

Abstract
Anorexia nervosa (AN) is a serious eating disorder characterized by self-starvation and extreme weight loss. Pseudoatrophic brain changes are often readily visible in individual brain scans, and AN may be a valuable model disorder to study structural neuroplasticity. Structural magnetic resonance imaging studies have found reduced gray matter volume and cortical thinning in acutely underweight patients to normalize following successful treatment. However, some well-controlled studies have found regionally greater gray matter and persistence of structural alterations following long-term recovery. Findings from diffusion tensor imaging studies of white matter integrity and connectivity are also inconsistent. Furthermore, despite the severity of AN, the number of existing structural neuroimaging studies is still relatively low, and our knowledge of the underlying cellular and molecular mechanisms for macrostructural brain changes is rudimentary. We critically review the current state of structural neuroimaging in AN and discuss the potential neurobiological basis of structural brain alterations in the disorder, highlighting impediments to progress, recent developments, and promising future directions. In particular, we argue for the utility of more standardized data collection, adopting a connectomics approach to understanding brain network architecture, employing advanced magnetic resonance imaging methods that quantify biomarkers of brain tissue microstructure, integrating data from multiple imaging modalities, strategic longitudinal observation during weight restoration, and large-scale data pooling. Our overarching objective is to motivate carefully controlled research of brain structure in eating disorders, which will ultimately help predict therapeutic response and improve treatment.
Keywords: Anorexia nervosa, Connectome, Diffusion tensor imaging, Eating disorders, Magnetic resonance imaging, White matter microstructure
Anorexia nervosa (AN) is an eating disorder that typically develops during adolescence and affects more women than men (8:1 ratio) (1). It is the third most common chronic illness in adolescents with the highest mortality ratio of all psychiatric disorders (2). The diagnostic criteria include energy-intake restriction, significantly low body weight, fear of gaining weight, and a distorted body image (3). Clinically, a restricting type marked by food restriction can be distinguished from an impulsive binge-eating/purging type, where affected individuals eat large amounts of food in a short time period (bingeing) and/or attempt to counteract weight gain by vomiting or laxative use (purging). In fact, impulse control may serve as a trait-based model to characterize individuals with eating disorders along a continuum (4). The pathophysiology remains unknown, with many interacting developmental, genetic, environmental, and neurobiological factors (1,5,6). Understanding the underlying neurobiology will be key for developing more effective treatments (7–9).
Researchers have long searched for clues in brain structure (10). Advances in neuroimaging, including the analysis of T1-weighted (11–13) and diffusion-weighted magnetic resonance images (14,15) have yielded important insights (16,17). One well-established finding is that gray matter (GM) reduction associated with acute malnutrition is largely reversible with weight restoration, at least in nonchronic cases in younger patients (Figure 1) (18–20). Other studies have found regionally specific GM alterations (21–23) and compromised white matter (WM) integrity or connectivity independent from acute malnourishment (24,25), but findings are inconsistent (26–30). Relationships with clinical variables are similarly diverse (31,32). These discrepancies may be due to differences in methods and cohorts. Compared with other psychiatric disorders, few large-scale studies exist, and our understanding of the microstructural basis for macrostructural brain changes in AN is still rudimentary.
Figure 1.
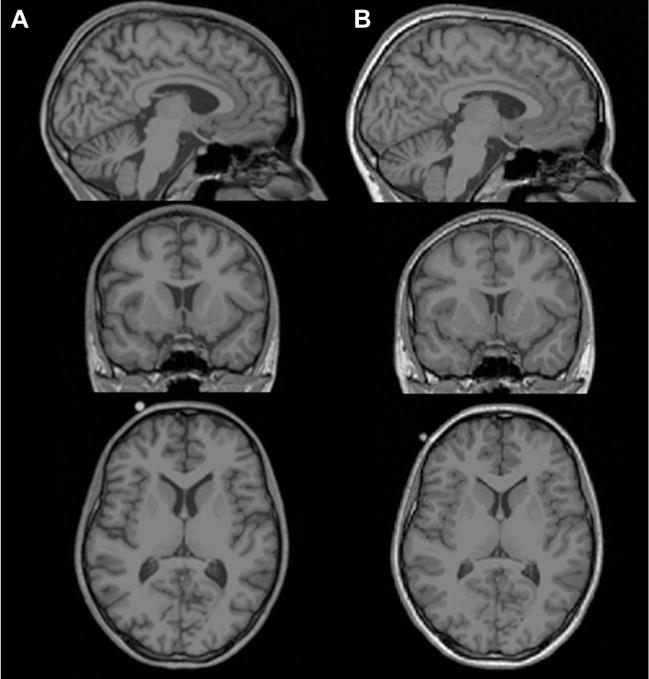
Rapid macrostructural brain changes in anorexia nervosa during weight restoration. Selected sagittal (top panel), coronal (middle panel) and axial (bottom panel) T1-weighted magnetic resonance images of (A) an acutely underweight adolescent patient with anorexia nervosa at admission to an inpatient eating disorder treatment (age, 15.6 years; body mass index, 16.2) and (B) the same patient 14 weeks later at discharge following weight restoration therapy (body mass index, 19.5). The images demonstrate widespread sulcal enlargement and marked ventricle dilation in illness and rapid normalization following nutritional rehabilitation. To illustrate the dynamic alteration in brain structure in anorexia nervosa objectively, this patient was chosen from the longitudinal sample from Bernardoni et al. (20) based on her standardized body mass index change score between admission and discharge, which was equal to the sample mean. Note that changes apparent in single-subject raw magnetic resonance images may not be representative of changes detected in group analyses of processed images, which may include regionally increased brain mass both in the underweight stage and after long-term recovery (22,49).
The main goals of this review are to 1) critically appraise the current state of structural neuroimaging in AN, 2) discuss cellular and molecular processes that may underlie morphological alterations in AN, and 3) outline a blueprint for future research elucidating the structural architecture of the brain in the disorder. While AN can be seen as a model disorder for understanding neuroadaptation to changing metabolic situations, the primary objective of this article is to motivate well-controlled research to help predict therapeutic response and eventually improve treatment. A cornerstone of this endeavor is promoting more standardized study protocols and large-scale data pooling and encouraging future studies with advanced technologies that integrate endocrine, metabolic, and (epi)genetic biomarkers.
STRUCTURAL NEUROIMAGING TECHNIQUES IN AN RESEARCH
Brain imaging can evaluate the structure and function of the living brain, pointing to mechanisms related to pathophysiology. A benefit of studying brain structure is that it tends to be more stable and less dependent on acute emotional and motivational states compared with functional neuroimaging. While early computed tomography studies in AN (33) were limited in their ability to detect localized alterations, recent structural magnetic resonance imaging (MRI) studies employed more sophisticated scanning and analysis procedures.
Volumetry based on T1-weighted MRI is one of the most frequently used techniques to study brain macrostructure. The introduction of voxel-based morphometry revolutionized structural neuroimaging, as it enabled automated quantification of GM and WM density from MRI intensity gradients (11). Most studies of AN focused on cortical and/or subcortical volume (26–29). However, cortical GM volume is a composite measure comprising two independent components: cortical thickness and surface area (34). Recent AN studies evaluated cortical thickness (20,35), folding, or gyrification (36,37).
Brain WM volume is often altered in psychiatric disease, including AN (28,29), but little is known about specific alterations in various disorders (38). Going beyond WM volume, several recent AN studies (39–41), including longitudinal observations (42–44), employed diffusion tensor imaging (DTI) (Supplemental Table S1), which provides information on WM integrity by quantifying water diffusivity along axons (45). The most common metric is fractional anisotropy (FA), a scalar value that describes the degree of anisotropic and/or directional diffusion associated with axon diameter, fiber packing density, membrane permeability, and myelination. FA is typically interpreted as a marker for WM integrity, but it is affected by several processes and cannot provide a definitive mechanistic explanation (46). To better characterize dynamic (pathological) microstructural abnormalities, DTI studies often include additional measures, including mean diffusivity—an overall measure of water diffusion—and measures of diffusion perpendicular or parallel to WM tracts (radial and axial diffusivity). Recent studies used DTI tractography to investigate WM fiber connectivity (25,43,47) measured by so-called streamlines that identify connection density and quality.
CURRENT STATE OF STRUCTURAL NEUROIMAGING RESEARCH IN AN
The increasing number of structural neuroimaging studies in AN (Figure 2) have been qualitatively reviewed and quantitatively meta-analyzed (Supplement) (26–30). The most comprehensive meta-analysis (28,29) concluded that globally reduced GM and WM volume in acutely underweight patients with AN normalizes during weight restoration, suggesting that alterations may merely reflect malnourishment. However, some well-controlled studies found larger frontal and subcortical volumes (22,39,48,49), and some regions, including cingulate gyrus (21,48,50) and cerebellum (31,32,51–53), appear to be altered more frequently than others. Furthermore, adolescents and adults may differ in the persistence and prognostic implications of structural alterations. Reports relating GM differences to clinical variables or cognitive function (19,22,32,54,55) are also heterogeneous—the only reliable predictor is body mass index (20,31,35,56). Similarly diverse findings from the small DTI literature in AN (Supplemental Table S1) hint at altered anisotropy and/or diffusivity in a range of WM tracts, including the cingulum, corpus callosum, superior longitudinal fasciculus, and, most frequently, fornix (17,30). However, the fornix is particularly susceptible to artifacts (57,58). Moreover, some studies report both increased and decreased FA in varying brain regions (40,42) or no differences at all (41).
Figure 2.
