

Abstract
The olfactory organs of vertebrates are not only extraordinary chemosensory organs but also a powerful defense system against infection. Nasopharynx-associated lymphoid tissue (NALT) has been traditionally considered as the first line of defense against inhaled antigens in birds and mammals. Novel work in early vertebrates such as teleost fish has expanded our view of nasal immune systems, now recognized to fight both water-borne and air-borne pathogens reaching the olfactory epithelium. Like other mucosa-associated lymphoid tissues (MALT), NALT of birds and mammals is composed of organized lymphoid tissue (O-NALT) (i.e., tonsils) as well as a diffuse network of immune cells, known as diffuse NALT (D-NALT). In teleosts, only D-NALT is present and shares most of the canonical features of other teleost MALT. This review focuses on the evolution of NALT in vertebrates with an emphasis on the most recent findings in teleosts and lungfish. Whereas teleost are currently the most ancient group where NALT has been found, lungfish appear to be the earliest group to have evolved primitive O-NALT structures.
Keywords: Evolution, Nasal immunity, NALT, Mucosal immunity, Vertebrates, Teleost fish
1. Historical aspects
Tonsillectomy or the surgical removal of tonsils is a very ancient practice. Approximately 2000 years ago, Aulus Cornelius Celsus, a Roman writer and physician, described tonsil surgery by using his fingers to remove tonsils (Koempel et al., 2006; Younis and Lazar, 2002). Today, 530,000 children under the age of 15 have theirs tonsils or adenoids removed in the US every year and it is still one of the most common surgical procedures in children in this country (Cullen et al., 2009; Roland et al., 2011).
The first attempt to nasally vaccinate humans against smallpox was reported in the Golden Mirror of Medicine, Chinese medical text in 1742. Nasal vaccination was done by using powdered scabs that were blown to the nose or filling the nose with a vesicle smeared cotton (Plotkin, 2014). Thus, tonsillectomy and nasal vaccination precede our understanding of nasal immune systems.
Anatomically, the human Waldeyer’s ring was first described in 1884 by von Waldeyer-Hartz as a ring of lymphoid tissue in the pharyngeal wall (Cocquyt et al., 2005; Perry and Whyte, 1998). Nasopharynx-associated lymphoid tissue (NALT) was first described as a paired of lymphoid cells accumulations in the nasal passage of rat in 1947 (Kelemen, 1947) whilst the mouse NALT was first described few decades later (Belal et al., 1977). In the subsequent years, the NALT of other mammals such as monkeys (Harkema et al., 1987; Loo and Chin, 1974) and horses (Mair et al., 1987, 1988) were described. However these studies did not include functional aspects of the nasal immune system (Kuper et al., 1992).
One of the major breakthroughs in nasal immunity field took place in the early 2000, when the first nasal vaccine for use in humans against influenza virus was licensed in the USA (FluMist) (Chen et al., 2006). To date, this vaccine remains the only nasal vaccine licensed for human use. The effectiveness and availability of this vaccine has helped the NALT community to expand basic scientific knowledge on nasal immune responses. Intranasal vaccination offers a number of advantages over other vaccination routes (Neutra and Kozlowski, 2006). Apart from the fact that is needle free and requires small amounts of antigen, intranasal delivery has been shown to stimulate not only local nasal immunity but also systemic immune responses as well as mucosal immune responses in distant mucosal sites (Fukuyama et al., 2012; Lycke, 2012; Neutra and Kozlowski, 2006; Pabst, 2015).
2. Anatomy of NALT
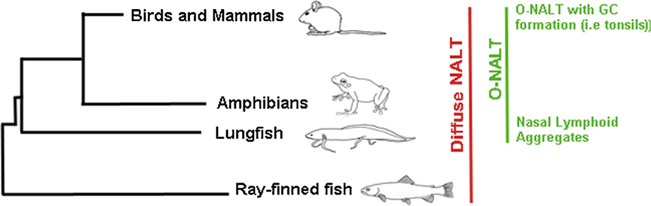
In endotherms, mucosa-associated lymphoid tissues (MALT) comprise a network of secondary lymphoid tissues that contain both well-organized lymphoid structures (organized MALT, O-MALT) and scattered or disseminated lymphoid cells (diffuse MALT, D-MALT) (Brandtzaeg and Pabst, 2004). Examples of O-MALT include the Peyer’s patches in the gut or the tonsils in the nasopharyngeal cavity. Generally speaking we know very little about NALT (both organized and diffuse) compared to the gut-associated lymphoid tissue (GALT). Furthermore, most mammalian studies focus on organized NALT (O-NALT), whereas diffuse NALT (D-NALT) has received very little attention. It is worth noting that the term “non-NALT” can also be found in the literature instead of D-NALT (Asanuma et al., 1997; Lee et al., 2015; Tamura et al., 1998). Additionally, “nasal passage (NP) leucocytes” is another term that can be found in the mammalian literature referring to D-NALT (Hiroi et al., 1998; Rodríguez-Monroy et al., 2007; Shikina et al., 2004). Despite the fact that the Society for mucosal immunology does not recommend the use of O-MALT and D-MALT (Brandtzaeg et al., 2008), numerous mammalian immunologists continue to use this terminology. Moreover, from the evolutionary immunologist point of view, O-NALT and D-NALT are useful terms considering that early vertebrates lack O-NALT (Fig. 1). In the next part of this review, we will summarize the basic anatomical aspects of NALT in different vertebrate groups.
