

Abstract
Rodent incisors grow permanently and the homeostasis of enamel production is maintained by a continuous supply of epithelial progenitors from putative stem cells in the cervical loop. We herein report that Runx1 regulates the Lgr5-expressing epithelial stem cells and their subsequent continuous differentiation into ameloblasts. Mice deficient in epithelial Runx1 demonstrate remarkable shortening of the incisors with underdevelopment of the cervical loop and enamel defects. In this mutant cervical loop, the proliferation of the dental epithelium was significantly disturbed and the expression of Lgr5 and enamel matrix proteins was remarkably downregulated. Interestingly, the expression of Socs3, an inhibitor of Stat3 signaling, was upregulated and Stat3 phosphorylation was suppressed specifically in the mutant cervical loop. The expression of Lgr5 and the enamel matrix protein in the wild-type incisor germs is disturbed by pharmaceutical Stat3 inhibition in vitro., of. Conversely, pharmaceutical activation of Stat3 rescues the defective phenotypes of the Runx1 mutant with upregulated Lgr5 and enamel matrix protein genes. The present results provide the first evidence of the role of Runx1 regulates the Lgr5-expressing epithelial stem cells and differentiation of ameloblast progenitors in the developing incisors. Our study also demonstrates that Stat3 modulates the Runx1-Lgr5 axis in the cervical loop.
Introduction
The rodent incisor represents a special type of tooth, as it grows continuously throughout the lifetime of the animal. Its regenerative capacity implies the presence of a stem cell pool1. The cervical loop, which is located at the proximal end of the incisor, has been shown to contain the epithelial stem cell niche that houses the progenitors of ameloblasts, which secrete enamel matrix at the labial surface of the incisors2,3. Many signaling molecules and growth factors have been implicated in the mediation of these interactions4, and several molecules are specifically involved in the maintenance of the continuous regeneration of the incisors5.
The Runx family genes are important transcriptional regulators in the embryonic development of various tissues and the pathogenesis of various diseases. Among the Runx family genes, Runx1 is recognized as a human hematopoietic stem cell factor6. In addition, Runx1 is also important for skin stem cells as well as skin cancer development. Furthermore, the epithelial-targeted deletion of Runx1 affects the hair shaft structure and the homeostasis of hair follicle stem cells7,8. On the other hand, our previous study demonstrated that the cervical loop did not fully develop in the incisors of mice deficient in epithelial Cbfb, which resulted in short incisors9. Cbfb is a non-DNA binding regulatory subunit and forms a heterodimer with the Runx proteins, as the complex works as a transcription factor10, However, the specific alpha unit of the RUNX protein that is required in the development of the incisors has remained unclear. In addition, the molecular mechanism underlying the regulation of stem cells in the incisors by Runx signaling is still largely unknown.
In mammals, JAK/STAT signaling is essential for a wide array of cytokines and growth factors11,12. Upon activation, phosphorylated Stats dimerize and translocate to the nucleus where they modulate the expression of target genes. Among the various functions of the Stat family, Stat3 is involved in the regulation of the self-renewal of embryonic stem cells, cancer stem cells, hematopoietic stem cells and the stem cells in hair follicles. Indeed, Lif activates Stat3 activation in ES cells13. In hair growth, the depletion of stat3 results in decreases in the stem cell colony and prolongs the hair regeneration cycle14. In hair development, Stat3 activity is regulated by Runx1 and the depletion of Runx1 leads to the inhibition of Stat3 phosphorylation in the bulge of the hair follicle and results in hair cycle dysregulation7,15. In addition, the overexpression of Runx1 leads to Stat3 activation and is necessary for the regulation of cellular proliferation and tumorigenesis in the skin16. STAT3 is involved in human breast cancer, in which high STAT3 levels are correlated with a poorer survival17. However, the possible involvement of Stat3 in the stem cell homeostasis in the growing incisors has not been investigated.
Lgr5 is a general stem cell marker and has been identified as a Wnt target genes in the intestinal stem cell niche18,19. In the growing incisors, the expression of Lgr5 is localized in the stem cell compartment in the cervical loop epithelium20,21. However, canonical Wnt signaling is exclusively evident in the mesenchymal tissue of the reporter mice, but not in the cervical loop epithelium22. On the other hand, Fgf10/Fgfr2b signaling plays essential roles in the regulation of the cervical loop stem cells1,20,21,23. In this signaling axis, the inhibition of FGFR or its downstream signal transduction pathways was shown to diminish the Lgr5-expressing cells in the cervical loop24. However, the molecular mechanism through which the Lgr5 expression is regulated in the cervical loop remains to be elucidated.
We herein report that Runx1 is involved in the regulation of epithelial stem cells and their subsequent differentiation into ameloblasts in the rodent incisors, using mice with epithelial-specific deletion of Runx1. We show that Runx1 deficiency results in the marked shortening of the incisors with the underdevelopment of the cervical loop. Indeed, the expression levels of Lgr5 and Sox2 (general stem cell markers) are significantly downregulated due to Runx1 deficiency. Runx1 mutant mice also show enamel defects with the significant downregulation of amelogenin and ameloblastin (ameloblast differentiation markers). We show that Stat3 phosphorylation of the mutant incisors is disturbed by Runx1 deficiency, specifically in the cervical loop epithelium. Indeed, the application of Stat3 inhibitors remarkably suppressed the expression of Lgr5 and enamel matrix protein of explanted incisors in in vitro culture. Conversely, treatment with Stat3 activator partially rescued the suppressed Lgr5 and Amelogenin expression in Runx1 mutant incisors in culture. Thus, we demonstrate that Runx1 is involved in the regulation of Lgr5-positive epithelial stem cells and ameloblast progenitors, at least in part, through the regulation of Stat3 phosphorylation in the cervical loop of the growing incisors.
Results
The expression of Runxs mRNA in the growing incisors
The previous study reported that Runx1 mRNA is expressed in the dental epithelium25. The present in situ hybridization analysis revealed that Runx1 was present in the basal epithelial cells (inner enamel epithelium) and the outer enamel epithelium in the cervical loop at P0 (Fig. 1A). RT-PCR analysis demonstrated that Runx1 expression was not detected in the dental mesenchyme. Runx2 mRNA demonstrated similar expression pattern both in the cervical loop and in the inner enamel epithelium (Fig. 1A) and Runx2 transcripts were also present in the mesenchyme, which was supported by PCR analysis (Fig. 1A). Runx3 and Cbfb mRNA transcripts were also detected both in the dental epithelium and mesenchyme, which was supported by PCR analysis (Fig. 1A).
Figure 1.

