

Abstract
During interracial encounters, well-intentioned European Americans sometimes engage in subtle displays of anxiety, which can be interpreted as signs of racial bias by African American partners. In the present research, same-race and cross-race stranger dyads (N = 123) engaged in getting-acquainted tasks, during which measures of sympathetic nervous system responses (preejection period, PEP) and heart rate variability were continuously collected. PEP scores showed that African American partners had stronger physiological linkage to European American partners who evidenced greater anxiety—greater cortisol reactivity, behavioral tension, and self-reported discomfort—which suggests greater physiological responsiveness to momentary changes in partners’ affective states when those partners were anxious. European Americans showed physiological linkage to African American and European American partners, but linkage did not vary as a function of their partner’s anxiety. Using physiological linkage offers a novel approach to understanding how affective responses unfold during dynamic intergroup interactions.
Keywords: interracial anxiety, physiological linkage, dyadic interaction, open data
In the United States, engaging in overtly prejudiced behaviors toward African Americans is largely condemned (Dovidio & Gaertner, 2004). For European Americans, racial bias often operates at a nonconscious level, resulting in subtle expressions that are difficult to control, including nonverbal displays of anxiety and discomfort and, in some cases, exaggerated (disingenuous) positivity (Mendes & Koslov, 2013; Richeson & Shelton, 2005). In interracial encounters, European Americans’ expressions of bias can be ambiguous in meaning and open to interpretation. For instance, nonverbal anxiety could signal dislike, distrust, or just general anxiety over not wanting to say or do something that could be interpreted as biased.
For African Americans, vigilance during interracial interactions is linked to motivation to detect bias across contexts, including practitioner-patient interactions (Cooper et al., 2012; Penner et al., 2016), teacher-student interactions (Casteel, 1998), and casual interactions (Mendes, Major, McCoy, & Blascovich, 2008). Minorities are adept at detecting racial bias, expressed through tone of voice, speech hesitancies, and physical gestures (Dovidio, Kawakami, & Gaertner, 2002). In contrast, European Americans may be less focused on behavioral signals of bias in their African American partners, arguably because minority-group members are more concerned with being the target of prejudice, whereas majority-group members are concerned with appearing likeable (Richeson & Shelton, 2005). In same-race encounters, behaviors that signal anxiety may not be interpreted as negatively as they are in cross-race ones. People are less sensitive to information suggesting that their same-race partner is anxious, and anxious partner behaviors often have weaker effects on outcomes of the interaction, which suggests that they carry less weight in these encounters (West, Pearson, & Stern, 2014).
The present research tested a novel pathway through which subtle displays of anxiety get “under the skin” for African Americans during interactions with European Americans. Specifically, we examined physiological linkage during a dyadic social interaction as a function of whether the dyad was composed of same-race or different-race individuals and how linkage is moderated by individuals’ and their partners’ anxiety. We propose that for African Americans, being in tune with the anxiety of their European American interaction partners can lead them to be more sensitive to tracking the ebb and flow of the affective intensity of their partners.
Anxiety can manifest in many forms; in this work, we attempted to capture anxiety three ways: using cortisol reactivity, behavioral tension, and self-reports of discomfort. Each measure represented distinct information that the partner could detect. Cortisol increases occur during situations that are effortful, socially evaluative, stressful, and threatening (Knapp, Hall, & Horgan, 2014; Lovallo & Thomas, 2000) and can be detected as anxiety by observers (Gray, Mendes, & Denny-Brown, 2008). Cortisol increases during interactions are associated with feelings of uncertainty about the partner (Loving, Gleason, & Pope, 2009; Priem & Solomon, 2011) or when experiencing negative social evaluation (Dickerson & Kemeny, 2004). Displays of behavioral tension—clenching, stiffness, and immobility—communicate discomfort and, particularly within interracial encounters, can convey racial bias (Dovidio et al., 2002). Lastly, self-reported discomfort requires individuals to be willing to report their felt state. This full suite of information provides potentially nonoverlapping information regarding a partner’s level of anxiety, especially since behavioral, physiological, and self-reported anxiety do not always correlate (Blascovich, Mendes, & Seery, 2002). For ease of clarity, we refer to all of these variables as “partner anxiety” throughout, although we considered each channel of anxiety separately.
As a secondary hypothesis, we tested whether African Americans who have stronger expectations of being treated negatively because of their race would also be more strongly linked to their European American partners. Minorities who expect discrimination during cross-race contact are attuned to information that could convey discrimination (Mendoza-Denton, Downey, Purdie, Davis, & Pietrzak, 2002), and greater expectations of discrimination are associated with greater discomfort, less trust, and higher cortisol reactivity during first-time cross-race encounters (Mendoza-Denton et al., 2002; Page-Gould, Mendoza-Denton, & Tropp, 2008). We tested this hypothesis using race-based rejection expectations of African Americans, which were measured prior to the study.
To test our hypotheses, we examined physiological linkage between partners who engaged in a cross-race (European American–African American) or same-race (European American–European American) interaction. Linkage was operationalized as the effect of partners’ physiological response (preejection period, PEP) at a given minute on their partners’ PEP response in the subsequent minute. We theorized that for African Americans, physiological linkage of PEP responses would be stronger when their European American partners had greater cortisol reactivity, appeared tenser, and reported more discomfort and when the African American participants had higher race-based rejection expectations.
Physiological linkage provides an optimal method for testing sensitivity to ebbs and flows in a partner’s affective states, given that being physiologically linked to a partner, and specifically showing conditional physiological reactivity, is related to social sensitivity (Guastello, Pincus, & Gunderson, 2006; Hess & Blairy, 2001) and accuracy in detecting emotions (Guastello et al., 2006; Levenson & Ruef, 1997), which require attentiveness toward the partner (Schoebi, 2008). Thus, physiological linkage provides an optimal methodological approach to testing the hypothesis that African Americans are more tuned in to European Americans who are anxious than to European Americans who are not.