Fig. 1.
Schematic diagram of the evolution of nasal immune systems in vertebrates.
2.1. Rodents and humans
The anatomy and structure of O-NALT have been widely studied in rodents (Mestecky et al., 2005). Murine NALT is composed of a pair of organized mucosal organs located on the roof of the soft palate at the entrance to the nasopharyngeal duct (Liang et al., 2001). NALT in mouse is considered by some to be analogous to the Waldeyer’s ring in human which consists of the adenoids and tonsils (Kuper et al., 1992). Thus, in both mice and humans, O-NALT is strategically placed in the upper airways to combat air-borne antigens. Perhaps, this anatomical observation historically hindered the investigation of NALT in non-terrestrial vertebrates.
It is important to highlight that O-NALT of rodents and humans appears to be significantly different anatomically speaking. In rodents, NALT is located in a single localization bilateral at the entrance of the nasopharyngeal duct whereas in human studies conducted in children, O-NALT was mostly found in the middle concha and it consisted of disseminated lymphoid, subepithelial follicles (Debertin et al., 2003). The results from this study indicated that children have O-NALT structures in addition to a Waldeyer’s ring (Debertin et al., 2003). Additionally, these differences under-score the fact that mice may not be the best models for human nasal immunity studies.
Similar to the Peyer’s patches in the gut, O-NALT structures are located underlying specialized portions of the epithelium known as follicle-associated epithelium (FAE). Additionally, high endothelial venules (HEVs) control lymphocyte trafficking into O-NALT (Kiyono and Fukuyama, 2004). O-NALT structures also have distinct B-lymphocyte and T-cell zones (Bailey et al., 2005; Brandtzaeg and Pabst, 2004). Germinal center formation occurs in O-NALT in response to infection or antigenic stimulation (Zuercher et al., 2002).
As mentioned earlier, both mice and humans also possess diffuse lymphoid cells situated on the mucosa of the nasal passages called (D-NALT) (Liang et al., 2001). D-NALT includes myeloid cells and lymphoid cells (both B and T cells). The similarities and differences between mammalian O-NALT and D-NALT are summarized in Table 1.
Table 1.
The differences between mice O-NALT and D-NALT.
| O-NALT | D-NALT | References | |
|---|---|---|---|
| Number of lymphocytes | + | ++ | Rodríguez-Monroy et al. (2007) |
| T/B cells ratio | 0.76–1.2 | 0.33–1.0 | Asanuma et al. (1997), Rodríguez-Monroy et al. (2007) |
| Percentage of B cellsa | 47–79% | 55–74% | Liang et al. (2001), Rodríguez-Monroy et al. (2007) |
| Plasma cells | + | ++ | Rodríguez-Monroy et al., (2007) |
| IgM+ B cells | 47 or 85% | 0.5–9% | Rodríguez-Monroy et al. (2007), Shikina et al. (2004), Smith et al. (2013) |
| IgA+ B cells | 1–1.6% | 7.7–10.8% | |
| B220+hi B cells | Present | Present | Rodríguez-Monroy et al. (2007) |
| B220+hi B220+low B cells | Absent | Present | |
| IgM/IgG/IgA secreting cells ratio (Uninfected) | 10/3/3 | 10/3/3 | Asanuma et al. (1997) |
| IgM/IgG/IgA secreting cells ratio (infected with influenza)b | 4/50/40 | 1/25/10 | |
| IgM/IgG/IgA secreting cells ratio(immunized with Cry1Ac protoxin)c | 1/80/125 | 1/125/350 | Rodriguez-Monroy and Moreno-Fierros (2010) |
| IgM/IgG/IgA secreting cells ratio(immunized with cholera toxin) | 1/30/65 | 1/90/160 | |
| IgA isotype class switching | Present | Absent | Shikina et al. (2004) |
| Class switch recombination-associated molecules | Present | Absent | |
| Long-lasting, specific effector antibody responsee | Absent | Present | Liang et al. (2001) |
| Frequency of AFCse | + | ++ | Liang et al. (2001) |
| Generation of virus-specific antibody forming cells (AFCs)e | + | + | |
| Infected/uninfected IFN-γ production ratiod | ~1000 | ~700 | Asanuma et al. (1997) |
| Infected/uninfected IL4 production ratio | 1 | 2 | |
| CD3+ T cells | 30–40% | 13–20% | Rodríguez-Monroy et al. (2007), Smith et al.(2013) |
| CD4+/CD8+ T cells ratio | 3–4.4 | 1.5–6.4 | Asanuma et al. (1997), Heritage et al. (1997), Rodríguez-Monroy et al. (2007), Smith et al.(2013) |
| αβ /γδ of CD3+ T cells ratio | 49–100 | 2.5–3 | Heritage et al. (1997) |
| αβ /γδ of CD4+ CD8- T cells ratio | 49–100 | 49–100 | |
| αβ /γδ ofCD4- CD8+ T cells ratio | 49–100 | 9–19 | |
| αβ /γδ ofCD4- CD8- T cells ratio | 0–1 | 0–1 | |
| Type of CD4+ T cells | Th0 | Th2 | Hiroi et al.(1998), Liang et al. (2001) |
Summary of major immune characteristics of mammalian organized and diffuse NALT in mice: ++ = high, + = low.