The incisor phenotype of K14-Cre/Runx1fl/fl mice. (A) Expression of Runx1, Runx2, Runx3 and Cbfb in sagittal sections of P0 incisors and gene expression was confirmed by RT-PCR from dissected epithelium and mesenchyme. Runx1 transcripts were detected in the dental epithelium. In the cervical loop, Runx1 was expressed in the basal epithelial cells (inner enamel epithelium) and the outer enamel epithelium in the cervical loop. Scale bar: 500 μm. (B) The Runx1 mutant incisors demonstrated a column shape and lost their transparency, although the control incisors developed fine edged shape. In the micro-CT analysis, the incisors of the Runx1 mutant mice had an abnormal shape, extreme shortening and enamel defects. On the other hand, significant differences were not detected in the molar root length or morphology. (C) HE staining of incisor sagittal sections from P0 Runx1 mutant mice confirmed the shortening of the Runx1 mutant incisors. At higher magnification, hypoplasia of the cervical loop (arrow) was evident and ameloblast precursors lost their polarity and had a flattened shape in Runx1 mutants (arrowhead). Scale bar: 100 μm. (D) HE staining of incisor frontal sections from P28 Runx1 mutant mice confirmed the lack of enamel matrix formation. At higher magnification, enamel matrix was not evident between the dentin and the periodontal ligament (arrow) in the upper incisors of Runx1 mutants. (E) Double immunostaining for Ki67 (green) and K14 (red) revealed that the epithelium in the cervical loop of the Runx1 mutant was significantly less proliferative (arrow). Nuclei were counterstained with DAPI (blue). Scale bar: 200 μm. (F) TUNEL staining revealed that the number of TUNEL-positive cells was significantly reduced at the cervical loop of the Runx1 mutant incisors (arrowhead). cl, cervical loop; pu, dental pulp; en, enamel; am, ameloblasts; od, odontoblasts. Scale bar: 200 μm.
The incisor phenotypes in Runx1 mutants
To analyze the functional role of Runx1 in the dental epithelium of the growing incisors, we examined the incisal phenotypes of conditional knockout (K14-Cre/Runx1fl/fl) mice, in which Runx1 expression was disrupted in the epithelium using Cre recombination26. The efficiency of the K14-Cre recombination of our mice was previously confirmed using Rosa26R reporter mice and X-gal staining9.
At P28, Runx1 mutants demonstrated column-shaped incisors with a loss of transparency, while the control mice developed fine-edged incisors (Fig. 1B). A micro-CT analysis revealed that the Runx1 mutant mice had abnormally shaped and extremely short incisors in comparison to the control mice (Fig. 1B). The labial surface of the mutant incisors was lacking enamel (Fig. 1B). On the other hand, there were no evident changes in the morphology of the mutant molars (Fig. 1B).
A histological analysis confirmed the shortening of the Runx1 mutant incisors at P0 (Fig. 1C). Under higher magnification, the mutant cervical loop demonstrated hypoplasia with a remarkably reduced stellate reticulum region. In wild-type incisors, ameloblast precursors started to be polarized and their nuclei aligned at the apical side of the cells; such polarization was not observed in the Runx1 mutant incisors (Fig. 1C). Enamel matrix was not evident on the labial surface of the Runx1 mutant incisors at E28 (Fig. 1D).
Double immunostaining of Ki67 and Keratin 14 (K14) revealed numerous proliferative cells in the epithelial layer in the cervical loop of control incisors. In contrast, the number of Ki67-immunoreactive proliferating epithelial cells was markedly reduced in the Runx1 mutant incisors (Fig. 1E), indicating that the proliferative activity in the cervical loop was markedly disturbed by Runx1 deficiency.
It is established that apoptotic cells are localized at the stellate reticulum of the cervical loop of the growing incisors23. TUNEL staining revealed that the number of apoptotic cells in the cervical loop was significantly decreased by Runx1 deficiency (Fig. 1F).
Downstream targets of Runx1 signaling in the incisors
To investigate how Runx1 deficiency leads to the defective incisor growth, we evaluated the possible downstream target genes by in situ hybridization and a qPCR. It is established that Lgr5 and Sox2 are specific markers of the epithelial stem cells in the cervical loop in the continuously growing mouse incisor2,27. In the present study, in situ hybridization and qPCR revealed that the expression levels of both Lgr5 and Sox2 were downregulated in the Runx1 mutant incisors and the downregulation of Lgr5 was more pronounced than that of Sox2 (Fig. 2A). The present in situ hybridization analysis and qPCR also demonstrated the significant downregulation of Amelogenin (Amel), Enamelin (Enam), Ameloblastin (Ambn) in the Runx1 mutant incisors (Fig. 2B). Mmp20 expression was not deviated significantly (Fig. 2B).
Figure 2.

The downstream target molecules of Runx1 signaling in the growing incisors. (A) In situ hybridization analyses demonstrated that the expression of Lgr5 and Sox2 (general stem cell markers) was significantly downregulated in the Runx1 mutants. A qPCR confirmed the downregulation of the Lgr5 and Sox2 expression in the mutants. *p < 0.05. Scale bar: 100 μm. (B) An in situ hybridization analysis demonstrated that the expression of Amelogenin (Amel), an enamel matrix protein, was significantly downregulated in the Runx1 mutants. A qPCR also confirmed the significant downregulation of Amel, Amelogenin (Amel), Enamelin (Enam), Ameloblastin (Ambn). The expression of Mmp20 was not significantly affected. *p < 0.05. Scale bar: 500 μm. (C) A qPCR A demonstrated that the Bcl-2 expression was upregulated in the Runx1 mutant cervical loop. Bax was not affected. *p < 0.05. cl, cervical loop.
In order to evaluate the possible cause of decreases in apoptotic cells at the labial side of the cervical loop, we evaluated the Bcl-2 and Bax expression. qPCR of the cervical loop showed that anti-apoptotic molecule Bcl-2 was upregulated in the Runx1 mutant cervical loop (Fig. 2C). Indeed, such upregulation of Bcl-2 due to Runx1 deficiency has also been observed in hematopoietic stem cells28
Stat3 phosphorylation in Runx1 mutant incisors
Runx1 acts as a tumor promoter in the formation and maintenance of mouse skin cancer through its promotion of Stat3 activation15,29 and Runx1 stimulates the Stat pathway through the repression of suppressor of cytokine signaling 3 (Socs3), a major negative feedback regulator of Stat3-activating cytokines13. This Stat3/Socs3 pathway plays a key role in promoting hair cell regeneration through stem cell activation, cell division, and differentiation in zebrafish30. Hence, we assumed that Runx1 deficiency might affect Stat3 signaling in the pathogenesis of defective phenotypes in the Runx1 mutant incisor.
Our in situ hybridization analysis and qPCR showed that Stat3 was expressed in the cervical region and was not affected by Runx1 deficiency (Fig. 3A).
Figure 3.

