

Dear Editor,
Dysregulation of hepatic lipid and glucose production results in obesity, diabetes and nonalcoholic fatty liver disease. De novo lipogenesis and gluconeogenesis in the liver contribute, at least in part, to the dynamic homeostasis of lipid and glucose levels (Rui, 2014; Han and Wang, 2017). These processes are regulated at the transcriptional level by different transcription factors in response to various environmental cues, including hormones, nutrition and stress. Regulators of lipogenesis include the sterol regulatory element-binding proteins (SREBPs), which are members of the basic helix-loop-helix leucine zipper transcription factor family (Goldstein et al., 2006). There are three closely related SREBPs, SREBP1a, SREBP1c and SREBP2, among which SREBP1c and SREBP2 are the major isoforms expressed in the liver. SREBP1c mainly regulates fatty acid synthesis, while SREBP2 controls cholesterol production (Goldstein et al., 2006). SREBPs are synthesized as inactive precursors bound to the endoplasmic reticulum (ER), where they associate with the sterol cleavage activating protein (SCAP) (Goldstein et al., 2006). Upon sensing sterol demand, the SREBP/SCAP complex buds from the ER in a COPII-dependent manner and is transported to the Golgi, where SREBP processing occurs. The mature SREBP then translocates to the nucleus and activates the transcription of lipid biosynthesis genes. In the presence of excess sterol levels, the SREBP/SCAP complex binds to insulin-induced gene (INSIG) and is restricted to the ER (Goldstein et al., 2006). Thus, SREBP processing, shuttling of SREBP to the nucleus and transcriptional control of genes involved in cholesterol and fatty acid synthesis are tightly regulated in response to nutrition conditions.
SREBP1c processing is regulated not only by nutrient status but also by hormonal signaling during feeding and fasting (Horton et al., 1998; Li et al., 2010). These features distinguish it from SREBP2, processing of which is regulated by sterol levels. During feeding, insulin signaling promotes SREBP1c activity through two parallel pathways, one dependent on mTOR and the other dependent on Insig2a (Yecies et al., 2011; Han and Wang, 2017). The Insig2 gene produces two alternatively spliced transcripts, Insig2a and Insig2b, which have different 5′-UTRs but share the same coding region (Yabe et al., 2003). Previous studies have shown that expression of Insig2a, the predominant Insig2 isoform in liver, is enhanced during fasting and attenuated during feeding (Yabe et al., 2003). Furthermore, insulin signaling decreases Insig2a expression with concomitant enhancement of SREBP1c activity (Yabe et al., 2003; Yellaturu et al., 2009). Together, these results suggest that Insig2a is a potent regulator of SREBP1c activity during both feeding and fasting in the liver. Although both fasting and/or glucagon signaling can attenuate SREBP1c activity (Horton et al., 1998; Yabe et al., 2003), the molecular mechanism linking fasting to Insig2a expression is unclear.
In contrast to SREBP1c-mediated hepatic lipogenesis, gluconeogenesis is activated during fasting and shut down during feeding. Gluconeogenesis is partially regulated by cAMP response element-binding protein (CREB) and CREB regulated transcription coactivators (CRTCs) (Altarejos and Montminy, 2011). CRTC2, which is highly expressed in the liver, is dephosphorylated and shuttled to the nucleus during fasting, where it binds to CREB and induces gluconeogenic gene expression (Altarejos and Montminy, 2011). During feeding, CRTC2 is phosphorylated and sequestered in the cytoplasm through phosphorylation-dependent interactions with 14-3-3 proteins (Altarejos and Montminy, 2011). Our previous results showed that CRTC2 mediates mTOR-controlled SREBP1c processing and lipogenesis during feeding (Han et al., 2015). However, it is unclear whether CRTC2 affects SREBP1c processing during fasting.
To investigate the effect of CRTC2 on SREBP1c processing during fasting, we fasted mice for 12 h and evaluated SREBP1c processing. We found that higher levels of mature SREBP1c accumulate in Crtc2−/− mice than in Crtc2+/+ mice following fasting (Fig. 1A). Expression of two SREBP1c-targeted lipogenic genes (Fasn, Scd1) was also elevated in Crtc2−/− mice as judged by real-time quantitative PCR (qPCR) (Fig. 1B). In contrast to SREBP1c, levels of SCAP are comparable in Crtc2+/+ and Crtc2−/− mice under fasted conditions (Fig. 1A). In addition to SCAP, INSIG family proteins, including INSIG1 and INSIG2, also play an important role in controlling SREBP1c processing (Yabe et al., 2002; Yabe et al., 2003). Therefore, we evaluated the hepatic expression of Insig1, Insig2a and Insig2b in Crtc2−/− mice. Insig2a, but not Insig1 or Insig2b, was dramatically decreased during fasting in Crtc2−/− mice, which is similar to attenuated expression of gluconeogenic genes (Pck1 and G6pc) (Figs. 1B and S1). In addition, fasting-induced effect on SREBP1c processing and Insig2a expression was blocked by H89, a PKA inhibitor (Fig. S2). Together, these results indicate that CRTC2 inhibits SREBP1c activity during fasting.