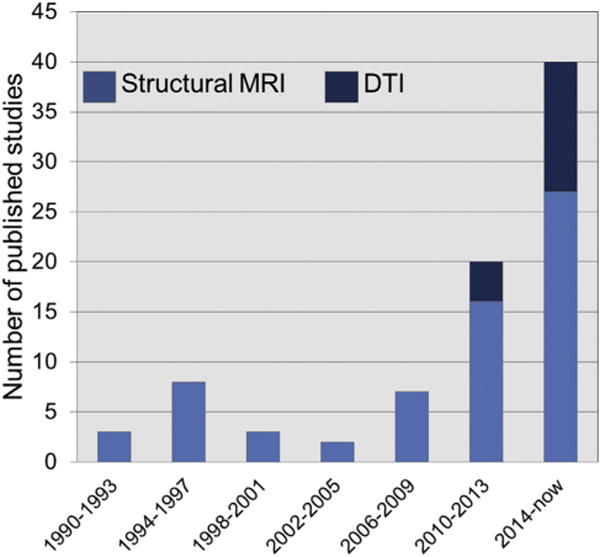
Structural magnetic resonance imaging (MRI) and diffusion tensor imaging (DTI) studies in anorexia nervosa. Graph depicts the increasing number of structural MRI and DTI studies in anorexia nervosa samples (n > 2 patients) published since the first known study in 1990 until July 2017 as identified with systematic PubMed and Scopus searches using the following terms: anorexia nervosa, eating disorders, MRI, magnetic resonance imaging, neuroimaging, DTI, diffusion tensor imaging, diffusion-weighted imaging. References for all studies counted in the graph are presented in the Supplement.
Thus, further research, particularly in weight-recovered individuals and accounting for potential differences between clinical subtypes, is needed (25,43,59,60). Recent analyses of WM structural and effective connectivity (43,44,47,60) point to alterations in reward-regulating frontostriatal circuitry, but a coherent pattern has yet to emerge (Figure 3). As discussed below, these inconsistencies are likely due to several factors, some of which are methodological, which should be addressed in future studies.
Figure 3.
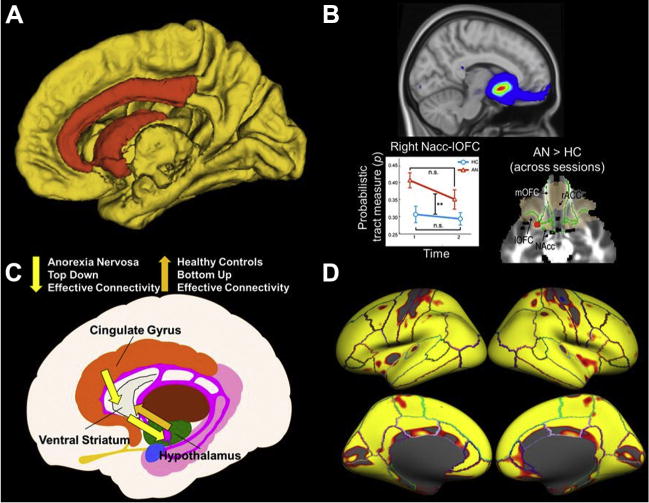
State-of-the-art structural neuroimaging research in anorexia nervosa (AN). (A) Using a connectomics approach (135,136) to compare network modularity (139) in diffusion tensor imaging data (whole-brain white matter tractography) acquired from weight-restored patients with AN (n = 24) and healthy control (HC) subjects (n = 31), Zhang et al. (60) found abnormal community structure in AN in a network composed of the caudate, pallidum, nucleus accumbens, and rostral and caudal anterior cingulate and posterior cingulate (orange regions). (B) In a longitudinal multimodal analysis (138,143) of diffusion tensor imaging and resting-state functional magnetic resonance imaging data collected in acutely underweight patients with AN (n = 22) and HC subjects (n = 18), Cha et al. (43) found increased structural connectivity, increased white matter fractional anisotropy, and increased functional connectivity as well as altered effective connectivity in a reward-related frontoaccumbal network in AN before treatment. Structural connectivity between the nucleus accumbens (NAcc) and left orbitofrontal cortex (lOFC) (top) continued to be abnormally increased relative to HC subjects following weight restoration (bottom left), and fractional anisotropy remained elevated (bottom right; red region). (C) In another multimodal analysis of diffusion tensor imaging and functional magnetic resonance imaging data during taste processing collected in acutely underweight patients with AN (n = 26), Frank et al. (47) found altered (increased and decreased) structural connectivity in food reward–regulating circuitry in patients with AN relative to HC subjects (n = 26) and an inverted pattern of effective connectivity between the hypothalamus and ventral striatum driven by the anterior cingulate. (D) In a longitudinal investigation of acutely underweight patients with AN (n = 35), Bernardoni et al. (20) found a global increase of cortical gray matter thickness (84% of the cortical surface) following brief weight restoration therapy (≈3 months following admission to a specialized nutritional rehabilitation program) and relative normalization compared with both long-term weight recovered patients with AN (n = 34) and HC subjects (n = 75). The heat map depicts regions of the lateral (top) and medial (bottom) surface of the brain in both hemispheres showing significantly increased cortical thickness between baseline (within 96 hours after admission) and follow-up measurement after a minimum 10% body mass index increase. ACC, anterior cingulate cortex; mOFC, medial orbitofrontal cortex.
Sample Size
Most studies have had small sample sizes (n < 20 patients) and low statistical power, inflating the likelihood of false-positive and false-negative results (61,62). A few recent studies compiled samples including more than 40 patients with AN (19,20,31), but none have conducted power calculations to determine sample sizes needed to detect reliable effects. Some studies have minimized the threat of unreliable findings by controlling for potential confounds (discussed below) and using stringent statistical thresholds controlled for multiple comparisons. Although the diverse results to date may reflect true heterogeneity in AN, progress will likely come from better-powered studies. As large samples are hard to recruit in a disorder often characterized by lack of illness insight and compliance, researchers should consider pooling data (see Advancing the Neuroscience of AN Through Collective Analysis below).
Nutritional Status
Modern neuroimaging can detect day-to-day changes in brain structure (63), and diet can influence brain morphometry (64). Studies should therefore carefully control and assess dietary intake, for example, by scanning at the same time of day after a standardized nutritional regimen. While some studies are vague regarding the nutritional status (37,40), others report scanning within the first days of patient care in a highly controlled environment (19,20,41–43), and still others report scanning after initial realimentation (1–2 weeks) (22,35,39). The former strategy captures AN in the natural disease state, but the latter reduces the effects of extreme malnutrition. Both strategies offer advantages, but future research should clarify the impact of initial realimentation by tracking body mass index change before scanning and longitudinal observation. Reversal of pseudoatrophy after weight restoration may be more rapid than previously thought (20,65).
Hydration
Hydration affects brain structure (66), and fluid intake is often abnormal in patients with AN (67). Three studies asked whether brain abnormalities in acute AN may reflect dehydration (22,24) by measuring urine specific gravity but found no evidence of dehydration or relationships with brain morphology (19,20,42). However, since self-induced dehydration and hyperhydration, together with impaired osmoregulation (68), are common in AN, and patients may “cheat the scale” by drinking excessively, targeted study is still needed, for instance, of water content (69).
Brain Development
The brain undergoes plastic changes across the life span (70), and it is challenging to disentangle developmental effects from insults due to malnutrition or neuroendocrine dysregulation in AN (71–73). Neurodevelopmental trajectories may be interrupted in AN, but the effects of malnutrition are at least one order of magnitude faster than age-related changes (20). While many AN studies attempt to account for maturation affects by covarying for age, others have adopted the often favorable strategy of focusing on homogeneous age ranges in separate analyses (19,22,39,41).
Extraneous Variables
In addition to controlling for eating disorder–specific factors, including clinical subtype (and subtype history), age of illness onset, and illness duration, future studies should control for other potential confounds, such as comorbidity (1,5), medication (74) including oral contraceptives (75), menstrual cycle (76), physical activity (77), and, in adolescents, pubertal stage (78). For analyses, use of covariates should be kept to a minimum to avoid overfitting statistical models. Depending on specific research questions, subanalysis of the influence of variables that are present only in the clinical group is important, both within-group (e.g., duration of illness) and between-(sub) group (e.g., medication) analyses.
Enhancing Reproducibility and Generalizability
Researchers are encouraged to follow best practices in neuroimaging (79) and to take proposed guidelines (Table 1) into consideration, many of which apply generally to eating disorders.
Table 1.
Factors to Consider in AN Structural Neuroimaging Research (Author Consensus)
| Category | Variable |
|---|---|
| Demographic Variables | Sample size (sufficient statistical power) |
| Age range (control for age effects and/or pairwise matching or restrict age range in analysis) | |
| In adolescents: pubertal stage (e.g., via self-report based on Tanner stage) | |
| Clinical Variables | AN subtype and subtype history (restricting vs. binge-eating/purging type) |
| Duration of illness | |
| Psychiatric and severe medical comorbidities (standardized assessment) | |
| Medication including oral contraceptives | |
| Time and weight gain between start of realimentation and scanning | |
| Nutritional regimen (e.g., scan before breakfast and after standard meal and/or dietary intake assessment) | |
| Hydration (e.g., by measuring urine specific gravity or using bioelectrical impedance analysis) | |
| Physical activity (self-report) | |
| Menstrual cycle (self-report) | |
| Study Design and Technology | Scan at standardized time of day |
| Analyze and control for head motion during MRI (especially for DTI) | |
| Use up-to-date MRI sequences (resolution, signal-to-noise) | |
| Control for different scanner platforms and changes in MRI sequences if applicable | |
| Statistics | Use up-to-date image analysis software |
| Systematic quality control of imaging data and outlier analysis (raw images and post-processing) | |
| Conservative use of covariates and appropriate analysis of potential confounds that occur only in the clinical group | |
| Control for multiple comparisons |
AN, anorexia nervosa; DTI, diffusion tensor imaging; MRI, magnetic resonance imaging.