(A) An in situ hybridization analysis demonstrated that the mRNA expression of Stat3 was evident in the cervical loop and the alveolar bone in the Runx1 mutant and control mice. A qPCR demonstrated that the Stat3 expression was not affected significantly by Runx1 deficiency. Scale bar: 200 μm. (B) Stat3 immunoreactivity was widely distributed in both the epithelium and the mesenchyme of the incisors and the surrounding bone. Stat3 immunoreactivity was not affected by Runx1 deficiency. In contrast, the pStat3 immunoreactivity of the cervical loop epithelium and the differentiating ameloblast was intense in control mice. In the Runx1 mutants, immunoreactivity to pStat3 was remarkably disturbed at the cervical loop epithelium (arrowhead), but not in the odontoblast layers or on the alveolar bone surface (asterisks). Scale bar: 200 μm. (C) An in situ hybridization analysis demonstrated that the mRNA expression of Socs3 was evident in the cervical loop and upregulated in Runx1 mutants. A qPCR demonstrated that the Socs3 expression was significantly upregulated by Runx1 deficiency. Scale bar: 200 μm. *p < 0.05. cl, cervical loop; pu, dental pulp; ab, alveolar bone.
The Stat3 activity was evaluated based on the state of phosphorylation. Stat3 immunohistochemistry demonstrated that the Stat3 immunoreactivity was widely detected in both the epithelium and the mesenchyme of the incisors and the surrounding bone (Fig. 3B). The spatial distribution of Stat3 immunoreactivity demonstrated similar distribution with regard to the expression of Stat3 mRNA (Fig. 3B). Furthermore, the spatial distribution of Stat3 immunoreactivity was not deviated in Runx mutants, indicating that Runx1 deficiency did not affect Stat3 expression (Fig. 3B).
The distribution of phospho-Stat3 (pStat3) immunoreactivity was similar to that of Stat3 and immunoreactivity to pStat3 was specifically intense in the cervical loop (Fig. 3B). Interestingly, pStat3 immunoreactivity was specifically disturbed at the cervical loop in the Runx1 mutants, whereas pStat3 immunoreactivity on the alveolar bone surface and odontoblasts were not affected (Fig. 3B). These findings indicated that Runx1 could regulate Stat3 activity specifically in the cervical loop epithelium through the modulation of Stat3 phosphorylation.
Our in situ hybridization analyses indicated the mRNA expression of Socs3 in the cervical loop and the significant upregulation of Socs3 expression in the Runx1 mutant (Fig. 3C). Such upregulation of Socs3 was also confirmed by qPCR (Fig. 3C). These findings suggested that Runx1 signaling affects Stat3 phosphorylation through, at least in part, the modulation of the Socs3 expression.
The inhibition of Stat3 phosphorylation in the growing incisors
To evaluate the functional roles of Stat3 signaling, AG490, a selective Jak2/Stat3 inhibitor that prevents Stat3 phosphorylation, and S3I-201, a direct Stat3 inhibitor that blocks Stat3 dimerization and DNA-binding31 was applied to explanted lower incisors at E16.0 and cultured for 6 days. A Western blotting analysis confirmed that the application of the Stat3 inhibitors Ag490 or S3I-201 suppressed immunoreactivity to pStat3, while the Stat3 immunoreactivity was not affected (Fig. 4A). The scanned full blots are presented in Supplemental Fig. S5.
Figure 4.
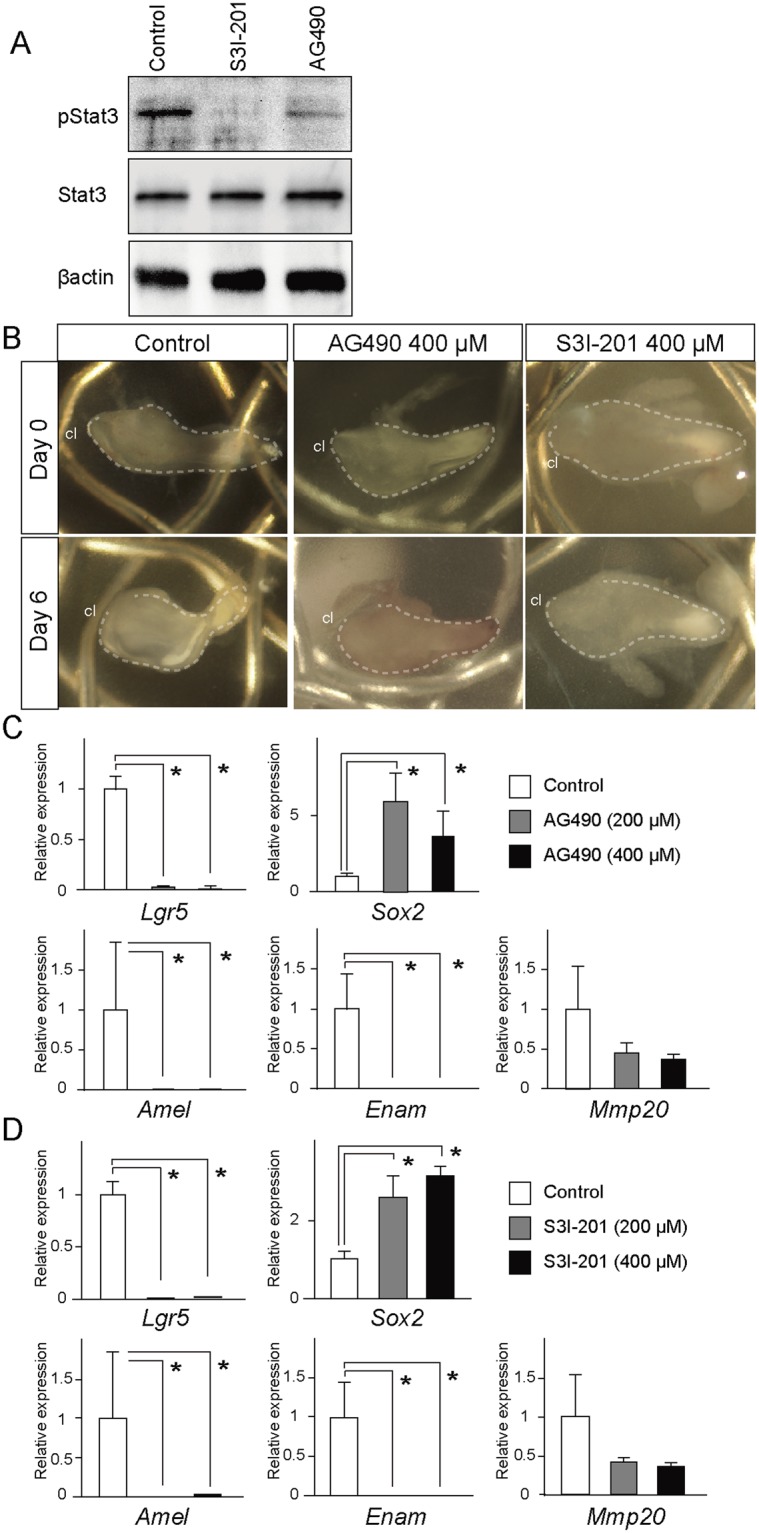
In vitro culture of the incisors and Stat3 inhibitor treatment. (A) A Western blot analysis showed that the application of Stat3 inhibitors of Ag490 or S3I-201 suppressed immunoreactivity to pStat3 but not to Stat3. (B) AG-490 or S3I-201, specific pharmaceutical Stat3 inhibitors, were applied on explanted lower incisors at E16.0 and cultured for 6 days. The size of the incisor explants did not change with Ag490 or S3I-201 treatment. (C) A qPCR analysis of the lower incisors showed that Ag490 treatment resulted in the marked downregulation of the Lgr5 expression, while the expression of Sox2 was upregulated. Ag490 treatment also resulted in the downregulation of the expression of Amelogenin, while AG490 significantly upregulated the Socs3 expression. *p < 0.05. (D) The Treatment of S3I-201 also specifically downregulated the expression levels of Lgr5 and Amelogenin, whereas the Sox2 expression did not deviate and the Socs3 expression was upregulated by S3I-201 treatment. *p < 0.05. cl, cervical loop.
In vitro culture allowed for the continuous monitoring of the explants and the cervical loops in the incisor were translucent and visible during the culture period. After six days of culture, some explants became bent and marked elongation was not observed (Fig. 4B). Furthermore, our histological observation showed that some tissue damages was observed at the tip of the incisal explants (Fig. S2A); however, most of the tissue, including the cervical loop regions, showed no tissue degradation or deterioration of morphology. Ki67 immunohistochemistry further demonstrated that proliferative activity was retained in both the dental epithelium and mesenchyme of the explants after long-term culture (Fig. S2B). These finding indicate that incisor explants remained viable over the course of culture.
qPCR of the lower incisors showed that Ag490 treatment resulted in the marked downregulation of the Lgr5 expression, whereas the expression of Sox2 was significantly upregulated (Fig. 4C). Whole-mount in situ hybridization also confirmed the upregulated Sox2 expression in the cervical loop in the AG490-treated samples (Fig. S2C). Ag490 treatment also resulted in the downregulation of Amelogenin and Enamelin, whereas the expression of Mmp20 was not significantly affected (Fig. 4C). Treatment of S3I-201 also specifically downregulated the expression of Lgr5 and enamel matrix protein genes, whereas the expression of Sox2 was upregulated (Fig. 4D). Treatment of S3I-201 also specifically downregulated the expression of Amelogenin and Enamelin (Fig. 4D). Taken together, these findings suggest that Stat3 activation is involved in the induction of Lgr5 and the expression of enamel matrix protein genes.
Rescue of the defective Runx1 mutant incisor phenotype by Stat3 activation
To confirm the role of Stat3 activation in the induction of the Lgr5 and enamel matrix protein genes under Runx1 signaling, we evaluated the possibility of rescuing the defective Runx1 mutant incisor phenotype, which showed the suppressed expression of Lgr5, by applying a pharmaceutical Stat3 activator to the mutant explants. A previous study demonstrated that ZnCl2 stimulation activates Stat3 phosphorylation and that activated Stat3 promotes embryonic stem cell pluripotency through the upregulation of genes such as Oct4 and Sox232.
The Western blotting analysis of the cervical loop in the present study confirmed that the application of the Stat3 activator ZnCl2 upregulated immunoreactivity to pStat3, while the Stat3 immunoreactivity was not affected (Fig. 5A). The scanned full blots of four samples are presented in Supplemental Fig. S6.
Figure 5.
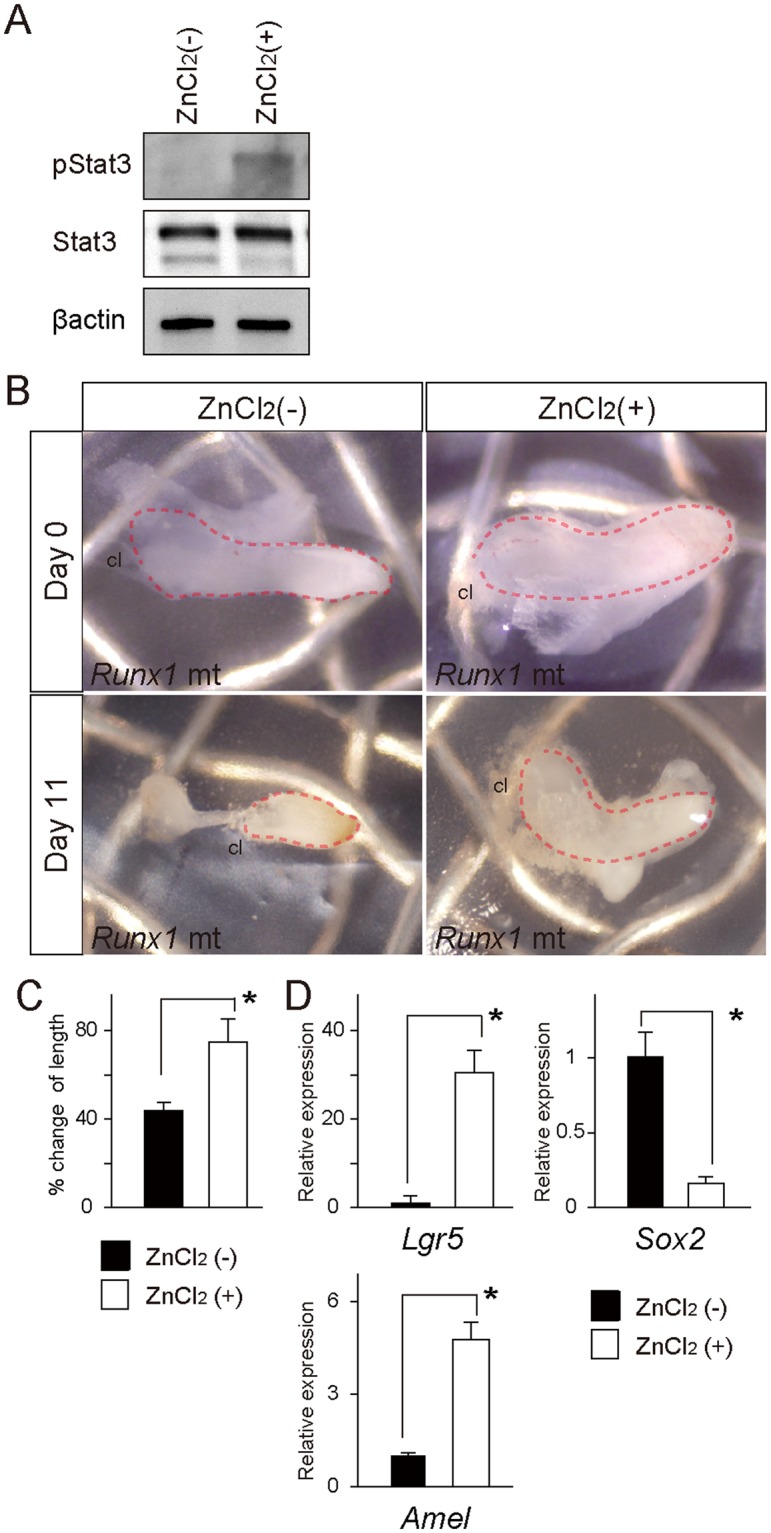
In vitro culture of the Runx1 mutant incisors and ZnCl2 treatment (A) A Western blot analysis showed that the application of Stat3 activator of ZnCl2 upregulated immunoreactivity to pStat3, but not to Stat3. (B) After 11 days of culture, the Runx1 mutant incisor explants became shrunk and bent, while the ZnCl2-treated Runx1 mutants were bigger than the mutants without ZnCl2-treatment. (C) The percent-change in the length of the explants revealed that ZnCl2 treatment rescued the shrinkage of the mutant incisors. *p < 0.05. (D) A qPCR of the Runx1 mutant incisors demonstrated that ZnCl2 treatment resulted in increases in the Lgr5 expression, while expression of Sox2 was downregulated. ZnCl2 treatment also rescued the downregulated expression of Amelogenin and downregulated the Socs3 expression. *p < 0.05. cl, cervical loop.
In our organ culture, the Runx1 mutant incisor explants shrank and became bent
after 11 days of culture (8/8 cases; Fig. 5B). Interestingly, ZnCl2 treatment increased the length of the mutant explants markedly (8/8 cases; Fig. 5B). The percent-change in the length indicated that ZnCl2 partially rescued the developmental defects of the Runx1 mutants (Fig. 5C). Our histological observation revealed that tissue degradation was evident at the tip of the incisal explants (Fig. S3A), while the proximal region of the tissue, including the cervical loop regions, showed no tissue damage. Ki67 immunohistochemistry further demonstrated that the proliferative activity was retained in both the dental epithelium and the mesenchyme of the explants after long-term culture (Fig. S3B). These findings indicate that the incisor explants of the Runx1 mutants remained viable over the course of culture.
qPCR demonstrated that ZnCl2 treatment significantly upregulated the expression of Lgr5 and Amelogenin (Fig. 5D). The expression of Sox2 was downregulated by ZnCl2 treatment, while the expression of Socs3 was downregulated (Fig. 5D). These findings suggest that a Stat3 activator specifically rescued the suppressed Lgr5 expression in the Runx1 mutant incisor.
Discussion
The Rodent incisors is a self-renewing organ that provides a remarkable example of a requirement for a continuous supply of progenitors from stem cells. The present results provide the first genetic evidence to support that Runx1 has a specific function in regulation of the Lgr5-expressing epithelial stem cells in the growing incisors through Stat3 activation. Furthermore, defective phenotypes of Runx1 mutant were rescued by pharmaceutical Stat3 activation, suggesting that Stat3 phosphorylation regulate the epithelial stem cell, which mediate the Runx1-Lgr5 axis to regulate the homeostasis of the continuous growth of the incisors.
The defective phenotype of the Runx1 mutant incisor was similar to that of Cbfb mutants. Since Cbfb acts as a binding partner for all Runx proteins (Runx1, Runx2 and Runx3)33, the targeted inactivation of its expression abrogates the activity of all Runx complexes34. In wild-type mice, the expression of Runx1 and Runx2 overlapped in the enamel epithelium in the incisors and Runx3 was also evident in the dental epithelium; hence, it was assumed that there might be redundancy among the functions of Runx1, Runx2 and Runx3. In this study, the Runx1 mutant phenotype was almost identical to the Cbfb mutant one, indicating that Runx1 has dominant roles in homeostasis and in the enamel formation of the growing incisors.
To assess how Runx1 signaling affects the epithelial stem cells in the growing incisors, we specifically examined the possible involvement of Stat3 signaling. It is established that Stat3 is essential in the maintenance of stem cells and cancer stem cells. The previous study demonstrated that Runx1 binds directly to the Socs3 promoters to represses their transcription in keratinocytes, and ultimately upregulates Stat3 activity by enhancing phosphorylation16; hence, we assumed that the Stat3 activity might be affected in the Runx1 mutant incisors. qPCR and in situ hybridization in the present study showed that the Socs3 expression was upregulated in association with Runx1 depletion. These findings suggested that Socs3 might contribute—at least in part—to the regulation of Stat3 activity under the control of Runx1 signaling. The self-renewal of stem cells has a regulatory mechanism that is similar to that of the proliferation of cancer cells and STAT3 is an essential component of this mechanism in embryonic stem cells and various types of stem cells and cancer stem cells. Indeed, Runx1 stimulates Stat3 signaling in three epithelial cancer cell types through the transcriptional repression of a Stat3 inhibitor, SOCS3, which triggers cancer cell growth. Although we did not identify the specific kinase or molecule that regulates the phosphorylation of Stat3 in Runx1 signaling, our findings on the Socs3 expression suggested that Runx1 might be involved directly or indirectly in the downregulation of the Socs3 expression, which might explain how Runx1 regulates the Stat3 phosphorylation.
Lgr5 is an established marker of epithelial stem cells in the intestinal crypt and in the hair follicle bulge and germ, and was identified as a Wnt target gene in the intestinal stem cell niche19,35,36. In the growing incisors, Lgr5-expressing cells were identified as actively cycling stem cells in the cervical regions; however, Wnt reporter was not present in the cervical loop epithelium, but was evident in the mesenchyme underlying the cervical loop. Immunostaining revealed that Stat3 is specifically activated in the cervical loop epithelium; thus, we explored the possible association between the Stat3 activity and the stem cell marker genes. It was of interest that in vitro culture with the Stat3-inhibitor Ag-490 or S3I-201 specifically suppressed the expression of Lgr5, but not Sox2. Furthermore, the reintroduction of Stat3 singling using ZnCl2, which activated Stat3 signaling, rescued the suppressed expression of Lgr5 in the Runx1 mutant incisors. These findings clearly indicated that Runx1 signaling could regulate the expression of Lgr5—at least in part—through the modification of Stat3 signaling in the cervical regions. The induction of Lgr5 by the activation of Stat3 is also observed in the progression of skin-related basal cell carcinomas37. On the other hand, the Runx1 mutant mice showed the downregulated expression of Sox2, as well as the expression of Lgr5. It is likely that Runx1 deficiency might also affect regulatory pathways other than the Stat3 signaling pathway, which could influence the stem cells in the cervical loop.
Mice harboring an epidermal-specific constitutive Stat3 deletion were generated using K5-Cre mice. This mouse demonstrated defective wound healing, hair cycling and thymocyte survival; however, tooth phenotypes has not been reported. It is possible that other Stat family might also contribute the homeostasis of the growing incisors and have functional redundancy with Stat3. Another possible explanation is related to the difference in the manner of Stat3 deletion. In the previous study using stat3 mutants, the Cre-mediated deletion of the Stat3 gene was driven by the Keratin5 (K5) promoter. K14-Cre is another well-known Cre driver that is used to generate conditional null mutant mice, and it was of interest that the Dicer null mutants driven by K14 and K5, respectively, do not exhibit identical appendage phenotypes. Indeed, K14-Dicer mutants exhibited more severe phenotypes at birth than K5-Dicer mutants and this difference could be due to the earlier onset of the K14-Cre driver38, or more efficient deletion using the K14 promoter. It is possible that the K14-Cre driver is appropriate for evaluating the function of Stat3 in the growing incisors. We are currently investigating the incisal phenotypes using K14-Cre driven Stat3 conditional null mice.
In conclusion, we found that Runx1 is involved in the regulation of Lgr5-positive epithelial stem cells that differentiate into ameloblasts, through regulation of phosphorylation of Stat3 in the cervical loop of the growing incisors. Furthermore, defective phenotypes of Runx1 mutants were rescued by treatment with a Stat3 activator, suggesting that Stat3 phosphorylation regulates epithelial stem cell, which mediate the Runx1-Lgr5 axis to regulate the homeostasis of the continuous growth of the incisors. Collectively, our study reveals an essential role for Runx1 and Stat3 activation epithelial stem cell function in the growing incisors (Fig. 6).
Figure 6.

A schematic illustration of the key findings of this article. (A) In the cervical loop of the wild type incisors, Runx1 regulates Stat3 phosphorylation, which further regulates the expression of Lgr5 and enamel matrix protein. (B) In the Runx1 mutant incisors, Stat3 phosphorylation was significantly disturbed with the downregulated expression of Lgr5 and enamel matrix proteins. (C) The pharmaceutical Inhibition of Stat3 signaling resulted in the significant downregulation of Lgr5 and enamel matrix proteins. (D) Pharmaceutical Stat3 activation upregulated the expression Lgr5 and enamel matrix proteins in Runx1 mutants.
Materials and Methods
Animals
All of the animal experiments were performed in strict accordance with the guidelines of the animal care and use committee of the Osaka University Graduate School of Dentistry, Osaka, Japan. The protocol was approved by the Committee on the Ethics of Animal Experiments of Osaka University Graduate School of Dentistry. Mice were housed in the animal facility at the Department of Dentistry, Osaka University. Welfare guidelines and procedures were performed with the approval of the Osaka University Graduate School of Dentistry Animal Committee. C57/BL6 mice were used as wild-type (WT) mice. Mice expressing Cre recombinase under the control of keratin 14 promoter (K14-Cre; strain Tg(KRT14-cre)1Amc) were bred with floxed Runx1 mice (Runx1tm3Spe) to produce epithelium-specific Runx1-knockout (K14-Cre/Runx1fl/fl) mice, as described previously9,26. We used their littermates or knockout embryos that did not carry the K14-Cre/Runx1fl/fl genotype as controls. K14-Cre/Runx1fl/fl mice were fed powder food.
The histological analysis
The mouse embryonic heads were dissected in BGJb Medium (Gibco). This tissue was fixed in 4% paraformaldehyde at 4 °C overnight, equilibrated in graded sucrose, and embedded in Tissue-Tek (OCT compound, Sakura).
The Micro-Computed Tomography (Micro-CT) Analysis
The lower incisors of the P28 control and conditional knockout mice were scanned with micro-CT (R_mCT2, Rigaku, Tokyo, Japan) at settings of 90 kV and 160 µA. The SimpleViewer software program (Rigaku) was used for the image analysis.
The in situ hybridization (ISH) analysis
The digoxigenin-labeled RNA probes used in this study were prepared using a DIG RNA labeling kit (Roche) according to the manufacturer’s protocol using each cDNA clone as the template.
The probes were synthesized from fragments of Runx1, Runx2, Runx3, Sox2, Lgr5, Stat3 and Amelogenin (Allen Institute for Brain Science) and were amplified with T7 and SP6 adaptor primers through a PCR. After hybridization, the expression patterns for each mRNA were detected and visualized according to their immunoreactivity with anti-digoxigenin alkaline phosphatase-conjugated Fab fragments (Roche), as previously reported9. At least three embryos of each genotype were used for each analysis.
Immunohistochemistry
After the removal of the OCT compound, immunofluorescence staining was performed on 20-µm sections using polyclonal rabbit-anti-Ki67 (1:400, ab15580, Abcam), monoclonal anti-mouse-K14 (1:200, ab7880, Abcam), monoclonal anti-rabbit-phospho-STAT3 (pStat3, 1:200, #9145, Cell Signaling Technology), monoclonal anti-mouse-STAT3 (Stat3, 1:200, #9139, Cell Signaling Technology) overnight at 4 °C. Then, Alexa488-conjugated donkey-anti-rabbit IgG (1:400, A21206, Molecular Probes) or Alexa546-conjugated goat-anti-mouse IgG (1:400, A11003, Molecular Probes) was used as a secondary antibody for 3 h at room temperature. The sections were then counterstained with DAPI (1:500, Dojindo) and mounted with fluorescent mounting medium (Dako). At least three embryos of each genotype were used for each analysis.
Laser microdissection
The mandible was freshly embedded in OCT compound and frozen immediately. Tissues were serially sectioned at a thickness of 25 μm at −20 °C using a cryostat (CM 1950, Leica). The tissue sections were mounted thawed on a film-coated slide. We stained these slides with cresyl violet. Sections of the cervical loop and the neighboring incisal epithelial tissues were micro-dissected respectively (about 300 μm in length each) using a Leica Micro Laser System (LMD6500; Leica) and collected in a tube, as shown in Supplemental Fig. S1. Representative samples Pre- and Post-laser microdissection from sections of the cervical loop of the incisor are shown in Supplemental Fig. S1. Sections from each genotype were pooled for extraction of mRNA.
Western blotting
The dissected tissues were lysed with RIPA buffer (nacalai tesque) supplemented with protease and phosphatase inhibitors (nacalai tesque). The lysates were centrifuged and the supernatant was heated in denaturing Laemmli buffer (Bio-rad Laboratories). Proteins were separated by SDS-PAGE and transferred to polyvinylidene difluoride membranes (Bio-rad Laboratories). The membranes were incubated with anti-Stat3 (1:1000, #9139, Cell Signaling Technology), anti-pStat3 (1:1000, #9145, Cell Signaling Technology), or beta-actin (1:2000, Sigma). The bound antibodies were detected with HRP-linked antibody (1:1,000, Cell Signaling Technology) and an ECL detection kit (Bio-rad Laboratories).
RNA extraction and the real-time RT-PCR
We used the laser-microdissected tissues of the control and K14-Cre/Runx1fl/fl mice to extract total RNA. Total RNA was isolated using ISOGEN (Nippon Gene) according to the manufacturer’s instructions. Total RNA was reverse transcribed to cDNA using an oligo (dT) with avian myeloblastosis virus reverse transcriptase (Takara). For the real-time RT-PCR, the cDNA was amplified with TaqDNA Polymerase (Toyobo) using a light cycler (Roche). The thermal profile for all SYBR Green PCRs was 95 °C for 30 s followed by 50 cycles of 3-step amplification, including 95 °C for 5 s, 55 °C for 10 s, 72 °C for 15 s, and a melting step, including 95 °C for 1 s, 73 °C for 15 s, 95 °C for 1 s, and 40 °C for 30 s for cooling. Data was normalized to Gapdh and expressed as relative value to the average of control group. The primer sequences are shown in Supplemental Table S1. At least three embryos of each genotype were used for each analysis.
In vitro culturing of the incisors and Stat3 inhibitor treatment
Two different types of Stat3 inhibitors were applied in the in vitro cultures39. To exclude possible effects of serum on Stat3 signaling, the present tooth model was established using chemically-defined BGJB medium (Gibco) without serum supplementation. The lower incisor was dissected from the E16.0 embryo and cultured on a track-etched polycarbonate membrane filter (Nuclepore) in Trowell-type organ culture with serumless, chemically-defined medium (BGJB, Gibco) with or without AG490 (200–400 µM; Sigma-Aldrich) or Stat3 Inhibitor VI, S3I-201 (200–400 µM; Sigma-Aldrich). Trowell-type culture enables the long-term culture of tooth germ explants, and previous in vitro culture has shown that the explanted tooth germs remained viable for four weeks using serumless chemically-defined BGJB medium40. Tissues were harvested after 6 days of culturing, then processed for a qPCR and Western blotting.
In vitro culture of the Runx1 mutant incisors and ZnCl2 treatment
To activate the phosphorylation of Stat3, ZnCl2 was applied in the in vitro cultures. The lower incisor of the Runx1 mutant was dissected from the E16.0 embryo and cultured on a track-etched polycarbonate membrane filter (Nuclepore) in Trowell type organ culture with serumless, chemically-defined medium (BGJb, Gibco) with or without ZnCl2 (Sigma-Aldrich). The tissues were harvested after 11 days of culturing, then processed for a qPCR and Western blotting.
In addition, in order to quantify the morphological changes associated with ZnCl2 treatment in Runx1 mutants, we measured the length of the incisal explants 11 days after culture. Since the explants became bent after long-term culture, the curved incisor length was traced and measured at the midpoint of the labial and lingual surface of the specimens using ImageJ software (NIH). The percent-change in the length of the explants was calculated to quantify the morphological changes associated with ZnCl2 treatment.
Electronic supplementary material
Acknowledgements
We thank Ms. Yuriko Nogami and Ryoko Hoshino for the excellent care and maintenance of our mouse colony and her valuable assistance in the histological, molecular and protein work. This work was supported by grants-in-aid for scientific research program from the Japan Society for the Promotion of Science (#15H02577 and #24249093, to TY).
Author Contributions
H.K. and T.Y. designed the study. S.E.S., H.K., H.O., T.I. and T.S. performed and/or analyzed experiments. I.T. and K.K. provided experimental reagents and participated in the discussions. S.E.S. and T.Y. wrote the manuscript with input from all authors. All authors read and approved the final manuscript
Competing Interests
The authors declare no competing interests.
Footnotes
Electronic supplementary material
Supplementary information accompanies this paper at 10.1038/s41598-018-29317-6.
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Harada H, et al. Localization of putative stem cells in dental epithelium and their association with Notch and FGF signaling. J Cell Biol. 1999;147:105–120. doi: 10.1083/jcb.147.1.105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Juuri E, et al. Sox2+stem cells contribute to all epithelial lineages of the tooth via Sfrp5+progenitors. Dev Cell. 2012;23:317–328. doi: 10.1016/j.devcel.2012.05.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Seidel K, et al. Hedgehog signaling regulates the generation of ameloblast progenitors in the continuously growing mouse incisor. Development. 2010;137:3753–3761. doi: 10.1242/dev.056358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kuang-Hsien HJ, Mushegyan V, Klein OD. On the cutting edge of organ renewal: Identification, regulation, and evolution of incisor stem cells. Genesis. 2014;52:79–92. doi: 10.1002/dvg.22732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Mitsiadis TA, Barrandon O, Rochat A, Barrandon Y, De Bari C. Stem cell niches in mammals. Exp Cell Res. 2007;313:3377–3385. doi: 10.1016/j.yexcr.2007.07.027. [DOI] [PubMed] [Google Scholar]
- 6.Mangan JK, Speck NA. RUNX1 mutations in clonal myeloid disorders: from conventional cytogenetics to next generation sequencing, a story 40 years in the making. Crit Rev Oncog. 2011;16:77–91. doi: 10.1615/CritRevOncog.v16.i1-2.80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Osorio KM, Lilja KC, Tumbar T. Runx1 modulates adult hair follicle stem cell emergence and maintenance from distinct embryonic skin compartments. J Cell Biol. 2011;193:235–250. doi: 10.1083/jcb.201006068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Raveh E, et al. Dynamic expression of Runx1 in skin affects hair structure. Mech Dev. 2006;123:842–850. doi: 10.1016/j.mod.2006.08.002. [DOI] [PubMed] [Google Scholar]
- 9.Kurosaka H, et al. Core binding factor beta functions in the maintenance of stem cells and orchestrates continuous proliferation and differentiation in mouse incisors. Stem Cells. 2011;29:1792–1803. doi: 10.1002/stem.722. [DOI] [PubMed] [Google Scholar]
- 10.Huang G, et al. Dimerization with PEBP2beta protects RUNX1/AML1 from ubiquitin-proteasome-mediated degradation. EMBO J. 2001;20:723–733. doi: 10.1093/emboj/20.4.723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Rawlings JS, Rosler KM, Harrison DA. The JAK/STAT signaling pathway. J Cell Sci. 2004;117:1281–1283. doi: 10.1242/jcs.00963. [DOI] [PubMed] [Google Scholar]
- 12.Reich NC. STATs get their move on. JAKSTAT. 2013;2:e27080. doi: 10.4161/jkst.27080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cartwright P, et al. LIF/STAT3 controls ES cell self-renewal and pluripotency by a Myc-dependent mechanism. Development. 2005;132:885–896. doi: 10.1242/dev.01670. [DOI] [PubMed] [Google Scholar]
- 14.Sano S, et al. Keratinocyte-specific ablation of Stat3 exhibits impaired skin remodeling, but does not affect skin morphogenesis. EMBO J. 1999;18:4657–4668. doi: 10.1093/emboj/18.17.4657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Osorio KM, et al. Runx1 modulates developmental, but not injury-driven, hair follicle stem cell activation. Development. 2008;135:1059–1068. doi: 10.1242/dev.012799. [DOI] [PubMed] [Google Scholar]
- 16.Scheitz CJ, Lee TS, McDermitt DJ, Tumbar T. Defining a tissue stem cell-driven Runx1/Stat3 signalling axis in epithelial cancer. EMBO J. 2012;31:4124–4139. doi: 10.1038/emboj.2012.270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Diaz N, et al. Activation of stat3 in primary tumors from high-risk breast cancer patients is associated with elevated levels of activated SRC and survivin expression. Clin Cancer Res. 2006;12:20–28. doi: 10.1158/1078-0432.CCR-04-1749. [DOI] [PubMed] [Google Scholar]
- 18.Baker BJ, Akhtar LN, Benveniste EN. SOCS1 and SOCS3 in the control of CNS immunity. Trends Immunol. 2009;30:392–400. doi: 10.1016/j.it.2009.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Van der Flier LG, et al. The Intestinal Wnt/TCF Signature. Gastroenterology. 2007;132:628–632. doi: 10.1053/j.gastro.2006.08.039. [DOI] [PubMed] [Google Scholar]
- 20.Harada H, et al. FGF10 maintains stem cell compartment in developing mouse incisors. Development. 2002;129:1533–1541. doi: 10.1242/dev.129.6.1533. [DOI] [PubMed] [Google Scholar]
- 21.Parsa S, et al. Signaling by FGFR2b controls the regenerative capacity of adult mouse incisors. Development. 2010;137:3743–3752. doi: 10.1242/dev.051672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Suomalainen M, Thesleff I. Patterns of Wnt pathway activity in the mouse incisor indicate absence of Wnt/beta-catenin signaling in the epithelial stem cells. Dev Dyn. 2010;239:364–372. doi: 10.1002/dvdy.22106. [DOI] [PubMed] [Google Scholar]
- 23.Yang Z, Balic A, Michon F, Juuri E, Thesleff I. Mesenchymal Wnt/β-Catenin Signaling Controls Epithelial Stem Cell Homeostasis in Teeth by Inhibiting the Antiapoptotic Effect of Fgf10. Stem Cells. 2015;33:1670–1681. doi: 10.1002/stem.1972. [DOI] [PubMed] [Google Scholar]
- 24.Chang JY, et al. Fibroblast growth factor signaling is essential for self-renewal of dental epithelial stem cells. J Biol Chem. 2013;288:28952–28961. doi: 10.1074/jbc.M113.506873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Yamashiro T, Aberg T, Levanon D, Groner Y, Thesleff I. Expression of Runx1, -2 and −3 during tooth, palate and craniofacial bone development. Mech Dev. 2002;119(Suppl 1):S107–110. doi: 10.1016/S0925-4773(03)00101-1. [DOI] [PubMed] [Google Scholar]
- 26.Ono Minagi H, et al. Runx1 mediates the development of the granular convoluted tubules in the submandibular glands. PLoS One. 2017;12:e0184395. doi: 10.1371/journal.pone.0184395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Chang JY, et al. Self-renewal and multilineage differentiation of mouse dental epithelial stem cells. Stem Cell Res. 2013;11:990–1002. doi: 10.1016/j.scr.2013.06.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Motoda L, et al. Runx1 protects hematopoietic stem/progenitor cells from oncogenic insult. Stem Cells. 2007;25:2976–2986. doi: 10.1634/stemcells.2007-0061. [DOI] [PubMed] [Google Scholar]
- 29.Hoi CS, et al. Runx1 directly promotes proliferation of hair follicle stem cells and epithelial tumor formation in mouse skin. Mol Cell Biol. 2010;30:2518–2536. doi: 10.1128/MCB.01308-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Liang J, et al. The stat3/socs3a pathway is a key regulator of hair cell regeneration in zebrafish. [corrected] J Neurosci. 2012;32:10662–10673. doi: 10.1523/JNEUROSCI.5785-10.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Siddiquee K, et al. Selective chemical probe inhibitor of Stat3, identified through structure-based virtual screening, induces antitumor activity. Proc Natl Acad Sci USA. 2007;104:7391–7396. doi: 10.1073/pnas.0609757104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Hu J, et al. Zinc Chloride Transiently Maintains Mouse Embryonic Stem Cell Pluripotency by Activating Stat3 Signaling. PLoS One. 2016;11:e0148994. doi: 10.1371/journal.pone.0148994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Levanon D, et al. AML1, AML2, and AML3, the human members of the runt domain gene-family: cDNA structure, expression, and chromosomal localization. Genomics. 1994;23:425–432. doi: 10.1006/geno.1994.1519. [DOI] [PubMed] [Google Scholar]
- 34.Wang Q, et al. The CBFbeta subunit is essential for CBFalpha2 (AML1) function in vivo. Cell. 1996;87:697–708. doi: 10.1016/S0092-8674(00)81389-6. [DOI] [PubMed] [Google Scholar]
- 35.Barker N, et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature. 2007;449:1003–1007. doi: 10.1038/nature06196. [DOI] [PubMed] [Google Scholar]
- 36.Jaks V, et al. Lgr5 marks cycling, yet long-lived, hair follicle stem cells. Nat Genet. 2008;40:1291–1299. doi: 10.1038/ng.239. [DOI] [PubMed] [Google Scholar]
- 37.Jia J, et al. LGR5 expression is controled by IKKα in basal cell carcinoma through activating STAT3 signaling pathway. Oncotarget. 2016;7:27280–27294. doi: 10.18632/oncotarget.8465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Lyle S, et al. Dicer cooperates with p53 to suppress DNA damage and skin carcinogenesis in mice. PLoS One. 2014;9:e100920. doi: 10.1371/journal.pone.0100920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Gurbuz V, et al. Effects of AG490 and S3I-201 on regulation of the JAK/STAT3 signaling pathway in relation to angiogenesis in TRAIL-resistant prostate cancer cells in vitro. Oncol Lett. 2014;7:755–763. doi: 10.3892/ol.2014.1795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Slavkin HC, et al. Cartilage, bone and tooth induction during early embryonic mouse mandibular morphogenesis using serumless, chemically-defined medium. Connect Tissue Res. 1990;24:41–51. doi: 10.3109/03008209009152421. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.