Overview of the Present Study
In this study, newly acquainted individuals engaged in either an interracial (African American–European American) or intraracial (European American–European American) interaction. Dyads completed a series of cooperative tasks designed to vary in affective intensity and cognitive demand, which thereby allowed us to test the extent to which participants tracked the ups and downs of the affective state of their partner. Because the tasks encouraged attention toward the partner, we expected physiological linkage for all participants. However, we hypothesized that for cross-race dyads, physiological linkage would increase for African Americans as a function of their European American partner’s anxiety (and their own race-based rejection expectations), but this would not be the case for European Americans. Lastly, we examined whether participants in cross-race dyads would show larger decreases in respiratory sinus arrhythmia (RSA; a measure of heart rate variability) than those in same-race dyads, which would provide converging evidence that participants in cross-race interactions exert more mental effort during interracial interactions, possibly because of increased attunement.
Method1
Design and participants
Participants were recruited through a combination of ads, snowball sampling, and outreach through community LISTSERVs; 258 participants (129 same-sex dyads) were initially recruited for participation in the study. Four dyads did not complete the study: In two dyads, 1 participant chose to leave midway through the study; another two dyads revealed that they knew each other when they were introduced. Two additional dyads with a partner age difference exceeding the a priori cutoff (maximum of 3 years) were removed from analyses. All other missing data were due to technical difficulties with either physiological collection or experimenter error.
Of the remaining 123 dyads, 42 were same-race (White-White; 48% male, 52% female), and 81 were mixed-race (White-Black; 49% male, 51% female). The mean age of the participants was 23.44 years (SD = 4.30), and their ages ranged from 18 to 40. All participants had lived in the United States since at least age 7 and considered English their primary language (see the Supplemental Material available online for more sample details).
We planned on running a minimum of 100 dyads with a secondary goal of running an additional 20% to account for missing data. For laboratory data in which physiological responses were obtained, we typically observed approximately 10% data missing completely at random (and given that we ran dyads, we doubled this value).
Procedure
Prior to arrival (at least 48 hr in advance), participants completed an online screening questionnaire that included the race-based rejection-sensitivity scale (and the anxiety subscale; details are given below). Participants were scheduled during afternoon hours to control for diurnal changes in cortisol, and on arrival, participants were placed in separate experimental rooms; they were not informed during recruitment or when providing initial informed consent that this was a two-person study. Participants provided a baseline saliva sample that was assayed for cortisol, and an experimenter attached physiological sensors to the participant. Participants relaxed alone in a seated position for a 5-min baseline recording.2 After baseline measurements, participants were told that for the rest of the study they would be interacting with another participant, and they provided a second consent at this time to continue. Participants then moved to a large room where they were introduced and asked whether they knew each other. For participants who knew each other previously, the study ended then. If the participants never met, the study continued.
We designed the study so that the different tasks the dyad completed would have distinct emotional and cognitive demands that would manifest varied physiological reactions. We also designed the protocol so that the study was pleasant, cooperative, and engaging over the 90-min period that the dyad interacted. The first task consisted of a 6-min “getting-to-know-you” conversation with the aid of a list of questions. After this, participants completed a first-impressions questionnaire to indicate their level of initial discomfort with their partner. Next, participants completed a task we designed that used tactile finger spelling. For 4 min, participants alternated spelling out words using American Sign Language (ASL; an ASL sheet was provided); but rather than seeing each other’s hands, they placed their hands in a box and felt each other’s hands to determine the words being spelled (see Koslov, 2010; Stern & West, 2014). The last task was a cooperative word-guessing game based on the game Taboo. In this task, participants took 2-min turns trying to get their partner to guess words, without being able to use any of five taboo words that were listed on their prompt cards (e.g., if the word to be guessed was “birthday,” the clue giver could not say “happy,” “anniversary,” “candles,” “cake,” or “presents”). The participants received 25 cents for every word guessed correctly and lost 25 cents for each taboo word accidentally spoken. The game lasted for 8 min, which gave each participant two turns as a guesser and two as a prompter. Random assignment determined who guessed first.
After the last task, a curtain was drawn between the participants, who were told that their interaction was over. Participants were given a second-impressions questionnaire in which self-reported discomfort again was measured. After a recovery recording of 3 min, participants returned to their original experiment room, and the second saliva sample was collected. Both participants were debriefed separately by their experimenters and then compensated and escorted out of the laboratory.
Measures
Autonomic nervous system (ANS) responses
Participants’ ANS responses were obtained during resting baseline and during all tasks with the partner. We employed electrocardiography (ECG) and impedance cardiography to obtain measurements of sympathetic nervous system (SNS) activity, specifically PEP, which is the amount of time between the left ventricle contracting (ECG waveform) and the aortic valve opening (Δz/Δt waveform). PEP is a useful measure for examining physiological influence because it is measured unobtrusively and continuously, fluctuations can be linked to momentary changes in affective states, and it is responsive to changes in affective states in a short time frame (within 3 to 5 s; Kraus & Mendes, 2013; Mendes, 2016). It represents a pure measure of SNS activation because it is calculated during the heart cycle when there are only sympathetic influences (during systole). This measure has been used to examine physiological influence in prior dyadic-interaction studies (Kraus & Mendes, 2013; Waters, West, Karnilowicz, & Mendes, 2017; Waters, West, & Mendes, 2014).
We also estimated heart rate variability, specifically RSA, from the ECG waveform. RSA provides a measure of parasympathetic nervous system activation and is calculated as the variability in heart rate during respiration. To process PEP and RSA, we first visually inspected the waveform data, edited artifacts, and then used MindWare software (Impedance Cardiography 2.6 and Heart Rate Variability 2.6; Gahanna, OH) to calculate PEP and RSA following standard guidelines from the Society for Psychophysiological Research (Berntson et al., 1997; Sherwood et al., 1990).
