dandelion extract for 4 days showed a significant increase in proliferation, while the group treated with 40 mg/ml for 4–5 days showed a significant reduction in proliferation. *P < 0.05. CON: Control.
Lipopolysaccharide inhibited the proliferation of rat skeletal muscle cells
Exponentially growing muscle cells (2 × 103 cells/100 μl) were seeded into 96-well plates and incubated with 5 mg/ml dandelion extract for 4 days. Variability in cell numbers was detected using CCK-8 after treatment with various groups (0, LPS with 5 mg/ml dandelion extract, and LPS) [Figure 2]. Our data showed a significant decrease in rat skeletal muscle cell number and activity 4 days after treatment with LPS compared with LPS + dandelion extract group (t = −131.959, P < 0.01), and also showed a significant increase in rat skeletal muscle cell number and activity 4 days after treatment with LPS and dandelion extract compared with control group (t = 19.466, P < 0.01).
Figure 2.
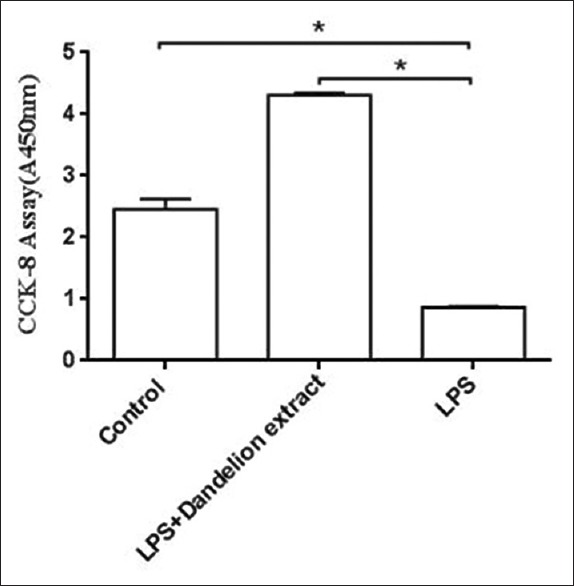
The effect of different groups on the proliferation of rat skeletal muscle cells. Compared with the control group, the group treated with LPS and 5 mg/ml dandelion extract for 4 days showed a significant increase in proliferation, while the group treated with LPS showed a significant reduction in proliferation compared with LPS + dandelion extract group. *P < 0.01. CON: Control; LPS: Lipopolysaccharide.
Histological analysis of the effect of dandelion extract on rat skeletal muscle cells through crystal violet staining
Rat skeletal muscle cells were treated with 5 or 40 mg/ml dandelion extract for 4 days, and the cells were then stained with crystal violet. Cell images were obtained using a microscope with a 100 × magnification [Figure 3]. When cells were treated with 5 mg/ml extract (B and b), cells numbers were significantly increased compared with the control group (A and a). However, when cells were treated with 40 mg/ml extract (C and c), cells numbers decreased significantly compared with the control group. These results suggest that 5 mg/ml dandelion extract can enhance muscle cell proliferation.
Figure 3.

The effect of 0, 5, and 40 mg/ml dandelion extract on the proliferation of rat skeletal muscle cells. The effect of 0, 5, and 40 mg/ml dandelion extract on the proliferation of rat skeletal muscle cells. After 4 days of treatment with 5 mg/ml dandelion extract a significant increase in proliferation of cells was seen, whereas 40 mg/ml extract had a strong inhibitory effect on proliferation of cells (A-C: images captured with camera; a-c, ×100). CON: Control.
Dandelion extract suppressed the release of inflammatory factors
The expression of IL-1β, IL-6, and TNF-α proteins was detected by Western blotting analysis [Figure 4]. IL-1β, IL-6, and TNF-α in LPS groups were significantly increased than that in control groups, respectively (t = 9.118, P < 0.01; t = 4.346, P < 0.05; t = 15.806, P < 0.01), and IL-1β, IL-6, and TNF-α in LPS + dandelion groups were significant decreased than that in LPS groups, respectively (t = −3.710, P < 0.01; t = −3.348, P < 0.01; t = −2.823, P < 0.05). LPS-induced IL-1β, IL-6, and TNF-α expression was significantly inhibited by dandelion extract (5 mg/ml) treatment.
Figure 4.