POTENTIAL NEUROBIOLOGICAL MECHANISMS UNDERLYING STRUCTURAL BRAIN ALTERATIONS IN AN
Alterations in brain structure may result from multiple processes, but the underlying mechanisms remain poorly understood (80). In this section, we discuss possible explanations for brain morphological changes in AN, including 1) intracellular fluid moving into extracellular spaces because of altered oncotic (colloid osmotic) pressure or dehydration (81); 2) loss of glial or neuronal cells (e.g., apoptosis); 3) macronutrient and/or micronutrient deficiency (82,83); and 4) suppression of leptin (84), reduction of gonadal and thyroid hormones as well as neurotrophins, and cortisol increase (72). The last-mentioned possibilities could contribute to inadequate protein biosynthesis, which may lead to a) changes in or loss of dendritic spines, fiber reorganization, and delayed synaptogenesis (neuronal remodeling) and b) increased lipid catabolism and/or decreased myelination and shrinkage of glial cells (glial remodeling).
Dehydration and Oncotic Pressure
As noted above, targeted studies are needed to confirm preliminary evidence indicating that dehydration may not explain GM reduction in acute AN (19,20,42). Additional measurement methods, such as bioelectrical impedance analysis (with vector analysis), may also be helpful (85). It would also be important to differentiate between hypovolemia that is due to dehydration and hypovolemia that is due to loss of electrolytes, and researchers need to take into account that hemoconcentration that is due to volume depletion in AN can be masked by anemia (86). Fluctuations in GM and/or WM characteristics might also be explained by changes in oncotic pressure related to fluid shifts between the circulatory system and the interstitial space. Indeed, free abdominal fluid and pericardial effusion are often found in acutely underweight patients with AN (87). Although serum protein values are normal in AN (88,89), albumin levels have been associated with alterations in MRI signal intensity (90), and exploration of relationships with morphological changes may be worthwhile.
Apoptosis of Brain Cells
Postmortem histological data from an AN case study suggest an abnormally slender neuron type with a long basal dendritic field and reduced spine density and/or altered spine morphology indicative of cellular degeneration (91). Animal models of AN show reduced proliferation of brain cells but not massive cell loss (92). If apoptosis were the underlying cause of GM loss in acute AN, normalization following weight restoration would be unlikely unless neurogenesis occurred. However, the rapid GM increase during therapy (roughly 4% globally in only 3 months) (20) is hardly compatible with the comparatively sluggish turnover rate documented in select neuronal subpopulations in adults (1%–2% annually) (93). Preliminary findings from studies using neuronal and glial damage and/or activation markers, such as neuron-specific enolase and glial fibrillary acid protein (GFAP) (94), speak against a massive acute loss of brain cells. However, the normal age-dependent increase in peripheral GFAP (95), which may reflect expansion of mature, differentiated astrocytes (96), is absent in acute AN. Thus, reductions in size or changes in morphology of glia or neurons and dendrites seem more likely candidates for brain changes in AN.
Dietary Intake: Macronutrients and Micronutrients
Patients with AN disproportionally avoid consuming fats (97,98). Consequently, plasma concentrations of several lipids (including essential fatty acids) are lower or altered in patients with acute (99,100) but normal in recovered patients (101). Lipids (especially cholesterol, glycerophospholipids, and sphingolipids) are major components of brain structure (102). Many fatty acids can be produced endogenously, but some polyunsaturated fatty acids that are crucial for fluidity of neuronal membranes and for modification of the number and affinity of membrane bound enzymes, receptors, and ionic channels must be provided by diet (103). Deficiency of polyunsaturated fatty acids has been associated with structural brain alterations (64), but such relationships have yet to be explored in AN.
Insufficient intake of fats and other macronutrients may also be associated with inadequate micronutrient intake. Few studies have investigated intake or plasma levels of vitamins and trace elements (including iron), but there may be a reduction of several micronutrients, especially vitamin B12 and zinc (104). Zinc deficiency has been suggested to play a causal role in AN (105), but a recent large study found reduced levels in only 3% of the adult patients, and other vitamin or mineral deficiencies were found in only approximately 45% of patients—less than expected given frequent dermatological symptoms in AN suggestive of micronutrient deficiency (106).
Similarly, iron deficiency is surprisingly uncommon in AN (107). Iron is one of the many constituents of myelin, and iron concentration is related to WM fiber integrity (108). For example, higher serum transferrin levels are associated with lower fractional anisotropy in various WM tracts (109). Although the consequences of micronutrient deficiencies on neurological function have long been known (e.g., vitamin B1 and Wernicke encephalopathy), the study of how nutrients affect brain structure is relatively new (82,83,110) and has been neglected in AN research. Because evaluating micronutrient intake using questionnaires may be unreliable (111), measuring plasma levels may be preferable.
Endocrine System
The hypothalamic-pituitary-adrenal axis is hyperactive in AN, and the “hunger hormone” ghrelin is elevated, whereas the hypothalamic-pituitary-thyroid and hypothalamic-pituitary-gonadal axes and the adipocyte-derived hormone leptin is suppressed (71,72). Cortisol concentrations are elevated in AN (112), and cortisol is known to impact brain structure (113). Suppression of the hypothalamic-pituitary-thyroid and hypothalamic-pituitary-gonadal axis in AN has been attributed to restricted energy intake or increased energy expenditure and may be an adaptation to low energy availability—i.e., sexual reproduction would cost energy required for vital body functions. Beyond their role in sexual differentiation, gonadal steroids are critical for brain development, especially during adolescence (114). Estradiol, for example, protects neurons from excitotoxins and oxidative stress (115) and promotes dendritic growth, neurotransmitter synthesis, and neuronal depolarization (116). Despite endocrine dysregulation in AN, few studies have investigated relationships with brain structure. One computed tomography study (117) found associations between the width of cortical sulci and hypercortisolemia and hypogonadotropic hypogonadism, which is compatible with an MRI study reporting a link between follicle-stimulating hormone and regional GM volumes (51). Another study found an increase in global GM volume following weight recovery to correlate negatively with blood cortisol levels (54). Given established relationships between leptin and GM volume in healthy individuals (118), future studies might seek to clarify the relative contribution of leptin deficiency to structural brain anomalies in AN.
NEUROTROPHIC FACTORS
Neurotrophin brain-derived neurotrophic factor (BDNF), nerve growth factor, and glial-derived neurotrophic factor are proteins that modulate neuronal or glial development or regeneration and have been repeatedly related to brain changes. BDNF promotes the growth, differentiation, and survival of neurons (119) and plays a role in synaptic plasticity (120) as well as food intake and weight regulation (121). Reduced BDNF levels in acute AN are well established (122,123), as are variations in GM and WM morphometry as a function of BDNF polymorphisms (124). However, studies testing associations between circulating BDNF (and nerve growth factor and glial-derived neurotrophic factor) and brain structure in AN are lacking. Future studies should control for phasic effects of exercise on BDNF levels (125), as up to 70% of patients with AN report excessive exercise (126).
Glial and/or Neuronal Remodeling
Even if the factors discussed above do not cause apoptosis, they may contribute to structural brain alterations in AN by affecting the size or shape of neurons and glia. This hypothesis is supported by findings of deficits in somal size and density as well as dendritic branching and spine density of hippocampal pyramidal cells in protein-malnourished rats (127). Similarly, short apical dendrites, fewer spines, and dendritic spine abnormalities have been found in infants with severe malnutrition (128). Such mechanisms of neuronal remodeling have been demonstrated to underlie MRI-detectable brain volume increases following cognitive training in rodents (129). Thus, neuronal-glial remodeling seems to be a plausible explanation of structural brain changes in AN.
The most established animal model, the activity-based anorexia model (130,131), offers clues to specific mechanisms in AN. As noted above, one study showed decreased cell proliferation suggestive of reduced gliogenesis (92). Also in line with the glial remodeling hypothesis, another study (132) found chronically starved rats to have reduced cerebral cortex and corpus callosum volumes and a lower immunoreactive surface area for GFAP-positive astrocytes but not neurons or oligodendrocytes. Similarly, using an alternative AN rat model, acute dehydration-induced anorexia, another study found a reduced number of GFAP-positive astrocytes in the corpus callosum (133). Although most research focuses on neurons, astrocytes also regulate neuronal activity, protect against inflammation, support the blood-brain barrier, and supply energy to neurons (134). Smaller or less mature (remodeled), fewer, and less functional astrocytes may thus worsen the already precarious metabolic situation of neurons in acutely underweight patients with AN.
FUTURE DIRECTIONS OF STRUCTURAL NEUROIMAGING RESEARCH IN AN
Controlling for confounds is critical, but progress will likely be driven by exploring brain network architecture—the so-called structural connectome (135,136)—employing acquisition methods that quantify biomarkers of brain tissue microstructure (quantitative MRI [qMRI]) (137), integrating data from multiple modalities (multimodal imaging) (138) as well as longitudinal observation during weight restoration and large-scale data pooling. Conventional analytic approaches neglect the network complexity of the underlying neurocircuitry. Investigating interregional correlations of GM morphology (139) or systems-level changes in connectivity by modeling the neural network architecture of GM brain regions (nodes) highly interconnected by WM fiber bundles (edges) (135) may be a promising way to bring clarity to the diverse regional abnormalities in GM and WM integrity reported in AN. Several recent studies have adopted a connectomics approach to both DTI (Figure 3) (25,43,47,60) and functional MRI data (140,141), but we are far from a comprehensive understanding of altered connectivity in AN.
As T1-weighted MRI data are uninformative regarding underlying tissue properties and DTI data in AN are inconclusive (Supplemental Table S1), more sensitive measures are needed. Rapidly developing qMRI methods are promising in this respect. By decoupling different contrast mechanisms from the overall MRI signal, qMRI directly measures surrogate markers of microscopic tissue properties (137). For example, quantitative mapping of R1 (water proton relaxation rate) indexes myelination (142) and may help in understanding changes in WM volume and integrity in AN. Similarly, qMRI methods might clarify the potential contribution of hydration and/or dietary intake as noted above by assessing, for instance, water (69) and iron content (108). The only known qMRI study in AN found, in addition to both increased and decreased fractional anisotropy in a number of WM tracts, reduced R1 in several tracts, suggesting that impaired WM integrity reflects myelin loss (40).