C57BL/6 mice have higher B cells percentage compared to BALB/c mice.
7 days post infection (dpi) with influenza.
BALB/c mice immunized intranasally.
7 dpi with influenza.
shows responses after influenza virus infection.
2.2. Other mammals
In this section, we are going to focus on reports pertaining four groups of mammals: cattle, sheep, canines and rabbits. The studies in these species have been motivated by the importance of nasal vaccination in veterinary medicine.
In cattle, the tonsil (O-NALT) was first described in 1992 (Schuh and Oliphant, 1992). Cattle tonsils are located at the entry of the pharynx and are equivalent to the Waldeyer’s ring in humans (Rebelatto et al., 2000). During development, adenoid can be detected at 95 days of gestation. Moreover, ciliated, microvillus cells and a loose accumulation of mononuclear cells in lamina propia is visible at 120–150 days of gestation. Small lymphoid follicles form at 4–5 months of gestation following by the appearance of goblet cells after 5 months of gestation (Schuh and Oliphant, 1992). Tonsils are well developed at birth in cattle. However, germinal center formation and increase in MHC class II expression only occur in the late natal and early post-natal period (Schuh and Oliphant, 1992). Moreover, cattle infected with foot-and-mouth disease virus (FMDV) showed increased expression of TLR-4 in NALT, indicating the importance of type I IFN responses in NALT against FMDV (Zhang et al., 2006).
Sheep O-NALT structures, similar to horse O-NALT, are clustered posterior to the opening of the Eustachian tube (Mair et al., 1988; Stanley et al., 2001). Thus, ovine NALT is highly organized and consists of discrete B and T cells areas similar to those found of humans and rodents. Furthermore, it has been shown that sheep NALT is covered by ciliated and non-ciliated cells which play an important role in antigen uptaking and processing (Stanley et al., 2001).
Peeters et al. reported the absence of typical O-NALT structures in the nasal mucosa of dogs without respiratory disease (Peeters et al., 2005). Waldeyer’s ring in the dog consists of the lingual tonsil, the palatine tonsils, the soft palate tonsil and the pharyngeal tonsil or adenoid (Billen et al., 2006). The nasopharyngeal mucosa in dogs appears uniform and the nasopharyngeal tonsil is not obvious (Billen et al., 2006). The latter might be due to the fact that dogs breathe through both the nose and the mouth; therefore, exposure of the canine nasal and nasopharyngeal mucosa to inhaled antigens is decreased. This may explain why the pharyngeal tonsil is less developed in dogs than horses, cattle, sheep and pigs (Billen et al., 2006).
Casteleyn et al. (2010) histologically examined the presence of NALT in rabbits by sectioning the nasal cavity. Rabbits appear to have well organized NALT in their nasal cavities including clustered I and II lymphoid follicles separated by interfollicular regions as well as isolated lymphoid follicles. Interestingly, in the middle third of rabbit nasal cavity, NALT occupied the largest space. The rabbit and human nasal cavities occupy a similar volume considering their respective body masses. However, in comparison with rodents, O-NALT in the rabbit is more abundant. Therefore the similarities between human and rabbit nasal cavities suggest that the rabbit is a better model for intranasal vaccine development than rodents are (Casteleyn et al., 2010). Additionally, rabbits have a D-NALT characterized by intraepithelial and lamina propria lymphocytes (Casteleyn et al., 2010).
2.3. Birds
Most of our knowledge on avian NALT comes from studies in chicken and duck. The nasal cavity in chickens is cone-shaped and separated into the right and left sides by a cartilaginiform nasal septum. The majority of the nasal cavity is occupied by the turbinates which play a major role in preventing the entry of dust and microbes (Kang et al., 2013). Lymphoid nodules are the main O-NALT structure in chickens. Nodules are made of B cells with developed germinal centers, surrounded by a coat of CD4+ T cells.