Figure 1.
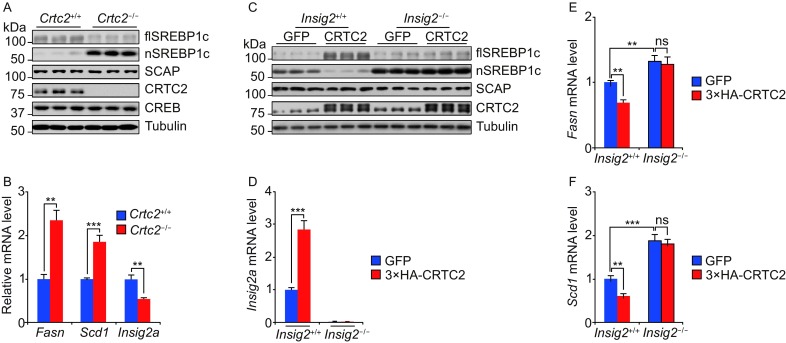
CRTC2 inhibits SREBP1c processing via INSIG2 during fasting. (A and B) Immunoblots showing hepatic amounts of full-length, inactive SREBP1c (flSREBP1c) and cleaved, active SREBP1c (nSREBP1c) in liver extracts (A) and qPCR results showing expression of lipogenic genes (Fasn, Scd1 and Insig2a) (B) in 12 h fasted Crtc2+/+ and Crtc2−/− mice. (C–F) Effect of 3×HA-CRTC2 overexpression on SREBP1c maturation (C), Insig2a expression (D), Fasn expression (E) and Scd1 expression (F) in 12 h fasted Insig2+/+ and Insig2−/− mice. ns, no significant statistical difference. Data are shown as mean ± s.e.m. **P < 0.01, ***P < 0.001, n = 8
We next asked whether CRTC2 modulates SREBP1c processing via INSIG2. Adenoviral expression of HA-tagged CRTC2 (3× HA-CRTC2) enhanced Insig2a expression and thus attenuated SREBP1c processing in wild-type mice during fasting (Fig. 1C–F). The inhibitory effect of CRTC2 on SREBP1c processing was abolished in Insig2−/− mice (Fig. 1C–F). These results demonstrate that CRTC2 regulates SREBP1c processing in an INSIG2-dependent manner during fasting.
Considering that Insig2a is a predominant isoform of Insig2 in the liver (Yabe et al., 2003) and CRTC2-dependent transcriptional effect on Insig2a expression, we tested whether the CRTC2/CREB complex directly regulates Insig2a expression. Overexpression of wild-type (WT) and constitutively active (CRTC2/AA) (Han et al., 2015) CRTC2 in the liver increased Insig2a expression; knockdown of Crtc2 by short hairpin RNA decreased its expression (Fig. 2A and 2B), which is consistent with the results from Crtc2 knockout mice (Fig. 1B). In addition, overexpression of ACREB, a dominant-negative form of CREB (Herzig et al., 2003), decreased Insig2a expression (Fig. 2C). Together, these results suggest that the CRTC2/CREB complex, as a whole, controls Insig2a expression.
Figure 2.