Combining and integrating data obtained with different modalities such as qMRI and DTI (40) promises to foster a greater understanding of the underlying mechanisms of structural brain changes in AN than can be achieved using one method alone (138). For example, two recent multimodal imaging studies combining diffusion tractography and functional MRI found evidence linking alterations in WM structural connectivity and functional and effective connectivity between brain regions implicated in aberrant reward processing in AN (Figure 3) (43,47). To date, no AN studies have capitalized on the cross-information provided by integrating data from multiple modalities (e.g., GM density and WM integrity) as enabled, for instance, with joint independent component analysis (143).
Whereas the majority of previous structural neuroimaging studies in AN have been cross-sectional, a positive recent development has been the increase of longitudinal studies (20,42–44), which can be particularly informative regarding the biological mechanisms underlying morphological alterations associated with weight rehabilitation. For example, the rapid increase of cortical thickness following brief weight restoration therapy (3–4 months) speaks against hypotheses of apoptosis and neurogenesis and for a neuronal and/or glial remodeling explanation (20). Studying brain structure in AN at multiple time points during recovery is also potentially informative regarding therapeutic response, which may help find better treatments.
ADVANCING THE NEUROSCIENCE OF AN THROUGH COLLECTIVE ANALYSIS
As noted above, sample sizes in the structural neuroimaging literature on AN are typically small and do not, on their own, have adequate statistical power to answer more subtle research questions. For example, no study to date has compiled samples including individuals across the entire eating disorder spectrum to account for the transdiagnostic nature of AN and how brain structure might be related to symptom fluctuation along a restrictive-impulsive continuum or based on subtype history (4,144). Data from different studies are also often difficult to compare owing to methodological heterogeneity, but they are nonetheless invaluable because they provide clinical phenotyping. In 2009, researchers expert in large-scale neuroimaging or genetics formed a network called Enhancing NeuroImaging Genetics through Meta-Analysis (ENIGMA) (http://enigma.usc.edu/) (145). ENIGMA has now pooled MRI and genome-wide genotyping data from greater than 30,000 individuals to discover more than 20 genetic loci affecting brain structure. In psychiatric disorders, ENIGMA has completed some of the largest collaborative neuroimaging studies to date including well over 3000 datasets each, with studies in progress in 18 conditions (146).
ENIGMA has used both a mega-analysis model, where all phenotypic data are analyzed at a central site, and a meta-analysis model, where analyses are run cooperatively across consortia sites with a pre-agreed protocol. For example, universal pre-processing, including quality control and analysis of T1-weighted MRI, is run locally using FreeSurfer (http://freesurfer.net/) with pre-agreed covariates. Subsequently, site-specific regression coefficients are combined and meta-analyzed, weighting the results based on the sample sizes of each contributing cohort. Once a network is established, all members can submit proposals with new research questions.
In 2016, an ENIGMA AN consortium (http://enigma.usc.edu/ongoing/enigma-anorexia/) dedicated to understanding structural brain changes in patients over the course of weight restoration and recovery was founded. While harmonizing acquisition and analysis of neuroimaging data across different scanners is generally challenging, and it is unclear, for instance, how cultural factors might influence brain structure in different AN subtypes, increased statistical power achieved by data pooling will allow for the examination of the contributing or modulating factors discussed in this review as well as the specificity of morphological alterations compared with other (eating) disorders.
The ENIGMA screen of genetic loci that affect the brain has revealed overlap between loci associated with brain morphometry and schizophrenia (147). The Psychiatric Genetics Consortium recently identified a genome-wide significant locus in AN on chromosome 12 (148). Additional analysis suggests positive genetic correlations with schizophrenia, educational achievement, and neuroticism and negative correlations with high body mass index and metabolic markers (148–150). Future studies could jointly screen the genome for loci associated with AN and brain measures, allowing, for the first time, causal inferences about brain differences related to genetic risk for AN.
CONCLUSIONS
Progress in understanding structural brain alterations in AN (and eating disorders generally) has been hindered by small sample sizes, methodological heterogeneity, and inadequate control for confounding variables (Table 1). In addition to outlining some steps future studies should take to avoid the shortcomings in the previous literature, this review focused on the largely overlooked question of what neurobiological mechanisms (e.g., glial remodeling) may underlie macromorphological changes in AN. Recent momentum in the field, including the application of innovative technologies and advanced analysis, is encouraging. The quest to identify brain regions and biological processes involved in AN will be further expedited by collective pooling of imaging, clinical, and genetic data. Harmonized global data pooling as in ENIGMA offers a new source of power to test associations between brain measures, treatments, and outcomes and to establish the reproducibility and generality of findings, which is especially important in a severe illness such as AN with low prevalence.
Supplementary Material
Acknowledgments
This work was supported by the Deutsche Forschungsgemeinschaft Grant Nos. EH 367/5 and SFB 940 (to JAK and SE), Swiss Anorexia Nervosa Foundation (SE), Roland Ernst Stiftung (SE), National Institute of Mental Health Grant Nos. MH096777 and MH103436 (to GKWF), National Institutes of Health Grant No. EB020403 (to PMT and ENIGMA Consortium), National Institutes of Health Big Data to Knowledge program (PMT and ENIGMA Consortium), Kavli Foundation (PMT and ENIGMA Consortium), and other private and federal funders worldwide (PMT and ENIGMA Consortium).
We thank Simone Schöps, Arne Doose, and Wibke Naumann for their assistance.
Footnotes
DISCLOSURES
The authors report no biomedical financial interests or potential conflicts of interest.
Supplementary material cited in this article is available online at http://dx.doi.org/10.1016/j.biopsych.2017.08.011.
References
- 1.Zipfel S, Giel KE, Bulik CM, Hay P, Schmidt U. Anorexia nervosa: Aetiology, assessment, and treatment. Lancet Psychiatry. 2015;2:1099–1111. doi: 10.1016/S2215-0366(15)00356-9. [DOI] [PubMed] [Google Scholar]
- 2.Papadopoulos FC, Ekbom A, Brandt L, Ekselius L. Excess mortality, causes of death and prognostic factors in anorexia nervosa. Br J Psychiatry J Ment Sci. 2009;194:10–17. doi: 10.1192/bjp.bp.108.054742. [DOI] [PubMed] [Google Scholar]
- 3.American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders. 5th. Arlington, VA: American Psychiatric Publishing; 2013. (DSM-5) [Google Scholar]
- 4.Brooks SJ, Rask-Andersen M, Benedict C, Schiöth HB. A debate on current eating disorder diagnoses in light of neurobiological findings: Is it time for a spectrum model? BMC Psychiatry. 2012;12:76. doi: 10.1186/1471-244X-12-76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Treasure J, Zipfel S, Micali N, Wade T, Stice E, Claudino A, et al. Anorexia nervosa. Nat Rev Dis Primer. 2015;1:15074. doi: 10.1038/nrdp.2015.74. [DOI] [PubMed] [Google Scholar]
- 6.Frank GKW. The perfect storm—a bio-psycho-social risk model for developing and maintaining eating disorders. Front Behav Neurosci. 2016;10:44. doi: 10.3389/fnbeh.2016.00044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Frank GKW, Shott ME. The role of psychotropic medications in the management of anorexia nervosa: Rationale, evidence and future prospects. CNS Drugs. 2016;30:419–442. doi: 10.1007/s40263-016-0335-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hill L, Peck SK, Wierenga CE, Kaye WH. Applying neurobiology to the treatment of adults with anorexia nervosa. J Eat Disord. 2016;4:31. doi: 10.1186/s40337-016-0119-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kaye WH, Wierenga CE, Bailer UF, Simmons AN, Bischoff-Grethe A. Nothing tastes as good as skinny feels: The neurobiology of anorexia nervosa. Trends Neurosci. 2013;36:110–120. doi: 10.1016/j.tins.2013.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Heidrich R, Schmidt-Matthias H. Encephalographic findings in anorexia nervosa. Arch Psychiatr Nervenkrankh Ver Mit Z Gesamte Neurol Psychiatr. 1961;202:183–201. doi: 10.1007/BF00356099. [DOI] [PubMed] [Google Scholar]
- 11.Ashburner J, Friston KJ. Voxel-based morphometry—the methods. Neuroimage. 2000;11:805–821. doi: 10.1006/nimg.2000.0582. [DOI] [PubMed] [Google Scholar]
- 12.Dale AM, Fischl B, Sereno MI. Cortical surface-based analysis. I. Segmentation and surface reconstruction. Neuroimage. 1999;9:179–194. doi: 10.1006/nimg.1998.0395. [DOI] [PubMed] [Google Scholar]
- 13.Fischl B, Sereno MI, Dale AM. Cortical surface-based analysis. II. Inflation, flattening, and a surface-based coordinate system. Neuroimage. 1999;9:195–207. doi: 10.1006/nimg.1998.0396. [DOI] [PubMed] [Google Scholar]
- 14.Smith SM, Jenkinson M, Johansen-Berg H, Rueckert D, Nichols TE, Mackay CE, et al. Tract-based spatial statistics: Voxelwise analysis of multi-subject diffusion data. Neuroimage. 2006;31:1487–1505. doi: 10.1016/j.neuroimage.2006.02.024. [DOI] [PubMed] [Google Scholar]
- 15.Behrens TEJ, Woolrich MW, Jenkinson M, Johansen-Berg H, Nunes RG, Clare S, et al. Characterization and propagation of uncertainty in diffusion-weighted MR imaging. Magn Reson Med. 2003;50:1077–1088. doi: 10.1002/mrm.10609. [DOI] [PubMed] [Google Scholar]
- 16.Phillipou A, Rossell SL, Castle DJ. The neurobiology of anorexia nervosa: A systematic review. Aust N Z J Psychiatry. 2014;48:128–152. doi: 10.1177/0004867413509693. [DOI] [PubMed] [Google Scholar]
- 17.Frank GKW. Recent advances in neuroimaging to model eating disorder neurobiology. Curr Psychiatry Rep. 2015;17:559. doi: 10.1007/s11920-015-0559-z. [DOI] [PubMed] [Google Scholar]
- 18.Wagner A, Greer P, Bailer UF, Frank GK, Henry SE, Putnam K, et al. Normal brain tissue volumes after long-term recovery in anorexia and bulimia nervosa. Biol Psychiatry. 2006;59:291–293. doi: 10.1016/j.biopsych.2005.06.014. [DOI] [PubMed] [Google Scholar]
- 19.King JA, Geisler D, Ritschel F, Boehm I, Seidel M, Roschinski B, et al. Global cortical thinning in acute anorexia nervosa normalizes following long-term weight restoration. Biol Psychiatry. 2015;77:624–632. doi: 10.1016/j.biopsych.2014.09.005. [DOI] [PubMed] [Google Scholar]
- 20.Bernardoni F, King JA, Geisler D, Stein E, Jaite C, Nätsch D, et al. Weight restoration therapy rapidly reverses cortical thinning in anorexia nervosa: A longitudinal study. Neuroimage. 2016;130:214–222. doi: 10.1016/j.neuroimage.2016.02.003. [DOI] [PubMed] [Google Scholar]
- 21.Mühlau M, Gaser C, Ilg R, Conrad B, Leibl C, Cebulla MH, et al. Gray matter decrease of the anterior cingulate cortex in anorexia nervosa. Am J Psychiatry. 2007;164:1850–1857. doi: 10.1176/appi.ajp.2007.06111861. [DOI] [PubMed] [Google Scholar]
- 22.Frank GK, Shott ME, Hagman JO, Mittal VA. Alterations in brain structures related to taste reward circuitry in ill and recovered anorexia nervosa and in bulimia nervosa. Am J Psychiatry. 2013;170:1152–1160. doi: 10.1176/appi.ajp.2013.12101294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Roberto CA, Mayer LES, Brickman AM, Barnes A, Muraskin J, Yeung L-K, et al. Brain tissue volume changes following weight gain in adults with anorexia nervosa. Int J Eat Disord. 2011;44:406–411. doi: 10.1002/eat.20840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Via E, Zalesky A, Sánchez I, Forcano L, Harrison BJ, Pujol J, et al. Disruption of brain white matter microstructure in women with anorexia nervosa. J Psychiatry Neurosci. 2014;39:367–375. doi: 10.1503/jpn.130135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Shott ME, Pryor TL, Yang TT, Frank GKW. Greater insula white matter fiber connectivity in women recovered from anorexia nervosa. Neuropsychopharmacology. 2016;41:498–507. doi: 10.1038/npp.2015.172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Van den Eynde F, Suda M, Broadbent H, Guillaume S, Van den Eynde M, Steiger H, et al. Structural magnetic resonance imaging in eating disorders: A systematic review of voxel-based morphometry studies. Eur Eat Disord Rev. 2012;20:94–105. doi: 10.1002/erv.1163. [DOI] [PubMed] [Google Scholar]
- 27.Titova OE, Hjorth OC, Schiöth HB, Brooks SJ. Anorexia nervosa is linked to reduced brain structure in reward and somatosensory regions: A meta-analysis of VBM studies. BMC Psychiatry. 2013;13:110. doi: 10.1186/1471-244X-13-110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Seitz J, Bühren K, von Polier GG, Heussen N, Herpertz-Dahlmann B, Konrad K. Morphological changes in the brain of acutely ill and weight-recovered patients with anorexia nervosa. A meta-analysis and qualitative review. Z Kinder Jugendpsychiatr Psychother. 2014;42:7–17. doi: 10.1024/1422-4917/a000265. quiz 17–18. [DOI] [PubMed] [Google Scholar]
- 29.Seitz J, Herpertz-Dahlmann B, Konrad K. Brain morphological changes in adolescent and adult patients with anorexia nervosa. J Neural Transm (Vienna) 2016;123:949–959. doi: 10.1007/s00702-016-1567-9. [DOI] [PubMed] [Google Scholar]
- 30.Martin Monzon B, Hay P, Foroughi N, Touyz S. White matter alterations in anorexia nervosa: A systematic review of diffusion tensor imaging studies. World J Psychiatry. 2016;6:177–186. doi: 10.5498/wjp.v6.i1.177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Seitz J, Walter M, Mainz V, Herpertz-Dahlmann B, Konrad K, von Polier G. Brain volume reduction predicts weight development in adolescent patients with anorexia nervosa. J Psychiatr Res. 2015;68:228–237. doi: 10.1016/j.jpsychires.2015.06.019. [DOI] [PubMed] [Google Scholar]
- 32.Fonville L, Giampietro V, Williams SCR, Simmons A, Tchanturia K. Alterations in brain structure in adults with anorexia nervosa and the impact of illness duration. Psychol Med. 2014;44:1965–1975. doi: 10.1017/S0033291713002389. [DOI] [PubMed] [Google Scholar]
- 33.Kohlmeyer K, Lehmkuhl G, Poutska F. Computed tomography of anorexia nervosa. AJNR Am J Neuroradiol. 1983;4:437–438. [PMC free article] [PubMed] [Google Scholar]
- 34.Winkler AM, Kochunov P, Blangero J, Almasy L, Zilles K, Fox PT, et al. Cortical thickness or grey matter volume? The importance of selecting the phenotype for imaging genetics studies. Neuroimage. 2010;53:1135–1146. doi: 10.1016/j.neuroimage.2009.12.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lavagnino L, Amianto F, Mwangi B, D’Agata F, Spalatro A, Zunta Soares GB, et al. The relationship between cortical thickness and body mass index differs between women with anorexia nervosa and healthy controls. Psychiatry Res. 2016;248:105–109. doi: 10.1016/j.pscychresns.2016.01.002. [DOI] [PubMed] [Google Scholar]
- 36.Schultz CC, Wagner G, de la Cruz F, Berger S, Reichenbach JR, Sauer H, Bär KJ. Evidence for alterations of cortical folding in anorexia nervosa. Eur Arch Psychiatry Clin Neurosci. 2017;267:41–49. doi: 10.1007/s00406-015-0666-1. [DOI] [PubMed] [Google Scholar]
- 37.Favaro A, Tenconi E, Degortes D, Manara R, Santonastaso P. Gyrification brain abnormalities as predictors of outcome in anorexia nervosa. Hum Brain Mapp. 2015;36:5113–5122. doi: 10.1002/hbm.22998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Giedd JN, Raznahan A, Alexander-Bloch A, Schmitt E, Gogtay N, Rapoport JL. Child psychiatry branch of the National Institute of Mental Health longitudinal structural magnetic resonance imaging study of human brain development. Neuropsychopharmacology. 2015;40:43–49. doi: 10.1038/npp.2014.236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Frank GK, Shott ME, Hagman JO, Yang TT. Localized brain volume and white matter integrity alterations in adolescent anorexia nervosa. J Am Acad Child Adolesc Psychiatry. 2013;52:1066–1075.e5. doi: 10.1016/j.jaac.2013.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Travis KE, Golden NH, Feldman HM, Solomon M, Nguyen J, Mezer A, et al. Abnormal white matter properties in adolescent girls with anorexia nervosa. Neuroimage Clin. 2015;9:648–659. doi: 10.1016/j.nicl.2015.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Pfuhl G, King JA, Geisler D, Roschinski B, Ritschel F, Seidel M, et al. Preserved white matter microstructure in young patients with anorexia nervosa? Hum Brain Mapp. 2016;37:4069–4083. doi: 10.1002/hbm.23296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Vogel K, Timmers I, Kumar V, Nickl-Jockschat T, Bastiani M, Roebroek A, et al. White matter microstructural changes in adolescent anorexia nervosa including an exploratory longitudinal study. Neuroimage Clin. 2016;11:614–621. doi: 10.1016/j.nicl.2016.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Cha J, Ide JS, Bowman FD, Simpson HB, Posner J, Steinglass JE. Abnormal reward circuitry in anorexia nervosa: A longitudinal, multimodal MRI study. Hum Brain Mapp. 2016;37:3835–3846. doi: 10.1002/hbm.23279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Olivo G, Wiemerslage L, Swenne I, Zhukowsky C, Salonen-Ros H, Larsson E-M, et al. Limbic-thalamo-cortical projections and reward-related circuitry integrity affects eating behavior: A longitudinal DTI study in adolescents with restrictive eating disorders. PloS One. 2017;12:e0172129. doi: 10.1371/journal.pone.0172129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Le Bihan D. Looking into the functional architecture of the brain with diffusion MRI. Nat Rev Neurosci. 2003;4:469–480. doi: 10.1038/nrn1119. [DOI] [PubMed] [Google Scholar]