Anxiety
We measured anxiety in three ways: using (a) cortisol reactivity to compare levels from before and after the interaction; (b) observer-rated nonverbal behaviors of tension during the social interaction; and (c) self-reported discomfort. Given that none of these measures shared method variance, we expected little to no overlap between them. Cortisol is released in a pulsatile fashion characterized by dumps, then breaks (Lovallo & Thomas, 2000). Self- and observer-rated behaviors were measured at discrete time points during the interaction. In support of this expectation, there was little to no correlation across the measures (rs between −.05 and −.13; see the Supplemental Material).
Saliva samples were collected using passive drool into IBL tubes (Tecan, Morrisville, NC), and samples were stored at −80° C. Once the study was completed, all saliva samples were shipped on dry ice to Clemens Kirschbaum’s laboratory in Dresden, Germany, to be assayed for cortisol using commercial immunoassay kits. Interassay coefficients were less than 10%. Cortisol reactivity was calculated as the second cortisol assessment (postinteraction) minus the first cortisol assessment (baseline).
To assess behavioral displays of tension, two trained coders unaware of the partners’ race rated the nonverbal behaviors of participants by watching videos that displayed only the participant during the getting-acquainted conversation portion of the study. All intraclass correlation coefficients (ICCs) between coders were statistically reliable (p < .001), which indicates acceptable agreement. We followed the guidelines of Heyman, Lorber, Eddy, and West (2014) to train coders. Coders rated three behaviors that were averaged to create a composite of nonverbal behavioral discomfort (measured from 0, not at all, to 5, mostly). These behaviors were how tense the participant appeared (erect position and clenched limbs; M = 1.17, SD = 1.53, ICC = .27), how rigid the participant appeared (stiffness and lack of movement; M = 1.39, SD = 1.42, ICC = .22), and how much the participant leaned forward (with an open posture; M = 2.22, SD = 1.15, ICC = .34; reverse-coded such that higher scores indicate less leaning forward; α = .82).
To measure discomfort, we asked participants to respond to the item “How comfortable do you feel with your partner?” (on a scale from 1 to 7) at two time points in the interaction: after the first getting-acquainted segment and at the end of the study. These two items were reverse-coded and averaged to create a composite of discomfort (r = .56, M = 2.52, SD = 0.90; effects were similar when each item was used individually though stronger after the entire interaction; see the Supplemental Material).
Race-based rejection sensitivity
African American participants read six scenarios that Mendoza-Denton and colleagues (2002) pretested to elicit race-based rejection concerns in African Americans (given the heterogeneity of our sample, we removed an additional six scenarios from the original scale that were specific to college settings). Participants first indicated their concern that each outcome would occur because of their race on a scale from 1 (very unconcerned) to 6 (very concerned) that measured race-based rejection anxiety (α = .89). Second, they indicated the likelihood that the other person would reject them because of their race on a scale from 1 (very unlikely) to 6 (very likely) that measures race-based rejection expectations (α = .89). We included race-based anxiety in our analyses to rule out the possibility that any negative reactions to potential discrimination—anxiety or expectation—were driving our effects. Rather, it is the component of the scale that is mostly strongly tied to vigilance in interracial encounters—race-based rejection expectations—that potentiates physiological linkage. The two constructs were highly correlated (r = .72), which is not surprising given that participants were responding to the same scenarios for both sets of items.
Analysis strategy
To examine the extent to which individuals showed physiological linkage with their partners from one time point to the next, we estimated a stability and influence model (Kashy & Kenny, 2000), in which participants’ PEP score at one point was treated as a function of their own score at the prior time point (the stability path) and their partner’s score at that prior time point (the influence path, referred to as linkage from here on). We note that the structuring of the data (as a person period pairwise file; see West, 2013) allowed us to estimate partner effects for both interaction partners in the model.
The stability and linkage paths were moderated by two contrast variables. Dyad race compared individuals in cross-race dyads (African Americans with European American partners and European Americans with African American partners; both coded −2/3) with those in same-race dyads (European Americans with European American partners; coded 1/3). Person race compared European Americans with African Americans within cross-race dyads (European Americans with European American partners were coded 0, African Americans with European American partners were coded −1/2, and European Americans with African American partners were coded 1/2). The main effects of these contrasts were also included.
The model allowed us to examine how much individuals showed physiological linkage to their partners while adjusting for stability in physiological states, thereby providing a conservative estimate of linkage. Because the data contained indistinguishable dyads measured over time (and at the same time points for both dyad members), we specified a two-level crossed model using multilevel modeling (see the Supplemental Material for SAS PROC MIXED syntax). The degrees of freedom were estimated using the Satterwaite method and could be fractional (Kenny, Kashy, & Cook, 2006). For each model, we included body mass index as a covariate (given its direct effect on cardiac functioning; see Waters et al., 2014, for the same approach).
For the sake of parsimony, we ran separate models for each of the three measures of anxiety. Following the recommendations of Kenny and Ledermann (2010), we fully saturated the models such that they contained both actor and partners’ anxiety as moderators of both stability and physiological linkage; this strategy allowed us to isolate the effects of partners’ anxiety while adjusting for any empirical overlap between participants’ and partners’ anxiety and to look at linkage while adjusting for stability. Of key theoretical interest was the three-way interaction among partners’ prior PEP (time x − 1) score, person race, and partners’ anxiety. This interaction tested whether the strength of the linkage path varied between European Americans and African Americans in cross-race dyads as a function of the partner’s anxiety. Given that there is no single recommended measure of effect size for effect estimates in multilevel models, and transforming variables by standardizing them can alter the random effects of the model (because random effects are not invariant to linear transformations of explanatory variables and can change dramatically when variables are standardized; see Hox, 2002), we present the unstandardized coefficients and confidence intervals (CIs) for the unstandardized effects only.
We present the results for cortisol reactivity first, followed by the effects for behavioral tension, self-reported discomfort, and race-based rejection expectations. For parsimony, we discuss only the main effects of stability and physiological linkage for the cortisol model but note that these effects were observed across all models. We discuss only significant effects for each model that are of theoretical interest, but we report all effects in the Supplemental Material.
Results
Cortisol reactivity
As expected, participants’ PEP scores were stable from a given minute to the next, as indicated by a main effect of the participant’s PEP prior score on his or her current PEP score, b = 0.69, 95% CI = [0.66, 0.71], SE = 0.013, t(112) = 52.88, p < .001. Participants also showed positive linkage to their partners’ physiology, as indicated by a main effect of the partner’s prior PEP score (time x − 1) on the participant’s current PEP score (time x), b = 0.06, 95% CI = [0.04, 0.08], SE = 0.011, t(147) = 5.63, p < .001. These two effects show that a strong predictor of participants’ SNS activation at time x was their SNS activation at time x − 1. Beyond this effect of stability, the more novel finding here is that participants overall showed linkage to their partners’ physiological response. Increases or decreases in partners’ SNS activation predicted participants’ SNS change in the same direction in the subsequent bin.
Moreover, the hypothesized three-way interaction among partners’ prior PEP score, person race, and partners’ cortisol reactivity on participants’ current PEP score was found, b = −0.02, 95% CI = [−0.03, −0.004], SE = 0.006, t(288) = −2.77, p = .006. The effect of partners’ cortisol reactivity on physiological linkage differed between European Americans and African Americans in cross-race dyads. We next examined the effect of partners’ cortisol reactivity on physiological linkage for the three types of dyad models. No other effects for stability were significant, ps > .21 (see the Supplemental Material for all other significant effects). Physiological linkage paths were about one tenth the size of stability paths, as a point of comparison.
Turning to the two-way interaction between partners’ prior PEP score and partners’ cortisol reactivity, we found that the interaction was significant for African Americans with European American partners, b = 0.009, 95% CI = [0.0005, 0.02], SE = 0.004, t(260) = 2.09, p = .040. As seen in Figure 1, the higher the European American partner’s cortisol reactivity, the stronger the physiological linkage from European Americans to their African American partners. In other words, African Americans were more strongly linked to European Americans who had greater cortisol reactivity.
Fig. 1.
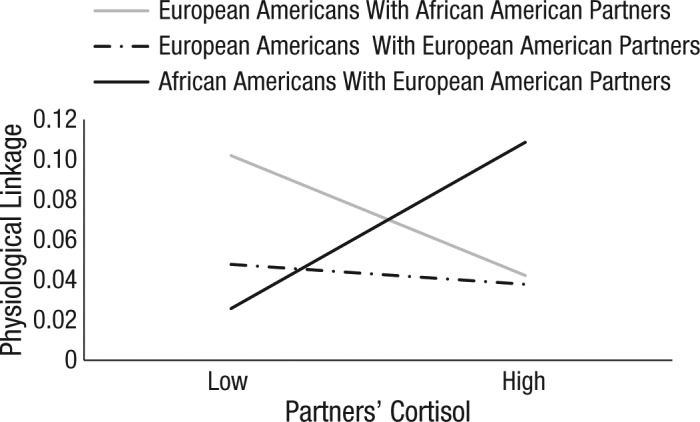
Mean predicted physiological linkage as a function of partners’ cortisol reactivity and dyad type. Physiological linkage was the effect of partners’ preejection period (PEP) score at time x − 1 on participants’ PEP score at time x. High and low refer to values 1 standard deviation above and below the mean, respectively.
For European Americans with African American partners, we found that the two-way interaction between partners’ prior PEP score and partners’ cortisol reactivity was not significant, b = −0.006, 95% CI = [−0.01, 0.003], SE = 0.004, t(288) = −1.39, p = .160. Physiological linkage was significant overall for these individuals, b = 0.07, 95% CI = [0.04, 0.11], SE = 0.02, t(234) = 4.27, p < .001, but did not vary as a function of partners’ cortisol reactivity (see Fig. 1).
Finally, examining European Americans with European American partners revealed that the two-way interaction between partners’ prior PEP score and partners’ cortisol reactivity was not significant, b = − 0.001, 95% CI = [−0.01, 0.008], SE = 0.0004, t(144) = −0.23, p = .820. As seen in Figure 1, physiological linkage was significantly different from zero overall, b = 0.04, 95% CI = [0.004, 0.08], SE = 0.02, t(205) = 2.15, p = .030, but did not vary as a function of the partner’s cortisol reactivity.
In sum, all participants showed physiological linkage to their partners, but African Americans showed stronger linkage to European American partners when those partners demonstrated greater cortisol reactivity.
Behavioral tension
Main effects of stability and physiological linkage are presented in the Supplemental Material and are consistent with the effects for partners’ cortisol. Consistent with the effect for cortisol reactivity, the hypothesized three-way interaction among partners’ prior PEP score, person race, and partners’ behavioral tension was significant, b = −0.07, 95% CI = [−0.14, −0.007], SE = 0.03, t(555) = −2.16, p = .030, which indicates that the effect of the partner’s behavior on physiological linkage differed between European Americans and African Americans in cross-race dyads.
Turning to the hypothesized two-way interaction between partners’ prior PEP score and partners’ tension, we found that for African Americans with European American partners, the interaction was significant, b = 0.05, 95% CI = [0.004, 0.09], SE = 0.02, t(306) = 2.18, p = .030. As seen in Figure 2, African Americans whose European American partners showed more behavioral tension (to observers coding the videos unaware of the actor’s race) showed stronger physiological linkage to those partners. However, this two-way interaction was not significant for European Americans with African American partners, b = −0.03, 95% CI = [−0.07, 0.02], SE = 0.02, t(504) = −1.16, p = .250, or for European Americans with European American partners, b = 0.03, 95% CI = [−0.04, 0.11], SE = 0.04, t(511) = 0.71, p = .480.
Fig. 2.
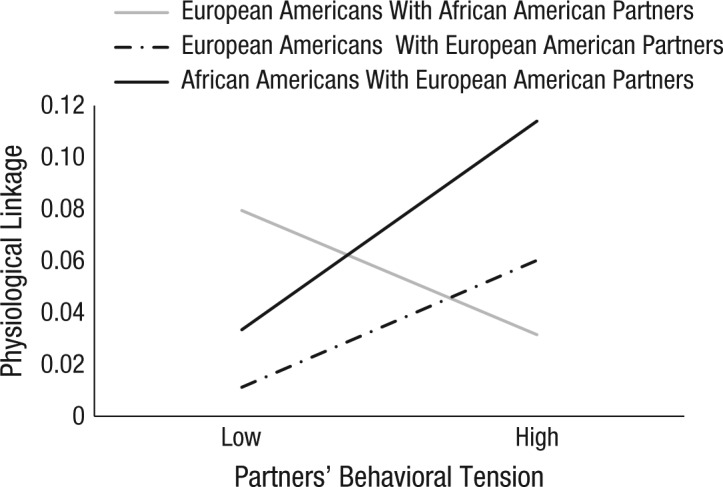
Mean predicted physiological linkage as a function of partners’ behavioral tension and dyad type. Physiological linkage was the effect of partners’ preejection period (PEP) score at time x – 1 on participants’ PEP score at time x. High and low refer to values 1 standard deviation above and below the mean, respectively.
Self-reported discomfort
Main effects of stability and physiological linkage are presented in the Supplemental Material and are consistent with the effects for partners’ cortisol and behavioral tension. Consistent with the effects for cortisol reactivity and behavioral tension, the hypothesized three-way interaction among partners’ prior PEP score, person race, and partners’ discomfort was significant, b = −0.09, 95% CI = [−0.15, −0.03], SE = 0.03, t(776) = −2.82, p = .005. No other effects were significant, ps > .18.
The hypothesized two-way interaction between partners’ prior PEP score and partners’ discomfort was significant for African Americans with European partners, b = 0.06, 95% CI = [0.007, 0.11], SE = 0.02, t(674) = 2.26, p = .020. As seen in Figure 3, African American participants showed stronger linkage to their European American partners the more uncomfortable those partners felt. However, this two-way interaction was not significant for European Americans with African American partners, b = −0.03, 95% CI = [−0.07, 0.006], SE = 0.02, t(509) = −1.65, p = .100, or for European Americans with European American partners, b = 0.02, 95% CI = [−0.02, 0.06], SE = 0.02, t(625) = 0.99, p = .330.
Fig. 3.
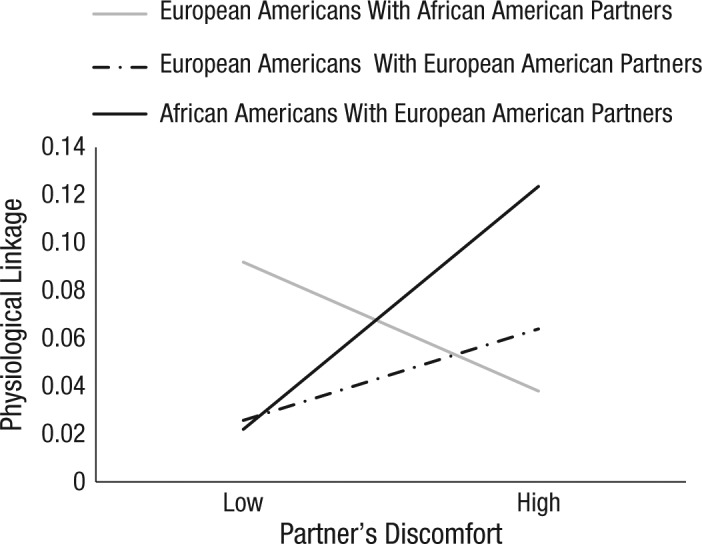
Mean predicted physiological linkage as a function of partners’ self-reported discomfort and dyad type. Physiological linkage was the effect of partners’ preejection period (PEP) score at time x – 1 on participants’ PEP score at time x. High and low refer to values 1 standard deviation above and below the mean, respectively.
African Americans’ race-based rejection expectations and anxiety
For race-based rejection expectations, the three-way interaction among partners’ prior PEP score, person race, and African Americans’ rejection expectations was significant, b = −0.03, 95% CI = [−0.05, −0.004], SE = 0.01, t(377) = −2.30, p = .020. This effect was driven by an opposite pattern of effects for European Americans and African Americans within cross-race encounters. For the interaction between partners’ prior PEP score and partners’ rejection expectation in European Americans, the higher their African American partners’ rejection-expectation score, the less they were influenced by their partner, b = −0.02, 95% CI = [−0.063, 0.008], SE = 0.02, t(1103) = −1.53, p = .120. However, for African Americans, the higher their own rejection-expectation score, the more they were influenced by their European American partners, b = 0.02, 95% CI = [−0.007, 0.06], SE = 0.01, t(336) = 1.59, p = .110. These results indicate that consistent with effects for the three anxiety measures, African Americans who had higher expectations of being treated negatively because of their race evidenced greater physiological linkage to their European American partners. None of the effects of African Americans’ race-based rejection anxiety on physiological linkage approached statistical significance (i.e., effects of their anxiety on their own or their partners’ linkage, ps > .30).
Post hoc analyses
We found that unlike African Americans, European Americans did not show physiological linkage to their partners as a function of their partners’ anxiety. To test the possibility that European American might be less attentive during the interaction than African Americans, we examined heart-rate-variability reactivity (indexed by RSA) during the most cognitively demanding portion of the interaction: a cooperative word game during which partners worked together and attended to each other’s behaviors to correctly guess a word from verbal cues. Although RSA reactivity can indicate multiple psychological processes (stress, emotion, motivation, and cognition; Mendes, 2016), during cognitively demanding tasks, decreases in RSA can be interpreted as a measure of attentional effort (Hansen, Johnson, & Thayer, 2003; Tattersall & Hockey, 1995). We reasoned that both European Americans and African Americans in cross-race interactions would demonstrate greater decreases in RSA reactivity (i.e., withdrawal of the vagal brake) than European Americans in same-race interactions, because they would be particularly motivated to tune in to their different-race partners, even if the motivations were different (Bergsieker, Shelton, & Richeson, 2010).
We estimated a two-level crossed model using generalized estimating equations. In the model, we included the main effect of person race (a three-level variable that compared European Americans with European American partners, African Americans with European American partners, and European Americans with African American partners), time, and their interactions. Results indicated a main effect of person race, Wald χ2(2) = 8.86, p = .012, and time, Wald χ2(3) = 20.46, p < .001. The Time × Person Race interaction was not significant, p > .250. Pairwise comparisons indicated that European Americans in same-race interactions showed less RSA withdrawal (M = −.26, SE = .06) than European Americans in cross-race interactions (M = −.46, SE = .06, p = .021) and African Americans in cross-race interactions (M = −.60, SE = .06, p = .004). European Americans with African American partners were not significantly different from African Americans with European American partners (p = .550). These data provide suggestive, albeit provisional, data that cross-race interactions might be associated with greater attentional effort or vigilance, compared with same-race interactions.
Discussion
Recently acquainted same-sex dyads showed physiological linkage during cooperative social tasks. Importantly, anxiety of the partners influenced linkage but only in cross-race dyads, and the relationship was asymmetric—European Americans’ greater anxiety facilitated linkage of their African American partners, but African Americans’ anxiety did not facilitate linkage of their European American partners (with data trending in the opposite direction). Specifically, African Americans showed stronger physiological linkage to their European American partners when those partners had higher cortisol reactivity, greater behavioral displays of tension, and higher self-ratings of interpersonal discomfort and when African Americans had higher race-based rejection expectations—a construct related to vigilance in detecting racial bias in interracial interactions. The findings for rejection expectations suggest that expecting to be treated negatively because of one’s race in general can predict an automatic process related to attunement (physiological linkage) in an actual encounter—a finding that to our knowledge has not been previously demonstrated. Moreover, rarely in studies of dyads do scholars observe parallel effects of partner-level variables measured during an encounter, and respondent-level variables measured several weeks prior to an encounter, on a single dyadic process. Future work should examine how partner- and respondent-level factors might work in congruence to produce physiological linkage.
In ancillary analyses, we observed that European Americans and African Americans showed greater heart-rate-variability reactivity (decreases in RSA) during the cooperative task than did in-group (European American–European American) dyads, which suggests greater attentional effort in intergroup than in same-race dyads. It might be that that European Americans expended cognitive effort and attention during the cross-race interaction, but for them, it was not critical to attend to information in partners that signals anxiety, or they misread the behavioral signals (Gray et al., 2008). Alternatively, RSA decreases might indicate greater negative affect or a combination of negative affect and attentional effort. While we were unable to differentiate the precise psychological processes of RSA decreases in this context, these data provide future research directions to examine within the intergroup context. Taken together, the RSA data, coupled with the generally weaker findings for physiological linkage in same-race dyads, provide evidence that individuals in same-race encounters may be less attuned to (or experiencing less negative affect with) their partners than those in cross-race ones.
These findings add to the literature on how subtle expressions of anxiety affect African Americans during interracial interactions. Physiological linkage to a partner is not a universally positive or negative experience for dyad members; it reflects a sensitivity to the fluxes and flows in the intensity of one’s partner’s affective states. Therefore, it is important to understand the affective state that one’s partner is experiencing to understand the possible outcomes of linkage. In interracial interactions in which European Americans are experiencing negative affect, such as stress and threat, linkage for African Americans might reflect the tendency to “catch” stress from their partners. If African Americans are chronically engaging with stressed partners and are particularly attuned to cues of stress, they may in turn experience elevated levels of stress as a result, which over time could accumulate to dysregulation.
This work has limitations that are important to highlight. First, we did not include same-race dyads with two African Americans because of a lack of feasibility—less than 5% of the population where this sample was drawn was African American. Given that we assigned participants to same-sex partners of similar age and occupation who did not know each other, it was not possible to recruit African American dyads. Therefore, we are limited in our ability to conclude that it is only during interracial interactions that African Americans are attuned to anxious behavior of partners. Despite this limitation, Whites and minorities within same-race encounters are often affected in similar ways by interacting with a partner who displays anxiety or is perceived to be anxious (e.g., Richeson, Trawalter, & Shelton, 2005; West et al., 2014) and that for African Americans, vigilance for detecting cues of bias are unique to the cross-race domain (Mendes et al., 2008; Richeson & Shelton, 2007).
Additionally, although we captured anxiety using multiple channels, the full suite of anxious cues that are detected by African Americans is unknown. The behavioral cues we measured—rigid body, tension, and failure to lean in—provide a glimpse into the possible observable signals; however, it is likely that there are multiple sensorial channels in which anxiety is manifested, including voice frequency and pitch, facial expressions, odor, and tactile information. In this study, participants had to touch each other’s hands during the tactile finger-spelling task, and in a recent study, we found that touch was an especially powerful predictor of physiological synchrony among close others (Waters et al., 2017). One possibility is that some channels of information are particularly potent cues of the partner’s anxiety during certain times of the interaction—the touch task, for example, conveys different information than the face-to-face getting-acquainted encounter.
Lastly, we theorized that vigilance to subtle indicators of bias underlies the process through which linkage is potentiated for African Americans, but we never measured vigilance directly. Although it might be difficult to directly measure attention to all cues in the partner that convey anxiety, future work could systematically vary the types of information conveyed by the partner and the access to this information to gain a more complete understanding of the role of vigilance in this process.
In summary, the present research identifies a novel physiological pathway through which interracial anxiety is transmitted during social interactions. Interracial anxiety can affect the lives of minorities to the extent that it influences voting behavior and attitudes associated with affordable housing, affirmative action, and employment opportunities, but one typically thinks of interracial anxiety as residing in the mind, body, and actions of the person who holds the beliefs. Here, we show that interracial anxiety does not simply affect the holder of bias, but also can directly affect the individual who is the target of the anxiety.