Chicken D-NALT, on the other hand, consists mostly of CD8+ T cells that can be found in the epithelium and in the lamina propria of the nasal mucosa (Kang et al., 2013; Ohshima and Hiramatsu, 2000). Additionally, scattered lymphoid cells are found in the paranasal organs of chickens (nasolacrimal ducts, lateral nasal glands and their ducts) (Bang and Bang, 1968; Kang et al., 2013).
NALT appears to be an important inductive site for the chicken’s mucosal immune system, however, the low absorption rates of antigen by the nasal mucosa may limit induction of effective nasal immune responses (Kang et al., 2013).
The nasal cavity in duck is cone-shaped and separated into right and left sides by a nasal septum (Kang et al., 2014). At the caudal regions of the nasal cavity there are two pairs of symmetrical lymphoid aggregates; one pair located on the dorsal side of the choanal cleft and the other pair on both sides of nasal septum (Kang et al., 2014). The FAE found in duck O-NALT, unlike that of rodents, contains almost no goblet cells. The use of liquid vaccines for ducklings and dry powder vaccine sprays for adult ducks has been recommended because liquid vaccines are unlikely to reach the NALT located on the nasal septum in adult ducks (Kang et al., 2014).
Like the chicken, ducks have diffuse lymphoid tissue and intraepithelial lymphocytes located within their nasal walls and turbinalia randomly (Kang et al., 2014).
2.4. Reptiles and amphibians
Unfortunately, NALT has been studied neither in reptiles nor in amphibians. This is despite the fact that chelonids are affected by upper respiratory tract disease (URTD) caused by Mycoplasma agassizii (Brown et al., 1999). The infection results in nasal discharge, nasal wheeze, conjunctivitis and inflammation of the oral mucosa. The contribution of NALT responses in M. agassizii infected chelonids should be investigated
Amphibians are known to have both diffuse and organized GALT and NALT (Goldstine et al., 1975). Tonsil-like structures were found in the lingual and sublingual regions of anuran amphibians (Myers, 1928). We further identified lymphoid aggregates associated with the olfactory epithelium of Lithobates sp. Tadpoles (Tacchi et al., 2015). Based on histological examination, these were present immediately underneath the olfactory epithelium but did not appear as organized as those found in lungfish. Contrary to birds and mammals, amphibian O-MALT is characterized by lymphoid nodules or aggregates that lack B and T cell zones or GC formation (Goldstine et al., 1975).
2.5. Sarcopterygian fishes
Sarcopterygian fish such as lungfish are the closest living sister species to all tetrapods (Zardoya and Meyer, 1996). Moreover, lungfish represent the transition from aquatic to terrestrial environments during vertebrate evolution and are exposed to both air-borne and water-borne antigens. We recently discovered that African lungfish (Protopterus sp.) may be the most ancient vertebrate where O-MALT, including O-NALT, is found (Fig. 1). O-NALT structures in lungfish are FAE-associated or embedded in the mucosa of the upper and lower jaw (Fig. 2). They are mostly composed of lymphocytes both B and T cells and do not display GC formation (Tacchi et al., 2015). In response to bacterial infection, lungfish O-NALT architecture and cellular composition changes, with an increase in the percentage of T and B cells compared to uninfected controls. Although not covered in the same study, we anticipate the presence of D-NALT in lungfish, but their characteristics need to be investigated. The discovery of these primordial forms of O-NALT in lungfish along with the ability to infect these animals intranasally in the laboratory makes this species an interesting system for investigating the structure, organogenesis and function of O-MALT in ectotherms.
Fig. 2.
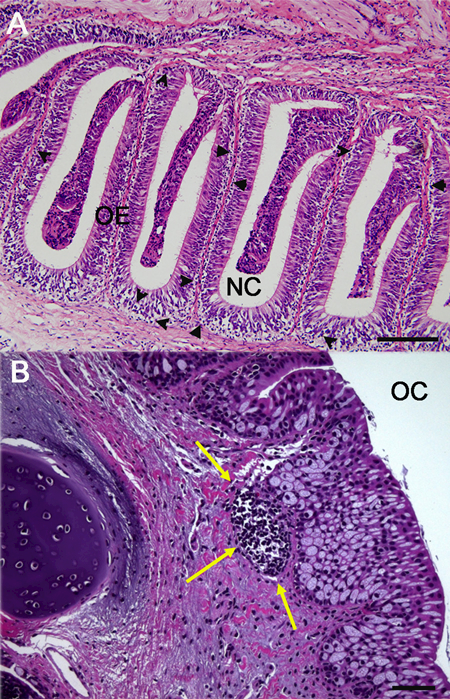
(A) Light micrograph (hematoxylin and eosin stain) of paraffin sections of the olfactory organ of an Australian eel (Anguila australis) showing the presence of diffuse NALT in the form of scattered lymphocytes (black arrow heads). (B) Light micrograph (hematoxylin and eosin stain) of the olfactory organ of the African lung-fish (Protopterus dolloi) showing the presence of a primordial lymphoid aggregate (yellow arrows). NC: nasal cavity; OC: oral cavity; OE: olfactory epithelium. Scale bar= 100 μm.
