

ABSTRACT
The tropism of human cytomegalovirus (HCMV) is influenced by the envelope glycoprotein complexes gH/gL/gO and gH/gL/UL128-131. During virion assembly, gO and the UL128-131 proteins compete for binding to gH/gL in the endoplasmic reticulum (ER). This assembly process clearly differs among strains, since Merlin (ME) virions contain abundant gH/gL/UL128-131 and little gH/gL/gO, whereas TR contains much higher levels of total gH/gL, mostly in the form of gH/gL/gO, but much lower levels of gH/gL/UL128-131 than ME. Remaining questions include (i) what are the mechanisms behind these assembly differences, and (ii) do differences reflect in vitro culture adaptations or natural genetic variations? Since the UL74(gO) open reading frame (ORF) differs in 25% of amino acids between TR and ME, we analyzed recombinant viruses in which the UL74(gO) ORF was swapped. TR virions were >40-fold more infectious than ME. Transcriptional repression of UL128-131 enhanced the infectivity of ME to the level of TR, despite still far lower levels of gH/gL/gO. Swapping the UL74(gO) ORF had no effect on either TR or ME. A quantitative immunoprecipitation approach revealed that gH/gL expression levels were within 4-fold between TR and ME, but the gO expression level was 20-fold lower for ME, which suggested differences in mRNA transcription, translation, or rapid ER-associated degradation of gO. trans-Complementation of gO expression during ME replication gave a 6-fold enhancement of infectivity beyond the 40-fold effect of UL128-131 repression alone. Overall, strain variations in the assembly of gH/gL complexes result from differences in the expression of gO and UL128-131, and selective advantages for reduced UL128-131 expression during fibroblast propagation are much stronger than those for higher gO expression.
IMPORTANCE Specific genetic differences between independently isolated HCMV strains may result from purifying selection on de novo mutations arising during propagation in culture or random sampling among the diversity of genotypes present in clinical specimens. Results presented indicate that while reduced UL128-131 expression may confer a powerful selective advantage during cell-free propagation of HCMV in fibroblast cultures, selective pressures for increased gO expression are much weaker. Thus, variation in gO expression among independent strains may represent natural genotype variability present in vivo. This may have important implications for virus-host interactions, such as immune recognition, and underscores the value of studying molecular mechanisms of replication using multiple HCMV strains.
KEYWORDS: genetic diversity, glycoproteins, human cytomegalovirus, tropism
INTRODUCTION
Human cytomegalovirus (HCMV) is widely spread throughout the world, found in approximately 60% of adults in developed countries and in 100% of adults in developing countries (reviewed in references 1–4). Immunocompromised individuals, such as those infected with HIV, or transplant recipients under antirejection treatments can suffer HCMV-related pathologies, including gastroenteritis, encephalitis, retinitis, and vasculopathies, which can accelerate allograft rejection. HCMV infection can also be acquired in utero, and this is a significant cause of congenital neurological impairments and sensorineural hearing loss. HCMV is transmitted mainly through body liquid, such as urine and saliva (5). Once infection is established, HCMV can spread throughout the body, infecting many of the major somatic cell types, including fibroblasts, smooth muscle cells, epithelial and endothelial cells, neurons, and leukocytes such as monocytes-macrophages and dendritic cells (6–9). HCMV does not replicate efficiently in transformed cells (10, 11); thus, most studies of the mechanisms governing HCMV tropism have involved dermal fibroblasts, retinal pigment epithelial cells, and umbilical cord endothelial cells, all of which can be easily cultured as normal, nontransformed cells.
Much focus has been on the gH/gL complexes, which, as for other herpesviruses, likely engage cell receptors and promote infection by contributing to the gB-mediated membrane fusion event or through activating cell signaling pathways (reviewed in references 12 and 13; 14). During virus assembly, the HCMV UL128-131 proteins and gO compete for binding to gH/gL to form the pentameric complex gH/gL/UL128-131 or the trimeric complex gH/gL/gO. Structural studies involving purified soluble complexes showed that gO and UL128 can each make a disulfide bond with cystine 144 of gL, and this was suggested to be the basis of the competitive assembly of the complexes (15). However, Stegmann et al. demonstrated that mutant gO lacking the cysteine implicated in the disulfide bond with gL formed intact and functional gH/gL/gO (16). This suggests that gO can engage in extensive noncovalent interactions with gH/gL. The gH/gL/UL128-131 complex is dispensable for infection of cultured fibroblasts and neuronal cells but is required for infection of epithelial and endothelial cells and monocytes-macrophages (17–21). In contrast, gH/gL/gO is critical for infection of all cell types (22–25). Both complexes likely interact with cell receptors. gH/gL/gO can bind platelet-derived growth factor receptor alpha (PDGFRα) through the gO subunit, and this interaction is critical for infection of fibroblasts (26–28). Epithelial and endothelial cells do not express PDGFRα, but blocking of gH/gL/gO with either neutralizing antibodies or soluble PDGFRα can inhibit infection of these cells, suggesting the existence of other gH/gL/gO receptors (26, 27). Receptors for gH/gL/UL128-131 might include epidermal growth factor receptor (EGFR) (also known as ErbB1) and β1 or β3 integrins, and these interactions may induce signaling cascades critical for infection of selected cell types, such as epithelial and endothelial cells and monocytes-macrophages (26, 29).
We recently reported that the amounts of gH/gL/gO and gH/gL/UL128-131 in the virion envelope can differ dramatically among commonly studied strains of HCMV and that this can affect the infectivity of the virions (25, 30). The salient results of those studies were that (i) Merlin (ME) virions contained gH/gL mostly in the form of gH/gL/UL128-131, whereas TR and TB 40/E (TB) virions had mostly gH/gL/gO; (ii) in terms of “total gH/gL,” the amount of gH/gL/gO in TR and TB virions was larger than the amount of gH/gL/UL128-131 in ME virions; (iii) the infectivity of all three strains on both fibroblasts and epithelial cells correlated with the amount of gH/gL/gO; and (iv) when the expression of UL128-131 was suppressed in ME, virions contained dramatically less gH/gL/UL128-131 but only slightly more gH/gL/gO. The latter point was especially curious since the model that gO and UL128-131 proteins compete for binding to gH/gL would predict that the fraction of gH/gL normally bound by UL128-131 would, in their absence, be bound by gO instead. This discrepancy could be explained by differences in the stoichiometric expression of gH/gL, gO, and UL128-131 between strains. An alternative hypothesis was suggested by the fact that there are at least eight alleles of the UL74 gene that encodes gO (31). The amino acid sequence of gO among these eight genotypes can vary between 10 and 30%, and this could affect competition with UL128-131 for binding to gH/gL. Both of these non-mutually-exclusive hypotheses were addressed in the experiments reported here.
(This article was submitted to an online preprint archive [32].)
RESULTS
Strains of HCMV display different patterns of glycoprotein expression and trafficking to virion assembly compartments.
The dramatic differences in the compositions of gH/gL complexes in TR and ME virions described previously by Zhou et al. (25, 30) suggested corresponding differences in glycoprotein expression and/or the trafficking of glycoproteins to virion assembly compartments (ACs). To address these possibilities, cells were infected for 2 days (Fig. 1A) or 5 days (Fig. 1B) with TR or ME, and steady-state amounts of viral proteins were compared by immunoblotting. At 2 days postinfection (dpi), immediate early protein 1/2 (IE1/2) levels were similar for both TR and ME, consistent with an equal multiplicity of infection. At 5 dpi, the levels of the virion structural proteins major capsid protein (MCP), gB, gH, and gL were also very similar between the two strains. In contrast, ME-infected cells contained dramatically more UL128-131 protein than did TR-infected cells. The UL148 protein was also included in these analyses because it was recently described as an endoplasmic reticulum (ER) chaperone protein that influences the ratio of gH/gL complexes (33). In TR-infected cells, an anti-UL148 antibody detected a prominent 35-kDa protein species, consistent with the previous description of the UL148 protein (33). This 35-kDa species was not detected in ME-infected cells. Instead, ME-infected cells contained two species that were less abundant and of higher and lower electrophoretic motilities than the single UL148 species detected in TR-infected cells. The basis of the apparent size difference was not characterized but could reflect differences in translational start/stop codon usage, splicing of the UL148 mRNA, or posttranslational modifications of the UL148 protein between strains. Overall, the pattern of expression of the UL128-131 and UL148 proteins correlated well with the previously described pentamer-rich nature of ME virions and the trimer-rich nature of TR virions (25, 30). Note that the expression of gO was not addressed in these analyses because the gO amino acid sequence differences between strains affect antibody recognition and preclude direct comparison (30).
FIG 1.
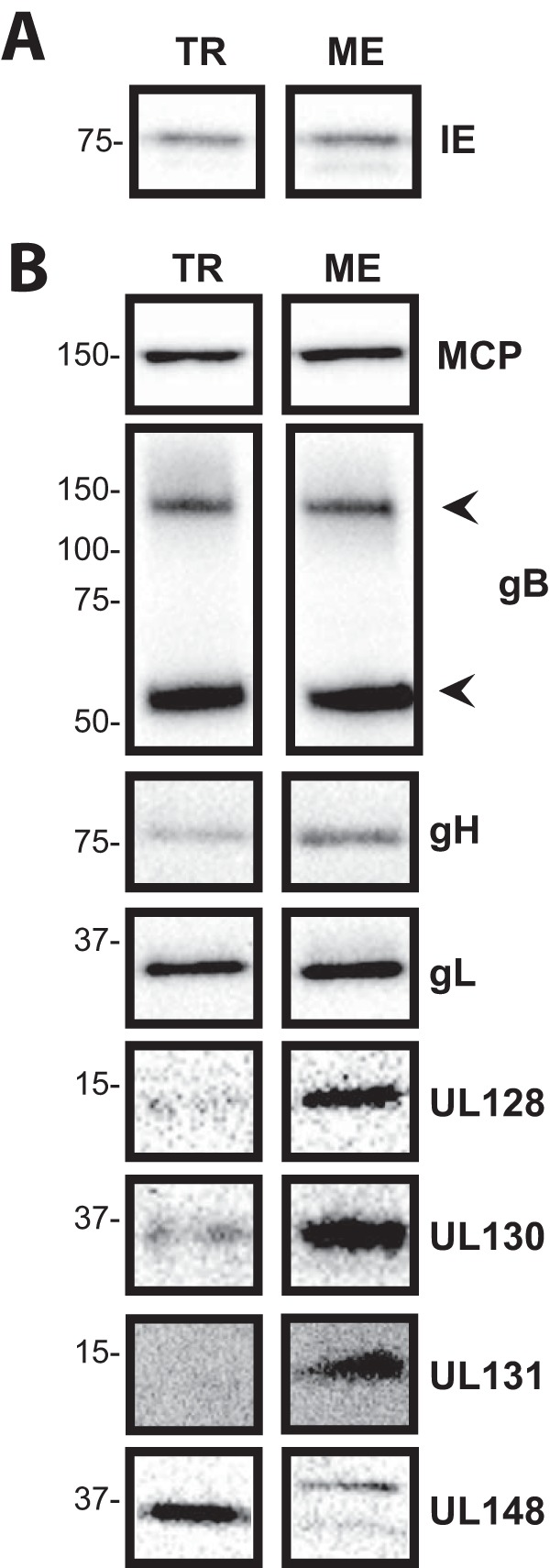
Comparison of protein expression between TR and ME. nHDF were infected with 1 PFU/cell of TR or ME. At day 2 (A) or day 5 (B), total cell extracts were separated by reducing SDS-PAGE and analyzed by immunoblot analysis probing for immediate early protein 1/2 (IE1/2), major capsid protein (MCP), gB, gH, gL, UL128, UL130, UL131, or UL148. Arrowheads indicate the positions of the cleaved 100-kDa and 55-kDa fragments of gB.
Trafficking of gH/gL from the ER to trans-Golgi network (TGN)-derived assembly compartments was assessed by treating the infected-cell extracts at 5 dpi with either endoglycosidase H (endo H) or peptide N-glycosidase F (PNGase F) and then analyzing gH and gL by immunoblotting (Fig. 2). The majority of gH and gL in TR-infected cells was endo H resistant, consistent with efficient transport from the ER to trans-Golgi network-derived ACs. In contrast, most of the gH and gL in ME-infected cells was sensitive to endo H digestion. In HFFFtet cells, which repress transcription from the UL128-131 locus (30, 34), there was even less endo H-resistant gH and gL. This suggested that the bulk of gH/gL trafficked to ACs in ME-infected neonatal human dermal fibroblasts (nHDF), which allow UL128-131 expression, represented gH/gL/UL128-131 and is consistent with previous observations that (i) the bulk of gH/gL in the ME virion is pentamers and (ii) the loss of gH/gL in the form of pentamers in ME-T virions due to the repression of the UL128-131 proteins is apparently not fully compensated for by the formation of complexes with gO (25, 30).
FIG 2.
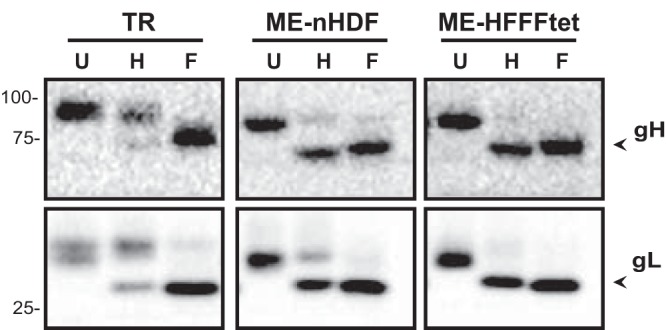
Analysis of ER-to-trans-Golgi compartment trafficking of glycoproteins in TR- or ME-infected cells. Extracts of nHDF infected with TR or ME or HFFFtet cells infected with ME were treated with endoglycosidase H (H) or PNGase F (F) or left untreated (U) and then separated by reducing SDS-PAGE and analyzed by immunoblot analysis probing for gH or gL. Arrowheads indicate the positions of the faster-migrating, deglycosylated species.
Differences in amino acid sequence of gO between TR and ME do not affect the infectivity of cell-free virus.
The predicted amino acid sequence of gO differs by 25% between TR and ME. This sequence divergence precluded direct comparison of gO expression levels because antibodies do not cross-react (30). Furthermore, these sequence differences could potentially affect the ability of the distinct gO isoforms to compete with the UL128-131 proteins for binding to gH/gL (thus influencing the amounts of gH/gL complexes in the mature virion envelope) or the function(s) of gO during entry, such as binding PDGFRα or other receptors. To address these possibilities, bacterial artificial chromosome (BAC) recombineering methods were used to replace the gO open reading frame (ORF) (UL74) of TR with the analogous sequences from ME, and visa versa, to generate recombinant viruses denoted TR_MEgO and ME_TRgO.
Zhou et al. demonstrated a positive correlation between the infectivity of HCMV virions and the amounts of gH/gL/gO in the virion envelope (25). To assess the effects of gO sequences on infectivity, cell-free virus stocks of the parental wild type and heterologous gO recombinants were analyzed by quantitative PCR (qPCR) to determine the number of virions, and infectivity was determined by a plaque assay. No difference in particles/PFU was observed between TR and the corresponding recombinant TR_MEgO (Fig. 3A) or between ME and the corresponding recombinant ME_TRgO (Fig. 3B). When ME-based HCMV was grown in HFFFtet cells, which repress UL128-131 expression, the resultant virions, ME-T and ME-T_TRgO, were dramatically more infectious, as shown previously (25, 34), but consistently, there were no differences due to the isoform of gO expressed (Fig. 3B). In parallel analyses, the amounts of gH/gL complexes were analyzed by nonreducing immunoblot probing for gL to detect intact, disulfide-linked gH/gL/gO and disulfide-linked gH/gL/UL128 (note that UL130 and UL131 are not disulfide linked to the intact pentamer complex and are thus separated by SDS-PAGE) (Fig. 4). Consistent with our previous reports (25, 30), TR virions contained much larger amounts of total gH/gL, mostly in the form of gH/gL/gO, whereas ME virions contained less gH/gL, mostly as gH/gL/UL128-131. Repression of the UL128-131 proteins (ME-T) drastically reduced the amount of gH/gL/UL128-131 and increased the amount of gH/gL/gO. However, note that the amount of gH/gL/gO in ME-T virions was still smaller than the amount of gH/gL/UL128-131 in ME virions, indicating that the repression of UL128-131 was not fully compensated for by gO. In no case did the expression of the heterologous gO isoform detectably influence the amounts of gH/gL complexes in HCMV virions. Together, these results suggest that the amino acid sequence differences between TR and ME gO do not influence gH/gL complex assembly or the function of gO in entry into fibroblasts.
FIG 3.
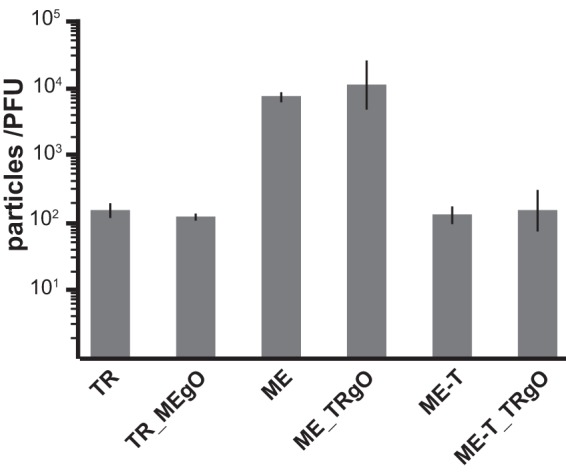
Specific infectivity of parental and TR-ME heterologous gO recombinants. Extracellular virions of TR, TR_MEgO, ME, ME_TRgO, ME-T, or ME-T_TRgO were analyzed by quantitative PCR for viral genomes, and PFU were determined by a plaque assay on nHDF. Average particle/PFU ratios from at least 4 independent experiments are plotted. Error bars represent standard deviations.
FIG 4.
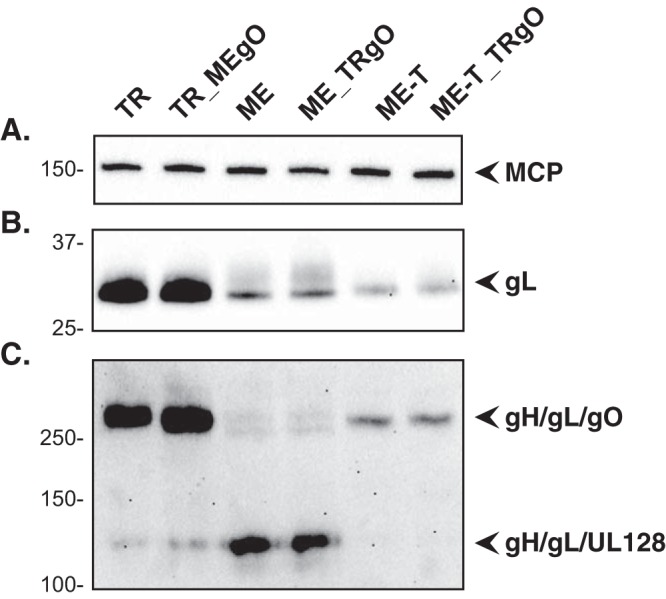
Immunoblot analysis of gH/gL complexes in parental viruses and TR-ME heterologous gO recombinants. Extracellular virion extracts of TR, TR_MEgO, ME, ME_TRgO, ME-T, or ME-T_TRgO were separated by reducing (A and B) or nonreducing (C) SDS-PAGE and analyzed by immunoblot probing for major capsid protein (A) or gL (B and C).
ME expresses less gO during replication than does TR.
The heterologous gO recombinants allowed comparison of gO expression levels between TR and ME. In the first analyses, cells infected with the parental virus or the heterologous gO recombinants were analyzed by reducing immunoblot analysis using TR- and ME-specific anti-gO antibodies (30) (Fig. 5). TR-specific gO antibodies detected two bands in TR-infected cells, a prominent species migrating just above the 100-kDa marker and a minor, more diffuse species migrating at approximately 130 to 140 kDa. The ME-specific antibodies detected similarly migrating bands in TR_MEgO-infected cells; however, their relative abundances appeared more equal. No similar bands were detected in cells infected with ME or ME_TRgO analyzed with either gO antiserum. The failure to detect either isoform of gO in cells infected with ME-based HCMV suggested that the protein expression level from the UL74 locus of ME was lower than that in TR.
FIG 5.
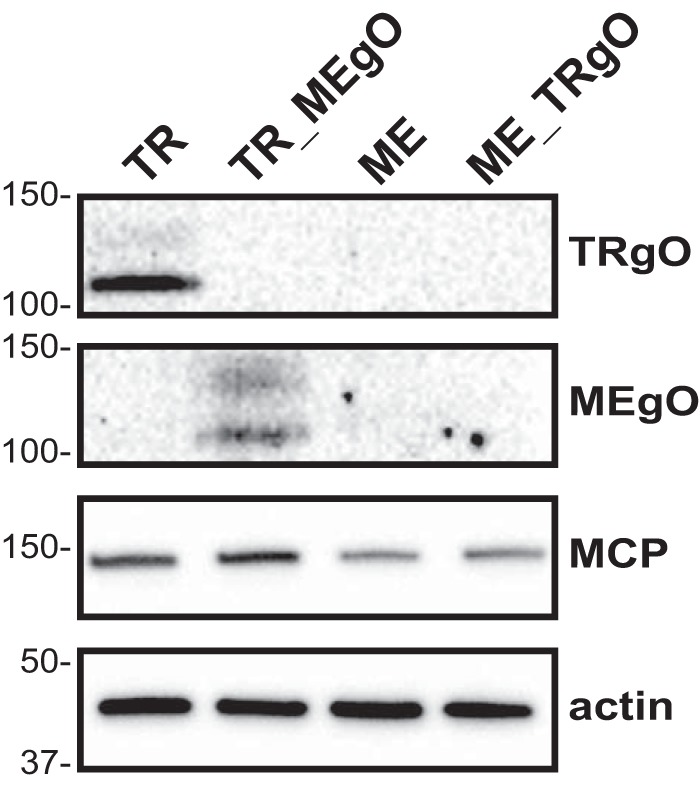
Immunoblot analysis of gO expression in cells infected with parental viruses and TR-ME heterologous gO recombinants. nHDF were infected with 1 PFU/cell of TR, TR_MEgO, ME, or ME_TRgO. At day 5, total cell extracts of infected cells were separated by reducing SDS-PAGE and analyzed by immunoblot probing for TRgO, MEgO, MCP, or actin.
To directly compare differences in glycoprotein expression between TR and ME, infected cells were labeled with [35S]methionine-cysteine for 15 min and then analyzed by immunoprecipitation with antipeptide antibodies specific for gH, gL, or gO, followed by SDS-PAGE and band density analysis (Fig. 6 and Tables 1 and 2). Two approaches were taken to allow direct quantitative comparisons of labeled proteins between extracts. First, cell extracts were denatured and reduced with SDS-dichlorodiphenyltrichloroethane (DDT) prior to immunoprecipitation to allow maximum epitope access by the antipeptide antibodies. Second, for each analysis, multiple immunoprecipitation reactions were performed in parallel with increasing amounts of protein extract input to ensure that antibodies were not limiting. In these experiments, expression levels of gH were nearly identical between TR and ME, and the gL expression level was approximately 4-fold higher for TR than for ME, but the gO expression level was strikingly 27-fold higher for TR than for ME (Fig. 6A and Table 1). To address the possibility that the MEgO-specific antibodies were simply less efficient at capturing MEgO from ME extracts, similar experiments were performed with the TR-ME heterologous gO recombinants (Fig. 6B and Table 2). Again, gH and gL levels were similar between TR_MEgO and ME_TRgO, but gO levels were approximately 20-fold higher for the TR-based virus. To address the hypothesis that differences in gO expression between TR and ME reflect differences in protein turnover, the [35S]methionine-cysteine label was chased for up to 6 h (Fig. 7). The patterns of gH detection over the chase time were very similar for both TR and ME samples. In both cases, the amounts of labeled gH dropped to 60% after 3 h and to 30 to 40% after 6 h. The pattern of gO detection for both TR and ME was comparable to that of gH detection. Together, these results confirmed that ME-infected cells express less gO than do TR-infected cells and suggested differences in early steps of expression, such as mRNA transcription, translation, or rapid ER-associated degradation, which can degrade proteins in the time scale of minutes (35).
FIG 6.
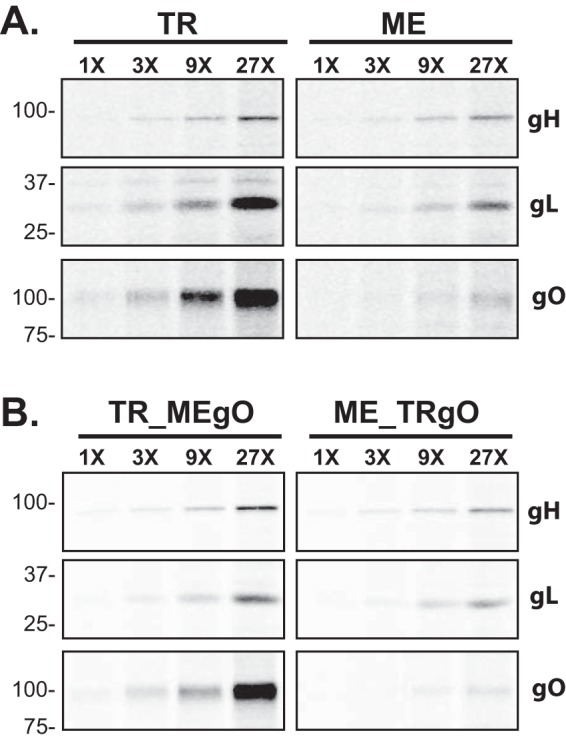
Quantitative comparison of glycoprotein expression in TR- and ME-infected cells. nHDF were infected with 1 PFU/cell of TR or ME (A) or TR_MEgO or ME_TRgO (B). At 5 dpi, infected cells were metabolically labeled with [35S]cysteine-methionine for 15 min, and membrane proteins were extracted in 1% Triton X-100. All samples were adjusted to 2% SDS–30 mM DTT, heated to 75°C for 10 min, cooled to room temperature, and then diluted 35-fold. Parallel immunoprecipitations were performed, in which equal amounts of anti-gH, gL, or gO (TR- or ME-specific) antibodies were reacted with 3-fold-increasing amounts of protein extract as the input, and precipitated proteins were analyzed by SDS-PAGE.
TABLE 1.
Quantitative comparison of glycoprotein expression in TR- and ME-infected cellsd
| Immunoprecipitation antibodya | Extract inputb (ml) | Strain |
Fold differencef | Mean fold difference (±SD)g | |||
|---|---|---|---|---|---|---|---|
| TR |
ME |
||||||
| Densityc | Adjusted densitye | Density | Adjusted density | ||||
| Anti-gH | 0.04 | ND | ND | ND | ND | ND | 1.4 (±0.1) |
| 0.13 | 136.7 | 4.6 | 106.6 | 3.4 | 1.3 | ||
| 0.40 | 476.9 | 15.9 | 337.8 | 10.9 | 1.5 | ||
| 1.20 | 1,200.7 | 40.0 | 872.9 | 28.2 | 1.4 | ||
| Anti-gL | 0.04 | 143.8 | 11.1 | ND | ND | ND | 4.0 (±1.2) |
| 0.13 | 679.1 | 52.2 | 127.2 | 9.8 | 5.3 | ||
| 0.40 | 1,627.3 | 125.2 | 509.3 | 39.2 | 3.2 | ||
| 1.20 | 6,071.3 | 467.0 | 1,809.9 | 139.2 | 3.4 | ||
| Anti-gO | 0.04 | 267.0 | 12.1 | ND | ND | ND | 27.2 (±9.4) |
| 0.13 | 1,008.6 | 45.8 | ND | ND | ND | ||
| 0.40 | 3,805.4 | 173.0 | 120.8 | 5.0 | 34.4 | ||
| 1.20 | 9,251.8 | 420.5 | 478.9 | 20.0 | 21.1 | ||
Seven microliters of rabbit antipeptide serum per immunoprecipitation reaction mixture.
Preparation of radiolabeled cell extracts is described in the legend to Fig. 6 and in Materials and Methods.
Pixel density of bands shown in Fig. 6A as determined using ImageJ version 1.48.
ND, band density not detected.
Density divided by the predicted number of methionine (met) and cysteine (cys) residues: TRgH (17 met, 13 cys), MEgH (17 met, 14 cys), TRgL (3 met, 10 cys), MEgL (3 met, 10 cys), TRgO (16 met, 6 cys), MEgO (18 met, 6 cys).
Adjusted density of TR divided by adjusted density of ME.
Average fold difference between TR and ME ± standard deviation.
TABLE 2.
Quantitative comparison of glycoprotein expression in TR_MEgO- and ME_TRgO-infected cellsd
| Immunoprecipitation antibodya | Extract inputb (ml) | Strain |
Fold differencef | Mean fold difference (±SD)g | |||
|---|---|---|---|---|---|---|---|
| TR_MEgO |
ME_TRgO |
||||||
| Densityc | Adjusted densitye | Density | Adjusted density | ||||
| Anti-gH | 0.04 | 68.4 | 2.3 | 29.5 | 1.0 | 2.4 | 1.6 (±0.5) |
| 0.13 | 181.6 | 6.1 | 163.1 | 5.3 | 1.1 | ||
| 0.40 | 539.5 | 18.0 | 410.6 | 13.2 | 1.4 | ||
| 1.20 | 1,697.7 | 56.6 | 1,064.0 | 34.3 | 1.6 | ||
| Anti-gL | 0.04 | ND | ND | ND | ND | ND | 1.5 (±0.4) |
| 0.13 | 196.6 | 15.1 | 153.8 | 11.8 | 1.3 | ||
| 0.40 | 645.0 | 49.6 | 508.3 | 39.1 | 1.3 | ||
| 1.20 | 2,547.5 | 196.0 | 1,269.6 | 97.7 | 2.0 | ||
| Anti-gO | 0.04 | 187.8 | 7.8 | ND | ND | ND | 19.7 (±1.7) |
| 0.13 | 945.8 | 39.4 | ND | ND | ND | ||
| 0.40 | 2,580.3 | 107.5 | 127.8 | 5.8 | 18.5 | ||
| 1.20 | 10,502.7 | 437.6 | 460.1 | 20.9 | 20.9 | ||
Seven microliters of rabbit antipeptide serum per immunoprecipitation reaction mixture.
Preparation of radiolabeled cell extracts is described in the legend to Fig. 6 and in Materials and Methods.
Pixel density of bands shown in Fig. 6B as determined using ImageJ version 1.48.
ND, band density not detected.
Density divided by the predicted number of methionine (met) and cysteine (cys) residues: TRgH (17 met, 13 cys), MEgH (17 met, 14 cys), TRgL (3 met, 10 cys), MEgL (3 met, 10 cys), TRgO (16 met, 6 cys), MEgO (18 met, 6 cys).
Adjusted density of TR_MEgO divided by adjusted density of ME_TRgO.
Average fold difference between TR_MEgO and ME_TRgO ± standard deviation.
FIG 7.
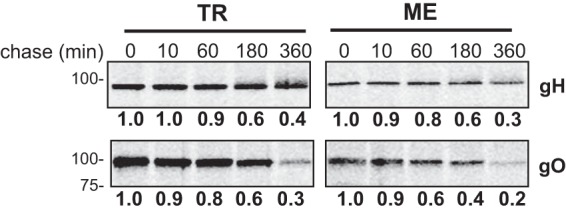
Analysis of glycoprotein turnover in TR- and ME-infected cells. nHDF were infected with 1 PFU/cell of TR or ME. At 5 dpi, infected cells were metabolically labeled with [35S]cysteine-methionine for 15 min, and the label was then chased for 0, 10, 60, 180, or 360 min. Membrane proteins were extracted in 1% Triton X-100, adjusted to 2% SDS–30 mM DTT, heated to 75°C for 10 min, cooled to room temperature, and then diluted 35-fold. Immunoprecipitation was performed with anti-gH and -gO (TR- or ME-specific) antibodies, and precipitated proteins were analyzed by SDS-PAGE. Band densities were determined relative to the 0-min chase time. Results shown are representative of data from 4 independent experiments.
Overexpression of gO during ME replication increases gH/gL/gO assembly and virus infectivity.
To directly test the hypothesis that the low abundance of gH/gL/gO in ME virions was due not simply to competition from the UL128-131 proteins but also to low gO expression, adenovirus (Ad) vectors were used to increase gO levels during ME replication. Ad vectors expressing green fluorescent protein (GFP) were used to control for potential effects of the Ad vectors themselves. Consistent with data from the above-described analyses, gO levels were below the limits of immunoblot detection in ME-infected nHDF or HFFFtet cells, but gO was readily detected in cells superinfected with AdMEgO (Fig. 8A). The overall expression of gL in ME-infected cells was reduced by the presence of either Ad vector (Fig. 8A). In the case of the control AdGFP, the lower intracellular gL level correlated with reduced levels of gH/gL/gO complexes in virions from HFFFtet cells (ME-T) (Fig. 8B), and this in turn correlated with reduced infectivity (i.e., increased particle/PFU ratio) (Fig. 9). The “Ad effect” on virion gH/gL levels and infectivity was less apparent in human foreskin fibroblast (HFF) cells (ME), perhaps masked by the overall larger amounts of gH/gL and the much lower infectivity of these virions (Fig. 8B and 9). Controlling for the Ad effect, AdMEgO expression in HFFFtet cells increased the amounts of gH/gL/gO in ME-T virions compared to AdGFP, and this resulted in a 6-fold enhancement of infectivity, beyond the 40-fold enhanced infectivity resulting from the repression of UL128-131 alone (Fig. 8B and 9). In contrast, AdMEgO expression had little effect on the virions from HFF cells.
FIG 8.
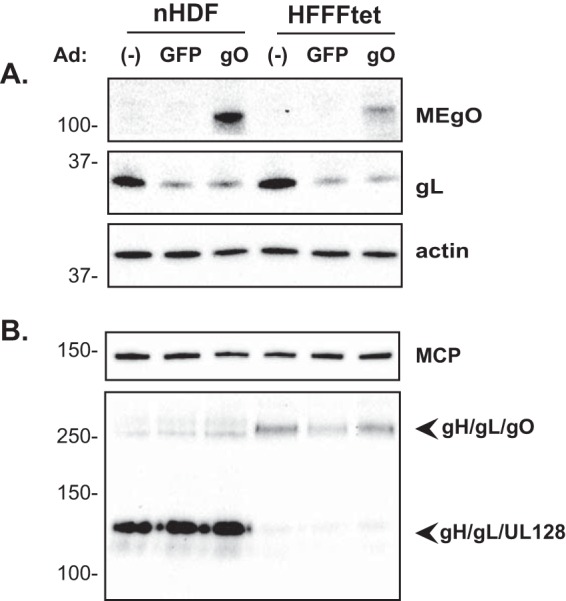
Ad vector overexpression of gO during ME replication. nHDF or HFFFtet cells were infected with ME for 2 days and then superinfected with Ad vectors expressing either GFP or MEgO for an additional 4 days. Extracts of infected cells (A) or extracellular virions (B) were separated by reducing (A and B, top) or nonreducing (B, bottom) SDS-PAGE and analyzed by immunoblot probing for MEgO, actin, MCP, or gL, as indicated to the right.
FIG 9.
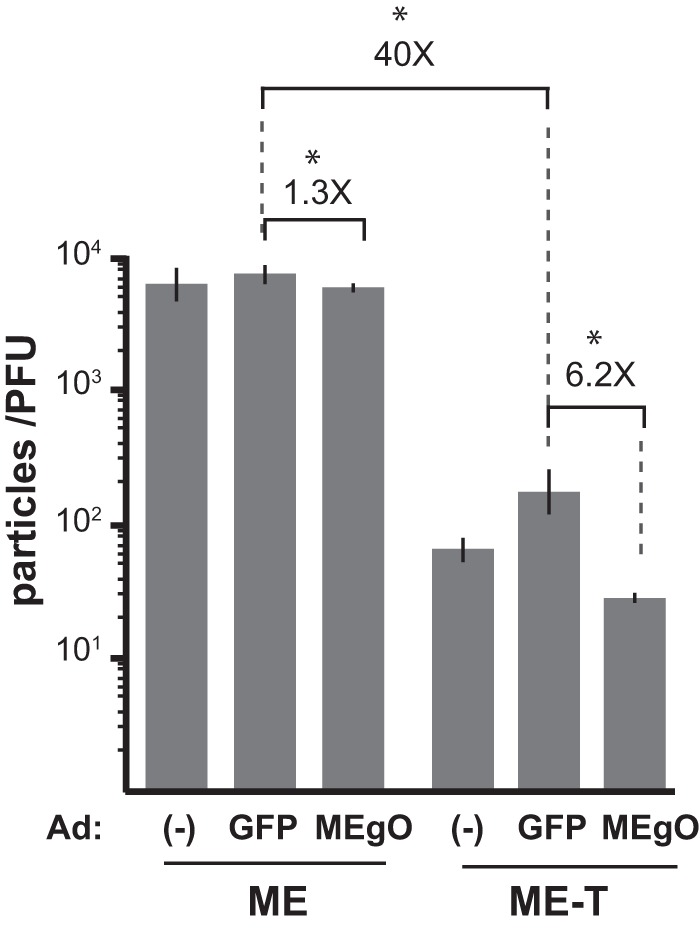
Specific infectivity of ME virions produced under conditions of gO overexpression. nHDF or HFFFtet cells were infected with ME for 2 days and then superinfected with Ad vectors expressing either GFP or MEgO for an additional 4 days. Extracellular virions from nHDF (ME) or HFFFtet (ME-T) cells were analyzed by quantitative PCR for viral genomes, and PFU were determined by a plaque assay on nHDF. Shown are average particle/PFU ratios of virions produced in 2 independent experiments, each analyzed in triplicate. Error bars represent the standard deviations. Asterisks above fold differences indicate a P value of <0.03 (determined by Student's unpaired t test [2 tailed]).
DISCUSSION
Recent population genetic studies have demonstrated a greater degree of genetic diversity of HCMV in clinical specimens than had been previously appreciated (36–38). The cell type and propagation methods likely narrow the resultant genotypes by purifying selection (39, 40). During propagation in cultured fibroblasts, inactivating mutations in the UL128-131 ORFs are rapidly selected in a BAC clone of ME, and this selective pressure can be relieved by transcriptional repression of the UL131 promoter, which reduces the expression of pentameric gH/gL/UL128-131 (34). In contrast, the UL128-131 ORFs are more stable in BAC clones of strains TR and TB (40, 41). The UL128-131 ORF of TB contains a single nucleotide polymorphism (SNP) relative to ME that reduces the splicing of the mRNA encoding the UL128 protein, which may help stabilize the UL128-131 ORFs through reduced expression of gH/gL/UL128-131 (41). However, TR is identical to ME at this nucleotide position, and recombinant ME in which the UL128-131 locus was replaced with the UL128-131 sequences from TR was as sensitive to the selective inactivation of the locus as wild-type ME (41). Together, these observations suggest that factors beyond the expression levels of the UL128-131 proteins can influence the selective pressures on the UL128-131 ORFs.
The results reported here demonstrated that TR and ME differ in the stoichiometry of expression of gO and UL128-131, and this seems to be a major factor determining the abundances of gH/gL/gO and gH/gL/UL128-131 in the virion envelope and the infectivity of cell-free virions. The steady-state levels of gH/gL in fibroblasts infected with TR and ME were found to be comparable, but ME-infected cells contained more UL128-131 than did TR-infected cells. In ME-infected cells, most of the gH/gL was in an ER-associated form, whereas TR-infected cells contained a large amount of Golgi compartment-associated gH/gL. This correlated well with previous observations that TR contained more total gH/gL than did ME virions (25, 30). The amount of Golgi compartment-associated gH/gL in ME-infected cells was reduced when the expression of the UL128-131 proteins was repressed, consistent with the observation that most of the gH/gL in ME virions was in the form of gH/gL/UL128-131 (25, 30). Comparison of gO expression levels between strains was complicated because the amino acid sequence differences between genotypes affected antibody recognition (30). To circumvent this caveat, HCMV recombinants were engineered, in which the UL74(gO) ORFs of TR were replaced with the homologous sequences of ME, and vice versa. This approach allowed the analysis of the expression of both gO isoforms in both genetic backgrounds, eliminating the possibility that the results were due to differences in antibody-antigen affinities. Immunoblot and radiolabeling experiments clearly demonstrated that ME-infected cells contained less gO than did TR-infected cells. The overexpression of gO during ME replication had no effect on the levels of gH/gL/gO or the infectivity of the virions unless UL128-131 proteins were also transcriptionally repressed, and even then, gH/gL/gO levels and infectivity were only modestly enhanced. Together, these results underscore the competition between gO and UL128-131 for binding to gH/gL and suggest that other factors may influence the efficiency of gH/gL/gO assembly.
The molecular mechanisms underpinning the discrepancy between TR and ME in the expression UL128-131 and gO remain unclear. As mentioned above, Murrell et al. described a SNP in the TB UL128-131 locus that affected mRNA splicing, in part explaining the lower expression levels of these proteins in TB (41). However, this splicing effect does not explain the difference in UL128-131 expression levels between TR and ME, since this nucleotide position is conserved between these strains. For gO, the radiolabeling analyses reported in Fig. 6 and 7 suggest that the differences are due to early events in UL74(gO) expression, such as transcription, mRNA processing/stability, translation, or rapid ER-associated degradation, occurring in the time scale of minutes (35). Attempts to analyze UL74(gO) mRNA levels between TR and ME by quantitative reverse transcription-PCR (RT-PCR) were complicated by the fact that HCMV genomes contain many overlapping RNA polymerase II (RNAPII) transcription units that vary between strains (42, 43). It is interesting that ME-infected cells contained less UL148 than did TR-infected cells. UL148 was first described as an ER-resident chaperone protein that promotes the assembly of gH/gL/gO (33). The mechanism may well involve interactions between UL148 and the cellular ER-associated degradation pathway (C. Nguyen, M. Siddiquey, H. Zhang, and J. Kamil, presented at the 42nd International Herpesvirus Workshop, Ghent, Belgium, 2017).
The TR-ME heterologous gO recombinant viruses also allowed analysis of the effects of gO amino acid sequence differences on the assembly of gH/gL complexes and the function of gO in entry. No differences were observed between TR and TR_MEgO or between ME and ME_TRgO in either the amounts of gH/gL complexes in virions or cell-free infectivity. These results argue against the notion that the amino acid sequence differences between gO genotypes affect interactions with gH/gL or the binding of the fibroblast entry receptor PDGFRα. Interestingly, Kalser et al. showed that replacing the endogenous gO protein of TB with the gO protein from Towne did not alter replication in cultured fibroblasts but enhanced replication in epithelial cell cultures (44). Thus, it may be that gO sequence variation affects interactions with receptors other than PDGFRα that mediate infection of epithelial cells.
Laib-Sampaio et al. reported that mutational disruption of UL74(gO) expression in ME had little effect on replication unless the UL128-131 locus was also disrupted (24). Those authors suggested that the spread of ME was mediated principally by gH/gL/UL128-131 in a cell-associated manner, but when UL128-131 was inactivated, spread could also occur in a cell-free manner, mediated by gH/gL/gO. This is in stark contrast to the dramatic phenotype reported for a gO-null TR mutant (22). Our finding that the level of expression of gO by ME is low compared to that by TR may provide a partial explanation for these different gO-null phenotypes.
It remains unclear whether the described difference in gO expression between TR and ME represents a bona fide variation that naturally exists between HCMV genotypes in vivo or reflects differential selection on de novo mutations that occurred during the independent isolation of these strains from clinical specimens. It seems clear that serial propagation of ME in cultured fibroblasts selects for de novo mutations that reduce or abolish the robust expression of the UL128-131 proteins (34, 40). The selective pressure that fixes these mutations in the culture population may be explained by data from the specific infectivity analyses reported here (Fig. 3 and 9) and by Zhou et al. (25). In both analyses, the specific infectivity of TR was measured at approximately 100 to 200 particles/PFU, whereas ME was 30- to 50-fold less infectious. Repression of the UL128-131 proteins enhanced the infectivity of ME (“ME-T”) to levels comparable to those of TR (approximately 100 particles/PFU). While the infectivities of ME-T and TR virions were comparable, ME-T virions still contained far less gH/gL/gO than did TR virions (Fig. 4) (25). Ad vector overexpression of gO enhanced the infectivity of ME only 6-fold beyond the enhancement due to UL128-131 repression alone (Fig. 8 and 9). Together, these observations seem to suggest that in vitro selective pressures for reduced UL128-131 expression are much more pronounced than any for enhanced gO expression. Thus, it is possible that the difference in gO expression between HCMV TR and ME is derived not from the selection of de novo mutations occurring during propagation in culture but from nonselective, random sampling of the multitude of different genotypes that likely preexist in clinical specimens (36–38). Distinguishing between these possibilities will require clear identification of the genomic sequences that determine gO expression levels.
MATERIALS AND METHODS
Cell lines.
Primary neonatal human dermal fibroblasts (nHDF; Thermo Fisher Scientific), MRC-5 fibroblasts (ATCC CCL-171; American Type Culture Collection), and HFFFtet cells (which express the tetracycline [Tet] repressor protein; provided by Richard Stanton) (34) were grown in Dulbecco's modified Eagle's medium (DMEM; Thermo Fisher Scientific) supplemented with 6% heat-inactivated fetal bovine serum (FBS; Rocky Mountain Biologicals, Inc., Missoula, MT, USA) and 6% bovine growth serum (BGS; Rocky Mountain Biologicals, Inc., Missoula, MT, USA).
Human cytomegaloviruses.
All HCMV strains were derived from bacterial artificial chromosome (BAC) clones. The BAC clone of TR was provided by Jay Nelson (Oregon Health and Sciences University, Portland, OR, USA) (45). The BAC clone of Merlin (ME) (pAL1393), which carries tetracycline operator sequences in the transcriptional promoter of UL130 and UL131, was provided by Richard Stanton. (34). Infectious HCMV was recovered by electroporation of BAC DNA into MRC-5 fibroblasts, as described previously by Wille et al. (22). Cell-free HCMV stocks were produced by infecting HFF or HFFFtet cells at 2 PFU per cell. At 8 to 10 days postinfection (when cells were still visually intact), culture supernatants were harvested, and cellular contaminants were removed by centrifugation at 1,000 × g for 10 min and again at 6,000 × g for 10 min. Stocks were judged to be cell free by the lack of calnexin and actin by Western blot analyses and then stored at −80°C. Freeze-thaw cycles were avoided. PFU were determined by plating a series of 10-fold dilutions of each stock onto replicate cultures of HFF for 2 h at 37°C and replacing the inoculum with DMEM supplemented with 5% FBS and 0.6% SeaPlaque agarose (to limit cell-free spread). Plaques were counted by light microscopy 3 weeks after infection.
Heterologous UL74(gO) recombinant HCMV.
A two-step BAC recombineering process was performed as previously described (34). In the first step, the endogenous UL74 ORF from the start codon to the stop codon of both TR and ME was replaced by a selectable marker. Briefly, cultures of Escherichia coli SW102 containing either the BAC clone of TR or ME were grown at 32°C until an optical density at 600 nm (OD600) of 0.55 was reached. Recombination genes were induced by incubation at 42°C for 15 min. The purified PCR product containing the KanR/LacZ/RpsL selectable marker cassette flanked by sequences homologous to 80 bp upstream and downstream of the TR or ME UL74 ORF was electroporated into the bacteria, and cultures were incubated for 1 h at 32°C and then selected on medium containing kanamycin (15 μg/ml), isopropyl-β-d-thiogalactopyranoside (IPTG) (50 μM), 5-bromo-4-chloro-3-indolyl-β-d-galactopyranoside (X-gal) (20 μg/ml), and chloramphenicol (12.5 μg/ml). First-step primer sequences were 5′-CTTGGTGGACTATGCTTAACGCTCTCATTCTCATGGGAGCTTTTTGTATCGTATTACGACATTGCTGTTTCCAGAACTCCTGTGACGGAAGATCACTTCG-3′ and 5′-CGACCAGAATCAGCAGTGAGTACACGCAGGCAAGCCAAACCACAAGGCAGACGGACGGTGCGGGGTCTCCTCCTCTGTCCTGAGGTTCTTATGGCTCTTG-3′ for TR and 5′-CCTGGTGGACTATGCTTAACGCTCTCATTCTGATGGGAGCTTTTTGTATCGTATTACGACATTGCTGCTTCCAGAACTCCTGTGACGGAAGATCACTTCG-3′ and 5′-CGACCAGAATCAGCAGTGAGTACACGCAGGCAAACCAAACCACAAGGCAGACGGACGGTGCGGGGTCTCCTCCTCTGTACTGAGGTTCTTATGGCTCTTG-3′ for ME.
In the second step, the selectable marker cassette in the TR and ME first-step intermediate BACs was replaced with the UL74(gO) sequence from the heterologous strain. Briefly, E. coli cultures were prepared for recombination as described above for step 1 and electroporated with purified PCR products containing the UL74 ORF from the TR or ME strain flanked by sequence homologous to 80 bp upstream and downstream of the opposite strain. Transformed E. coli cells were selected for the removal of the KanR/LacZ/RpsL cassette by growth on medium containing streptomycin (1.5 mg/ml), IPTG (50 μM), X-gal (20 μg/ml), and chloramphenicol (12.5 μg/ml). Primers used to generate the second-step PCR product were 5′-GCCTGGTGGACTATGCTTAACGCTCTCATTCTGATGGGAGCTTTTTGTATCGTATTACGACATTGCTGCTTCCAGAACTTTACTGCAACCACCACCAAAG-3′ and 5′-CGACCAGAATCAGCAGTGAGTACACGCAGGCAAACCAAACCACAAGGCAGACGGACGGTGCGGGGTCTCCTCCTCTGTAATGGGGAGAAAAGGAGAGATG-3′ for the transformation of TR UL74 into ME and 5′-GGCTTGGTGGACTATGCTTAACGCTCTCATTCTCATGGGAGCTTTTTGTATCGTATTACGACATTGCTGTTTCCAGAACTTTACTGCGACCACCACCAAA-3′ and 5′-CAGAATCAGCAGTGAGTACACGCAGGCAAGCCAAACCACAAGGCAGACGGACGGTGCGGGGTCTCCTCCTCTGTCATGGGGAAAAAAGAGATGATAATGG for the transformation of ME UL74 into TR.
The final heterologous UL74(gO) recombinants were verified by Sanger sequencing of PCR products using the following primers: 5′-GATGATTTTTACAAGGCACATTGTACATC-3′ and 5′-AACTAGGTCGTCTTGGAAGC-3′ for TRΔMEgO and 5′-CTCACAATGATTTTTACAATGCG-3′ and 5′-AACTAGGTCGTCTTGGAAGC-3′ for MEΔTRgO.
Antibodies.
Rabbit polyclonal antipeptide antibodies specific for TBgO and MEgO were described previously (30). Rabbit polyclonal antibodies specific for UL148 were described previously (33). Rabbit polyclonal anti-peptide antibodies against gH, gL, UL130, and UL131 were provided by David Johnson (Oregon Health and Sciences University, Portland, OR, USA) (46). Anti-UL128 monoclonal antibody (MAb) 4B10 was provided by Tom Shenk (Princeton University, Princeton, NJ, USA) (47). MAb 28-4 directed against major capsid protein (MCP) and MAb 27-156 directed against gB were provided by Bill Britt (48, 49). MAb CH160 against cytomegalovirus (CMV) immediate early protein 1 (IE1) and IE2 was purchased from Abcam (Cambridge, MA, USA).
Immunoblotting.
HCMV-infected cells or cell-free virions were solubilized in 2% SDS–20 mM Tris-buffered saline (TBS) (pH 6.8). Insoluble material was cleared by centrifugation at 16,000 × g for 15 min, and extracts were then boiled for 10 min. For endoglycosidase H (endo H) or peptide N-glycosidase F (PNGase F) treatment assays, proteins were extracted in 1% Triton X-100 (TX100) plus 0.5% sodium deoxycholate (DOC) in 20 mM Tris (pH 6.8) plus 100 mM NaCl (TBS-TX-DOC). Extracts were clarified by centrifugation at 16,000 × g for 15 min and treated with endo H or PNGase F according to the manufacturer's instructions (New England BioLabs). For reducing blots, dithiothreitol (DTT) was added to extracts to a final concentration of 25 mM. After separation by SDS-PAGE, proteins were transferred onto polyvinylidene difluoride (PVDF) membranes (Whatman) in a buffer containing 10 mM NaHCO3 and 3 mM Na2CO3 (pH 9.9) plus 10% methanol. Transferred proteins were probed with MAbs or rabbit polyclonal antibodies, anti-rabbit or anti-mouse secondary antibodies conjugated with horseradish peroxidase (Sigma-Aldrich), and Pierce ECL-Western blotting substrate (Thermo Fisher Scientific). Chemiluminescence was detected using a Bio-Rad ChemiDoc MP imaging system.
Radiolabeling proteins.
Cell cultures were incubated in labeling medium (Met-Cys-free DMEM plus 2% dialyzed FBS lacking methionine and cysteine) for 2 h at 37°C, and [35S]methionine-cysteine was then added to 1 mCi/ml (EasyTag Express 35S protein labeling mix; PerkinElmer). For chase experiments, label medium was removed, and cultures were washed twice in DMEM plus 2% FBS supplemented with a 10-fold excess of nonradioactive methionine and cysteine and then incubated in this medium for the indicated times.
Immunoprecipitation.
Cell extracts were harvested in TBS-TX-DOC supplemented with 0.5% bovine serum albumin (BSA) and 1 mM phenylmethylsulfonyl fluoride (PMSF), clarified by centrifugation at 16,000 × g for 15 min, adjusted to 2% SDS–30 mM DTT, and heated at 75°C for 15 min. The extracts were then diluted 35-fold with TBS-TX-DOC supplemented with 0.5% BSA and 10 mM iodoacetamide, incubated on ice for 15 min, and precleared with protein A-agarose beads (Invitrogen/Thermo Fisher Scientific) for at 4°C for 2 h. Immunoprecipitation reactions were set up with specific antibodies and protein A-agarose beads, and the mixtures were incubated overnight at 4°C. Protein A-agarose beads were washed 3 times with TBS-TX-DOC, and proteins were eluted with 2% SDS and 30 mM DTT in TBS at room temperature (RT) for 15 min, followed by 75°C for 10 min. Eluted proteins were separated by SDS-PAGE and analyzed with a Typhoon FLA-9500 imager (GE Healthcare Life Sciences). Band densities were determined using ImageJ version 1.48 software.
Quantitative PCR.
Viral genomes were determined as described previously (25). Briefly, cell-free HCMV stocks were treated with DNase I before extraction of viral genomic DNA (PureLink viral RNA/DNA minikit; Life Technologies/Thermo Fisher Scientific). Primers specific for sequences within UL83 were used with the MyiQ real-time PCR detection system (Bio-Rad).
Superinfection of HCMV-infected cells with replication-defective adenovirus vectors.
The construction of Ad vectors expressing MEgO or GFP was described previously (30). Two days after HCMV infection, cells were superinfected with 20 PFU/cell of AdMEgO or AdGFP. Six days later, cell-free HCMV was collected from the supernatant culture by centrifugation, and cells were harvested for immunoblotting.
ACKNOWLEDGMENTS
We are grateful to Bill Britt, David Johnson, Jay Nelson, and Tom Shenk for generously supplying HCMV BAC clones, antibodies, and cell lines, as indicated in Materials and Methods, and members of the Ryckman laboratory for support and insightful discussions.
This work was supported by a grant from the National Institutes of Health to B.J.R. (R01AI097274).
Experiments were designed by B.J.R., L.Z., and M.Z. and performed by L.Z. and M.Z. Data were analyzed and the manuscript was prepared by B.J.R., L.Z., J.K., and R.S.
REFERENCES
- 1.Boppana SB, Fowler KB, Pass RF, Rivera LB, Bradford RD, Lakeman FD, Britt WJ. 2005. Congenital cytomegalovirus infection: association between virus burden in infancy and hearing loss. J Pediatr 146:817–823. doi: 10.1016/j.jpeds.2005.01.059. [DOI] [PubMed] [Google Scholar]
- 2.Britt WJ. 2008. Manifestations of human cytomegalovirus infection: proposed mechanisms of acute and chronic disease. Curr Top Microbiol Immunol 325:417–470. [DOI] [PubMed] [Google Scholar]
- 3.Griffiths P, Baraniak I, Reeves M. 2015. The pathogenesis of human cytomegalovirus. J Pathol 235:288–297. doi: 10.1002/path.4437. [DOI] [PubMed] [Google Scholar]
- 4.Streblow DN, Orloff SL, Nelson JA. 2007. Acceleration of allograft failure by cytomegalovirus. Curr Opin Immunol 19:577–582. doi: 10.1016/j.coi.2007.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cannon MJ, Hyde TB, Schmid DS. 2011. Review of cytomegalovirus shedding in bodily fluids and relevance to congenital cytomegalovirus infection. Rev Med Virol 21:240–255. doi: 10.1002/rmv.695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Plachter B, Sinzger C, Jahn G. 1996. Cell types involved in replication and distribution of human cytomegalovirus. Adv Virus Res 46:195–261. doi: 10.1016/S0065-3527(08)60073-1. [DOI] [PubMed] [Google Scholar]
- 7.Sinzger C, Grefte A, Plachter B, Gouw AS, The TH, Jahn G. 1995. Fibroblasts, epithelial cells, endothelial cells and smooth muscle cells are major targets of human cytomegalovirus infection in lung and gastrointestinal tissues. J Gen Virol 76:741–750. doi: 10.1099/0022-1317-76-4-741. [DOI] [PubMed] [Google Scholar]
- 8.Sinzger C, Jahn G. 1996. Human cytomegalovirus cell tropism and pathogenesis. Intervirology 39:302–319. doi: 10.1159/000150502. [DOI] [PubMed] [Google Scholar]
- 9.Sinzger C, Digel M, Jahn G. 2008. Cytomegalovirus cell tropism. Curr Top Microbiol Immunol 325:63–83. [DOI] [PubMed] [Google Scholar]
- 10.Xu S, Schafer X, Munger J. 2016. Expression of oncogenic alleles induces multiple blocks to human cytomegalovirus infection. J Virol 90:4346–4356. doi: 10.1128/JVI.00179-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Smith JD. 1986. Human cytomegalovirus: demonstration of permissive epithelial cells and nonpermissive fibroblastic cells in a survey of human cell lines. J Virol 60:583–588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Connolly SA, Jackson JO, Jardetzky TS, Longnecker R. 2011. Fusing structure and function: a structural view of the herpesvirus entry machinery. Nat Rev Microbiol 9:369–381. doi: 10.1038/nrmicro2548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Campadelli-Fiume G, Collins-McMillen D, Gianni T, Yurochko AD. 2016. Integrins as herpesvirus receptors and mediators of the host signalosome. Annu Rev Virol 3:215–236. doi: 10.1146/annurev-virology-110615-035618. [DOI] [PubMed] [Google Scholar]
- 14.Heldwein EE. 2016. gH/gL supercomplexes at early stages of herpesvirus entry. Curr Opin Virol 18:1–8. doi: 10.1016/j.coviro.2016.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ciferri C, Chandramouli S, Donnarumma D, Nikitin PA, Cianfrocco MA, Gerrein R, Feire AL, Barnett SW, Lilja AE, Rappuoli R, Norais N, Settembre EC, Carfi A. 2015. Structural and biochemical studies of HCMV gH/gL/gO and pentamer reveal mutually exclusive cell entry complexes. Proc Natl Acad Sci U S A 112:1767–1772. doi: 10.1073/pnas.1424818112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Stegmann C, Abdellatif ME, Laib Sampaio K, Walther P, Sinzger C. 2017. Importance of highly conserved peptide sites of human cytomegalovirus gO for formation of the gH/gL/gO complex. J Virol 91:e01339-16. doi: 10.1128/JVI.01339-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hahn G, Revello MG, Patrone M, Percivalle E, Campanini G, Sarasini A, Wagner M, Gallina A, Milanesi G, Koszinowski U, Baldanti F, Gerna G. 2004. Human cytomegalovirus UL131-128 genes are indispensable for virus growth in endothelial cells and virus transfer to leukocytes. J Virol 78:10023–10033. doi: 10.1128/JVI.78.18.10023-10033.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ryckman BJ, Jarvis MA, Drummond DD, Nelson JA, Johnson DC. 2006. Human cytomegalovirus entry into epithelial and endothelial cells depends on genes UL128 to UL150 and occurs by endocytosis and low-pH fusion. J Virol 80:710–722. doi: 10.1128/JVI.80.2.710-722.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wang D, Shenk T. 2005. Human cytomegalovirus UL131 open reading frame is required for epithelial cell tropism. J Virol 79:10330–10338. doi: 10.1128/JVI.79.16.10330-10338.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Luo MH, Schwartz PH, Fortunato EA. 2008. Neonatal neural progenitor cells and their neuronal and glial cell derivatives are fully permissive for human cytomegalovirus infection. J Virol 82:9994–10007. doi: 10.1128/JVI.00943-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gerna G, Percivalle E, Lilleri D, Lozza L, Fornara C, Hahn G, Baldanti F, Revello MG. 2005. Dendritic-cell infection by human cytomegalovirus is restricted to strains carrying functional UL131-128 genes and mediates efficient viral antigen presentation to CD8+ T cells. J Gen Virol 86:275–284. doi: 10.1099/vir.0.80474-0. [DOI] [PubMed] [Google Scholar]
- 22.Wille PT, Knoche AJ, Nelson JA, Jarvis MA, Johnson DC. 2010. A human cytomegalovirus gO-null mutant fails to incorporate gH/gL into the virion envelope and is unable to enter fibroblasts and epithelial and endothelial cells. J Virol 84:2585–2596. doi: 10.1128/JVI.02249-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Jiang XJ, Adler B, Sampaio KL, Digel M, Jahn G, Ettischer N, Stierhof YD, Scrivano L, Koszinowski U, Mach M, Sinzger C. 2008. UL74 of human cytomegalovirus contributes to virus release by promoting secondary envelopment of virions. J Virol 82:2802–2812. doi: 10.1128/JVI.01550-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Laib Sampaio K, Stegmann C, Brizic I, Adler B, Stanton RJ, Sinzger C. 2016. The contribution of pUL74 to growth of human cytomegalovirus is masked in the presence of RL13 and UL128 expression. J Gen Virol 97:1917–1927. doi: 10.1099/jgv.0.000475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Zhou M, Lanchy JM, Ryckman BJ. 2015. Human cytomegalovirus gH/gL/gO promotes the fusion step of entry into all cell types, whereas gH/gL/UL128-131 broadens virus tropism through a distinct mechanism. J Virol 89:8999–9009. doi: 10.1128/JVI.01325-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kabanova A, Marcandalli J, Zhou T, Bianchi S, Baxa U, Tsybovsky Y, Lilleri D, Silacci-Fregni C, Foglierini M, Fernandez-Rodriguez BM, Druz A, Zhang B, Geiger R, Pagani M, Sallusto F, Kwong PD, Corti D, Lanzavecchia A, Perez L. 2016. Platelet-derived growth factor-alpha receptor is the cellular receptor for human cytomegalovirus gHgLgO trimer. Nat Microbiol 1:16082. doi: 10.1038/nmicrobiol.2016.82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Stegmann C, Hochdorfer D, Lieber D, Subramanian N, Stöhr D, Laib Sampaio K, Sinzger C. 2017. A derivative of platelet-derived growth factor receptor alpha binds to the trimer of human cytomegalovirus and inhibits entry into fibroblasts and endothelial cells. PLoS Pathog 13:e1006273. doi: 10.1371/journal.ppat.1006273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Wu Y, Prager A, Boos S, Resch M, Brizic I, Mach M, Wildner S, Scrivano L, Adler B. 2017. Human cytomegalovirus glycoprotein complex gH/gL/gO uses PDGFR-α as a key for entry. PLoS Pathog 13:e1006281. doi: 10.1371/journal.ppat.1006281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Nogalski MT, Chan GC, Stevenson EV, Collins-McMillen DK, Yurochko AD. 2013. The HCMV gH/gL/UL128-131 complex triggers the specific cellular activation required for efficient viral internalization into target monocytes. PLoS Pathog 9:e1003463. doi: 10.1371/journal.ppat.1003463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Zhou M, Yu Q, Wechsler A, Ryckman BJ. 2013. Comparative analysis of gO isoforms reveals that strains of human cytomegalovirus differ in the ratio of gH/gL/gO and gH/gL/UL128-131 in the virion envelope. J Virol 87:9680–9690. doi: 10.1128/JVI.01167-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Rasmussen L, Geissler A, Cowan C, Chase A, Winters M. 2002. The genes encoding the gCIII complex of human cytomegalovirus exist in highly diverse combinations in clinical isolates. J Virol 76:10841–10848. doi: 10.1128/JVI.76.21.10841-10848.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Zhang L, Zhou M, Stanton R, Kamil J, Ryckman BJ. 2018. Expression levels of glycoprotein O (gO) vary between strains of human cytomegalovirus, influencing the assembly of gH/gL complexes and virion infectivity. bioRxiv doi: 10.1101/299222. [DOI] [PMC free article] [PubMed]
- 33.Li G, Nguyen CC, Ryckman BJ, Britt WJ, Kamil JP. 2015. A viral regulator of glycoprotein complexes contributes to human cytomegalovirus cell tropism. Proc Natl Acad Sci U S A 112:4471–4476. doi: 10.1073/pnas.1419875112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Stanton RJ, Baluchova K, Dargan DJ, Cunningham C, Sheehy O, Seirafian S, McSharry BP, Neale ML, Davies JA, Tomasec P, Davison AJ, Wilkinson GW. 2010. Reconstruction of the complete human cytomegalovirus genome in a BAC reveals RL13 to be a potent inhibitor of replication. J Clin Invest 120:3191–3208. doi: 10.1172/JCI42955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wiertz EJ, Jones TR, Sun L, Bogyo M, Geuze HJ, Ploegh HL. 1996. The human cytomegalovirus US11 gene product dislocates MHC class I heavy chains from the endoplasmic reticulum to the cytosol. Cell 84:769–779. doi: 10.1016/S0092-8674(00)81054-5. [DOI] [PubMed] [Google Scholar]
- 36.Renzette N, Bhattacharjee B, Jensen JD, Gibson L, Kowalik TF. 2011. Extensive genome-wide variability of human cytomegalovirus in congenitally infected infants. PLoS Pathog 7:e1001344. doi: 10.1371/journal.ppat.1001344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Renzette N, Gibson L, Bhattacharjee B, Fisher D, Schleiss MR, Jensen JD, Kowalik TF. 2013. Rapid intrahost evolution of human cytomegalovirus is shaped by demography and positive selection. PLoS Genet 9:e1003735. doi: 10.1371/journal.pgen.1003735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sijmons S, Thys K, Mbong Ngwese M, Van Damme E, Dvorak J, Van Loock M, Li G, Tachezy R, Busson L, Aerssens J, Van Ranst M, Maes P. 2015. High-throughput analysis of human cytomegalovirus genome diversity highlights the widespread occurrence of gene-disrupting mutations and pervasive recombination. J Virol 89:7673–7695. doi: 10.1128/JVI.00578-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Dargan DJ, Douglas E, Cunningham C, Jamieson F, Stanton RJ, Baluchova K, McSharry BP, Tomasec P, Emery VC, Percivalle E, Sarasini A, Gerna G, Wilkinson GW, Davison AJ. 2010. Sequential mutations associated with adaptation of human cytomegalovirus to growth in cell culture. J Gen Virol 91:1535–1546. doi: 10.1099/vir.0.018994-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Murrell I, Wilkie GS, Davison AJ, Statkute E, Fielding CA, Tomasec P, Wilkinson GW, Stanton RJ. 2016. Genetic stability of bacterial artificial chromosome-derived human cytomegalovirus during culture in vitro. J Virol 90:3929–3943. doi: 10.1128/JVI.02858-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Murrell I, Tomasec P, Wilkie GS, Dargan DJ, Davison AJ, Stanton RJ. 2013. Impact of sequence variation in the UL128 locus on production of human cytomegalovirus in fibroblast and epithelial cells. J Virol 87:10489–10500. doi: 10.1128/JVI.01546-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Balázs Z, Tombácz D, Szűcs A, Csabai Z, Megyeri K, Petrov AN, Snyder M, Boldogkői Z. 2017. Long-read sequencing of human cytomegalovirus transcriptome reveals RNA isoforms carrying distinct coding potentials. Sci Rep 7:15989. doi: 10.1038/s41598-017-16262-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Gatherer D, Seirafian S, Cunningham C, Holton M, Dargan DJ, Baluchova K, Hector RD, Galbraith J, Herzyk P, Wilkinson GW, Davison AJ. 2011. High-resolution human cytomegalovirus transcriptome. Proc Natl Acad Sci U S A 108:19755–19760. doi: 10.1073/pnas.1115861108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kalser J, Adler B, Mach M, Kropff B, Puchhammer-Stöckl E, Görzer I. 2017. Differences in growth properties among two human cytomegalovirus glycoprotein O genotypes. Front Microbiol 8:1609. doi: 10.3389/fmicb.2017.01609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Murphy E, Yu D, Grimwood J, Schmutz J, Dickson M, Jarvis MA, Hahn G, Nelson JA, Myers RM, Shenk TE. 2003. Coding potential of laboratory and clinical strains of human cytomegalovirus. Proc Natl Acad Sci U S A 100:14976–14981. doi: 10.1073/pnas.2136652100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ryckman BJ, Rainish BL, Chase MC, Borton JA, Nelson JA, Jarvis MA, Johnson DC. 2008. Characterization of the human cytomegalovirus gH/gL/UL128-131 complex that mediates entry into epithelial and endothelial cells. J Virol 82:60–70. doi: 10.1128/JVI.01910-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wang D, Shenk T. 2005. Human cytomegalovirus virion protein complex required for epithelial and endothelial cell tropism. Proc Natl Acad Sci U S A 102:18153–18158. doi: 10.1073/pnas.0509201102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Schoppel K, Hassfurther E, Britt W, Ohlin M, Borrebaeck CA, Mach M. 1996. Antibodies specific for the antigenic domain 1 of glycoprotein B (gpUL55) of human cytomegalovirus bind to different substructures. Virology 216:133–145. doi: 10.1006/viro.1996.0040. [DOI] [PubMed] [Google Scholar]
- 49.Chee M, Rudolph SA, Plachter B, Barrell B, Jahn G. 1989. Identification of the major capsid protein gene of human cytomegalovirus. J Virol 63:1345–1353. [DOI] [PMC free article] [PubMed] [Google Scholar]