Supplementary Material
A more detailed description of the method is provided in the Supplemental Material available online. Data and syntax are available at http://www.psych.nyu.edu/westlab/lab-resources.html and https://osf.io/f78gs/.
Participants were randomly assigned to a task with a high cognitive load (counting backwards from a four-digit number by steps of seven) or a low cognitive load (counting backwards from a four-digit number by steps of one) prior to meeting each other. We ran the analyses presented here with and without controlling for this covariate, and there were no appreciable changes in any of the results.
Footnotes
Action Editor: D. Stephen Lindsay served as action editor for this article.
Declaration of Conflicting Interests: The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding: This research was supported by grants from the National Science Foundation (BCS 1430799) and National Institute on Aging (R24AG048024).
Supplemental Material: Additional supporting information can be found at http://journals.sagepub.com/doi/suppl/10.1177/0956797617722551
Open Practices:

All data have been made publicly available via the Open Science Framework and can be accessed at https://osf.io/kq897. The complete Open Practices Disclosure for this article can be found at http://journals.sagepub.com/doi/suppl/10.1177/0956797617722551. This article has received the badge for Open Data. More information about the Open Practices badges can be found at http://www.psychologicalscience.org/publications/badges.
References
- Bergsieker H. B., Shelton J. N., Richeson J. A. (2010). To be liked versus respected: Divergent goals in interracial interactions. Journal of Personality and Social Psychology, 99, 248–264. doi: 10.1037/a0018474 [DOI] [PubMed] [Google Scholar]
- Berntson G. G., Bigger J. T., Jr., Eckberg D. L., Grossman P., Kaufmann P. G., Malik M., , . . . van der Molen M. W. (1997). Heart rate variability: Origins, methods, and interpretive caveats. Psychophysiology, 34, 623–648. [DOI] [PubMed] [Google Scholar]
- Blascovich J., Mendes W. B., Seery M. D. (2002). Intergroup encounters and threat: A multi-method approach. In Mackie D. M., Smith E. R. (Eds.), From prejudice to intergroup emotions: Differentiated reactions to social groups (pp. 89–109). New York, NY: Psychology Press. [Google Scholar]
- Casteel C. A. (1998). Teacher-student interactions and race in integrated classrooms. The Journal of Educational Research, 92, 115–120. doi: 10.1080/00220679809597583 [DOI] [Google Scholar]
- Cooper L. A., Roter D. L., Carson K. A., Beach M. C., Sabin J. A., Greenwald A. G., Inui T. S. (2012). The associations of clinicians’ implicit attitudes about race with medical visit communication and patient ratings of interpersonal care. American Journal of Public Health, 102, 979–987. doi: 10.2105/AJPH.2011.300558 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dickerson S. S., Kemeny M. E. (2004). Acute stressors and cortisol responses: A theoretical integration and synthesis of laboratory research. Psychological Bulletin, 130, 355–391. doi:10.1037.0033-2909.130.3.355 [DOI] [PubMed] [Google Scholar]
- Dovidio J. F., Gaertner S. L. (2004). Aversive racism. In Zanna M. P. (Ed.), Advances in experimental social psychology (Vol. 36, pp. 1–52). San Diego, CA: Academic Press. doi: 10.1016/S0065-2601(04)36001-6 [DOI] [Google Scholar]
- Dovidio J. F., Kawakami K., Gaertner S. L. (2002). Implicit and explicit prejudice and interracial interaction. Journal of Personality and Social Psychology, 82, 62–68. doi: 10.1037/0022-3514.82.1.62 [DOI] [PubMed] [Google Scholar]
- Gray H., Mendes W. B., Denny-Brown C. (2008). An in-group advantage in detecting intergroup anxiety. Psychological Science, 19, 1233–1237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guastello S. J., Pincus D., Gunderson P. R. (2006). Electrodermal arousal between participants in a conversation: Nonlinear dynamics and linkage effects. Nonlinear Dynamics, Psychology, and Life Sciences, 10, 365–399. [PubMed] [Google Scholar]
- Hansen A. L., Johnson B. H., Thayer J. F. (2003). Vagal influence on working memory and attention. International Journal of Psychophysiology, 48, 263–274. doi: 10.1016/S0167-8760(03)00073-4 [DOI] [PubMed] [Google Scholar]
- Hess U., Blairy S. (2001). Facial mimicry and emotional contagion to dynamic emotional facial expressions and their influence on decoding accuracy. International Journal of Psychophysiology, 40, 129–141. doi: 10.1016/S0167-8760(00)00161-6 [DOI] [PubMed] [Google Scholar]
- Heyman R. E., Lorber M. F., Eddy J. M., West T. V. (2014). Behavioral observation and coding. In Reis H. T., Judd C. M. (Eds.), Handbook of research methods in social and personality psychology (2nd ed., pp. 345–372). New York, NY: Cambridge University Press. [Google Scholar]
- Hox J. (2002). Multilevel analysis: Techniques and applications. Mahwah, NJ: Erlbaum. [Google Scholar]
- Kashy D. A., Kenny D. A. (2000). The analysis of data from dyads and groups. In Reis H. T., Judd C. M. (Eds.), Handbook of research methods in social and personality psychology (1st ed., pp. 451–477). New York, NY: Cambridge University Press. [Google Scholar]
- Kenny D. A., Kashy D. A., Cook W. L. (2006). Dyadic data analysis. New York, NY: Guilford Press. [Google Scholar]
- Kenny D. A., Ledermann T. (2010). Detecting, measuring, and testing dyadic patterns in the actor–partner interdependence model. Journal of Family Psychology, 24, 359–366. doi: 10.1037/a0019651 [DOI] [PubMed] [Google Scholar]
- Knapp M. L., Hall J. A., Horgan T. G. (2014). Nonverbal communication in human interaction. Boston, MA: Wadsworth Cengage Learning. [Google Scholar]
- Koslov K. R. (2010). Pounding hearts and brittle smiles: The effect of resource depletion on outgroup positivity (Doctoral dissertation). Available from ProQuest Dissertations and Theses Database. (UMI No. 3414826) [Google Scholar]
- Kraus M. W., Mendes W. B. (2013). Sartorial symbols of social class elicit class-consistent behavioral and physiological responses: A dyadic approach. Journal of Experimental Psychology: General, 143, 2330–2340. doi: 10.1037/xge0000023 [DOI] [PubMed] [Google Scholar]
- Levenson R. W., Ruef A. M. (1997). Physiological aspects of emotional knowledge and rapport. In Iches W. J. (Ed.), Empathic accuracy (pp. 44–72). New York, NY: Guilford Press. [Google Scholar]
- Lovallo W. R., Thomas T. L. (2000). Stress hormones in psychophysiological research. In Cacioppo J. T., Tassinary L. G., Berntson G. G. (Eds.), Handbook of psychophysiology (pp. 342–367). Cambridge, England: Cambridge University Press. [Google Scholar]
- Loving T. J., Gleason M. E. J., Pope M. T. (2009). Transition novelty moderates daters’ cortisol responses when talking about marriage. Personal Relationships, 16, 187–203. doi: 10.1111/j.1475-6811.2009.01218.x [DOI] [Google Scholar]
- Mendes W. B. (2016). Emotion and the autonomic nervous system. In Barrett L. F., Lewis M., Haviland-Jones J. (Eds.), Handbook of emotions (pp. 166–181). New York, NY: Guilford Press. [Google Scholar]
- Mendes W. B., Koslov K. (2013). Brittle smiles: Positive biases toward stigmatized and outgroup targets. Journal of Experimental Psychology: General, 142, 923–933. doi: 10.1037/a0029663 [DOI] [PubMed] [Google Scholar]
- Mendes W. B., Major B., McCoy S., Blascovich J. (2008). How attributional ambiguity shapes physiological and emotional responses to social rejection and acceptance. Journal of Personality and Social Psychology, 94, 278–291. doi: 10.1037/0022-3514.94.2.278 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mendoza-Denton R., Downey G., Purdie V. J., Davis A., Pietrzak J. (2002). Sensitivity to status-based rejection. Journal of Personality and Social Psychology, 83, 896–918. doi: 10.1037/0022-3514.83.4.896 [DOI] [PubMed] [Google Scholar]
- Page-Gould E., Mendoza-Denton R., Tropp L. R. (2008). With a little help from my cross-group friend. Journal of Personality and Social Psychology, 95, 1080–1094. doi: 10.1037/0022-3514.95.5.1080 [DOI] [PubMed] [Google Scholar]
- Penner L. A., Dovidio J. F., Gonzalez R., Albrecht T. L., Chapman R., Foster T., , . . . Eggly S. (2016). The effects of oncologist implicit racial bias in racially discordant oncology interactions. Journal of Clinical Oncology, 34, 2874–2881. doi: 10.1200/JCO.2015.66.3658 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Priem J. S., Solomon D. H. (2011). Relational uncertainty and cortisol responses to hurtful and supportive messages from a dating partner. Personal Relationships, 18, 198–223. doi: 10.1111/j.1475-6811.2011.01353.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richeson J. A., Shelton J. N. (2005). Brief report: Thin slices of racial bias. Journal of Nonverbal Behavior, 29, 75–86. doi: 10.1007/s10919-004-0890-2 [DOI] [Google Scholar]
- Richeson J. A., Shelton J. N. (2007). Negotiating interracial interactions: Costs, consequences, and possibilities. Psychological Science, 16, 316–320. doi: 10.1111/j.1467-8721.2007.00528.x [DOI] [Google Scholar]
- Richeson J. A., Trawalter S., Shelton J. N. (2005). African Americans’ implicit racial attitudes and the depletion of executive function after interracial interactions. Social Cognition, 23, 336–352. doi: 10.1521/soco.2005.23.4.336 [DOI] [Google Scholar]
- Schoebi D. (2008). The coregulation of daily affect in marital relationships. Journal of Family Psychology, 22, 595–604. doi: 10.1037/0893-3200.22.3.595 [DOI] [PubMed] [Google Scholar]
- Sherwood A., Allen M. T., Fahrenberg J., Kelsey R. M., Lovallo W. R., van Doornen L. J. (1990). Methodological guidelines for impedance cardiography. Psychophysiology, 27, 1–23. doi: 10.1111/j.1469-8986.1990.tb02171.x [DOI] [PubMed] [Google Scholar]
- Stern C. D., West T. V. (2014). Circumventing anxiety during interpersonal encounters to provide interest in contact: An implementation intention approach. Journal of Experimental Social Psychology, 50, 82–93. doi: 10.1016/j.jesp.2013.09.008 [DOI] [Google Scholar]
- Tattersall A. J., Hockey G. R. (1995). Level of operator control and changes in heart rate variability during simulated flight maintenance. Human Factors, 37, 682–698. [DOI] [PubMed] [Google Scholar]
- Waters S. F., West T. V., Karnilowicz H. R., Mendes W. B. (2017). Affect contagion between mothers and infants: Exploring valence and touch. Journal of Experimental Psychology: General, 146, 1043–1051. doi: 10.1037/xge0000322 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waters S. F., West T. V., Mendes W. B. (2014). Stress contagion: Physiological covariation between mothers and infants. Psychological Science, 25, 934–942. doi: 10.1177/0956797613518352 [DOI] [PMC free article] [PubMed] [Google Scholar]
- West T. V. (2013). Repeated-measure methods with dyads. In Simpson J. A., Campbell L. (Eds.), The Oxford handbook of close relationships (pp. 731–749). New York, NY: Oxford University Press. [Google Scholar]
- West T. V., Pearson A. R., Stern C. (2014). Anxiety perseverance in intergroup interaction: When incidental explanations backfire. Journal of Personality and Social Psychology, 107, 825–843. doi: 10.1037/a0037941 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.