- 46.Jones DK, Cercignani M. Twenty-five pitfalls in the analysis of diffusion MRI data. NMR Biomed. 2010;23:803–820. doi: 10.1002/nbm.1543. [DOI] [PubMed] [Google Scholar]
- 47.Frank GKW, Shott ME, Riederer J, Pryor TL. Altered structural and effective connectivity in anorexia and bulimia nervosa in circuits that regulate energy and reward homeostasis. Transl Psychiatry. 2016;6:e932. doi: 10.1038/tp.2016.199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Brooks SJ, Barker GJ, O’Daly OG, Brammer M, Williams SCR, Benedict C, et al. Restraint of appetite and reduced regional brain volumes in anorexia nervosa: A voxel-based morphometric study. BMC Psychiatry. 2011;11:179. doi: 10.1186/1471-244X-11-179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Favaro A, Tenconi E, Degortes D, Manara R, Santonastaso P. Effects of obstetric complications on volume and functional connectivity of striatum in anorexia nervosa patients. Int J Eat Disord. 2014;47:686–695. doi: 10.1002/eat.22320. [DOI] [PubMed] [Google Scholar]
- 50.Friederich H-C, Walther S, Bendszus M, Biller A, Thomann P, Zeigermann S, et al. Grey matter abnormalities within cortico-limbic-striatal circuits in acute and weight-restored anorexia nervosa patients. Neuroimage. 2012;59:1106–1113. doi: 10.1016/j.neuroimage.2011.09.042. [DOI] [PubMed] [Google Scholar]
- 51.Mainz V, Schulte-Rüther M, Fink GR, Herpertz-Dahlmann B, Konrad K. Structural brain abnormalities in adolescent anorexia nervosa before and after weight recovery and associated hormonal changes. Psychosom Med. 2012;74:574–582. doi: 10.1097/PSY.0b013e31824ef10e. [DOI] [PubMed] [Google Scholar]
- 52.Boghi A, Sterpone S, Sales S, D’Agata F, Bradac GB, Zullo G, Munno D. In vivo evidence of global and focal brain alterations in anorexia nervosa. Psychiatry Res. 2011;192:154–159. doi: 10.1016/j.pscychresns.2010.12.008. [DOI] [PubMed] [Google Scholar]
- 53.Amianto F, Caroppo P, D’Agata F, Spalatro A, Lavagnino L, Caglio M, et al. Brain volumetric abnormalities in patients with anorexia and bulimia nervosa: A voxel-based morphometry study. Psychiatry Res. 2013;213:210–216. doi: 10.1016/j.pscychresns.2013.03.010. [DOI] [PubMed] [Google Scholar]
- 54.Castro-Fornieles J, Bargalló N, Lázaro L, Andrés S, Falcon C, Plana MT, Junqué C. A cross-sectional and follow-up voxel-based morphometric MRI study in adolescent anorexia nervosa. J Psychiatr Res. 2009;43:331–340. doi: 10.1016/j.jpsychires.2008.03.013. [DOI] [PubMed] [Google Scholar]
- 55.Suchan B, Busch M, Schulte D, Grönemeyer D, Grönermeyer D, Herpertz S, Vocks S. Reduction of gray matter density in the extrastriate body area in women with anorexia nervosa. Behav Brain Res. 2010;206:63–67. doi: 10.1016/j.bbr.2009.08.035. [DOI] [PubMed] [Google Scholar]
- 56.Bomba M, Riva A, Veggo F, Grimaldi M, Morzenti S, Neri F, Nacinovich R. Impact of speed and magnitude of weight loss on the development of brain trophic changes in adolescents with anorexia nervosa: A case control study. Ital J Pediatr. 2013;39:14. doi: 10.1186/1824-7288-39-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Jones DK, Knösche TR, Turner R. White matter integrity, fiber count, and other fallacies: The do’s and don’ts of diffusion MRI. Neuroimage. 2013;73:239–254. doi: 10.1016/j.neuroimage.2012.06.081. [DOI] [PubMed] [Google Scholar]
- 58.Kaufmann LK, Baur V, Hänggi J, Jäncke L, Piccirelli M, Kollias S, et al. Fornix underwater? Ventricular enlargement biases forniceal diffusion magnetic resonance imaging indices in anorexia nervosa. Biol Psychiatry Cogn Neurosci Neuroimaging. 2017;2:430–437. doi: 10.1016/j.bpsc.2017.03.014. [DOI] [PubMed] [Google Scholar]
- 59.Yau W-YW, Bischoff-Grethe A, Theilmann RJ, Torres L, Wagner A, Kaye WH, Fennema-Notestine C. Alterations in white matter microstructure in women recovered from anorexia nervosa. Int J Eat Disord. 2013;46:701–708. doi: 10.1002/eat.22154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Zhang A, Leow A, Zhan L, GadElkarim J, Moody T, Khalsa S, et al. Brain connectome modularity in weight-restored anorexia nervosa and body dysmorphic disorder. Psychol Med. 2016;46:2785–2797. doi: 10.1017/S0033291716001458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Button KS, Ioannidis JPA, Mokrysz C, Nosek BA, Flint J, Robinson ESJ, Munafò MR. Power failure: Why small sample size undermines the reliability of neuroscience. Nat Rev Neurosci. 2013;14:365–376. doi: 10.1038/nrn3475. [DOI] [PubMed] [Google Scholar]
- 62.Fusar-Poli P, Radua J, Frascarelli M, Mechelli A, Borgwardt S, Di Fabio F, et al. Evidence of reporting biases in voxel-based morphometry (VBM) studies of psychiatric and neurological disorders. Hum Brain Mapp. 2014;35:3052–3065. doi: 10.1002/hbm.22384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Trefler A, Sadeghi N, Thomas AG, Pierpaoli C, Baker CI, Thomas C. Impact of time-of-day on brain morphometric measures derived from T1-weighted magnetic resonance imaging. Neuroimage. 2016;133:41–52. doi: 10.1016/j.neuroimage.2016.02.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.McNamara RK, Asch RH, Lindquist DM, Krikorian R. Role of polyunsaturated fatty acids in human brain structure and function across the lifespan: An update on neuroimaging findings [published online ahead of 9] Prostaglandins Leukot Essent Fatty Acids. 2017 doi: 10.1016/j.plefa.2017.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Suda M, Narita K, Takei Y, Aoyama Y, Takahashi K, Yuki N, et al. Changes in gray matter volume with rapid body weight changes in anorexia nervosa: A voxel-based morphometric study. Biol Psychiatry. 2011;70:e35–e36. doi: 10.1016/j.biopsych.2011.03.034. [DOI] [PubMed] [Google Scholar]
- 66.Streitbürger D-P, Möller HE, Tittgemeyer M, Hund-Georgiadis M, Schroeter ML, Mueller K. Investigating structural brain changes of dehydration using voxel-based morphometry. PloS One. 2012;7:e44195. doi: 10.1371/journal.pone.0044195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Hart S, Abraham S, Luscombe G, Russell J. Fluid intake in patients with eating disorders. Int J Eat Disord. 2005;38:55–59. doi: 10.1002/eat.20155. [DOI] [PubMed] [Google Scholar]
- 68.Evrard F, da Cunha MP, Lambert M, Devuyst O. Impaired osmoregulation in anorexia nervosa: A case-control study. Nephrol Dial Transplant. 2004;19:3034–3039. doi: 10.1093/ndt/gfh507. [DOI] [PubMed] [Google Scholar]
- 69.Meyers SM, Tam R, Lee JS, Kolind SH, Vavasour IM, Mackie E, et al. Does hydration status affect MRI measures of brain volume or water content? J Magn Reson Imaging. 2016;44:296–304. doi: 10.1002/jmri.25168. [DOI] [PubMed] [Google Scholar]
- 70.Shaw P, Kabani NJ, Lerch JP, Eckstrand K, Lenroot R, Gogtay N, et al. Neurodevelopmental trajectories of the human cerebral cortex. J Neurosci. 2008;28:3586–3594. doi: 10.1523/JNEUROSCI.5309-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Bailer UF, Kaye WH. A review of neuropeptide and neuroendocrine dysregulation in anorexia and bulimia nervosa. Curr Drug Targets CNS Neurol Disord. 2003;2:53–59. doi: 10.2174/1568007033338689. [DOI] [PubMed] [Google Scholar]
- 72.Schorr M, Miller KK. The endocrine manifestations of anorexia nervosa: Mechanisms and management. Nat Rev Endocrinol. 2017;13:174–186. doi: 10.1038/nrendo.2016.175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Prince AC, Brooks SJ, Stahl D, Treasure J. Systematic review and meta-analysis of the baseline concentrations and physiologic responses of gut hormones to food in eating disorders. Am J Clin Nutr. 2009;89:755–765. doi: 10.3945/ajcn.2008.27056. [DOI] [PubMed] [Google Scholar]
- 74.Navari S, Dazzan P. Do antipsychotic drugs affect brain structure? A systematic and critical review of MRI findings. Psychol Med. 2009;39:1763–1777. doi: 10.1017/S0033291709005315. [DOI] [PubMed] [Google Scholar]
- 75.Pletzer B, Kronbichler M, Kerschbaum H. Differential effects of androgenic and anti-androgenic progestins on fusiform and frontal gray matter volume and face recognition performance. Brain Res. 2015;1596:108–115. doi: 10.1016/j.brainres.2014.11.025. [DOI] [PubMed] [Google Scholar]
- 76.Pletzer B, Kronbichler M, Aichhorn M, Bergmann J, Ladurner G, Kerschbaum HH. Menstrual cycle and hormonal contraceptive use modulate human brain structure. Brain Res. 2010;1348:55–62. doi: 10.1016/j.brainres.2010.06.019. [DOI] [PubMed] [Google Scholar]
- 77.Gümmer R, Giel KE, Schag K, Resmark G, Junne FP, Becker S, et al. High levels of physical activity in anorexia nervosa: A systematic review. Eur Eat Disord Rev. 2015;23:333–344. doi: 10.1002/erv.2377. [DOI] [PubMed] [Google Scholar]
- 78.Blakemore SJ, Burnett S, Dahl RE. The role of puberty in the developing adolescent brain. Hum Brain Mapp. 2010;31:926–933. doi: 10.1002/hbm.21052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Nichols TE, Das S, Eickhoff SB, Evans AC, Glatard T, Hanke M, et al. Best practices in data analysis and sharing in neuroimaging using MRI. Nat Neurosci. 2017;20:299–303. doi: 10.1038/nn.4500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Zatorre RJ, Fields RD, Johansen-Berg H. Plasticity in gray and white: Neuroimaging changes in brain structure during learning. Nat Neurosci. 2012;15:528–536. doi: 10.1038/nn.3045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Artmann H, Grau H, Adelmann M, Schleiffer R. Reversible and non-reversible enlargement of cerebrospinal fluid spaces in anorexia nervosa. Neuroradiology. 1985;27:304–312. doi: 10.1007/BF00339562. [DOI] [PubMed] [Google Scholar]
- 82.Bourre JM. Effects of nutrients (in food) on the structure and function of the nervous system: Update on dietary requirements for brain. Part 1: Micronutrients. J Nutr Health Aging. 2006;10:377–385. [PubMed] [Google Scholar]
- 83.Bourre JM. Effects of nutrients (in food) on the structure and function of the nervous system: Update on dietary requirements for brain. Part 2: Macronutrients. J Nutr Health Aging. 2006;10:386–399. [PubMed] [Google Scholar]
- 84.Matochik JA, London ED, Yildiz BO, Ozata M, Caglayan S, DePaoli AM, et al. Effect of leptin replacement on brain structure in genetically leptin-deficient adults. J Clin Endocrinol Metab. 2005;90:2851–2854. doi: 10.1210/jc.2004-1979. [DOI] [PubMed] [Google Scholar]
- 85.Haas V, Riedl A, Hofmann T, Nischan A, Burghardt R, Boschmann M, Klapp B. Bioimpedance and bioimpedance vector analysis in patients with anorexia nervosa. Eur Eat Disord Rev. 2012;20:400–405. doi: 10.1002/erv.1166. [DOI] [PubMed] [Google Scholar]
- 86.Caregaro L, Di Pascoli L, Favaro A, Nardi M, Santonastaso P. Sodium depletion and hemoconcentration: Overlooked complications in patients with anorexia nervosa? Nutrition. 2005;21:438–445. doi: 10.1016/j.nut.2004.08.022. [DOI] [PubMed] [Google Scholar]
- 87.Docx MKF, Gewillig M, Simons A, Vandenberghe P, Weyler J, Ramet J, Mertens L. Pericardial effusions in adolescent girls with anorexia nervosa: Clinical course and risk factors. Eat Disord. 2010;18:218–225. doi: 10.1080/10640261003719484. [DOI] [PubMed] [Google Scholar]
- 88.Swenne I. The significance of routine laboratory analyses in the assessment of teenage girls with eating disorders and weight loss. Eat Weight Disord. 2004;9:269–278. doi: 10.1007/BF03325081. [DOI] [PubMed] [Google Scholar]
- 89.Ehrlich S, Querfeld U, Pfeiffer E. Refeeding oedema: An important complication in the treatment of anorexia nervosa. Eur Child Adolesc Psychiatry. 2006;15:241–243. doi: 10.1007/s00787-006-0528-5. [DOI] [PubMed] [Google Scholar]
- 90.Pirker A, Kramer L, Voller B, Loader B, Auff E, Prayer D. Type of edema in posterior reversible encephalopathy syndrome depends on serum albumin levels: An MR imaging study in 28 patients. AJNR Am J Neuroradiol. 2011;32:527–531. doi: 10.3174/ajnr.A2332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Neumarker KJ, Dudeck U, Meyer U, Neumarker U, Schulz E, Schonheit B. Anorexia nervosa and sudden death in childhood: Clinical data and results obtained from quantitative neurohistological investigations of cortical neurons. Eur Arch Psychiatry Clin Neurosci. 1997;247:16–22. doi: 10.1007/BF02916248. [DOI] [PubMed] [Google Scholar]
- 92.Barbarich-Marsteller NC, Fornal CA, Takase LF, Bocarsly ME, Arner C, Walsh BT, et al. Activity-based anorexia is associated with reduced hippocampal cell proliferation in adolescent female rats. Behav Brain Res. 2013;236:251–257. doi: 10.1016/j.bbr.2012.08.047. [DOI] [PubMed] [Google Scholar]
- 93.Bergmann O, Spalding KL, Frisén J. Adult neurogenesis in humans. Cold Spring Harb Perspect Biol. 2015;7:a018994. doi: 10.1101/cshperspect.a018994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Ehrlich S, Burghardt R, Weiss D, Salbach-Andrae H, Craciun EM, Goldhahn K, et al. Glial and neuronal damage markers in patients with anorexia nervosa. J Neural Transm (Vienna) 2008;115:921–927. doi: 10.1007/s00702-008-0033-8. [DOI] [PubMed] [Google Scholar]
- 95.Rosengren LE, Ahlsén G, Belfrage M, Gillberg C, Haglid KG, Hamberger A. A sensitive ELISA for glial fibrillary acidic protein: Application in CSF of children. J Neurosci Methods. 1992;44:113–119. doi: 10.1016/0165-0270(92)90004-w. [DOI] [PubMed] [Google Scholar]
- 96.Nichols NR, Day JR, Laping NJ, Johnson SA, Finch CE. GFAP mRNA increases with age in rat and human brain. Neurobiol Aging. 1993;14:421–429. doi: 10.1016/0197-4580(93)90100-p. [DOI] [PubMed] [Google Scholar]
- 97.Misra M, Tsai P, Anderson EJ, Hubbard JL, Gallagher K, Soyka LA, et al. Nutrient intake in community-dwelling adolescent girls with anorexia nervosa and in healthy adolescents. Am J Clin Nutr. 2006;84:698–706. doi: 10.1093/ajcn/84.4.698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Fernstrom MH, Weltzin TE, Neuberger S, Srinivasagam N, Kaye WH. Twenty-four-hour food intake in patients with anorexia nervosa and in healthy control subjects. Biol Psychiatry. 1994;36:696–702. doi: 10.1016/0006-3223(94)91179-7. [DOI] [PubMed] [Google Scholar]
- 99.Holman RT, Adams CE, Nelson RA, Grater SJ, Jaskiewicz JA, Johnson SB, Erdman JW. Patients with anorexia nervosa demonstrate deficiencies of selected essential fatty acids, compensatory changes in nonessential fatty acids and decreased fluidity of plasma lipids. J Nutr. 1995;125:901–907. doi: 10.1093/jn/125.4.901. [DOI] [PubMed] [Google Scholar]
- 100.Shih PB. Integrating multi-omics biomarkers and postprandial metabolism to develop personalized treatment for anorexia nervosa. Prostaglandins Other Lipid Mediat. 2017;132:69–76. doi: 10.1016/j.prostaglandins.2017.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Shih PB, Yang J, Morisseau C, German JB, Zeeland AA, Armando AM, et al. Dysregulation of soluble epoxide hydrolase and lipidomic profiles in anorexia nervosa. Mol Psychiatry. 2016;21:537–546. doi: 10.1038/mp.2015.26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Piomelli D, Astarita G, Rapaka R. A neuroscientist’s guide to lipidomics. Nat Rev Neurosci. 2007;8:743–754. doi: 10.1038/nrn2233. [DOI] [PubMed] [Google Scholar]
- 103.Yehuda S, Rabinovitz S, Mostofsky DI. PUFA: Mediators for the nervous, endocrine, and immune systems. In: Mostofsky DI, Yehuda S, Salem N, editors. Fatty Acids: Physiologic and Behavioral Functions. Totowa, NJ: Humana Press; 2001. pp. 403–420. [Google Scholar]
- 104.Marzola E, Nasser JA, Hashim SA, Shih PA, Kaye WH. Nutritional rehabilitation in anorexia nervosa: Review of the literature and implications for treatment. BMC Psychiatry. 2013;13:290. doi: 10.1186/1471-244X-13-290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Birmingham CL, Goldner EM, Bakan R. Controlled trial of zinc supplementation in anorexia nervosa. Int J Eat Disord. 1994;15:251–255. [PubMed] [Google Scholar]
- 106.Achamrah N, Coëffier M, Rimbert A, Charles J, Folope V, Petit A, et al. Micronutrient status in 153 patients with anorexia nervosa. Nutrients. 2017;9:225. doi: 10.3390/nu9030225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Papillard-Marechal S, Sznajder M, Hurtado-Nedelec M, Alibay Y, Martin-Schmitt C, Dehoux M, et al. Iron metabolism in patients with anorexia nervosa: Elevated serum hepcidin concentrations in the absence of inflammation. Am J Clin Nutr. 2012;95:548–554. doi: 10.3945/ajcn.111.025817. [DOI] [PubMed] [Google Scholar]
- 108.Stüber C, Morawski M, Schäfer A, Labadie C, Wähnert M, Leuze C, et al. Myelin and iron concentration in the human brain: A quantitative study of MRI contrast. Neuroimage. 2014;93(part 1):95–106. doi: 10.1016/j.neuroimage.2014.02.026. [DOI] [PubMed] [Google Scholar]
- 109.Jahanshad N, Kohannim O, Hibar DP, Stein JL, McMahon KL, de Zubicaray GI, et al. Brain structure in healthy adults is related to serum transferrin and the H63D polymorphism in the HFE gene. Proc Natl Acad Sci U S A. 2012;109:E851–E859. doi: 10.1073/pnas.1105543109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Jahanshad N, Rajagopalan P, Thompson PM. Neuroimaging, nutrition, and iron-related genes. Cell Mol Life Sci. 2013;70:4449–4461. doi: 10.1007/s00018-013-1369-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Kipnis V, Midthune D, Freedman L, Bingham S, Day NE, Riboli E, et al. Bias in dietary-report instruments and its implications for nutritional epidemiology. Public Health Nutr. 2002;5:915–923. doi: 10.1079/PHN2002383. [DOI] [PubMed] [Google Scholar]
- 112.Warren MP. Endocrine manifestations of eating disorders. J Clin Endocrinol Metab. 2011;96:333–343. doi: 10.1210/jc.2009-2304. [DOI] [PubMed] [Google Scholar]
- 113.van der Werff SJ, Andela CD, Nienke Pannekoek J, Meijer OC, van Buchem MA, Rombouts SA, et al. Widespread reductions of white matter integrity in patients with long-term remission of Cushing’s disease. Neuroimage Clin. 2014;4:659–667. doi: 10.1016/j.nicl.2014.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.McCarthy MM. Estradiol and the developing brain. Physiol Rev. 2008;88:91–124. doi: 10.1152/physrev.00010.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Green PS, Simpkins JW. Neuroprotective effects of estrogens: Potential mechanisms of action. Int J Dev Neurosci. 2000;18:347–358. doi: 10.1016/s0736-5748(00)00017-4. [DOI] [PubMed] [Google Scholar]
- 116.Rao ML, Kölsch H. Effects of estrogen on brain development and neuroprotection—implications for negative symptoms in schizophrenia. Psychoneuroendocrinology. 2003;28(suppl 2):83–96. doi: 10.1016/s0306-4530(02)00126-9. [DOI] [PubMed] [Google Scholar]
- 117.Nogal P, Pniewska-Siark B, Lewinski A. Relation of trophic changes in the central nervous system, measured by the width of cortical sulci, to the clinical course of anorexia nervosa (II) Neuro Endocrinol Lett. 2008;29:879–883. [PubMed] [Google Scholar]
- 118.Pannacciulli N, Le DS, Chen K, Reiman EM, Krakoff J. Relationships between plasma leptin concentrations and human brain structure: A voxel-based morphometric study. Neurosci Lett. 2007;412:248–253. doi: 10.1016/j.neulet.2006.11.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Park H, Poo M. Neurotrophin regulation of neural circuit development and function. Nat Rev Neurosci. 2013;14:7–23. doi: 10.1038/nrn3379. [DOI] [PubMed] [Google Scholar]
- 120.Waterhouse EG, Xu B. New insights into the role of brain-derived neurotrophic factor in synaptic plasticity. Mol Cell Neurosci. 2009;42:81–89. doi: 10.1016/j.mcn.2009.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Kernie SG, Liebl DJ, Parada LF. BDNF regulates eating behavior and locomotor activity in mice. EMBO J. 2000;19:1290–1300. doi: 10.1093/emboj/19.6.1290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Ehrlich S, Salbach-Andrae H, Eckart S, Merle JV, Burghardt R, Pfeiffer E, et al. Serum brain-derived neurotrophic factor and peripheral indicators of the serotonin system in underweight and weight-recovered adolescent girls and women with anorexia nervosa. J Psychiatry Neurosci. 2009;34:323–329. [PMC free article] [PubMed] [Google Scholar]
- 123.Brandys MK, Kas MJH, van Elburg AA, Campbell IC, Adan RAH. A meta-analysis of circulating BDNF concentrations in anorexia nervosa. World J Biol Psychiatry. 2011;12:444–454. doi: 10.3109/15622975.2011.562244. [DOI] [PubMed] [Google Scholar]
- 124.Chiang MC, Barysheva M, Toga AW, Medland SE, Hansell NK, James MR, et al. BDNF gene effects on brain circuitry replicated in 455 twins. Neuroimage. 2011;55:448–454. doi: 10.1016/j.neuroimage.2010.12.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Szuhany KL, Bugatti M, Otto MW. A meta-analytic review of the effects of exercise on brain-derived neurotrophic factor. J Psychiatr Res. 2015;60:56–64. doi: 10.1016/j.jpsychires.2014.10.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Hebebrand J, Exner C, Hebebrand K, Holtkamp C, Casper RC, Remschmidt H, et al. Hyperactivity in patients with anorexia nervosa and in semistarved rats: Evidence for a pivotal role of hypoleptinemia. Physiol Behav. 2003;79:25–37. doi: 10.1016/s0031-9384(03)00102-1. [DOI] [PubMed] [Google Scholar]
- 127.Diaz-Cintra S, Garcia-Ruiz M, Corkidi G, Cintra L. Effects of prenatal malnutrition and postnatal nutritional rehabilitation on CA3 hippocampal pyramidal cells in rats of four ages. Brain Res. 1994;662:117–126. doi: 10.1016/0006-8993(94)90803-6. [DOI] [PubMed] [Google Scholar]
- 128.Benítez-Bribiesca L, De la Rosa-Alvarez I, Mansilla-Olivares A. Dendritic spine pathology in infants with severe protein-calorie malnutrition. Pediatrics. 1999;104:e21. doi: 10.1542/peds.104.2.e21. [DOI] [PubMed] [Google Scholar]
- 129.Lerch JP, Yiu AP, Martinez-Canabal A, Pekar T, Bohbot VD, Frankland PW, et al. Maze training in mice induces MRI-detectable brain shape changes specific to the type of learning. Neuroimage. 2011;54:2086–2095. doi: 10.1016/j.neuroimage.2010.09.086. [DOI] [PubMed] [Google Scholar]
- 130.Hall JF, Smith K, Schnitzer SB, Hanford PV. Elevation of activity level in the rat following transition from ad libitum to restricted feeding. J Comp Physiol Psychol. 1953;46:429–433. doi: 10.1037/h0062565. [DOI] [PubMed] [Google Scholar]
- 131.Routtenberg A, Kuznesof AW. Self-starvation of rats living in activity wheels on a restricted feeding schedule. J Comp Physiol Psychol. 1967;64:414–421. doi: 10.1037/h0025205. [DOI] [PubMed] [Google Scholar]
- 132.Frintrop L, Liesbrock J, Paulukat L, Johann S, Kas MJ, Tolba R, et al. Reduced astrocyte density underlying brain volume reduction in activity-based anorexia rats [published online ahead of print Jan 30] World J Biol Psychiatry. 2017 doi: 10.1080/15622975.2016.1273552. [DOI] [PubMed] [Google Scholar]
- 133.Reyes-Haro D, Labrada-Moncada FE, Miledi R, Martínez-Torres A. Dehydration-induced anorexia reduces astrocyte density in the rat corpus callosum. Neural Plast. 2015;2015:474917. doi: 10.1155/2015/474917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Elsayed M, Magistretti PJ. A new outlook on mental illnesses: Glial involvement beyond the glue. Front Cell Neurosci. 2015;9:468. doi: 10.3389/fncel.2015.00468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Sporns O, Tononi G, Kötter R. The human connectome: A structural description of the human brain. PLoS Comput Biol. 2005;1:e42. doi: 10.1371/journal.pcbi.0010042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Fornito A, Zalesky A, Breakspear M. The connectomics of brain disorders. Nat Rev Neurosci. 2015;16:159–172. doi: 10.1038/nrn3901. [DOI] [PubMed] [Google Scholar]
- 137.Tofts PL. Quantitative MRI of the Brain: Measuring Changes. Hoboken, NJ: Wiley; 2005. [Google Scholar]
- 138.Liu S, Cai W, Liu S, Zhang F, Fulham M, Feng D, et al. Multimodal neuroimaging computing: A review of the applications in neuropsychiatric disorders. Brain Inform. 2015;2:167–180. doi: 10.1007/s40708-015-0019-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Alexander-Bloch A, Giedd JN, Bullmore E. Imaging structural co-variance between human brain regions. Nat Rev Neurosci. 2013;14:322–336. doi: 10.1038/nrn3465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Geisler D, Borchardt V, Lord AR, Boehm I, Ritschel F, Zwipp J, et al. Abnormal functional global and local brain connectivity in female patients with anorexia nervosa. J Psychiatry Neurosci. 2016;41:6–15. doi: 10.1503/jpn.140310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Ehrlich S, Lord AR, Geisler D, Borchardt V, Boehm I, Seidel M, et al. Reduced functional connectivity in the thalamo-insular subnetwork in patients with acute anorexia nervosa. Hum Brain Mapp. 2015;36:1772–1781. doi: 10.1002/hbm.22736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Lutti A, Dick F, Sereno MI, Weiskopf N. Using high-resolution quantitative mapping of R1 as an index of cortical myelination. Neuroimage. 2014;93(part 2):176–188. doi: 10.1016/j.neuroimage.2013.06.005. [DOI] [PubMed] [Google Scholar]
- 143.Sui J, Adali T, Yu Q, Chen J, Calhoun VD. A review of multivariate methods for multimodal fusion of brain imaging data. J Neurosci Methods. 2012;204:68–81. doi: 10.1016/j.jneumeth.2011.10.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Godier LR, Park RJ. Compulsivity in anorexia nervosa: A transdiagnostic concept. Front Psychol. 2014;5:778. doi: 10.3389/fpsyg.2014.00778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Thompson PM, Stein JL, Medland SE, Hibar DP, Vasquez AA, Renteria ME, et al. The ENIGMA Consortium: Large-scale collaborative analyses of neuroimaging and genetic data. Brain Imaging Behav. 2014;8:153–182. doi: 10.1007/s11682-013-9269-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Thompson PM, Andreassen OA, Arias-Vasquez A, Bearden CE, Boedhoe PS, Brouwer RM, et al. ENIGMA and the individual: Predicting factors that affect the brain in 35 countries worldwide. Neuroimage. 2017;145:389–408. doi: 10.1016/j.neuroimage.2015.11.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Lee PH, Baker JT, Holmes AJ, Jahanshad N, Ge T, Jung JY, et al. Partitioning heritability analysis reveals a shared genetic basis of brain anatomy and schizophrenia. Mol Psychiatry. 2016;21:1680–1689. doi: 10.1038/mp.2016.164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Duncan L, Yilmaz Z, Gaspar H, Walters R, Goldstein J, Anttila V, et al. Significant locus and metabolic genetic correlations revealed in genome-wide association study of anorexia nervosa. Am J Psychiatry. 2017;174:850–858. doi: 10.1176/appi.ajp.2017.16121402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Bulik-Sullivan B, Finucane HK, Anttila V, Gusev A, Day FR, Loh PR, et al. An atlas of genetic correlations across human diseases and traits. Nat Genet. 2015;47:1236–1241. doi: 10.1038/ng.3406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Bulik CM, Kleiman SC, Yilmaz Z. Genetic epidemiology of eating disorders. Curr Opin Psychiatry. 2016;29:383–388. doi: 10.1097/YCO.0000000000000275. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.