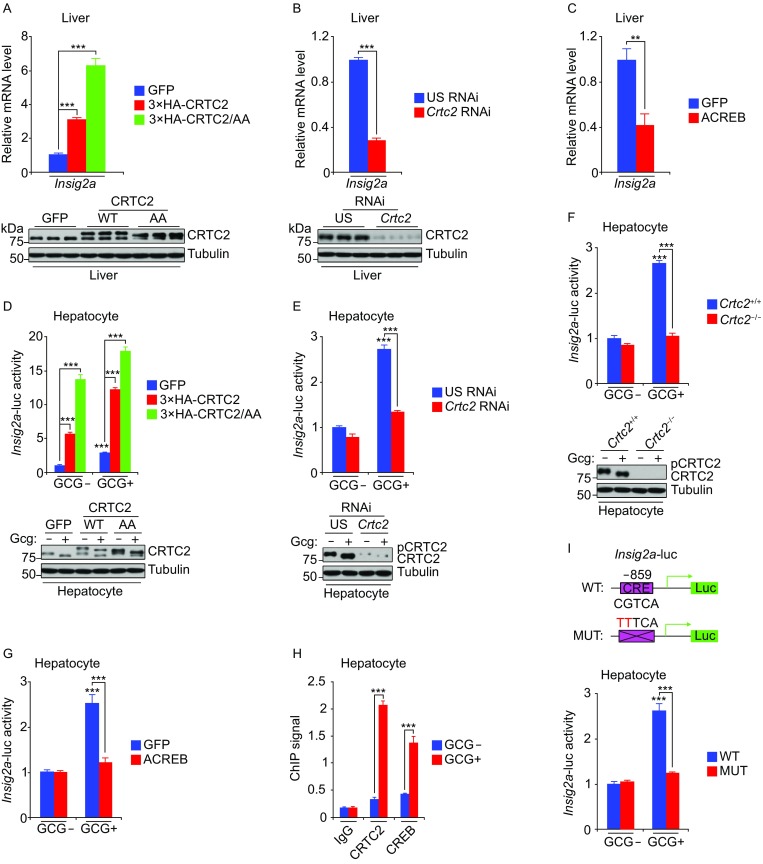
Insig2a is transcriptionally regulated by the CRTC2/CREB complex. (A–C) Effect of wild-type (WT) 3×HA-CRTC2 and its nuclear-localized mutant (FLAG-CRTC2/AA, CRTC2 with double Ala mutations at Ser171 and Ser275) (A), Crtc2 knockdown (B) or ACREB (C) on Insig2a expression in 12 h fasted mice. Bottom panels: Immunoblots showing the relative CRTC2 level. (D–G), Effect of 3×HA-CRTC2 and 3×HA-CRTC2/AA (D), CRTC2 deficiency by knockdown (E) or knockout (F), and ACREB (G) on Insig2a-luc activity in mouse primary hepatocytes. Bottom panels: Immunoblots showing the relative CRTC2 level. (H) Chromatin immunoprecipitation showing the occupancy of CRTC2 and CREB on the Insig2a promoter. (I) Top panel: Location of the wild-type (WT) CRE half-site and its mutant (MUT) within the mouse Insig2a promoter. Bottom panel: Relative luciferase activity with the wild-type or mutant Insig2a promoters in the presence or absence of glucagon. GCG, glucagon. US, unspecific. Data are shown as mean ± s.e.m. **P < 0.01, ***P < 0.001, n = 8
To further demonstrate whether the CRTC2/CREB complex autonomously regulates Insig2a, we used reporter assays in mouse primary hepatocytes. Overexpression of CRTC2 and CRTC2/AA enhanced Insig2a-luc activity following glucagon (Gcg) treatment, while Crtc2 deficiency by knockdown or knockout attenuated Insig2a-luc activity (Fig. 2D–F). As shown in Figure 2G, overexpression of ACREB decreased Insig2a-luc activity in hepatocytes, consistent with the results in liver (Fig. 2C). These results show that the CRTC2/CREB complex autonomously controls Insig2a expression.
Having obtained evidence for a regulatory effect of the CRTC2/CREB complex on Insig2a, we performed chromatin immunoprecipitation (ChIP) studies to determine whether Insig2a is a direct target of CRTC2/CREB. Supporting this notion, glucagon stimulation dramatically increased both CRTC2 and CREB occupancy on the Insig2a promoter (Fig. 2H). CREB regulates gene expression by binding to palindromic (TGACGTCA) or to half-site (TGACG/CGTCA) cAMP-responsive elements (CREs) positioned within 1 kb of the transcriptional start site (Altarejos and Montminy, 2011). When we mutated a CRE half-site located at position − 859 of the Insig2a promoter, the Insig2a-luc reporter lost its ability to respond to glucagon (Fig. 2I), indicating that this element is critical for glucagon-induced transcriptional activation of Insig2a. Taken together, these results suggest that fasting activates CRTC2/CREB-controlled Insig2a expression, thereby attenuating SREBP1c processing.
It is well known that fasting or glucagon attenuates SREBP1c processing (Horton et al., 1998; Shimomura et al., 2000; Yabe et al., 2003), but the mechanism is unclear. Our results showed that the CRTC2/CREB complex controls Insig2a expression and thus attenuates SREBP1c processing during fasting (Fig. S3). This highlights the important role of the CRTC2/CREB complex in SREBP1c maturation during fasting. Meanwhile, our results fill the gap in our understanding of how glucagon or fasting signaling modulates SREBP1c processing.
Footnotes
We thank Drs. Marc Montminy and Wei-min Tong for providing reagents. This work was supported by the National Natural Science Foundation of China (Grant No. 31500956).
All institutional and national guidelines for the care and use of laboratory animals were followed. Yuanyuan Zhang, Yi Liu, Liqun Chen, Yiguo Wang and Jinbo Han declare that they have no conflict of interest.
Yuanyuan Zhang and Jinbo Han designed the study and analyzed the data. Yuanyuan Zhang, Yi Liu and Liqun Chen performed the experiments. Jinbo Han wrote the paper. All authors reviewed and commented on the manuscript.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Footnotes
Electronic supplementary material
The online version of this article (10.1007/s13238-018-0538-3) contains supplementary material, which is available to authorized users.
References
- Altarejos JY, Montminy M. CREB and the CRTC co-activators: sensors for hormonal and metabolic signals. Nat Rev Mol Cell Biol. 2011;12:141–151. doi: 10.1038/nrm3072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen L, Wang K, Long A, Jia L, Zhang Y, Deng H, Li Y, Han J, Wang Y. Fasting-induced hormonal regulation of lysosomal function. Cell Res. 2017;27:748. doi: 10.1038/cr.2017.45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldstein JL, DeBose-Boyd RA, Brown MS. Protein sensors for membrane sterols. Cell. 2006;124:35–46. doi: 10.1016/j.cell.2005.12.022. [DOI] [PubMed] [Google Scholar]
- Han J, Wang Y. mTORC1 signaling in hepatic lipid metabolism. Protein Cell. 2017;9:145–151. doi: 10.1007/s13238-017-0409-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han J, Li E, Chen L, Zhang Y, Wei F, Liu J, Deng H, Wang Y. The CREB coactivator CRTC2 controls hepatic lipid metabolism by regulating SREBP1. Nature. 2015;524:243–246. doi: 10.1038/nature14557. [DOI] [PubMed] [Google Scholar]
- Herzig S, Hedrick S, Morantte I, Koo SH, Galimi F, Montminy M. CREB controls hepatic lipid metabolism through nuclear hormone receptor PPAR-gamma. Nature. 2003;426:190–193. doi: 10.1038/nature02110. [DOI] [PubMed] [Google Scholar]
- Horton JD, Bashmakov Y, Shimomura I, Shimano H. Regulation of sterol regulatory element binding proteins in livers of fasted and refed mice. Proc Natl Acad Sci USA. 1998;95:5987–5992. doi: 10.1073/pnas.95.11.5987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S, Brown MS, Goldstein JL. Bifurcation of insulin signaling pathway in rat liver: mTORC1 required for stimulation of lipogenesis, but not inhibition of gluconeogenesis. Proc Natl Acad Sci USA. 2010;107:3441–3446. doi: 10.1073/pnas.0914798107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rui L. Energy metabolism in the liver. Compr Physiol. 2014;4:177–197. doi: 10.1002/cphy.c130024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimomura I, Matsuda M, Hammer RE, Bashmakov Y, Brown MS, Goldstein JL. Decreased IRS-2 and increased SREBP-1c lead to mixed insulin resistance and sensitivity in livers of lipodystrophic and ob/ob mice. Mol Cell. 2000;6:77–86. doi: 10.1016/S1097-2765(05)00010-9. [DOI] [PubMed] [Google Scholar]
- Wang Y, Inoue H, Ravnskjaer K, Viste K, Miller N, Liu Y, Hedrick S, Vera L, Montminy M. Targeted disruption of the CREB coactivator Crtc2 increases insulin sensitivity. Proc Natl Acad Sci USA. 2010;107:3087–3092. doi: 10.1073/pnas.0914897107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yabe D, Brown MS, Goldstein JL. Insig-2, a second endoplasmic reticulum protein that binds SCAP and blocks export of sterol regulatory element-binding proteins. Proc Natl Acad Sci USA. 2002;99:12753–12758. doi: 10.1073/pnas.162488899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yabe D, Komuro R, Liang G, Goldstein JL, Brown MS. Liver-specific mRNA for Insig-2 down-regulated by insulin: implications for fatty acid synthesis. Proc Natl Acad Sci USA. 2003;100:3155–3160. doi: 10.1073/pnas.0130116100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yecies JL, Zhang HH, Menon S, Liu S, Yecies D, Lipovsky AI, Gorgun C, Kwiatkowski DJ, Hotamisligil GS, Lee CH, et al. Akt stimulates hepatic SREBP1c and lipogenesis through parallel mTORC1-dependent and independent pathways. Cell Metab. 2011;14:21–32. doi: 10.1016/j.cmet.2011.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yellaturu CR, Deng X, Park EA, Raghow R, Elam MB. Insulin enhances the biogenesis of nuclear sterol regulatory element-binding protein (SREBP)-1c by posttranscriptional down-regulation of Insig-2A and its dissociation from SREBP cleavage-activating protein (SCAP).SREBP-1c complex. J Biol Chem. 2009;284:31726–31734. doi: 10.1074/jbc.M109.050914. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.