The effect of dandelion extract on the expression of inflammatory factors (TNF-α, IL-6 and IL-1β) in the control, experimental and positive control groups, respectively. (a and b) Western blotting shows that 4 days after treatment with dandelion extract (5 mg/ml), a significant reduction in the levels of inflammatory factors was observed. *P < 0.05, †P < 0.01. CON: Control; TNF-α: Tumor necrosis factor-α; IL: Interleukin; GAPDH: Glyceraldehyde-3-phosphate dehydrogenase; LPS: Lipopolysaccharide.
Effect of dandelion extract on AKT protein
The levels of AKT and p-AKT were evaluated using Western blotting. As shown in Figure 5, compared with the control group, the protein levels of p-AKT in the LPS group were significantly increased than that in the control group (t = 4.432, P < 0.05). In dandelion extract (5 mg/ml) + LPS group, the protein levels of p-AKT were significantly decreased in compared with the LPS group (t = −3.618, P < 0.05).
Figure 5.
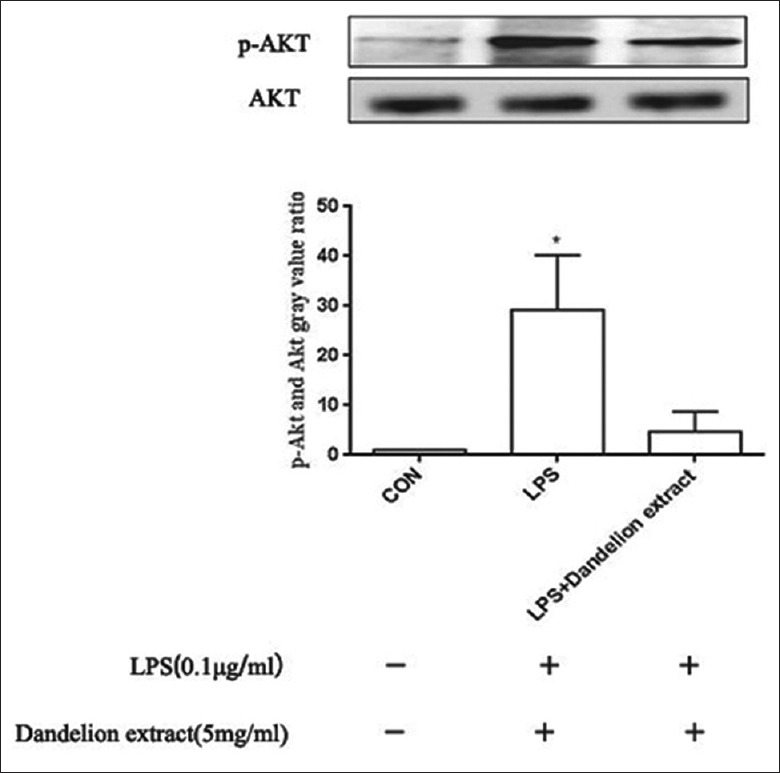
The effect of dandelion extract and LPS on the expression of p-AKT protein. Results are shown for the control, experimental, and negative control groups, respectively. Four days after treatment with dandelion extract (5 mg/ml), compared with the LPS group, a significant reduction in the levels of p-AKT protein was observed. *P < 0.05. CON: Control; LPS: Lipopolysaccharide.
Dandelion extract promoted the release of myogenic factors
Then, we explored the reason that how dandelion extract could promote proliferation of skeletal muscle cells. We detected the expression of myogenic factors including myogenin and MyoD1 proteins by Western blotting analysis [Figure 6]. Myogenin in LPS group were decreased than that in control group (t = 3.300, P < 0.01) and in LPS + dandelion group (t = −3.207, P < 0.01), MyoD1 in LPS group were significantly decreased than that in control group (t = 4.039, P < 0.05) and in LPS + dandelion group (t = −3.160, P < 0.05). LPS+ dandelion extract (5 mg/ml) also promoted myogenin and MyoD1 expression significantly than control group and LPS group.
Figure 6.

The effect of dandelion extract on the expression of myogenic factors (myogenin and MyoD1). (a and b) Western blotting shows that, 4 days after treatment with dandelion extract (5 mg/ml) a significant increment in the levels of MyoD1 protein was observed (*P < 0.05). (c and d) Western blotting shows that, 4 days after treatment with dandelion extract (5 mg/ml) a significant increment in the levels of myogenin protein was observed (†P < 0.01). CON: Control; GAPDH: Glyceraldehyde-3-phosphate dehydrogenase; LPS: Lipopolysaccharide.