2.6. Teleosts
Until recently, three MALT have been characterized in teleost fish: gut-associated lymphoid tissue (GALT), skin-associated lymphoid tissue (SALT) and gill-associated lymphoid tissue (Salinas et al., 2011). However, we have discovered that NALT is likely conserved in all jawed vertebrates as it was found in teleost fish. Teleost NALT resembles other teleost MALT in many regards such as the presence of diffuse lymphoid cells with no organized structures present, a predominant constitutive presence of IgT+ B cells and secreted IgT compared to systemic lymphoid tissues and the presence of an associated microbiota coated by mucosal immunoglobulins (Tacchi et al., 2014). This discovery broke the previous paradigm of nasal immune systems being a first line of defense against inhaled antigens.
Histological examination of the NALT of four different families of teleost fish showed that teleost NALT harbors diffuse lymphoid cells but lacks O-NALT (Tacchi et al., 2014). One of the key features of teleost MALT is the preponderant presence of IgT+ B cells and secreted IgT into mucosal secretions (Zhang et al., 2010). Trout D-NALT contains abundant B cells, 48.5% are IgM+ B cells and 51.5% are IgT+ B cells, following the canonical B cell composition of other trout MALT. Similarly, the ratio of secreted IgT and secreted IgM in nasal mucus is much higher than that in plasma, again highlighting that trout NALT shares the main features of other MALT in teleosts. In gut and skin, previous studies have shown that specific IgT but not IgM responses occur in the local mucosal environment in response to parasitic infections in rainbow trout (Xu et al., 2013; Zhang et al., 2010). Although we detected high levels of IgT in the nasal secretion of trout in the absence of antigenic stimulation, specific IgT responses in the olfactory organ of teleosts in response to infection or vaccination are yet to be investigated. We have demonstrated striking protection levels conferred by nasal vaccines against viral and bacterial pathogens of fish in rainbow trout however, the mechanisms underlying the observed protection are not well understood and may include both humoral and cellular immunity (LaPatra et al., 2015; Tacchi et al., 2014).
Using a live attenuated infectious virus (IHNV) model of nasal vaccination, we were able to show that trout NALT mounts strong innate and adaptive immune responses over the course of a 28 day experiment. Whereas the expression of innate immune genes peaked at day 4 post vaccination, expression of adaptive immune genes peaked at day 14. Additionally, nasal vaccination resulted in stimulation of immune genes not only in NALT but also in the head-kidney (Tacchi et al., 2014).
Altogether, our results clearly reveal that early bony vertebrates evolved olfactory organs with strong defense functions that serve as a primary mucosal immune barrier against pathogens. Moreover, the discovery of teleost NALT and detection of nasal immune responses in trout in response to vaccination underscores the fact that strong nasal immune responses in vertebrates can take place in the absence of O-NALT. Currently, surgical ablation of O-NALT in mice is used as a tool to evaluate the specific contribution of O-NALT to mammalian nasal immune responses. This method is complex and may result in incomplete removal of all O-NALT cells. Thus, we propose that teleost NALT is an excellent model for the study of D-NALT and can provide a unique tool to understand, for instance, how tonsillectomized humans respond to nasal vaccines.
3. Evolution of NALT-specific molecules in vertebrates
The mucosal immune system has evolved complex suites of immune molecules as well as cell adhesion and trafficking molecules that separate it from the systemic immune system. For instance, trafficking of lymphocytes to NALT in mice is governed by the PNad-L-selectin pair (Csencsits et al., 1999; Fukuyama et al., 2002), whereas the α4β7-MAdCAM-1 pair governs trafficking of lymphocytes to the gut (Berlin et al., 1995).
The development of lymphoid tissues mainly relies on the lymphotoxin-driven expression of homeostatic chemokines such as CXCL13 (Ansel et al., 2000; Randall and Mebius, 2014). CXCL13 also plays an important role in maintaining the lymphoid architecture of NALT such as formation of germinal centers (Rangel-Moreno et al., 2005). In NALT, however, additional chemokines like CCL20 and CCL9 which are important in dendritic cell and B cell migration are required for development (Zhao et al., 2003). Impaired expression of CCL19/CCL21 results in defects in influenza-specific CD8 T cells responses in NALT of lymphotoxin-α-deficient (Ltα−/−) and plt/plt mice (Rangel-Moreno et al., 2005).
We know very little about the role of chemokines in the nasal immune system of vertebrates other than mice and humans. In trout, we found CCL19 expression was upregulated around 50-fold at 4 days post vaccination with live attenuated IHNV suggesting a conserved role of this chemokine in vertebrate nasal immunity (Tacchi et al., 2014).
With respect to O-MALT evolution, it has been proposed that organization of lymphocytes in structures such as lymph nodes or O-MALT is an endotherm-restricted innovation. Lymphocyte organization requires the presence of certain molecular signals that create the adequate microenvironment for B and T cells. Most of the molecules that play a role in lymphocyte organization belong to the tumor necrosis factor superfamily (TNFSF) (Tacchi et al., 2015). In this respect, we recently provided new insights into the evolution of TNFSF and showed that it greatly diversified in African lungfish. Among TNFSF members, lymphotoxins are of particular relevance due to their function in the organization of lymphoid tissues. Interestingly, where lymphotoxin beta receptor (LTBR) is found from teleosts to humans, lymphotoxin alpha (LTA) and beta (LTB) are absent in teleosts but found in lungfish. In other words, TNFSF members known to be essential in lymphocyte organization in mammals first appeared in lungfish and are absent from teleost genomes. Moreover, a number of these TNFSF members were found to be expressed in African lungfish O-NALT (Tacchi et al., 2015).
ID2 is a transcription factor known to be essential for O-NALT formation. ID2 is involved in the induction of CD3-CD4+CD45+ cells (Yokota et al., 1999) and ID2 deficient mice could not develop NALT after birth (Fukuyama et al., 2002; Yokota et al., 1999). We found ID2 expressed both in trout and lungfish NALT and its expression was significantly modulated upon nasal vaccination with a viral vaccine or nasal bacterial infection, respectively (Tacchi et al., 2014). Important molecules in the formation of NALT in mice are summarized in Table 2.
Table 2.
Requirements for the formation of NALT in mice.
| KO mice with | Organogenesis of NALT | References |
|---|---|---|
| Id2 | Absent | Fukuyama et al. (2002), Yokota et al. (1999) |
| Ror-γt | Not required/well developed | Harmsen et al. (2002), Sun et al. (2000) |
| Ltα | Disorganized but developed with fewer number of lymphocytes | De Togni et al. (1994), Fukuyama et al. (2002), Harmsen et al. (2002) |
| aly/aly | Disorganized but developed with fewer number of lymphocytes | Fukuyama et al. (2002), Miyawaki et al. (1994) |
| IL-7rα | Disorganized but developed with decreased number of lymphocytes | Adachi et al.(1998), Fukuyama et al. (2002), Harmsen et al. (2002) |
| plt/plt | Developed with decreased number of lymphocytes | Fukuyama et al. (2006), Rangel-Moreno et al. (2005) |
| Microbiota | Developed with delay | Randall and Mebius (2014) |
| LTβ | Developed but disorganized | Fukuyama et al. (2002), Koni et al. (1997) |
| TRANCE | Developed but disorganized | Ache and Young (2005), Harmsen et al. (2002) |
| CXCL13/CCL19/CCL21 | Developed but disorganized | Ansel et al. (2000), Fukuyama et al. (2006), Rangel-Moreno et al. (2005) |
4. Nasal microbiota and immunity in vertebrates
Every animal mucosal surface is colonized by millions of microorganisms forming a very ancient and successful symbiosis between prokaryotes and metazoans. The microbiota controls the immunological development of the host by different mechanisms such as inhibition of pathogen colonization on mucosal surfaces and stimulation of the host immune system (Buffie and Pamer, 2013; Kiyono and Fukuyama, 2004; Randall and Mebius, 2014). Although the number of studies revealing the importance of the microbiota in GALT development and function is vast, information regarding its effects on NALT is limited.
The nasal microbiome of human was sequenced and characterized in 2010 and further studies continue to determine its role in health and disease (Dewhirst et al., 2010). In humans, over 96% of the nasal bacterial microbiome belongs to three phyla: Actinobacteria, Firmicutes and Proteobacteria (Yan et al., 2013). In other mammalian species such as pigs, the nasal microbiome is Proteobacteria, Firmicutes and Spirochaetes (Weese et al., 2014). Nasal-associated microbiota are also present in aquatic vertebrates as evidenced by our studies in rainbow trout (Tacchi et al., 2014). The bacterial community associated with the trout olfactory organ is very diverse. 16s rDNA pyrosequencing revealed the presence of 18 total bacterial phyla, the highest number of phyla present among all body sites. The bacterial community was dominated by Proteobacteria, Actinobacteria, Bacteriodetes and Firmicutes with the class Betaproteobacteria accounting for 15.1–53.6% of all sequences (Lowrey et al., 2015). Thus, although limited pyrosequencing data is available, it appears that Proteobacteria and Firmicutes may be the most dominant and conserved taxa among nasal microbiomes in vertebrates, further studies will confirm or reject this hypothesis. The trout nasal microbiome composition is distinct from that of any other trout mucosal surface but most closely resembles that of the skin.
The presence of unique bacterial communities in association with each mucosal anatomical site suggests a tight cross-talk between the microbiota and MALT in vertebrates. One of the ways by which the microbiota and the host immune system interact is by means of secretory immunoglobulins that help anchor microorganisms to the mucus layer, while tagging them in a process known as immune exclusion. As an example, trout nasal-associated bacteria, similar to gut and skin bacteria, are coated by secretory immunoglobulins. Particularly, in trout skin and gut mucus, the immunoglobulin isotype IgT has specialized in the role of bacteria coating (Xu et al., 2013; Zhang et al., 2010) with IgM coating significantly lower numbers of bacteria. In nasal mucus, however, an equal proportion of bacteria were coated with IgM or IgT and the proportion of bacteria coated with both isotypes was higher than that found in the gut and skin (Tacchi et al., 2014).
From mammalian studies it appears clear that the microbiota plays a key role in the form and function of NALT. For instance, the microbiota was shown to modulate local nasal T cell responses against Mycoplasma pulmonis infection (Henriksson et al., 2004). On the other hand, the composition of the nasal microbiome of humans is a determinant of Staphylococcus aureus colonization and carriage (Frank et al., 2010; Yan et al., 2013) and, in mice, it regulates the immune responses against respiratory influenza A virus infection (Ichinohe et al., 2011). Additionally, it has been shown that the number of M cells in NALT of specific pathogen free rats is lower compared to normally reared rats (Jeong et al., 2000) and the numbers of T and B cells in NALT are 2–3 times greater after an experimental infection (Asanuma et al., 1997). NALT, like most of the lymphoid organs, can develop in germ free mice. NALT development CXCL5-deficient mice in sterile conditions takes about 20 weeks, however by repeated intranasal application of heat inactivated Propionibacterium acnes suspension, NALT can be induced after 8 weeks (Krege et al., 2009). In the future, the specific roles of the nasal microbiota on the development and function of vertebrate NALT should be further elucidated.
5. How old is NALT?
Our current view of nasal immunity is limited and mostly restricted to mammalian studies. We provided the first evidence supporting the idea that nasal immunity is an ancient arm of the mucosal immune system of vertebrates. Furthermore, our studies underscore that D-NALT, as anticipated, precedes the appearance of O-NALT during evolution (Fig. 1). Currently, we can state that D-NALT is at least 380 MY old, although we anticipate the cartilaginous fish and even agnathans may be equipped with an equivalent immune system associated with their olfactory organs. How old is O-NALT is a separate question. We have provided evidence of primitive O-NALT structures in the African lungfish, indicating that the ancestor to all tetrapods first acquired this immunological innovation (Tacchi et al., 2015). Further studies should address the immune function of primitive O-NALT in ectotherms.
6. Concluding remarks
Olfaction is one of the most ancient and conserved sensory systems in vertebrates. It seems that the successful structure and function of olfactory organs evolved to not only provide the exquisite detection of odorants but also to ensure that pathogens do not gain entry into hosts via this route. We hope that our pioneer studies in NALT evolution motivate more studies in non-mammalian models that complete our understanding of nasal immune systems and their evolution. Future studies in teleosts may address whether specific IgT is the main immunoglobulin isotype in nasal adaptive immune responses and whether nasal vaccination can lead to protection in other distant MALT. Lungfish studies, on the other hand, should try to address what evolutionary advantages lymphocyte organization confers at mucosal sites in the absence of germinal center formation.
Acknowledgments
This work was funded by awards NSF IOS # 1456940 and USDA AFRI # 2DN70–2RDN7 to IS. Authors thank Victoria Hansen for the artwork.
References
- Ache BW, Young JM, 2005. Olfaction: diverse species, conserved principles. Neuron 48, 417–430. [DOI] [PubMed] [Google Scholar]
- Adachi S, Yoshida H, Honda K, Maki K, Saijo K, Ikuta K, Saito T, Nishikawa S-I, 1998. Essentia role of IL-7 receptor alpha in the formation of Peyer’s patch anlage. Int. Immunol 10 (1), 1–6. [DOI] [PubMed] [Google Scholar]
- Ansel KM, Ngo VN, Hyman PL, Luther SA, Förster R, Sedgwick JD, Browning JL, Lipp M, Cyster JG, 2000. A chemokine-driven positive feedback loop organizes lymphoid follicles. Nature 406, 309–314. [DOI] [PubMed] [Google Scholar]
- Asanuma H, Hodson Thompson A, Iwasaki T, Sato Y, Inaba Y, Aizawa C, Kurata T, Tamura S.-i.-, 1997. Isolation and characterization of mouse nasal-associated lymphoid tissue. J. Immunol. Methods 202, 123–131. [DOI] [PubMed] [Google Scholar]
- Bailey M, Haverson K, Inman C, Harris C,Jones P, Corfield G, Miller B, Stokes C, 2005. The development of the mucosal immune system pre-and post-weaning: balancing regulatory and effector function. Proc. Nutr. Soc 64, 451–457. [DOI] [PubMed] [Google Scholar]
- Bang BG, Bang F, 1968. Localized lymphoid tissues and plasma cells in paraocular and paranasal organ systems in chickens Am. J. Pathol 53, 735. [PMC free article] [PubMed] [Google Scholar]
- Belal A, El-Gohery Y, Talaat M, 1977. Nasal and paranasal pathology in experimental bilharziasis. J. Laryngol. Otol 91, 391–400. [DOI] [PubMed] [Google Scholar]
- Berlin C, Bargatze R, Campbell J, Von Andrian U, Szabo M, Hasslen S, Nelson R, Berg E, Erlandsen S, Butcher E, 1995. α4 integrins mediate lymphocyte attachment and rolling under physiologic flow. Cell 80, 413–422. [DOI] [PubMed] [Google Scholar]
- Billen F, Peeters D, Dehard S, Day M, Clercx C, 2006. Distribution of leucocyte subsets in the canine pharyngeal tonsil. J. Comp. Pathol 135, 63–73. [DOI] [PubMed] [Google Scholar]
- Brandtzaeg P, Kiyono H, Pabst R, Russell M, 2008. Terminology: nomenclature of mucosa-associated lymphoid tissue. Mucosal Immunol 1, 31–37. [DOI] [PubMed] [Google Scholar]
- Brandtzaeg P, Pabst R, 2004. Let’s go mucosal: communication on slippery ground. Trends Immunol 25, 570–577. [DOI] [PubMed] [Google Scholar]
- Brown M, McLaughlin G, Klein P, Crenshaw B, Schumacher I, Brown D, Jacobson E, 1999. Upper respiratory tract disease in the gopher tortoise is caused by Mycoplasma agassizii. J. Clin. Microbiol 37, 2262–2269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buffie CG, Pamer EG, 2013. Microbiota-mediated colonization resistance against intestinal pathogens. Nat. Rev. Immunol 13, 790–801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casteleyn C, Broos A, Simoens P, Van Den Broeck W, 2010. NALT (nasalcavity-associated lymphoid tissue) in the rabbit. Vet. Immunol. Immunopathol 133, 212–218. [DOI] [PubMed] [Google Scholar]
- Chen Z, Aspelund A, Kemble G, Jin H, 2006. Genetic mapping of the cold-adapted phenotype of B/Ann Arbor/1/66, the master donor virus for live attenuated influenza vaccines (FluMist®). Virology 345, 416–423. [DOI] [PubMed] [Google Scholar]
- Cocquyt G, Baten T, Simoens P, Van Den Broeck W, 2005. Anatomical localisation and histology of the ovine tonsils. Vet. Immunol. Immunopathol 107, 79–86. [DOI] [PubMed] [Google Scholar]
- Csencsits KL,Jutila MA, Pascual DW, 1999. Nasal-associated lymphoid tissue: phenotypic and functional evidence for the primary role of peripheral node addressin in naive lymphocyte adhesion to high endothelial venules in a mucosal site. J. Immunol 163, 1382–1389. [PubMed] [Google Scholar]
- Cullen KA, Hall MJ, Golosinskiy A, Statistics N.C.f.H, 2009. Ambulatory surgery in the United States, 2006 US Department of Health and Human Services, Centers for Disease Control and Prevention, National Center for Health Statistics, Maryland. [Google Scholar]
- De Togni P, Goellner J, Ruddle NH, Streeter PR, Fick A, Mariathasan S, Smith SC, Carlson R, Shornick LP, Strauss-Schoenberger J, 1994. Abnormal development of peripheral lymphoid organs in mice deficient in lymphotoxin. Science 264, 703–707. [DOI] [PubMed] [Google Scholar]
- Debertin A, Tschernig T, Tönjes H, Kleemann W, Tröger H, Pabst R, 2003. Nasal--associated lymphoid tissue (NALT): frequency and localization in young children. Clin. Exp. Immunol 134, 503–507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dewhirst FE, Chen T, Izard J, Paster BJ, Tanner AC, Yu W-H, Lakshmanan A, Wade WG, 2010The humanoral microbiome. J. Bacteriol 192, 5002–5017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank DN, Feazel LM, Bessesen MT, Price CS,Janoff EN, Pace NR, 2010. The human nasal microbiota and Staphylococcus aureus carriage. PLoS One 5, e10598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fukuyama S, Hiroi T, Yokota Y, Rennert PD, Yanagita M, Kinoshita N, Terawaki S, Shikina T, Yamamoto M, Kurono Y, 2002. