

Abstract
The microbial diversity and function of terrestrial lichens have been well studied, but knowledge about the non-photosynthetic bacteria associated with marine lichens is still scarce. 16S rRNA gene Illumina sequencing was used to assess the culture-independent bacterial diversity in the strictly marine cyanolichen species Lichina pygmaea and Lichina confinis, and the maritime chlorolichen species Xanthoria aureola which occupy different areas on the littoral zone. Inland terrestrial cyanolichens from Austria were also analysed as for the marine lichens to examine further the impact of habitat/lichen species on the associated bacterial communities. The L. confinis and L. pygmaea communities were significantly different from those of the maritime Xanthoria aureola lichen found higher up on the littoral zone and these latter communities were more similar to those of the inland terrestrial lichens. The strictly marine lichens were dominated by the Bacteroidetes phylum accounting for 50% of the sequences, whereas Alphaproteobacteria, notably Sphingomonas, dominated the maritime and the inland terrestrial lichens. Bacterial communities associated with the two Lichina species were significantly different sharing only 33 core OTUs, half of which were affiliated to the Bacteroidetes genera Rubricoccus, Tunicatimonas and Lewinella, suggesting an important role of these species in the marine Lichina lichen symbiosis. Marine cyanolichens showed a higher abundance of OTUs likely affiliated to moderately thermophilic and/or radiation resistant bacteria belonging to the Phyla Chloroflexi, Thermi, and the families Rhodothermaceae and Rubrobacteraceae when compared to those of inland terrestrial lichens. This most likely reflects the exposed and highly variable conditions to which they are subjected daily.
Keywords: 16SrRNA, Illumina, Marine cyanolichen, Symbiosis, Lichina
Introduction
The lichen symbiosis, commonly recognised as a partnership of a fungus (mycobiont), and a photosynthetic partner (photobiont) arose with the conquest of land in the lower Devonian, according to the first clear fossil evidence (Honegger, Edwards & Axe, 2013). The enclosure of the photobiont partner by protective layers of the fungal partner gave rise to a new morphological structure. This symbiotic structure is called the lichen thallus, which apparently mediates a high degree of tolerance to desiccation (Kranner et al., 2008), and allows many lichens to thrive as poikilohydric organisms in environments characterised by periodic changes in environmental conditions. Therefore, lichens are typically found in habitats where other organisms struggle to persist such as the intertidal belt of coastal rocks, where lichens develop characteristically coloured belts.
Bacteria were already found in the oldest microfossils that can be reliably assigned to lichen thalli (Honegger, Edwards & Axe, 2013), an observation that fits well with the observations of bacterial colonisation of extant lichens (Cardinale et al., 2008). Recent works suggest that the ubiquitous presence of bacteria in lichen thalli contributes to a more complex functional network beyond the interaction of fungi and algae (Aschenbrenner et al., 2016). Bacteria were first cultured from lichens many years ago and were originally supposed to be involved in nitrogen-fixation (Henkel & Yuzhakova, 1936). However, due to the low culturability of bacteria from many environments (Ferguson, Buckley & Palumbo, 1984) and the tendency of culture methods to select for opportunistic species which rarely dominate in the natural environment (Eilers, Pernthaler & Amann, 2000), their diversity was only recently fully recognised.
Culture-independent molecular studies of lichen microbial diversity as well as microscopic observations revealed that bacteria belonging to the Alphaproteobacteria were the dominant microbial group associated with the lichens (Cardinale, Puglia & Grube, 2006; Liba et al., 2006; Grube et al., 2009; Bjelland et al., 2011; Hodkinson et al., 2012; Cardinale et al., 2012a). In these studies, high abundances of Alphaproteobacteria were generally observed on the surface structures of the lichen thalli although some were observed in the fungal hyphae. More recently, the application of high-throughput sequencing techniques to lichen bacterial community analysis confirmed that the composition of the lichen bacterial communities could be more influenced by the mycobiont species than by their sample site (Bates et al., 2011), by the photobiont (Hodkinson et al., 2012), and also by the position in the lichen thallus (Mushegian et al., 2011).
Lichens exhibit clear specificity for substrate and microhabitat conditions, and a clear example of habitat specialisation can be observed for marine lichens, which show vertical zonation in four characteristic belts along rocky coastlines (also known as sublittoral, littoral, supra-littoral and terrestrial zones (Fletcher, 1973a; Fletcher, 1973b)). The common littoral lichen Lichina pygmaea is immersed for several hours each day whereas Lichina confinis occurs higher up in the littoral zone, where it is perpetually subjected to splashing and sea-spray and submerged only during short periods of high tides. Xanthoria spp. can also occur in this zone and in the xeric supralittoral zone, which is exposed to sea-spray during storms but not submerged in seawater. Therefore, marine lichen species certainly experience different levels of stress, ranging from direct sunlight exposure, temperature, salinity and wind variation according to their position in the littoral zone (Delmail et al., 2013).
The genus Lichina belongs to the class Lichinomycetes and is composed of both strictly marine and non-marine species (Schultz, 2017). The marine species L. confinis and L. pygmaea harbour cyanobacterial photobionts closely related to strains of the genus Rivularia (Ortiz-Álvarez et al., 2015). Even though the two lichen species above show a similar distribution range, their cyanobionts belong to separate groups that do not overlap at the OTU or even at the haplotype level (Ortiz-Álvarez et al., 2015). Apart from the cyanobacterial photobiont, the composition of the bacteria associated with marine lichens is poorly studied when compared to those of inland lichens. So far, only Hydropunctaria maura, which forms the black belt on littoral zones, was included in a culture-independent study (Bjelland et al., 2011). Bacterial communities of this lichen were different from inland terrestrial lichens, with higher relative abundances of Actinobacteria, Bacteroidetes, Deinococcus, and Chloroflexi. A recent study of Icelandic marine lichens focused on culturable bacteria, but also revealed by fingerprinting (DGGE) analysis of 16S rRNA genes that the bacterial communities were different among different marine lichen species (Sigurbjornsdottir et al., 2014).
Such differences could be of interest for bioprospecting approaches since lichens are known to be a rich source of natural products (Boustie & Grube, 2005; Parrot et al., 2016). However, lichen-associated bacteria have only recently been discovered as an additional contributor to the lichen chemical diversity, and even though only few lichen-associated bacteria have been studied carefully in this respect, many of them produce secondary metabolites. Some of their compounds have pronounced bioactive properties (see Suzuki et al., 2016 for a review). Nevertheless, the role of these compounds in the symbiosis is not well understood at this stage. So far, bioprospecting of the marine lichens L. pygmaea and L. confinis (and Roccella fuciformis) was carried out by a cultivation-based approach targeting Actinobacteria that led to the isolation of 247 bacterial species including 51 different Actinobacteria (Parrot et al., 2015).
To address the paucity of knowledge on the total diversity and variation of bacteria associated with marine lichens and to provide a base for future bioprospecting efforts, the uncultured microbial communities of the cyanolichen species L. confinis and L. pygmaea and the chlorolichen species X. aureola and Xanthoria parietina were characterised by Illumina sequencing of 16S rRNA gene fragments. In addition, other inland terrestrial cyanolichen species were characterised in parallel to allow us to address the following questions: (i) what are the major bacterial groups associated with the marine and maritime lichens occupying different heights on the littoral zone; (ii) how does the culturable fraction of bacteria from Lichina spp. and Lathagrium auriforme compare to the total bacterial community associated with these species; and (iii) do inland terrestrial cyanolichens share a significant proportion of their symbionts with marine cyanolichens?
Materials & Methods
Sampling design and sample collection
The marine cyanolichens Lichina pygmaea and Lichina confinis were collected on 16 October 2013 from the Houssaye Point, Erquy on the coast of Brittany, France (Table 1, Fig. 1). Lichens were sampled in triplicate in a 3 m radius zone that we defined as a sample cluster. Two other sample clusters of triplicates were sampled further along the littoral zone. For L. confinis, sample clusters 1 and 2 were 22 m apart and clusters 2 and 3 were 14.4 m apart. For L. pygmaea, clusters 1 and 2, and 2 and 3, were both separated by 6 m. Duplicate seawater samples were also taken using sterile 50 ml centrifuge tubes. Similarly, one cluster of three Xanthoria aureola samples was also taken from Houssaye Point and another cluster of Xanthoria parietina was sampled from the rocky coastline of Banyuls-sur-mer on the Mediterranean Sea (Table 1, Table S1). The inland terrestrial cyanolichens were sampled at Kesselfalklamm, Austria. Lathagrium auriforme and Scytinium lichenoides were sampled in the shaded humid zone near to a river whereas Lathagrium cristatum and Lathagrium fuscovirens were collected from rocks in an exposed dry zone (Table 1, Table S1, Fig. 1). Lichens were sampled using a sterile scalpel and gloves, placed in sterile bags and stored at −80 °C.
Table 1. Lichen species analysed in this study.
The different lichen species sampled in this study are presented with details on their corresponding mycobiont and photobiont partners, their habitat and the location of the sampling sites.
| Lichen species | Mycobiont | Photobiont | Habitat | Date | Location |
|---|---|---|---|---|---|
| Lichina confinis | Lichinomycetes, Lichinales, Lichinaceae | Cyanobacteria: Rivularia sp | Upper eulittoral | 16.10.13 | Erquy, France (Atlantic Ocean) |
| Lichina pygmaea | Lichinomycetes, Lichinales, Lichinaceae | Cyanobacteria: Rivularia sp | Lower eulittoral | 16.10.13 | Erquy, France (Atlantic Ocean) |
| Xanthoria aureola | Lecanoromycetes, Teloschistales, Teloschistaceae | Green alga: Trebouxia sp | Mesic-supralittoral | 16.10.13 | Erquy, France (Atlantic Ocean) |
| Xanthoria parietina | Lecanoromycetes, Teloschistales, Teloschistaceae | Green alga: Trebouxia sp | Xeric-supralittoral | 06.03.14 | Banyuls sur mer, France (Mediterranean Sea) |
| Lathagrium auriforme | Lecanoromycetes, Peltigerales Collemataceae | Cyanobacteria: Nostoc sp. | Terrestrial | 26.11.13 | Kesselfalklamm, Austria |
| Lathagrium cristatum | Lecanoromycetes, Peltigerales Collemataceae | Cyanobacteria: Nostoc sp. | Terrestrial | 26.11.13 | Kesselfalklamm, Austria |
| Lathagrium fuscovirens | Lecanoromycetes, Peltigerales Collemataceae | Cyanobacteria: Nostoc sp. | Terrestrial | 26.11.13 | Kesselfalklamm, Austria |
| Scytinium lichenoides | Lecanoromycetes, Peltigerales Collemataceae | Cyanobacteria: Nostoc sp. | Terrestrial | 26.11.13 | Kesselfalklamm, Austria |
Figure 1. Examples of lichens collected in this study.

(A) Lichina pygmaea (B) Lichina confinis (C) Xanthoria aureola (Brittany) (D) Lathagrium auriforme (E) L. confinis sample cluster (F) X. aureola sample cluster. Photos by Delphine Parrot.
The morphological identification of the lichens was verified by sequencing the marker genes ITS-5.8S rRNA (White et al., 1990) for the marine and maritime lichens and the genes beta-tubulin (Myllys, Lohtander & Tehler, 2001) and nuclear LSU rRNA (Vilgalys & Hester, 1990; Otálora et al., 2010) for the inland terrestrial lichens (Supplementary Information, Table S2).
DNA extraction
Genomic DNA was extracted using the DNeasy PowerPlant Pro DNA Isolation Kit (Qiagen S.A.S., Courtaboeuf, France). Lysis was achieved by first grinding the lichens (several pieces selected randomly from each thallus amounting to approximately 0.1 g of material) to a powder with liquid nitrogen using a sterile pestle and mortar. The powder was added to the lysis tubes of the kit following the protocol according to the manufacturer’s instructions, including a heating step of 65 °C for 10 min before the addition of the RNase A solution. Homogenisation was achieved using a standard vortex equipped with a flat-bed vortex pad. DNA quality was verified by agarose gel electrophoresis and DNA was quantified using the Quant-iT™ PicoGreen® dsDNA Assay Kit (Invitrogen, Thermo-Fisher Scientific Inc., Courtaboeuf, France) according to manufacturer’s instructions.
PCR amplification of 16S rRNA genes and illumina sequencing
Bacterial 16S rRNA gene fragments were amplified using the primer pair 341F (CCTACGGGNGGCWGCAG) and 805R (GACTACHVGGGTATCTAATCC) which show the best combination of domain and phylum coverage (Klindworth et al., 2012). The 341F primers were tagged at the 5′end with different 7 bp tags for each sample (Table S3) that were chosen from a set of tags designed to be robust to substitution, deletion and insertion errors incurred in massively parallel sequencing (Faircloth & Glenn, 2012). We also included in parallel a control consisting of DNA from a synthetic mock community (Mock) of 20 bacterial species containing equimolar (even) rRNA operon counts (HM-782D; Genomic DNA from Microbial Mock Community B, Even, Low Concentration, BEI Resources, Manassas, VA. This standard is now obtainable from LGC Standards S.a.r.l., Molsheim, France; reference ATCC® MSA-1002™). Between 1-10 ng of each DNA sample (or 1 µl of Mock DNA) were amplified in duplicate 10 µl reactions containing 1X KAPA 2G Fast Ready Mix (Merck, Darmstadt, Germany) and 0.5 µM of each primer. The PCR cycling conditions were 95 °C for 3 min followed by 25 cycles of 95 °C for 15s, 55 °C for 15s and 72 °C for 2s, and a final extension of 72 °C for 30s. Duplicate reactions were pooled, and PCR amplification verified by gel electrophoresis. To normalise the samples before pooling and sequencing, the Sequalprep Normalization Plate (96) kit (Invitrogen) was used according to manufacturer’s instructions. After binding, washing and elution, the 37 environmental samples and the mock DNA sample were pooled into one tube. The clean-up of the PCR amplicon pool was achieved with the Wizard SV Gel and PCR Clean-Up System (Promega, Charbonnières-les-Bains, France) according to manufacturer’s instructions with elution in 30 µl of molecular biology grade water (Merck). DNA was quantified with the Quant-iT PicoGreen dsDNA Assay kit (Invitrogen) according to manufacturer’s instructions. Approximately 350 ng DNA was pooled with 700 ng of barcoded PCR products from a different project (sequencing the 16S rRNA genes of marine bacterioplankton) and sent out to the sequencing company Fasteris (Geneva, Switzerland) for library preparation and sequencing. Library preparation involved ligation on PCR using the TruSeq DNA Sample Preparation Kit (Illumina, San Diego, CA, USA) according to manufacturer’s instructions except that five PCR cycles were used instead of 10 cycles. The library was sequenced on one Illumina MiSeq run using the 2 × 300 bp protocol with MiSeq version 3.0 chemistry and the basecalling pipeline MiSeq Control Software 2.4.1.3, RTA 1.18.54.0 and CASAVA-1.8.2. The error rate was measured by spiking the library with about 0.5% of a PhiX library and mapping the reads onto the PhiX reference genome.
Sequence pre-processing
Paired-end Illumina sequence data was processed with a custom pipeline (Supplementary Information) using mainly the USEARCH-64 version 8 package (Drive5; Tiburon, CA, USA) with some commands from QIIME version 1.9.1 (Caporaso et al., 2010) and mothur version 1.33.3 (Schloss et al., 2009). Analysis of the Mock data set first allowed us to calibrate the different steps of the sequence processing pipeline ensuring that the assigned OTUs reflected as closely as possible the defined composition of the sample and allowed us to reveal potential sequencing artifacts (errors, contamination). To minimise errors and reduce over-inflation of diversity, the following criteria were chosen in the pre-processing steps; (a) zero mismatches allowed in the overlapping region when merging the paired end reads, (b) quality filtering was carried out after merging using a stringent expected error of 1.0 (Edgar & Flyvbjerg, 2015), (c) zero mismatches allowed in the barcode when demultiplexing, (d) exact matches to both primers required, (e) removal of singleton sequences that are expected to have errors and that increase the number of spurious OTUs (Edgar, 2013). Sequence data was submitted to the European Nucleotide Archive under the study number: PRJEB23513.
Sequence data analysis
OTUs were defined with the UPARSE algorithm, part of the USEARCH-64 package, which clusters sequences with >97% identity to OTU centroid sequences (representative sequences that were selected based on their rank of read abundance after dereplication) and that simultaneously removes chimeras (Edgar, 2013). Subsequent analysis steps were performed in QIIME or MACQIIME version 1.9.1. OTU taxonomy was assigned with the RDP classifier using the GreenGenes OTU database (gg_13_8_otus) defined at 97% identity. For the lichen dataset, the OTU table was filtered to remove eukaryotic, archaeal and organelle sequences (chloroplast, mitochondria). Due to the presence of the lichen photobiont, a large number of cyanobacterial sequences were recovered. The OTU table was filtered to separate non-cyanobacteria sequences from the cyanobacteria sequences. The representative sequences were aligned using the PYNAST aligner in QIIME and the 13_8 GreenGenes alignment (Supplementary Information), and the OTU table was further pruned by removing non-aligning OTUs. Data exploration and analysis was carried out with QIIME (see Supplementary Information) and with the packages Phyloseq (McMurdie & Holmes, 2013) and vegan (Oksanen et al., 2013) in R (R Core Team, 2015). One-way ANOSIM tests were performed using the vegan package in R to determine significant differences between groups. To examine the patterns of phylogenetic relatedness of the lichen-associated bacterial communities, the mean pairwise distance (MPD) and the mean nearest taxon distance (MNTD) were calculated using the Picante package in R (Kembel et al., 2010). Whereas MPD calculates the mean pairwise distance between all OTUs in each community, MNTD calculates the mean distance between each OTU in the community and its closest relative. To test the significance of the phylogenetic community structure observed, the standardised effect size (SES) was then calculated for the MPD and MNTD to compare the observed pattern to that of a null model of phylogeny. We used the null model of randomly shuffling the tip labels. SES(MPD) and SES(MNTD) are equivalent to −1 times the Net Relatedness Index (NRI) and the Nearest Taxon Index (NTI) respectively, reported previously (Webb, Ackerly & Kembel, 2008).
Since analysis of the Mock data revealed the presence of contaminant and/or sequences from a library from another project, potentially due to cross-talk during the Illumina sequencing, OTUs exhibiting a low uniform abundance amongst all the samples were filtered out by calculating between-sample variance on a relative abundance OTU table and removing those OTUs with a variance of <1 ×10−6.
Comparison of sequences with cultured species, and the NBCI database
The 16S rRNA gene sequences of 247 bacterial strains previously isolated from the lichens L. pygmaea, L. confinis, and L. auriforme (Parrot et al., 2015) were compared to the sequences from this study by blastn (Altschul et al., 1997) queries against a database of the representative OTU sequences in the current study using the standalone version of NCBI blast. The bacterial strains were isolated from the lichens in France and Austria 18 months and 12 months respectively before those sampled in this study, but they were sampled at exactly the same sites. Note that in the previous study (Parrot et al., 2015), the lichen L. auriforme was cited with its previous name; C. auriforme (Collema auriforme).
In addition, the sequences of the top 30 OTUS from the strict marine or maritime-inland terrestrial lichens were subjected to blastn queries against the NCBI Genbank nt database.
Results
Control of sequence quality and data treatment
The mock community sequences were parsed out and analysed separately from the other sequences obtained from the lichen and seawater samples. Analysis of the 20-species mock community allowed us to check for potential contamination and also to refine our bioinformatic analysis pipeline to obtain the expected number and identity of OTUs. Using the criteria described in the methods and the algorithm UPARSE of the USEARCH package for clustering OTUs at 97% identity, we obtained 19 OTUs (identical to the reference sequence Edgar, 2013) which perfectly corresponded to the mock community (the two Staphylococcus sp. strains cluster together) plus 17 other OTUs. The 19 mock OTUs accounted for 99.5% of the sequences. Of the remaining 17 OTUs, four were affiliated to the Cyanobacteria class and three to chloroplasts that were the dominant OTUs found respectively in the lichen samples (this study) and in samples from the offshore bacterioplankton study loaded in the same MiSeq run. This is evidence of the sequencing artifact termed “cross-talk” which can be due to index misassignment where reads from one sample are wrongly assigned the indexes from a different sample. The remaining 10 mock OTUs accounted for 0.24% of the reads.
After the different sequence processing steps on the lichen and seawater samples, a total of 509,656 good quality sequences obtained were clustered into 93 cyanobacterial OTUs and 2,519 bacterial OTUs. Examination of the OTU tables of the study samples also revealed cross-talk OTUs as observed in the mock sample that were mainly attributed to chloroplasts and cyanobacteria. This problem was partly resolved by removal of these lineages for subsequent analysis of the heterotrophic bacterial diversity but there were nevertheless some bacterioplankton OTUs present at consistently low abundances in all the lichen samples, which are dominant in the samples from different study (e.g., Pelagibacteraceae). Therefore, for certain analyses, the OTU table was filtered for low variance OTUs which allowed the removal of spurious OTUs. This data treatment would also have removed non-spurious low abundance OTUs but as our analysis was focused on the dominant (most abundant) OTUs and not the rare OTUs, this would not affect our major findings.
As expected for cyanolichens, a high proportion of the sequences were affiliated to the Cyanobacteria lineage (including chloroplasts) and ranged from 30–60% for the marine lichens and from 40–85% for the inland terrestrial lichens (Fig. S1). The cyanobacterial sequences were then removed for further analyses. The number of non-cyanobacteria bacterial sequences per sample ranged from 2,759 for L. auriforme to >14,000 sequences for X. aureola.
Bacterial communities associated with lichens in different areas of the littoral zone
For clarity, we use the term “marine lichens” to refer to the two Lichina species that are strictly marine, “maritime lichens” to refer to the two Xanthoria species and “inland terrestrial lichens” the other lichens isolated from Austria.
The alpha diversity of the non-cyanobacteria bacterial communities associated with the marine and maritime lichens and in the adjacent seawater was compared using different species diversity and richness indices calculated with a rarefied OTU table (Fig. 2). The marine and maritime lichens showed a significantly lower diversity than the inland terrestrial lichens (t33 = 3.35, p = 0.001 for Inverse Simpson). L. pygmaea bacterial communities exhibited higher species richness than those of L. confinis (observed OTUs and ACE) but showed lower diversity as indicated by the Shannon and Inverse Simpson diversity metrics.
Figure 2. Alpha diversity of bacterial communities associated with marine, maritime and inland terrestrial lichens, and in seawater.

The richness ((A) Observed OTUs and (B) Abundance-based Coverage Estimator (ACE) of species richness), and diversity ((C) Shannon diversity and (D) Simpson diversity) metrics for the lichen species or seawater bacterial communities are displayed by position on the littoral zone for the marine and maritime lichens and the lichens from Austria are plotted as the Terrestrial category.
The taxonomic diversity at the phylum level of the non-cyanobacteria bacteria associated with the different lichens is shown in Fig. 3. Bacteroidetes and Proteobacteria were the dominant phyla in the marine and maritime lichens although the Lichina lichens showed a higher relative abundance of Bacteroidetes (>50% of the sequences) and Chloroflexi compared to the Xanthoria species. The two chlorolichen Xanthoria species collected from either the Mediterranean Sea or Atlantic Ocean coastlines were also associated with the candidate phylum FBP that was either absent or associated at very low abundance in the Lichina spp. Interestingly, the phylum Thermi comprising many thermophilic species were predominantly associated with the Lichina and Xanthoria lichens.
Figure 3. Phylogenetic diversity of the bacteria associated with different lichen species or in seawater.

The relative abundances of the major bacterial phyla and Proteobacteria classes associated with lichens or in seawater are expressed as percentages of the total sequences. Phyla that accounted for less than 2% of the total sequences were grouped together as “Other”.
A comparison of the lichen bacterial community beta diversity as assessed by Bray–Curtis dissimilarity of a rarefied OTU table and hierarchical clustering is shown in Fig. 4A. The dendrogram shows a clear separation of the marine cyanolichens from the seawater samples and also from the maritime lichens whose bacterial communities formed a separate cluster with the inland terrestrial cyanolichens. The data were then reanalysed, focusing on the most abundant OTUs associated with the lichens. This involved removing the seawater samples from the dataset and filtering out low variance OTUs thus reducing the OTU number from 2,519 to 367. Despite this filtering step, the clustering of the lichen communities of the reduced dataset (Fig. S2) was similar to that of the full dataset (Fig. 4A). ANOSIM tests confirmed that the separation of marine versus the other lichen groups, and also the separation of the L. confinis and L. pygmaea groups was highly significant (R = 1, p = 0.001 for both tests). The diversity of the bacterial communities in the replicate samples of the maritime chlorolichens X. aureola and X. parietina clustered according to sampling location (Atlantic Ocean or Mediterranean Sea coast).
Figure 4. Beta diversity of bacterial communities associated with the lichens or seawater assessed by Bray Curtis dissimilarity and average linkage clustering.
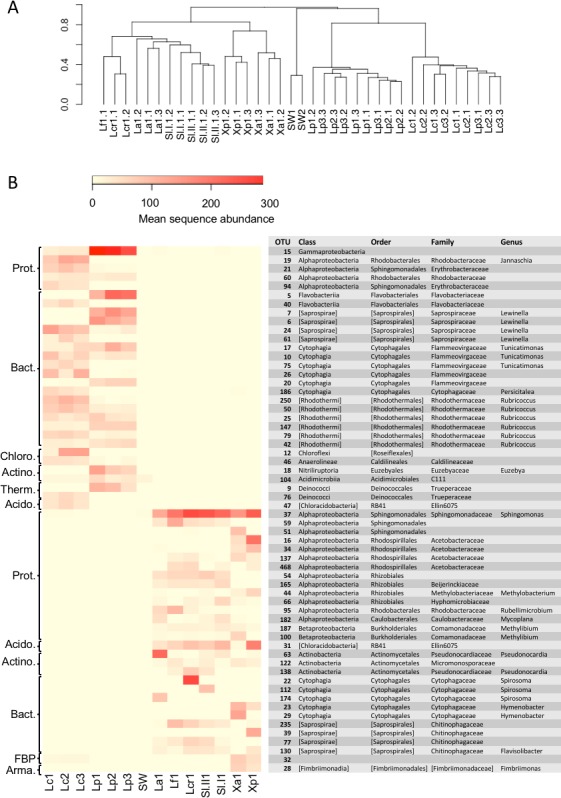
(A) Beta diversity of bacterial communities associated with the lichens or seawater assessed by Bray Curtis dissimilarity and average linkage clustering (B) Heatmap showing the relative abundance of the 30 most abundant OTUs among either the marine or the maritime-inland terrestrial lichens. The mean sequence abundance was calculated for each lichen sample cluster or the seawater samples. Lf, L. fuscovirens; Lcr, L. cristatum; La, L. auriforme; Sl, S. lichenoides; Xp, X. parietina; Xa, X. aureola; SW, seawater; Lc, L. confinis; Lp, L. pygmaea.
At the OTU level, the marine versus the maritime-inland terrestrial lichens were associated with very different bacterial communities as illustrated in Fig. 4B, which shows the 30 most abundant OTUs for either group. The closest relatives of the OTU representative sequences assessed by blastn are presented in Table S4. Over 80% of the representative OTU sequences showed >97% identity hits to previously deposited sequences. OTUs with a lower % identity belonged mainly to the Bacteroidetes phylum. Interestingly, one Chloroflexi OTU (OTU_12) that was recovered from L. confinis showed only a very low 91% identity to its closest previously described relative (Table S4) and half of the closest relatives for the marine lichen OTUs originated from marine intertidal outcrops (Couradeau et al., 2017).
A gammaproteobacterial OTU dominated in L. pygmaea and although this OTU was also present in L. confinis, it was not the most abundant. L. confinis showed a more even distribution of the OTUs compared to L. pygmaea as reflected by the diversity metrics. Despite the relative spatial proximity of X. aureola and the Lichina lichens on the Atlantic coast, there were very few shared OTUs among those genera. One OTU affiliated to the candidate phylum FBP was present at low abundance in L. confinis and Xanthoria sp. but not in L. pygmaea. The microbial communities recovered from the L. pygmaea samples also showed very few OTUs in common with the seawater that was sampled in proximity to this lichen. This suggests that the L. pygmaea communities were specifically associated to the lichen and very different from the free-living seawater communities.
Interestingly the Lichina lichens were associated with several OTUs belonging to the Rhodothermaceae family (Bacteroidetes) and the Deinococcus-Thermus and Chloroflexi phyla of which some members are meso- or thermophilic, highly resistant to UV radiation (Deinococcus) and phototrophic (Chloroflexi). Although the Lichina communities showed a similar genus-level composition with closely related OTUs (eg Lewinella, Tunicatimonas, Rubricoccus) present in both lichens, these OTUs often showed an inverse pattern of abundance between the two species, where OTUs in genera present in both species tended to have higher counts in only one of the species (Fig. 4).
Phylogenetic analysis has already been used to show that environmental filtering rather than competitive effects structure marine bacterial communities (Pontarp et al., 2012). Similarly, to examine the potential ecological processes that could have influenced the community structure of the non-cyanobacteria bacteria in association with the lichen species, the level of phylogenetic clustering (indicative of environmental filtering) or over-dispersion (competitive effects) was estimated from a calculation of the Net Relatedness Index (NRI) and the Nearest Taxon Index (NTI; (Webb, Ackerly & Kembel, 2008). With the exception of the NRI for communities associated with L. confinis, there was evidence of phylogenetic clustering of the bacterial OTUs for the marine and maritime lichens as observed by a positive NRI and NTI (Table 2).
Table 2. Levels of phylogenetic clustering of the lichen associated bacterial communities as measured by the Nearest Taxon Index (NTI) and the Net Relatedness Index (NRI).
| Lichen species | N ∘ taxa | NRI | runs > null | NTI | runs > null |
|---|---|---|---|---|---|
| Coastal Brittany lichens | |||||
| L. confinis | 271 | 1.42 | 905 | 2.30** | 991 |
| L. pygmaea | 281 | 3.49** | 998 | 1.68* | 951 |
| X. aureola | 224 | 3.13** | 997 | 2.15* | 978 |
| Inland Austrian lichens | |||||
| S. Lichenoides | 294 | 2.15* | 983 | 0.12 | 552 |
| L. auriforme | 260 | 2.53** | 992 | 1.73 | 944 |
| L. cristatum | 226 | 1.32 | 902 | 0.92 | 825 |
| L. fuscovirens | 205 | 1.05 | 859 | 1.61 | 938 |
Notes.
The values of each metric for the coastal or inland lichens are presented together with the number of runs where the observed NRI/NTI values were greater than those of the null model.
p < 0.05.
p < 0.01.
Shared and specific bacterial OTUs in the marine and maritime lichens
To explore the degree of overlap between the marine and maritime lichen bacterial communities in more detail, the OTUs consistently shared between the species, and potentially indicative of a core microbiome were identified (subsequently referred to as core OTUs). These OTUs were grouped at the family (or order when identification not possible) level and represented as relative abundance (Fig. 5). Setting a very conservative threshold of 100% presence in all replicates of the marine lichen species, only 33 OTUs were shared by both species (Fig. 5A). Of these, 15 were attributed to the Bacteroidetes families Saprospiraceae, Flammeovirgaceae and Rhodothermaceae (genera Lewinella, Tunicatimonas and Rubricoccus respectively). These OTUs accounted for around 60% of the sequences amongst the core OTUs and showed similar relative abundances between the two Lichina species (Fig. 5A). Other families included the Chloroflexi Caldilineaceae, and the Alphaproteobacteria families Rhodobacteraceae and Erythrobacteraceae that were more abundant in L. confinis, and a Gammaproteobacteria OTU in L. pygmaea.
Figure 5. Comparison of relative abundance of core or specific OTUs present in L. confinis and L. pygmaea.

Comparison of relative abundance of core OTUs present in (A) all replicates of both L. confinis and L. pygmaea or (B) the core OTUs specific for either species grouped at the family level. When family level assignment was not available, the next higher taxonomic level assigned was used. (C) Venn diagram showing the number of core OTUs shared or specific to L. confinis and L. pygmaea.
When considering OTUs associated to each lichen species, 38 and 42 OTUs were specific (i.e., present at 100% of the replicates) for L. confinis and L. pygmaea respectively, and their relative abundances are shown in Figs. 5B and 5C, grouped at the family (or order when identification was not possible at the family level) level. The distribution and relative abundance of the family level were very different between the two species. For L. confinis, the most noticeable differences included a higher relative abundance of Acidobacteria Ellin6075, Bacteroidetes Cytophagaceae and Flammeovirgaceae and the Chloroflexi order Rosiflexales. For L. pygmaea, the Actinobacteria family Euzebyaceae, and the Bacteroidetes families Saprospiraceae and Flavobacteriaceae dominated the specific core OTUs.
To determine the OTUs that showed statistically significant differential abundances between L. confinis and L. pygmaea, we used the DESeq2 negative binomial Wald test (Love, Huber & Anders, 2014). One third of the OTUs tested (120 out of 367 OTUs) showed a significantly different distribution between the two lichens (P < 0.001) and those OTUs with a mean abundance >20 are shown in Fig. 6. These OTUs were distributed in several shared families (left side of the plot) but we also observed differentially distributed OTUs that were distinct for each lichen. Interestingly, 29 out of the 33 core Lichina OTUs showed a statistically significant different distribution between the two species.
Figure 6. OTUs with significantly different relative abundances in L.confinis and L. pygmaea.

OTUs with significantly different relative abundances in L.confinis and L. pygmaea were assessed by the DESeq2 package using an adjusted p value < 0.001 and a base mean threshold > 20. NA : OTUs not assignable at the particular taxonomic level (order or family).
X. aureola collected above L. confinis on the littoral belt only shared one core OTU with the Lichina lichens that was assigned to the Gammaproteobacteria OTU, the most abundant OTU in L. pygmaea. X. aureola and X. parietina sampled from the Atlantic or Mediterranean French coasts shared 24 core OTUs (Fig. S3) that were dominated by the Proteobacteria (mostly Alpha- and Betaproteobacteria) accounting for over 50% of the sequences for each species. X. parietina, collected from the dryer Mediterranean coast, showed higher relative abundances of Acidobacteria and the family Acetobacteraceae (Alphaproteobacteria) compared to X. aureola.
Inland terrestrial cyanolichen-associated bacterial communities
Out of the four inland terrestrial lichen species, L. auriforme and S. lichenoides showed the highest diversity and species richness, followed by L. cristatum and L. fuscovirens (Fig. 2). As observed in previous surveys of terrestrial lichen bacterial diversity, these inland lichen species were also dominated by the phylum Proteobacteria, attributed mainly to the class Alphaproteobacteria (Fig. 3). The phylum distributions were generally similar amongst these species except the Bacteroidetes were almost twice as abundant in L. cristatum and L. fuscovirens, compared to the abundance in the other two species.
The Bray–Curtis dissimilarity dendrogram (Fig. 4A) shows that the bacterial communities associated with L. auriforme were closely related to those of S. lichenoides (both species were from the humid forest gorge) and this was confirmed with an ANOSIM test that showed no significant difference between the species groups (R = 0.239, p = 0.064). The other two species L. cristatum and L. fuscovirens from the dryer, rocky zone clustered apart and the separation of these two groups (humid forest versus dry/exposed) was also significant (R = 0.582, p = 0.017).
At the OTU level, the genus Sphingomonas (Alphaproteobacteria) was highly represented in the maritime and inland terrestrial lichens and remarkably absent in the marine lichens. Other OTUs present in all the inland terrestrial species included two OTUs assigned to the order Rhizobiales, a Betaproteobacteria OTU assigned as Methylibium, an Acidobacteria OTU and two Bacteroidetes OTUs belonging to the Chitinophagaceae. For the inland terrestrial lichens, significant NRI values were only observed for S. lichenoides and L. auriforme but none of these lichens showed significant phylogenetic clustering at the tree tips for the NTI (Table 2).
Comparison of culturable species and total bacterial diversity from coastal lichens and L. auriforme
The 16S rRNA gene sequences of previously isolated strains originating from L. confinis, L. pygmaea, and L. auriforme were compared to the sequences in this study to investigate the representation of the culturable fraction relative to the bacterial diversity assessed by the cultivation-independent molecular method. Thirty-four strains out of the 254 previously isolated strains had 16S rRNA sequence identities >97% to the OTUs identified in this study (Table S5). The majority of the OTUs that had 16S rRNA sequences with high identity (>99%) to the strains were of low abundance (<0.5% of total sequences) in our study. One exception included the abundant OTU_37 (Sphingomonas; ∼4–14% of total sequences) that shared 99% identity to a strain isolated from L. auriforme. One of the top 30 most abundant OTUs from the marine lichens (OTU_21, Erythrobacteraceae) shared 98% sequence identity with a strain isolated from L. pygmaea.
Diversity of lichen photobionts and non-photobiont cyanobacteria
The major photobiont of each of the cyanolichens was identified as the cyanobacterial OTU with the highest relative abundance among all the cyanobacterial sequences, and these identifications were confirmed with that reported in the literature (Otálora et al., 2010; Ortiz-Álvarez et al., 2015). The marine and inland terrestrial cyanolichens were associated with a different major photobiont. For L. confinis and L. pygmaea, the majority of the cyanobacterial reads clustered into a single Rivularia sp. OTU accounting for 51–82% of the reads for the L. confinis replicate samples and 25–79% for those of L. pygmaea. The remaining cyanobacterial sequences belonged principally to the order Chroococcales (including the family Xenococcaceae and the genus Chroococcidiopsis) and the family Pseudanabaenaceae (Fig. S4). For the inland terrestrial lichens, two abundant Nostoc OTUs were identified. The first OTU was attributed as the primary photobiont in L. auriforme, L. cristatum and L. fuscovirens, accounting for 92–98% of all cyanobacterial sequences and was also present (albeit at less than 5.5% of all cyanobacterial sequences) in S. lichenoides. The second Nostoc OTU was the major photobiont of S. lichenoides, accounting for 92–94% of total cyanobacterial sequences in four out of five replicates (67% in the fifth) but had a low abundance in the other terrestrial lichens (at less than 0.5%). Therefore, in contrast to the marine lichens, the inland terrestrial lichen photobionts dominated the cyanobacterial sequences. The non-photobiont OTUs of these lichens included, amongst others, the genus Calothrix and members of the Chroococcales and together they did not exceed 10% of the total cyanobacterial sequences (Fig. S4). As to be expected for chlorolichens, few cyanobacterial sequences (<5% of total reads) were obtained for the Xanthoria lichens but one chloroplast OTU was abundant in both species. Although only assigned as far as the order Chlorophyta, this OTU most likely represents the major photobiont of Xanthoria which is known to be the green alga Trebouxia.
Discussion
To date, the largest body of lichen research has been undeniably devoted to terrestrial species. We address this knowledge gap in applying, for the first time, next generation Illumina sequencing to characterise the bacterial communities associated to strict marine cyanolichens and a maritime chlorolichen, occupying different areas of the littoral zone. Whereas the two marine Lichina species, occurring in two neighbouring zones, share some common OTUs, many are specific to each species and there was very little overlap between the marine and maritime lichen bacterial communities.
Lichina bacterial communities
A comparison of the Lichina bacterial community diversity allowed us to identify potential core OTUs and specific OTUs for each species, reflecting the potential adaptation of L. confinis or L. pygmaea to their different littoral zones. The Lichina bacterial communities shared 15 of 33 core OTUs that were attributed to only three Bacteroidetes genera, Rubricoccus, Lewinella, and Tunicatimonas, as well as several Alphaproteobacteria OTUs from the Erythrobacteraceae and Rhodobacteraceae families, and a Chloroflexi OTU. Tunicatimonas pelagia which belongs to the Flammeovirgaceae family was initially isolated from a sea anemone (Yoon et al., 2012) and other members of this family are known to associate with corals (Apprill, Weber & Santoro, 2016). Lewinella belongs to the Saprospiraceae family that was found as an epiphyte of red algae (Miranda et al., 2013) and on filamentous bacteria identified as Chloroflexi (Xia et al., 2008), and isolates from this genus were also cultured from marine lichens (Sigurbjornsdottir et al., 2014; Parrot et al., 2015). Notably, the bacterial groups associated with the Lichina lichens were also similar to those found in the surface layer of hypersaline microbial mats (Schneider et al., 2013) and in epiphytic communities on the red macroalga Porphyra umbilicalis (Miranda et al., 2013). The co-occurrence of these different bacteria in microbial mats and on various marine hosts suggests that they have adapted to similar environmental conditions. In functioning as a consortium, these communities could potentially increase their robustness and that of their hosts by expanding their range of metabolic capacities and resistance mechanisms to stress (Hays et al., 2015).
Interestingly there were multiple Lewinella, Tunicatimonas and Rubricoccus OTUs in each lichen species and OTUs that were affiliated to the same genus were differentially distributed between the two Lichina species (e.g., see the Lewinella OTUs 6, 7, 24 and 61 in Fig. 4). This could suggest that these bacterial genera belong to ecologically differentiated pools as was already shown for the for the Rivularia cyanobionts of the same lichen species (Ortiz-Álvarez et al., 2015). This mechanism of niche adaptation is thought to occur in extreme environments and could also apply to the bacteria associated to these marine lichens, which became adapted to the intertidal or supratidal zones. The presence of these closely related clusters was also supported by the positive and significant NTI values (Webb et al., 2002) indicative of environmental filtering effects on the Lichina bacterial communities.
Rocky tidal zones are thought to be one of the most stressful habitats on earth (Miller, Harley & Denny, 2009) and both marine lichen species bacterial communities were associated with several groups of bacteria that are known to be heat- or radiation-resistant. These include several OTUs identified as core OTUs that were affiliated to the genus Rubricoccus of the Rhodothermaceae (Park et al., 2011), to the phylum Chloroflexi (that contains moderately thermophilic/thermophilic groups), and to the family Truperaceae in which the isolated species are thermophilic and nearly as radiation resistant as Deinococcus (Albuquerque et al., 2005).
Despite sharing 33 core OTUs, the two marine Lichina species L. confinis and L. pygmaea were associated with significantly different bacterial communities at the OTU level and remarkably the Bray Curtis dissimilarity distance between these communities was greater than that of the distance between the communities associated with two different inland terrestrial cyanolichen genera, Lathagrium auriforme and Scytinium lichenoides (Fig. 4A). This difference was also reflected in the high number of OTUs (>120) that displayed a significantly different abundance between the two species (Fig. 6). These significant differences were despite the relatively short distance that separated the species on the vertical gradient (height on the littoral belt). Other studies show evidence that height on the littoral zone is the most important factor explaining the rocky coastline species variance with slope, exposure, substrate and orientation explaining less (Chappuis et al., 2014). L. confinis occurs higher up on the littoral belt, being mostly exposed to the air and would be subject to higher temperatures and UV radiation. This could explain why some thermophilic groups such as Chloroflexi and the Actinobacterial genus Rubrobacter that include thermophilic species resistant to radiation (Ferreira et al., 1999) were four-fold and 13-fold respectively more abundant in L. confinis than L. pygmaea. The bacterial groups that were more abundant in L. pygmaea when compared to L. confinis included the Gammaproteobacteria OTU related to a seagrass epiphyte Granulosicoccus (Kurilenko et al., 2010) an OTU affiliated to the Actinobacterial genus Euzebya, recovered from a sea cucumber (Kurahashi et al., 2010) and several Flavobacteriaceae and Rhodobacteraceae OTUs. Marine epiphytic Actinobacteria, and Phaeobacter (Rhodobacteraceae), associated with a variety of marine plants and animals, are known to produce bioactive molecules (Rao et al., 2007; Valliappan, Sun & Li, 2014). Such molecules have been shown to inhibit settlement of algal spores, fungi and larvae and may contribute to the host’s defence against biofouling (Rao et al., 2007). As L. pygmaea is immersed for half the day, it may be more subject to biofouling than L. confinis and hence the biofilm communities covering the L. pygmaea may play an important role in the inhibition of attachment of undesirable organisms.
Despite the significantly different bacterial communities observed in the two vertically segregated Lichina species in the littoral belt, we did not find clear evidence for significant differences between the samples of each species across horizontal distances (minimum 6 m and maximum 37 m between clusters). This was indicated by beta-diversity analyses both on the whole or filtered dataset which included the most abundant OTUs and which revealed a lack of clustering of the replicates according to site. A lack of geographical influence on the lichen associated bacterial communities was also observed in other studies where sampling was carried out on a small spatial scale (Grube et al., 2009; Bates et al., 2011). In a study where lichen sampling was carried out at a much greater spatial scale, across the North American continent, a significant effect of geographical location on bacterial communities associated with a specific species was observed (Hodkinson et al., 2012).
Low culturability of marine lichen associated bacteria
Culturing of bacterial isolates from novel environments is essential for natural product discovery. The harsh environments, in which the Lichina lichen species are found, could provide a wealth of bioactive products, particularly from the potentially thermophilic/radiation resistant bacteria identified in our study. However, only 34 OTUs showed significant similarity to the 254 previously isolated cultured isolates (Parrot et al., 2015) even when lowering the similarity threshold to 97%. Moreover, three OTUs that were 100% identical to isolates were present at low abundances in the Illumina dataset. Whereas the Bacteroidetes phylum accounted for around 53% of the 16S rRNA sequences from the marine lichens, only four strains from this phylum were cultured. These results can be partly explained by the culturing strategy which was intentionally skewed towards the isolation of Actinobacteria by using Actinobacteria Isolation Agar (Parrot et al., 2015). As the bacterial isolates were obtained from lichens sampled 12–18 months prior to those that were analysed in this study (even though they were collected from the same site), we cannot rule out the influence of different sampling times on the bacterial composition. Nevertheless, the low culturability of bacteria from certain environments is well known (Amann, Ludwig & Schleifer, 1995) and our results further highlight the importance of parallel culture independent molecular methods to gain a more complete picture of the microbial diversity. Such results, combined with metagenomic approaches to target gene clusters of interest (Vester, Glaring & Stougaard, 2015) and novel isolation strategies incorporating lichen extracts into culture media (Biosca et al., 2016) should improve the bioprospecting of marine lichens. Nevertheless, the fact that some of the previously cultured strains were relatively close to the community described by our study, suggests that some of the bacteria associated with lichens might not be as recalcitrant to culture when compared to bacteria from other environments, highlighting the interest of these associations as sources of cultured microbial diversity.
Marine versus terrestrial lichens: challenging the paradigm of Alphaproteobacterial dominance
Here we show that bacterial communities in two strictly marine lichen species were completely different from those of two maritime and four inland terrestrial lichen species and that this difference is immediately visible at the phylum level with 50% of the sequences assigned to Bacteroidetes in Lichina, with a lower proportion of Proteobacteria (20–30%), the converse of the inland lichens from the humid wooded zone (15–19% Bacteroidetes and 50% Proteobacteria). At the OTU level, the differences were even more marked, with no overlap between the marine and inland terrestrial cyanolichens studied. The X. aureola communities (sampled above L. confinis on the rocky shore) only shared a single OTU with the Lichina lichens (Gammaproteobacteria OTU_15) which was only present at low abundance (0.05%). High abundances of Bacteroidetes were also observed in a different marine lichen Hydropunctaria maura (Bjelland et al., 2011) and supports the idea that this group of bacteria are major components of the bacterial communities associated with marine lichens, whereas Alphaproteobacteria are well confirmed by several studies as the dominant group associated with terrestrial lichens (Cardinale et al., 2008; Grube et al., 2009; Bates et al., 2011; Mushegian et al., 2011; Hodkinson et al., 2012). The marine and the maritime/inland terrestrial lichen mycobionts did belong to two different classes (Lichinomycetes and Lecanoromycetes respectively; see Table 1) so we cannot rule out that this significant mycobiont taxonomic separation could be partly responsible for the difference in bacterial community composition observed. Nevertheless, a survey of bacterial communities associated with different cyanolichens and chlorolichens showed that taxonomic dissimilarity of the mycobiont did not necessarily correlate with bacterial community dissimilarity (Hodkinson et al., 2012). In that study, although Dictyonema belongs to the division Basidiomycota while the other lichens belonged to the Ascomycota, the bacterial communities of Dictyonema were still composed of >50% Alphaproteobacteria distributed into the same four orders as for the majority of the other lichens. To be able to separate more clearly the influences of lichen taxonomy and habitat on the diversity of the associated bacterial symbionts, ideally, a terrestrial Lichina lichen species or another lichen belonging to the Lichinaceae family should be characterised in the future.
In marine environments Bacteroidetes are specialised in degrading polymeric organic matter and are adapted for an attached lifestyle by the production of adhesion proteins (Fernández-Gómez et al., 2013). Although Bacteroidetes are often reported attached to particles they are also known to be epiphytic on a variety of marine algae from temperate regions and often co-occur with Proteobacteria (Lachnit et al., 2011; Burke et al., 2011; Wahl et al., 2012; Miranda et al., 2013). Although high abundances of Alphaproteobacteria sequences were detected on the green alga Ulva australis from Botany Bay NSW Australia (Burke et al., 2011), Bacteroidetes was the most abundant group associated with the red alga Porphyra umbilicalis (Miranda et al., 2013) in Maine, US and with red and brown algae sampled during summer months in Kiel Bight, Germany (Lachnit et al., 2011). This suggests that Bacteroidetes may be important members of marine epiphytic and perhaps epilichenic communities, although this would need to be confirmed by microscopy-based (FISH) approaches.
Other differences between the marine and inland terrestrial lichens included the relative abundance of the bacterial (non-cyanobacterial) sequences compared to the cyanobacterial sequences that were as low as 20% in L. auriforme but accounted for 50–60% in the marine lichens (Fig. S1). The marine lichen H. maura also had a higher estimated concentration of bacterial cells in the lichen thallus compared to inland lichens (Bjelland et al., 2011). These observations suggest that bacteria associated with marine lichens might play a greater role in the functioning of the lichen symbiosis but this would need to be confirmed by microscopy analyses and for example, metabolite profiling.
Several roles of lichen-associated bacteria have been suggested and include nutrient supply (e.g., by nitrogen fixation and phosphate scavenging), nutrient recycling, protection against UV radiation, resistance to abiotic and biotic environmental stressors, and thallus degradation (Eymann et al., 2017). Unlike terrestrial lichens, marine lichens would need to be adapted to much more variable gradients of UV light, salinity (osmotic) stress, heat, desiccation and mechanical stresses from wave action and therefore we would expect the associated bacteria to play a role in resisting these particularly variable conditions. Although marine lichens can provide their own UV protection by the production of a parietin derivative in the case of Xanthoria and mycosporine-like amino acids in the case of the Lichina sp. (Roullier et al., 2011), it is possible that the bacteria that colonise the surface of the lichen like a biofilm (Cardinale et al., 2012b) could provide a protective layer conferred by other photoprotective pigments. The majority of the identified OTUs were affiliated to pigmented bacterial families, and several potentially phototrophic bacterial groups such as the aerobic anoxygenic phototropic (AAP; e.g., Chloroflexi, Erythrobacteraceae, Rhodobacteraceae) or rhodopsin-containing bacteria (e.g., Flavobacteriaceae, (Yoshizawa et al., 2014), Rhodothermaceae (Vavourakis et al., 2016), Deinococcus-Thermus) that contain carotenoids that may have a photoprotective role.
To alleviate salt stress and resist desiccation, marine lichens and their photobionts accumulate compatible solutes that are mainly sugars, sugar alcohols and complex sugars, lowering the water potential and hence reducing water movement out of the cells (Delmail et al., 2013). Indeed L. pygmaea which is immersed in seawater for half the day was shown to resist salt stress better than Xanthoria aureola, which is located higher up on the littoral belt and only receives sea spray (Delmail et al., 2013). Marine biofilms are considered to be analogous to a multicellular organism in that it functions like a “second skin”, replacing the hosts top layers as the new interface between the host and the environment (Wahl et al., 2012). Such a function could be imagined for the bacteria associated with L. pygmaea that could contribute to the osmotic adaptation of the lichen by maintaining a layer of cells in osmotic balance with the surrounding water. This could be achieved by different mechanisms including the synthesis and uptake of organic osmolytes and by using a sodium pumping rhodopsin as inferred from Bacteroidetes metagenomes recovered from hypersaline lakes (Vavourakis et al., 2016). While this is speculative, bacterial communities inhabiting the rhizosphere of coastal plants were shown to confer salt stress acclimatisation to the plants, potentially though the use of ion and compatible solute transporters (Yuan et al., 2016).
In cyanolichens or tripartite lichens, fixed carbon and nitrogen are supplied to the mycobiont by the photobiont whereas phosphate was shown to be a limiting macronutrient since cyanolichen growth was stimulated after immersing the thalli in a phosphate solution (McCune & Caldwell, 2009). This was also supported by metagenomic and proteomic analysis of two terrestrial lichens that attributed an important role of phosphate solubiliation genes to the associated microbial community, more specifically to the Alphaproteobacteria (Grube et al., 2015; Sigurbjornsdottir, Andresson & Vilhelmsson, 2015). The Lichina lichens may be less subject to phosphate limitation because L. pygmaea is immersed in seawater for half a day and L. confinis receives sea spray in a coastal area that is not particularly phosphorus limited. Nevertheless, some of the associated bacteria may play a role in phosphate uptake from the seawater or in the recycling and transfer of phosphate from senescing parts of the thallus to the growing apices as was shown for Cladonia (Hyvärinen & Crittenden, 2000). Such nutrient recycling may be carried out in part by the Bacteroidetes that are known to produce a wide range of enzymes capable of degrading polymeric organic matter including glycoside hydrolases and peptidases (Fernández-Gómez et al., 2013).
Conclusions
Here we show the striking differences between bacterial communities associated with marine and terrestrial lichens and even between marine lichens inhabiting different littoral zones. This represents a first piece in the puzzle in understanding the role of the associated bacteria in the marine lichen symbiosis, which is lagging much behind that of the terrestrial lichens. To begin to elucidate the functions of these communities, future work should include a multidisciplinary approach combining microscopy techniques to determine the spatial organisation of the core microbiome members and “omic” approaches to understand their functional role. An integrative study using metagenomics, metatranscriptomics and metabolomics could be performed on L. pygmaea during high and low tide to reveal a wealth of information on how the symbiotic partners interact and adapt to these extreme conditions. Moreover, such information would not only further our understanding of the marine lichen symbiosis but could also guide future bioprospecting efforts.
Supplemental Information
Supplementary information comprising supplementary figures and details of the data analysis pipeline.
Figure S1. Relative abundance (%) of bacterial and cyanobacterial sequences recovered from each lichen species. Xanthoria sp. Atl; from Brittany, and Xanthoria sp. Med. from the Mediterranean coast.
Figure S2. Bray-Curtis dissimilarity dendrogram using the filtered dataset (retaining high variance OTUs) described in the main text.
Figure S3. Comparison of relative abundance of core OTUs present in all replicates of two Xanthoria sp. sampled from the French Atlantic Ocean (Atl) and Mediterranean Sea (Med) coasts. OTUs are grouped at the family level when assigned or at the lowest taxonomic level assigned when family level was not available.
Figure S4. Distribution of the major groups of Cyanobacteria associated with the different lichen species with the groups presented at the lowest assignable taxonomic level.
List of lichens sampled with details of the different clusters, their geographical location and the temperature at the time of sampling.
Results of the blastn analyses of the sequenced marker genes used to check the morphological lichen identifications.
List of sample IDs for each lichen sample and their corresponding barcode used to tag the forward primer 341F.
List of the 30 most abundant OTUs for either the marine lichens or the maritime-inland terrestrial lichen group, presented together with their GreenGenes taxonomic identity and the closest relative obtained by blastn analysis.
List of lichen bacterial 16S rRNA OTUs and their taxonomy that showed >97% 16S rRNA gene sequence identity to bacterial isolates cultured from L. pygmaea, L. confinis and L. auriforme.
Acknowledgments
We thank the BIO2MAR platform (http://bio2mar.obs-banyuls.fr) for providing technical support and access to instrumentation. The following reagent was obtained through BEI Resources, NIAID, NIH as part of the Human Microbiome Project: Genomic DNA from Microbial Mock Community B (Even, Low Concentration), v5.1L, for 16S rRNA Gene Sequencing, HM-782D. We would also like to thank Dr. Nicolas Magain for his thorough and thoughtful review, particularly on the lichen taxonomy, which greatly helped to improve this paper.
Funding Statement
This study was funded by by the project MALICA (10-INBS-02-01) of the Agence Nationale de la Recherche, France and the INSA Rennes for the ministerial grant for the PhD of Delphine Parrot. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Additional Information and Declarations
Competing Interests
Marcelino T. Suzuki is an Academic Editor for PeerJ.
Author Contributions
Nyree J. West conceived and designed the experiments, performed the experiments, analyzed the data, contributed reagents/materials/analysis tools, prepared figures and/or tables, authored or reviewed drafts of the paper, approved the final draft.
Delphine Parrot conceived and designed the experiments, performed the experiments, contributed reagents/materials/analysis tools, prepared figures and/or tables, authored or reviewed drafts of the paper, approved the final draft.
Claire Fayet performed the experiments, authored or reviewed drafts of the paper, approved the final draft.
Martin Grube and Sophie Tomasi conceived and designed the experiments, performed the experiments, contributed reagents/materials/analysis tools, authored or reviewed drafts of the paper, approved the final draft.
Marcelino T. Suzuki conceived and designed the experiments, contributed reagents/materials/analysis tools, authored or reviewed drafts of the paper, approved the final draft.
DNA Deposition
The following information was supplied regarding the deposition of DNA sequences:
Sequence data is available at the European Nucleotide Archive under the study number: PRJEB23513.
Data Availability
The following information was supplied regarding data availability:
West, Nyree; Parrot, Delphine; Fayet, Claire; Grube, Martin; Tomasi, Sophie; Suzuki, Marcelino T. (2018): Lichen associated bacteria 16S rRNA Illumina sequences. figshare. Fileset. https://doi.org/10.6084/m9.figshare.5891509.v1.
References
- Albuquerque et al. (2005).Albuquerque L, Simões C, Nobre MF, Pino NM, Battista JR, Silva MT, Rainey FA, Da Costa MS. Truepera radiovictrix gen. nov., sp. nov., a new radiation resistant species and the proposal of Trueperaceae fam. nov. FEMS Microbiology Letters. 2005;247:161–169. doi: 10.1016/j.femsle.2005.05.002. [DOI] [PubMed] [Google Scholar]
- Altschul et al. (1997).Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Research. 1997;25:3389–3402. doi: 10.1093/nar/25.17.3389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amann, Ludwig & Schleifer (1995).Amann RI, Ludwig W, Schleifer KH. Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiological Reviews. 1995;59:143–169. doi: 10.1128/mr.59.1.143-169.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Apprill, Weber & Santoro (2016).Apprill A, Weber LG, Santoro AE. Distinguishing between microbial habitats unravels ecological complexity in coral microbiomes. mSystems. 2016;1:e00143-16. doi: 10.1128/mSystems.00143-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aschenbrenner et al. (2016).Aschenbrenner IA, Cernava T, Berg G, Grube M. Understanding microbial multi-species symbioses. Frontiers in Microbiology. 2016;7 doi: 10.3389/fmicb.2016.00180. Article 180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bates et al. (2011).Bates ST, Cropsey GWG, Caporaso JG, Knight R, Fierer N. Bacterial communities associated with the lichen symbiosis. Applied and Environmental Microbiology. 2011;77:1309–1314. doi: 10.1128/AEM.02257-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biosca et al. (2016).Biosca EG, Flores R, Santander RD, Díez-Gil JL, Barreno E. Innovative approaches using lichen enriched media to improve isolation and culturability of lichen associated bacteria. PLOS ONE. 2016;11:e0160328. doi: 10.1371/journal.pone.0160328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bjelland et al. (2011).Bjelland T, Grube M, Hoem S, Jorgensen SL, Daae FL, Thorseth IH, Ovreas L. Microbial metacommunities in the lichen-rock habitat. Environmental Microbiology Reports. 2011;3:434–442. doi: 10.1111/j.1758-2229.2010.00206.x. [DOI] [PubMed] [Google Scholar]
- Boustie & Grube (2005).Boustie J, Grube M. Lichens—a promising source of bioactive secondary metabolites. Plant Genetic Resources. 2005;3:273–287. doi: 10.1079/PGR200572. [DOI] [Google Scholar]
- Burke et al. (2011).Burke C, Thomas T, Lewis M, Steinberg P, Kjelleberg S. Composition, uniqueness and variability of the epiphytic bacterial community of the green alga Ulva australis. The ISME Journal. 2011;5:590–600. doi: 10.1038/ismej.2010.164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caporaso et al. (2010).Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, Fierer N, Peña AG, Goodrich JK, Gordon JI, Huttley GA, Kelley ST, Knights D, Koenig JE, Ley RE, Lozupone CA, McDonald D, Muegge BD, Pirrung M, Reeder J, Sevinsky JR, Turnbaugh PJ, Walters WA, Widmann J, Yatsunenko T, Zaneveld J, Knight R. QIIME allows analysis of high-throughput community sequencing data. Nature Methods. 2010;7:335–336. doi: 10.1038/nmeth.f.303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cardinale et al. (2008).Cardinale M, De Castro JV, Mueller H, Berg G, Grube M. In situ analysis of the bacterial community associated with the reindeer lichen Cladonia arbuscula reveals predominance of Alphaproteobacteria. FEMS Microbiology Ecology. 2008;66:63–71. doi: 10.1111/j.1574-6941.2008.00546.x. [DOI] [PubMed] [Google Scholar]
- Cardinale et al. (2012a).Cardinale M, Grube M, Castro JV, Mueller H, Berg G. Bacterial taxa associated with the lung lichen Lobaria pulmonaria are differentially shaped by geography and habitat. FEMS Microbiology Letters. 2012a;329:111–115. doi: 10.1111/j.1574-6968.2012.02508.x. [DOI] [PubMed] [Google Scholar]
- Cardinale, Puglia & Grube (2006).Cardinale M, Puglia AM, Grube M. Molecular analysis of lichen-associated bacterial communities. FEMS Microbiology Ecology. 2006;57:484–495. doi: 10.1111/j.1574-6941.2006.00133.x. [DOI] [PubMed] [Google Scholar]
- Cardinale et al. (2012b).Cardinale M, Steinova J, Rabensteiner J, Berg G, Grube M. Age, sun and substrate: triggers of bacterial communities in lichens. Environmental Microbiology Reports. 2012b;4:23–28. doi: 10.1111/j.1758-2229.2011.00272.x. [DOI] [PubMed] [Google Scholar]
- Chappuis et al. (2014).Chappuis E, Terradas M, Cefalì ME, Mariani S, Ballesteros E. Vertical zonation is the main distribution pattern of littoral assemblages on rocky shores at a regional scale. Estuarine, Coastal and Shelf Science. 2014;147:113–122. doi: 10.1016/j.ecss.2014.05.031. [DOI] [Google Scholar]
- Couradeau et al. (2017).Couradeau E, Roush D, Guida BS, Garcia-Pichel F. Diversity and mineral substrate preference in endolithic microbial communities from marine intertidal outcrops (Isla de Mona, Puerto Rico) Biogeosciences. 2017;14:311–324. doi: 10.5194/bg-14-311-2017. [DOI] [Google Scholar]
- Delmail et al. (2013).Delmail D, Grube M, Parrot D, Cook-Moreau J, Boustie J, Labrousse P, Tomasi S. Halotolerance in Lichens: symbiotic coalition against salt stress. In: Ahmad P, Azooz MM, Prasad MNV, editors. Ecophysiology and responses of plants under salt stress. Springer New York; 2013. pp. 115–148. [DOI] [Google Scholar]
- Edgar (2013).Edgar RC. UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nature Methods. 2013;10:996–998. doi: 10.1038/nmeth.2604. [DOI] [PubMed] [Google Scholar]
- Edgar & Flyvbjerg (2015).Edgar RC, Flyvbjerg H. Error filtering, pair assembly, and error correction for next-generation sequencing reads. Bioinformatics. 2015;31:3476–3482. doi: 10.1093/bioinformatics/btv401. [DOI] [PubMed] [Google Scholar]
- Eilers, Pernthaler & Amann (2000).Eilers H, Pernthaler J, Amann R. Succession of pelagic marine bacteria during enrichment: a close look at cultivation-induced shifts. Applied and Environmental Microbiology. 2000;66:4634–4640. doi: 10.1128/AEM.66.11.4634-4640.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eymann et al. (2017).Eymann C, Lassek C, Wegner U, Bernhardt J, Fritsch OA, Fuchs S, Otto A, Albrecht D, Schiefelbein U, Cernava T, Aschenbrenner I, Berg G, Grube M, Riedel K. Symbiotic interplay of fungi, algae, and bacteria within the lung lichen lobaria pulmonaria L. Hoffm. as assessed by state-of-the-art metaproteomics. Journal of Proteome Research. 2017;16:2160–2173. doi: 10.1021/acs.jproteome.6b00974. [DOI] [PubMed] [Google Scholar]
- Faircloth & Glenn (2012).Faircloth BC, Glenn TC. Not all sequence tags are created equal: designing and validating sequence identification tags robust to indels. PLOS ONE. 2012;7:e42543. doi: 10.1371/journal.pone.0042543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferguson, Buckley & Palumbo (1984).Ferguson RL, Buckley EN, Palumbo AV. Response of marine bacterioplankton to differential filtration and confinement. Applied and Environmental Microbiology. 1984;47:49–55. doi: 10.1128/aem.47.1.49-55.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernández-Gómez et al. (2013).Fernández-Gómez B, Richter M, Schüler M, Pinhassi J, Acinas SG, González JM, Pedrós-Alió C. Ecology of marine Bacteroidetes: a comparative genomics approach. The ISME Journal. 2013;7:1026–1037. doi: 10.1038/ismej.2012.169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferreira et al. (1999).Ferreira AC, Nobre MF, Moore E, Rainey FA, Battista JR, Costa MS da. Characterization and radiation resistance of new isolates of Rubrobacter radiotolerans and Rubrobacter xylanophilus. Extremophiles. 1999;3:235–238. doi: 10.1007/s007920050121. [DOI] [PubMed] [Google Scholar]
- Fletcher (1973a).Fletcher A. The ecology of marine (Littoral) Lichens on some rocky shores of Anglesey. The Lichenologist. 1973a;5:368–400. doi: 10.1017/S0024282973000459. [DOI] [Google Scholar]
- Fletcher (1973b).Fletcher A. The ecology of maritime (Supralittoral) Lichens on some rocky shores of Anglesey. The Lichenologist. 1973b;5:401–422. doi: 10.1017/S0024282973000460. [DOI] [Google Scholar]
- Grube et al. (2009).Grube M, Cardinale M, De Castro JV, Müller H, Berg G. Species-specific structural and functional diversity of bacterial communities in lichen symbioses. The ISME Journal. 2009;3:1105–1115. doi: 10.1038/ismej.2009.63. [DOI] [PubMed] [Google Scholar]
- Grube et al. (2015).Grube M, Cernava T, Soh J, Fuchs S, Aschenbrenner I, Lassek C, Wegner U, Becher D, Riedel K, Sensen CW, Berg G. Exploring functional contexts of symbiotic sustain within lichen-associated bacteria by comparative omics. The ISME Journal. 2015;9:412–424. doi: 10.1038/ismej.2014.138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hays et al. (2015).Hays SG, Patrick WG, Ziesack M, Oxman N, Silver PA. Better together: engineering and application of microbial symbioses. Current Opinion in Biotechnology. 2015;36:40–49. doi: 10.1016/j.copbio.2015.08.008. [DOI] [PubMed] [Google Scholar]
- Henkel & Yuzhakova (1936).Henkel PA, Yuzhakova L. Nitrogen-fixing bacteria in lichens. Izvestiya Biologicheskogo Nauchno-Issledovatel’skogo Instituta pri Permskom Gosudarstvennom Universitete. 1936;10:9–10. [Google Scholar]
- Hodkinson et al. (2012).Hodkinson BP, Gottel NR, Schadt CW, Lutzoni F. Photoautotrophic symbiont and geography are major factors affecting highly structured and diverse bacterial communities in the lichen microbiome. Environmental Microbiology. 2012;14:147–161. doi: 10.1111/j.1462-2920.2011.02560.x. [DOI] [PubMed] [Google Scholar]
- Honegger, Edwards & Axe (2013).Honegger R, Edwards D, Axe L. The earliest records of internally stratified cyanobacterial and algal lichens from the lower Devonian of the Welsh Borderland. The New Phytologist. 2013;197:264–275. doi: 10.1111/nph.12009. [DOI] [PubMed] [Google Scholar]
- Hyvärinen & Crittenden (2000).Hyvärinen M, Crittenden PD. 33P translocation in the thallus of the mat-forming lichen Cladonia portentosa. The New Phytologist. 2000;145:281–288. doi: 10.1046/j.1469-8137.2000.00581.x. [DOI] [Google Scholar]
- Kembel et al. (2010).Kembel SW, Cowan PD, Helmus MR, Cornwell WK, Morlon H, Ackerly DD, Blomberg SP, Webb CO. Picante: R tools for integrating phylogenies and ecology. Bioinformatics. 2010;26:1463–1464. doi: 10.1093/bioinformatics/btq166. [DOI] [PubMed] [Google Scholar]
- Klindworth et al. (2012).Klindworth A, Pruesse E, Schweer T, Peplies J, Quast C, Horn M, Glöckner FO. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Research. 2012;41:e1. doi: 10.1093/nar/gks808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kranner et al. (2008).Kranner I, Beckett R, Hochman A, Nash TH. Desiccation-tolerance in Lichens: a review. The Bryologist. 2008;111:576–593. doi: 10.1639/0007-2745-111.4.576. [DOI] [Google Scholar]
- Kurahashi et al. (2010).Kurahashi M, Fukunaga Y, Sakiyama Y, Harayama S, Yokota A. Euzebya tangerina gen. nov., sp. nov., a deeply branching marine actinobacterium isolated from the sea cucumber Holothuria edulis, and proposal of Euzebyaceae fam. nov. Euzebyales ord. nov. and Nitriliruptoridae subclassis nov. International Journal of Systematic and Evolutionary Microbiology. 2010;60:2314–2319. doi: 10.1099/ijs.0.016543-0. [DOI] [PubMed] [Google Scholar]
- Kurilenko et al. (2010).Kurilenko VV, Christen R, Zhukova NV, Kalinovskaya NI, Mikhailov VV, Crawford RJ, Ivanova EP. Granulosicoccus coccoides sp. nov., isolated from leaves of seagrass (Zostera marina) International Journal of Systematic and Evolutionary Microbiology. 2010;60:972–976. doi: 10.1099/ijs.0.013516-0. [DOI] [PubMed] [Google Scholar]
- Lachnit et al. (2011).Lachnit T, Meske D, Wahl M, Harder T, Schmitz R. Epibacterial community patterns on marine macroalgae are host-specific but temporally variable. Environmental Microbiology. 2011;13:655–665. doi: 10.1111/j.1462-2920.2010.02371.x. [DOI] [PubMed] [Google Scholar]
- Liba et al. (2006).Liba CM, Ferrara FIS, Manfio GP, Fantinatti-Garboggini F, Albuquerque RC, Pavan C, Ramos PL, Moreira-Filho CA, Barbosa HR. Nitrogen-fixing chemo-organotrophic bacteria isolated from cyanobacteria-deprived lichens and their ability to solubilize phosphate and to release amino acids and phytohormones. Journal of Applied Microbiology. 2006;101:1076–1086. doi: 10.1111/j.1365-2672.2006.03010.x. [DOI] [PubMed] [Google Scholar]
- Love, Huber & Anders (2014).Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biology. 2014;15 doi: 10.1186/s13059-014-0550-8. Article 550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCune & Caldwell (2009).McCune B, Caldwell BA. A single phosphorus treatment doubles growth of cyanobacterial lichen transplants. Ecology. 2009;90:567–570. doi: 10.1890/08-0344.1. [DOI] [PubMed] [Google Scholar]
- McMurdie & Holmes (2013).McMurdie PJ, Holmes S. phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLOS ONE. 2013;8:e61217. doi: 10.1371/journal.pone.0061217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller, Harley & Denny (2009).Miller LP, Harley CDG, Denny MW. The role of temperature and desiccation stress in limiting the local-scale distribution of the owl limpet, Lottia gigantea. Functional Ecology. 2009;23:756–767. doi: 10.1111/j.1365-2435.2009.01567.x. [DOI] [Google Scholar]
- Miranda et al. (2013).Miranda LN, Hutchison K, Grossman AR, Brawley SH. Diversity and abundance of the bacterial community of the red Macroalga Porphyra umbilicalis: did bacterial farmers produce Macroalgae? PLOS ONE. 2013;8:e58269. doi: 10.1371/journal.pone.0058269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mushegian et al. (2011).Mushegian AA, Peterson CN, Baker CCM, Pringle A. Bacterial diversity across individual Lichens. Applied and Environmental Microbiology. 2011;77:4249–4252. doi: 10.1128/AEM.02850-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Myllys, Lohtander & Tehler (2001).Myllys L, Lohtander K, Tehler A. β-Tubulin, ITS and group I intron sequences challenge the species pair concept in physcia aipolia and P. caesia. Mycologia. 2001;93:335–343. doi: 10.2307/3761655. [DOI] [Google Scholar]
- Oksanen et al. (2013).Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin P, O’Hara R, Simpson G, Solymos P, Stevens M, Wagner H. Vegan: community ecology package. R Package Version. 2.0-102013 CRAN.
- Ortiz-Álvarez et al. (2015).Ortiz-Álvarez R, De los Ríos A, Fernández-Mendoza F, Torralba-Burrial A, Pérez-Ortega S. Ecological specialization of two photobiont-specific maritime cyanolichen species of the genus Lichina. PLOS ONE. 2015;10:e0132718. doi: 10.1371/journal.pone.0132718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Otálora et al. (2010).Otálora MAG, Aragón G, Molina MC, Martínez I, Lutzoni F. Disentangling the Collema-Leptogium complex through a molecular phylogenetic study of the Collemataceae (Peltigerales, lichen-forming Ascomycota) Mycologia. 2010;102:279–290. doi: 10.3852/09-114. [DOI] [PubMed] [Google Scholar]
- Park et al. (2011).Park S, Yoshizawa S, Kogure K, Yokota A. Rubricoccus marinus gen. nov., sp. nov., of the family “Rhodothermaceae”, isolated from seawater. International Journal of Systematic and Evolutionary Microbiology. 2011;61:2069–2072. doi: 10.1099/ijs.0.026294-0. [DOI] [PubMed] [Google Scholar]
- Parrot et al. (2015).Parrot D, Antony-Babu S, Intertaglia L, Grube M, Tomasi S, Suzuki MT. Littoral lichens as a novel source of potentially bioactive Actinobacteria. Scientific Reports. 2015;5:15839. doi: 10.1038/srep15839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parrot et al. (2016).Parrot D, Legrave N, Delmail D, Grube M, Suzuki M, Tomasi S. Review—Lichen-associated bacteria as a hot spot of chemodiversity: focus on uncialamycin, a promising compound for future medicinal applications. Planta Medica. 2016;82:1143–1152. doi: 10.1055/s-0042-105571. [DOI] [PubMed] [Google Scholar]
- Pontarp et al. (2012).Pontarp M, Canbäck B, Tunlid A, Lundberg P. Phylogenetic analysis suggests that habitat filtering is structuring marine bacterial communities across the globe. Microbial Ecology. 2012;64:8–17. doi: 10.1007/s00248-011-0005-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- R Core Team (2015).R Core Team . R Foundation for Statistical Computing, 2014; Vienna: 2015. [Google Scholar]
- Rao et al. (2007).Rao D, Webb JS, Holmström C, Case R, Low A, Steinberg P, Kjelleberg S. Low densities of epiphytic bacteria from the Marine Alga Ulva australis inhibit settlement of fouling organisms. Applied and Environmental Microbiology. 2007;73:7844–7852. doi: 10.1128/AEM.01543-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schloss et al. (2009).Schloss PD, Westcott SL, Ryabin T, Hall JR, Hartmann M, Hollister EB, Lesniewski RA, Oakley BB, Parks DH, Robinson CJ, Sahl JW, Stres B, Thallinger GG, Van Horn DJ, Weber CF. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Applied and Environmental Microbiology. 2009;75:7537–7541. doi: 10.1128/AEM.01541-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider et al. (2013).Schneider D, Arp G, Reimer A, Reitner J, Daniel R. Phylogenetic analysis of a microbialite-forming microbial mat from a hypersaline lake of the Kiritimati Atoll, Central Pacific. PLOS ONE. 2013;8:e66662. doi: 10.1371/journal.pone.0066662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schultz (2017).Schultz M. Morphological and molecular data support Lichina intermedia as a distinct austral-marine species in the L. pygmaea group. The Lichenologist. 2017;49:321–332. doi: 10.1017/S0024282917000251. [DOI] [Google Scholar]
- Sigurbjornsdottir, Andresson & Vilhelmsson (2015).Sigurbjornsdottir MA, Andresson OS, Vilhelmsson O. Analysis of the Peltigera membranacea metagenome indicates that lichen-associated bacteria are involved in phosphate solubilization. Microbiology. 2015;161:989–996. doi: 10.1099/mic.0.000069. [DOI] [PubMed] [Google Scholar]
- Sigurbjornsdottir et al. (2014).Sigurbjornsdottir MA, Heidmarsson S, Jonsdottir AR, Vilhelmsson O. Novel bacteria associated with Arctic seashore lichens have potential roles in nutrient scavenging. Canadian Journal of Microbiology. 2014;60:307–317. doi: 10.1139/cjm-2013-0888. [DOI] [PubMed] [Google Scholar]
- Suzuki et al. (2016).Suzuki MT, Parrot D, Berg G, Grube M, Tomasi S. Lichens as natural sources of biotechnologically relevant bacteria. Applied Microbiology and Biotechnology. 2016;100:583–595. doi: 10.1007/s00253-015-7114-z. [DOI] [PubMed] [Google Scholar]
- Valliappan, Sun & Li (2014).Valliappan K, Sun W, Li Z-Y. Marine actinobacteria associated with marine organisms and their potentials in producing pharmaceutical natural products. Applied Microbiology and Biotechnology. 2014;98:7365–7377. doi: 10.1007/s00253-014-5954-6. [DOI] [PubMed] [Google Scholar]
- Vavourakis et al. (2016).Vavourakis CD, Ghai R, Rodriguez-Valera F, Sorokin DY, Tringe SG, Hugenholtz P, Muyzer G. Metagenomic insights into the uncultured diversity and physiology of microbes in four hypersaline soda lake brines. Frontiers in Microbiology. 2016;7 doi: 10.3389/fmicb.2016.00211. Article 211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vester, Glaring & Stougaard (2015).Vester JK, Glaring MA, Stougaard P. Improved cultivation and metagenomics as new tools for bioprospecting in cold environments. Extremophiles. 2015;19:17–29. doi: 10.1007/s00792-014-0704-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vilgalys & Hester (1990).Vilgalys R, Hester M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. Journal of Bacteriology. 1990;172:4238–4246. doi: 10.1128/jb.172.8.4238-4246.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wahl et al. (2012).Wahl M, Goecke F, Labes A, Dobretsov S, Weinberger F. The second skin: ecological role of epibiotic biofilms on marine organisms. Frontiers in Microbiology. 2012;3 doi: 10.3389/fmicb.2012.00292. Article 292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webb, Ackerly & Kembel (2008).Webb C, Ackerly D, Kembel S. Phylocom: software for the analysis of phylogenetic community structure and trait evolution. Bioinformatics. 2008;24:2098–2100. doi: 10.1093/bioinformatics/btn358. [DOI] [PubMed] [Google Scholar]
- Webb et al. (2002).Webb CO, Ackerly DD, McPeek MA, Donoghue MJ. Phylogenies and community ecology. Annual Review of Ecology and Systematics. 2002;33:475–505. doi: 10.1146/annurev.ecolsys.33.010802.150448. [DOI] [Google Scholar]
- White et al. (1990).White T, Bruns T, Lee S, Taylor J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis M, Gelfand D, Shinsky J, White T, editors. PCR protocols: a guide to methods and applications. Academic Press; London: 1990. pp. 315–322. [Google Scholar]
- Xia et al. (2008).Xia Y, Kong Y, Thomsen TR, Halkjær Nielsen P. Identification and ecophysiological characterization of epiphytic protein-hydrolyzing Saprospiraceae (“Candidatus Epiflobacter” spp.) in activated Sludge. Applied and Environmental Microbiology. 2008;74:2229–2238. doi: 10.1128/AEM.02502-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoon et al. (2012).Yoon J, Oku N, Park S, Katsuta A, Kasai H. Tunicatimonas pelagia gen. nov., sp. nov., a novel representative of the family Flammeovirgaceae isolated from a sea anemone by the differential growth screening method. Antonie Van Leeuwenhoek. 2012;101:133–140. doi: 10.1007/s10482-011-9626-6. [DOI] [PubMed] [Google Scholar]
- Yoshizawa et al. (2014).Yoshizawa S, Kumagai Y, Kim H, Ogura Y, Hayashi T, Iwasaki W, DeLong EF, Kogure K. Functional characterization of flavobacteria rhodopsins reveals a unique class of light-driven chloride pump in bacteria. Proceedings of the National Academy of Sciences of the United States of America. 2014;111:6732–6737. doi: 10.1073/pnas.1403051111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan et al. (2016).Yuan Z, Druzhinina IS, Labbé J, Redman R, Qin Y, Rodriguez R, Zhang C, Tuskan GA, Lin F. Specialized microbiome of a halophyte and its role in helping non-host plants to withstand salinity. Scientific Reports. 2016;6:32467. doi: 10.1038/srep32467. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary information comprising supplementary figures and details of the data analysis pipeline.
Figure S1. Relative abundance (%) of bacterial and cyanobacterial sequences recovered from each lichen species. Xanthoria sp. Atl; from Brittany, and Xanthoria sp. Med. from the Mediterranean coast.
Figure S2. Bray-Curtis dissimilarity dendrogram using the filtered dataset (retaining high variance OTUs) described in the main text.
Figure S3. Comparison of relative abundance of core OTUs present in all replicates of two Xanthoria sp. sampled from the French Atlantic Ocean (Atl) and Mediterranean Sea (Med) coasts. OTUs are grouped at the family level when assigned or at the lowest taxonomic level assigned when family level was not available.
Figure S4. Distribution of the major groups of Cyanobacteria associated with the different lichen species with the groups presented at the lowest assignable taxonomic level.
List of lichens sampled with details of the different clusters, their geographical location and the temperature at the time of sampling.
Results of the blastn analyses of the sequenced marker genes used to check the morphological lichen identifications.
List of sample IDs for each lichen sample and their corresponding barcode used to tag the forward primer 341F.
List of the 30 most abundant OTUs for either the marine lichens or the maritime-inland terrestrial lichen group, presented together with their GreenGenes taxonomic identity and the closest relative obtained by blastn analysis.
List of lichen bacterial 16S rRNA OTUs and their taxonomy that showed >97% 16S rRNA gene sequence identity to bacterial isolates cultured from L. pygmaea, L. confinis and L. auriforme.
Data Availability Statement
The following information was supplied regarding data availability:
West, Nyree; Parrot, Delphine; Fayet, Claire; Grube, Martin; Tomasi, Sophie; Suzuki, Marcelino T. (2018): Lichen associated bacteria 16S rRNA Illumina sequences. figshare. Fileset. https://doi.org/10.6084/m9.figshare.5891509.v1.