DISCUSSION
DOMS is commonly accompanied by a series of changes in muscle tissue structure, physiology, and biochemistry. Secondary inflammation can aggravate the clinical symptoms of DOMS, including muscle aches and pains, and muscle weakness and swelling. This affects the endurance ability and strength of the muscle and ultimately leads to a decrease in work capacity and power output and hence sport and exercise performance, overall.[21,22,23] The inflammatory response during DOMS is triggered by mechanical disruption of sarcomeres, T-tubules, myofibrils, cytoskeletal proteins, and the sarcoplasmic reticulum. Inflammatory responses occur within 1 h of sustaining muscle injury and reach a peak at 48 h, but the extensive infiltration of monocytes and macrophages continued for 2–4 days. The primary symptoms of DOMS are thought to be mediated by leukocytes and cytokines, which play important roles not only in clearing cell debris, but also in disrupting myofilaments, cytosolic structures, and the membrane of damaged muscles.[24]
In this study, we investigated whether the Chinese medicine, dandelion extract, can relieve skeletal muscle cell inflammation and stimulate skeletal muscle cell activity and proliferation. In the present study, we chose the commonly used LPS to induce an in vitro inflammatory model using primary skeletal muscle cells. LPS is a potent inducer of inflammation and a key mediator of the innate immune response. Stimulation of skeletal muscle cells by LPS leads to the production and secretion of cytokines and other inflammatory mediators, and subsequently promotes a proinflammatory response.[25] Dandelion has been widely used as a traditional medicine against various disorders.[26,27] It has been found the phenolic compounds in dandelion including: luteolin, chrysoeriol, chicoric acid, and chlorogenic acid.[6] Among them, luteolin and chicoric acid are the most abundant phenolic compounds. Luteolin plays a role as an antioxidant, an anti-inflammatory agent, and a free radical scavenger.[28] Chicoric acid could decrease LPS-stimulated nuclear factor (NF)-κB level and TNF-α production in macrophages.[29] Hence, we considered luteolin and chicoric acid may be the active ingredients of dandelion extract to anti-inflammatory in LPS-induced inflammatory reaction of skeletal muscle cells, in addition, also includes triterpenes and sterols. The ethanol reflux extraction method and high-performance liquid chromatography were employed for the preparation and analysis of dandelion extract which is a classic method to extract Chinese medicine. We further study found that LPS could inhibit cell activity and proliferation, and dandelion extract could against this inhibitory effect.
The inflammatory factors including IL-1β, IL-6, and TNF-α are a direct contributor to the persistent cellular infiltration at inflammatory sites and the induction of tissue destruction. Studies have shown that myocytes and other cells can release IL-1β, IL-6, and TNF-α when skeletal muscle is damaged.[30,31,32,33,34,35] The role of IL-6 in exercise-induced skeletal muscle injury was suggested in studies of eccentric exercise (a common model of exercise-induced skeletal muscle damage).[31] Tomiya et al.[32] produced eccentric contraction of muscle through electrical stimulation. IL-6 was detected in the muscle cytoplasm 12 h after eccentric exercise, and it was also detected in inflammatory cells and proliferating satellite cells,[33] suggesting that the cytokine is associated with the inflammatory responses in eccentric exercise-induced muscle damage. Tracey et al.[34] found an increase in TNF-α release during skeletal muscle injury. Moreover, TNF-α has been shown to be important in the pathological process by recruiting other inflammatory cells and stimulating pain and cartilage breakdown.[35] This cytokine mediates the early inflammatory response by regulating the production of other cytokines, including IL-1β and IL-6.[7]
In this study, we found that compared to the control group, dandelion extract significantly decreased LPS-induced expression of IL-1β, IL-6, and TNF-α in rat skeletal muscle cells, which is consistent with the study by Zhang et al.[36] However, the detecting targets including IL-1β, IL-6, TNF-α, and treatment time were different. These differences could be due to the different tissues with different blood supply, leading to the different concentration of drug administered. Another possible reason could be the different concentrations of treatments. Ma et al.[37] induced inflammation in the lung using LPS and found that the aqueous extract of dandelion significantly reduced the expression of IL-1β, IL-6, and TNF-α induced by LPS, which was consistent with our findings. Recently, Yang et al.[38] used dandelion to treat LPS-induced acute bronchitis in rats and found that the content of IL-6 and TNF-α in the bronchial tissue was significantly reduced after treatment, which agreed with our results. Another study published by Yang et al.[39] in 2016 also showed similar results. In the study by Xueshibojie et al.,[40] dandelion significantly reduced smoking-induced lung inflammation and decreased the expressions of IL-1β, IL-6, and TNF-α. Numerous studies have shown that dandelion, as a traditional Chinese medicine, has significantly reduced the expression of inflammatory cytokines IL-1β, IL-6, and TNF-α, and relieved the inflammatory response in the body. Our study found that dandelion extract can significantly reduce the expression of IL-1β, IL-6, and TNF-α, and alleviate the LPS-induced inflammatory response in skeletal muscle cells.
Our study also found that dandelion extract, is able to enhance the proliferation of rat skeletal muscle cells, and significantly increase the myogenin and MyoD1 protein expression. We showed that dandelion extract had the strongest effect at a concentration of 5 mg/ml. Interestingly, studies have shown that TNF plays a role in the regulation of muscle formation and is an essential regulator in the early stages of myoblast differentiation through the p38 MAPK pathway.[41,42] However, there are also studies that have shown that treatment of C2C12 myoblast-like cells with TNF-α were results in the loss of MyoD1 and myogenin proteins, and the inhibition of myotube formation. TNF-α further inhibits myogenesis by inducing the expression of nitric oxide synthase and the production of nitric oxide through the NF-kB pathway, which binds to the promoter site of MyoD1 and downregulates myogenic transcription, leading to a reduction in MyoD1 expression and activity.[43,44] Differences are thought to relate to the concentration of TNF-α used. A low concentration, 0.05 ng/ml, increased the activity and expression of p38, myogenin, and main histocompatibility complex, however, concentrations of 0.5–5 ng/ml had an inhibitory effect. Therefore, whether TNF-α has a stimulatory or inhibitory effect in muscle regeneration is concentration dependent. Low concentrations have a physiological effect and stimulate muscle regeneration; high concentrations have a pathological effect and inhibit muscle regeneration, which agreed with our findings. Our data showed that dandelion extract enhanced the proliferation of skeletal muscle cells at a concentration of 5 mg/ml, but inhibited it at a concentration of 40 mg/ml. In vivo study demonstrated that TNF-α is chemotactic to myoblasts and can promote satellite cell mitosis, suggesting a direct effect of TNF-α in muscle regeneration.[45] It has been suggested that TNF-α is not only an enhancement factor that initiates the proliferation of satellite cells, but also a sensory factor that promotes satellite cell entry into the cell cycle and hence the repair of muscle damage. Our results show that an appropriate concentration (5 mg/ml) of dandelion extract can partially inhibit the expression and secretion of TNF-α, and also could promote the myogenic factors such as myogenin and MyoD1, which may be related to the enhancement of skeletal muscle cell proliferation. That is, the dandelion extract at a concentration of 5 mg/ml can appropriately inhibit the expression of TNF-α, promote myogenic factors and then increase muscle cell proliferation.
In this study, we found that dandelion extract may exert some of its anti-inflammatory effects by affecting the activity of AKT in LPS-induced inflammatory reaction in skeletal muscle cells of rat. Although AKT activation is crucial for the recovery phase after skeletal muscle injury, moreover, dandelion extract can inhibit AKT in muscle cells after LPS induced. The mechanisms of dandelion extract in muscle cells without the effect of LPS are not clear; thus, the mechanisms need to be clarified in the future. As this is a preliminary study, we have not yet investigated the potential signaling pathways involved in the inflammatory process in skeletal muscle cells, so cannot elucidate the mechanism by which dandelion extract suppressed inflammation. We also did not examine the role of myogenic factors such as MyoD and myogenin in the promotion of skeletal muscle cell proliferation by the extract.
Taken together, our findings showed that dandelion extract can enhance the proliferation and activity of rat skeletal muscle cells and LPS can inhibit cell activity and proliferation. As mentioned earlier, dandelion can inhibit the expression of inflammatory cytokines, including IL-1β, IL-6, and TNF-α and promote the expression of myogenic cytokines, such as myogenin and MyoD1, suggesting that the TCM may have a synergistic effect to promote cell proliferation and alleviate the inflammatory response in the process of relieving DOMS. TNF-α may be the key cytokine in this synergistic effect. This study provides a potential direction for further investigations into the role of dandelion extract in rat skeletal muscle cells and suggests that dandelion is a promising drug for the treatment of skeletal muscle injury.
Financial support and sponsorship
This work was supported by grants from the National Key Research and Development Program (No. 2016YFC0107102 [S. P.]); the National Natural Science Foundation of China under Grant (No. 30871211, 81271538 [S. P.]).
Conflicts of interest
There are no conflicts of interest.
Footnotes
Edited by: Li-Shao Guo
REFERENCES
- 1.Guastadisegni C, Nicolini A, Balduzzi M, Ajmone-Cat MA, Minghetti L. Modulation of PGE2 and TNF-α by nitric oxide inresting and LPS-activated RAW 264.7 cells. Cytokine. 2002;19:175–80. doi: 10.1006/cyto.2002.1955. doi: 10.1006/cyto.2002.1955. [DOI] [PubMed] [Google Scholar]
- 2.Dunster JL. The macrophage and its role in inflammation and tissue repair: mathematical and systems biology approaches. Wiley Interdiscip Rev Syst Biol Med. 2016;8:87–99. doi: 10.1002/wsbm.1320. doi: 10.1002/wsbm.1320. [DOI] [PubMed] [Google Scholar]
- 3.Molofsky AB, Savage AK, Locksley RM. Interleukin-33 in tissue homeostasis, injury, and inflammation. Immunity. 2015;42:1005–19. doi: 10.1016/j.immuni.2015.06.006. doi: 10.1016/j.immuni.2015.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ahmad VU, Yasmeen S, Ali Z, Khan MA, Choudhary MI, Akhtar F, et al. Taraxacin, a new guaianolide from Taraxacum wallichii. J Nat Prod. 2000;63:1010–1. doi: 10.1021/np990495+. doi: 10.1021/np990495+ [DOI] [PubMed] [Google Scholar]
- 5.Seo SW, Koo HN, An HJ, Kwon KB, Lim BC, Seo EA, et al. Taraxacum officinale protects against cholecystokinin-induced acute pancreatitis in rats. World J Gastroenterol. 2005;11:597–9. doi: 10.3748/wjg.v11.i4.597. doi: 10.3748/wjg.v11.i4.597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Williams CA, Goldstone F, Greenham J. Flavonoids, cinnamic acids and coumarins from the different tissues and medicinal preparations of Taraxacum officinale. Phytochemistry. 1996;42:121–7. doi: 10.1016/0031-9422(95)00865-9. doi: 10.1016/0031-9422(95)00865-9. [DOI] [PubMed] [Google Scholar]
- 7.Koh YJ, Cha DS, Ko JS, Park HJ, Choi HD. Anti-inflammatory effect of Taraxacum officinale leaves on lipopolysaccharide-induced inflammatory responses in RAW 264.7 cells. J Med Food. 2010;13:870–8. doi: 10.1089/jmf.2009.1249. doi: 10.1089/jmf.2009.1249. [DOI] [PubMed] [Google Scholar]
- 8.Park CM, Ji YP, Song YS. Luteolin and chicoric acid, two major constituents of dandelion leaf, inhibit nitric oxide and lipid peroxide formation in lipopolysaccharide-stimulated RAW 264.7 cells. J Food Sci Nutr. 2010;15:92–7. doi: 10.3746/jfn.2010.15.2.092. [Google Scholar]
- 9.Budzianowski J. Coumarins, caffeoyltartaric acids and their artifactual methyl esters from Taraxacum officinale leaves. Planta Med. 1997;63:288. doi: 10.1055/s-2006-957681. doi: 10.1055/s-2006-957681. [DOI] [PubMed] [Google Scholar]
- 10.Hoult JR, Payá M. Pharmacological and biochemical actions of simple coumarins: Natural products with therapeutic potential. Gen Pharmacol. 1996;27:713–22. doi: 10.1016/0306-3623(95)02112-4. doi: 10.1016/0306-3623(95)02112-4. [DOI] [PubMed] [Google Scholar]
- 11.Leal LK, Ferreira AA, Bezerra GA, Matos FJ, Viana GS. Antinociceptive, anti-inflammatory and bronchodilator activities of Brazilian medicinal plants containing coumarin: A comparative study. J Ethnopharmacol. 2000;70:151–9. doi: 10.1016/s0378-8741(99)00165-8. doi: 10.1016/S0378-8741(99)00165-8. [DOI] [PubMed] [Google Scholar]
- 12.Cheng JF, Chen M, Wallace D, Tith S, Arrhenius T, Kashiwagi H, et al. Discovery and structure-activity relationship of coumarin derivatives as TNF-alpha inhibitors. Bioorg Med Chem Lett. 2004;14:2411–5. doi: 10.1016/j.bmcl.2004.03.022. doi: 10.1016/S0960-894X(04)00355-5. [DOI] [PubMed] [Google Scholar]
- 13.Shan J, Fu J, Zhao Z, Kong X, Huang H, Luo L, et al. Chlorogenic acid inhibits lipopolysaccharide-induced cyclooxygenase-2 expression in RAW264.7 cells through suppressing NF-kappaB and JNK/AP-1 activation. Int Immunopharmacol. 2009;9:1042–8. doi: 10.1016/j.intimp.2009.04.011. doi: 10.1016/j.intimp.2009.04.011. [DOI] [PubMed] [Google Scholar]
- 14.Simandi B, Kristo ST, Kéry A, Selmeczi LK, Kmecz I, Kemény S. Supercritical fluid extraction of dandelion leaves. J Supercrit Fluids. 2002;23:135–42. doi: 10.1016/S0896-8446(02)00012-8. [Google Scholar]
- 15.Akashi T, Furuno T, Takahashi T, Ayabe SI. Biosynthesis of triterpenoids in cultured cells and regenerated and wild plant organ of Taraxacum officinale. Phytochemistry. 1994;36:303–8. doi: 10.1016/S0031-9422(00)97065-1. [Google Scholar]
- 16.Ríos JL, Recio MC, Máñez S, Giner RM. Natural triterpenoids as anti-inflammatory agents. Stud Nat Prod Chem. 2000;22:93–143. doi: 10.1016/S1572-5995(00)80024-1. [Google Scholar]
- 17.Siqueira JM, Jr, Peters RR, Gazola AC, Krepsky PB, Farias MR, Rae GA. Anti-inflammatory effects of a triterpenoid isolated from Wilbrandia ebracteata cogn. Life Sci. 2007;80:1382–7. doi: 10.1016/j.lfs.2006.12.021. doi: 10.1016/j.lfs.2006.12.021. [DOI] [PubMed] [Google Scholar]
- 18.Banno N, Akihisa T, Yasukawa K, Tokuda H, Tabata K, Nakamura Y, et al. Anti-inflammatory activities of the triterpene acids from the resin of Boswellia carteri. J Ethnopharmacol. 2006;107:249–53. doi: 10.1016/j.jep.2006.03.006. doi: 10.1016/j.jep.2006.03.006. [DOI] [PubMed] [Google Scholar]
- 19.Ammar S, Edziri H, Mahjoub MA, Chatter R, Bouraoui A, Mighri Z, et al. Spasmolytic and anti-inflammatory effects of constituents from Hertia cheirifolia. Phytomedicine. 2009;16:1156–61. doi: 10.1016/j.phymed.2009.03.012. doi: 10.1016/j.phymed.2009.03.012. [DOI] [PubMed] [Google Scholar]
- 20.Akihisa T, Yasukawa K. Antitumor-promoting and anti-inflammatory activities of triterpenoids and sterols from plants and fungi. Stud Nat Prod Chem. 2001;25:43–87. doi: 10.1016/S1572-5995(01)80005-3. [Google Scholar]
- 21.Torres R, Ribeiro F, Alberto Duarte J, Cabri JM. Evidence of the physiotherapeutic interventions used currently after exercise-induced muscle damage: Systematic review and meta-analysis. Phys Ther Sport. 2012;13:101–14. doi: 10.1016/j.ptsp.2011.07.005. doi: 10.1016/j.ptsp.2011.07.005. [DOI] [PubMed] [Google Scholar]
- 22.Lavender AP, Nosaka K. Changes in fluctuation of isometric force following eccentric and concentric exercise of the elbow flexors. Eur J Appl Physiol. 2006;96:235–40. doi: 10.1007/s00421-005-0069-5. doi: 10.1007/s00421-005-0069-5. [DOI] [PubMed] [Google Scholar]
- 23.Graven-Nielsen T, Arendt-Nielsen L. Induction and assessment of muscle pain, referred pain, and muscular hyperalgesia. Curr Pain Headache Rep. 2003;7:443–51. doi: 10.1007/s11916-003-0060-y. doi: 10.1007/s11916-003-0060-y. [DOI] [PubMed] [Google Scholar]
- 24.Grounds MD. Reasons for the degeneration of ageing skeletal muscle: A central role for IGF-1 signalling. Biogerontology. 2002;3:19–24. doi: 10.1023/a:1015234709314. doi: 10.1023/A:1015234709314. [DOI] [PubMed] [Google Scholar]
- 25.Meng F, Lowell CA. Lipopolysaccharide (LPS)-induced macrophage activation and signal transduction in the absence of Src-family kinases Hck, Fgr, and Lyn. J Exp Med. 1997;185:1661–70. doi: 10.1084/jem.185.9.1661. doi: 10.1084/jem.185.9.1661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gargouri M, Magné C, Ben Amara I, Ben Saad H, El Feki A. Dandelion-enriched diet of mothers alleviates lead-induced damages in liver of newborn rats. Cell Mol Biol (Noisy-le-grand) 2017;63:67–75. doi: 10.14715/cmb/2017.63.2.10. doi: 10.14715/cmb/2017.63.2.10. [DOI] [PubMed] [Google Scholar]
- 27.Wirngo FE, Lambert MN, Jeppesen PB. The physiological effects of dandelion (Taraxacum officinale) in type 2 diabetes. Rev Diabet Stud. 2016;13:113–31. doi: 10.1900/RDS.2016.13.113. doi: 10.1900/RDS.2016.13.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Harris GK, Qian Y, Leonard SS, Sbarra DC, Shi X. Luteolin and chrysin differentially inhibit cyclooxygenase-2 expression and scavenge reactive oxygen species but similarly inhibit prostaglandin-E2 formation in RAW 264.7 cells. J Nutr. 2006;136:1517–21. doi: 10.1093/jn/136.6.1517. doi: 10.1093/jn/136.6.1517. [DOI] [PubMed] [Google Scholar]
- 29.Stevenson LM, Matthias A, Banbury L, Penman KG, Bone KM, Leach DL, et al. Modulation of macrophage immune responses by Echinacea. Molecules. 2005;10:1279–85. doi: 10.3390/10101279. doi: 10.3390/10101279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Butterfield TA, Best TM, Merrick MA. The dual roles of neutrophils and macrophages in inflammation: A critical balance between tissue damage and repair. J Athl Train. 2006;41:457–65. doi: 10.1016/S0162-0908(08)79217-1. [PMC free article] [PubMed] [Google Scholar]
- 31.Armstrong RB, Ogilvie RW, Schwane JA. Eccentric exercise-induced injury to rat skeletal muscle. J Appl Physiol Respir Environ Exerc Physiol. 1983;54:80–93. doi: 10.1152/jappl.1983.54.1.80. doi: 10.1152/jappl.1983.54.1.80. [DOI] [PubMed] [Google Scholar]
- 32.Tomiya A, Aizawa T, Nagatomi R, Sensui H, Kokubun S. Myofibers express IL-6 after eccentric exercise. Am J Sports Med. 2004;32:503–8. doi: 10.1177/0095399703258788. doi: 10.1177/0095399703258788. [DOI] [PubMed] [Google Scholar]
- 33.Chan MH, Carey AL, Watt MJ, Febbraio MA. Cytokine gene expression in human skeletal muscle during concentric contraction: Evidence that IL-8, like IL-6, is influenced by glycogen availability. Am J Physiol Regul Integr Comp Physiol. 2004;287:R322–7. doi: 10.1152/ajpregu.00030.2004. doi: 10.1152/ajpregu.00030.2004. [DOI] [PubMed] [Google Scholar]
- 34.Tracey KJ, Wei H, Manogue KR, Fong Y, Hesse DG, Nguyen HT, et al. Cachectin/tumor necrosis factor induces cachexia, anemia, and inflammation. J Exp Med. 1988;167:1211–27. doi: 10.1084/jem.167.3.1211. doi: 10.1084/jem.167.3.1211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Chao CC, Hu S. Tumor necrosis factor-alpha potentiates glutamate neurotoxicity in human fetal brain cell cultures. Dev Neurosci. 1994;16:172–9. doi: 10.1159/000112104. doi: 10.1159/000112104. [DOI] [PubMed] [Google Scholar]
- 36.Zhang X, Xiong H, Li H, Cheng Y. Protective effect of taraxasterol against LPS-induced endotoxic shock by modulating inflammatory responses in mice. Immunopharmacol Immunotoxicol. 2014;36:11–6. doi: 10.3109/08923973.2013.861482. doi: 10.3109/08923973.2013.861482. [DOI] [PubMed] [Google Scholar]
- 37.Ma C, Zhu L, Wang J, He H, Chang X, Gao J, et al. Anti-inflammatory effects of water extract of Taraxacum mongolicum hand.-Mazz on lipopolysaccharide-induced inflammation in acute lung injury by suppressing PI3K/Akt/mTOR signaling pathway. J Ethnopharmacol. 2015;168:349–55. doi: 10.1016/j.jep.2015.03.068. doi: 10.1016/j.jep.2015.03.068. [DOI] [PubMed] [Google Scholar]
- 38.Yang N, Li C, Tian G, Zhu M, Bu W, Chen J, et al. Organic acid component from taraxacum mongolicum hand.-Mazz alleviates inflammatory injury in lipopolysaccharide-induced acute tracheobronchitis of ICR mice through TLR4/NF-κB signaling pathway. Int Immunopharmacol. 2016;34:92–100. doi: 10.1016/j.intimp.2016.02.028. doi: 10.1016/j.intimp.2016.02.028. [DOI] [PubMed] [Google Scholar]
- 39.Yang N, Dong Z, Tian G, Zhu M, Li C, Bu W, et al. Protective effects of organic acid component from Taraxacum mongolicum hand-Mazz against LPS-induced inflammation: Regulating the TLR4/IKK/NF-κB signal pathway. J Ethnopharmacol. 2016;194:395–402. doi: 10.1016/j.jep.2016.08.044. doi: 10.1016/j.jep.2016.08.044. [DOI] [PubMed] [Google Scholar]
- 40.Xueshibojie L, Duo Y, Tiejun W. Taraxasterol inhibits cigarette smoke-induced lung inflammation by inhibiting reactive oxygen species-induced TLR4 trafficking to lipid rafts. Eur J Pharmacol. 2016;789:301–7. doi: 10.1016/j.ejphar.2016.07.047. doi: 10.1016/j.ejphar.2016.07.047. [DOI] [PubMed] [Google Scholar]
- 41.Chen SE, Gerken E, Zhang Y, Zhan M, Mohan RK, Li AS, et al. Role of TNF-{alpha} signaling in regeneration of cardiotoxin-injured muscle. Am J Physiol Cell Physiol. 2005;289:C1179–87. doi: 10.1152/ajpcell.00062.2005. doi: 10.1152/ajpcell.00062.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Chen SE, Jin B, Li YP. TNF-alpha regulates myogenesis and muscle regeneration by activating p38 MAPK. Am J Physiol Cell Physiol. 2007;292:C1660–71. doi: 10.1152/ajpcell.00486.2006. doi: 10.1152/ajpcell.00486.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Acharyya S, Ladner KJ, Nelsen LL, Damrauer J, Reiser PJ, Swoap S, et al. Cancer cachexia is regulated by selective targeting of skeletal muscle gene products. J Clin Invest. 2004;114:370–8. doi: 10.1172/JCI20174. doi: 10.1172/JCI200420174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Guttridge DC, Mayo MW, Madrid LV, Wang CY, Baldwin AS., Jr NF-kappaB-induced loss of myoD messenger RNA: Possible role in muscle decay and cachexia. Science. 2000;289:2363–6. doi: 10.1126/science.289.5488.2363. doi: 10.1126/science.289.5488.2363. [DOI] [PubMed] [Google Scholar]
- 45.Warren GL, Hulderman T, Jensen N, McKinstry M, Mishra M, Luster MI, et al. Physiological role of tumor necrosis factor alpha in traumatic muscle injury. FASEB J. 2002;16:1630–2. doi: 10.1096/fj.02-0187fje. doi: 10.1096/fj.02-0187fje. [DOI] [PubMed] [Google Scholar]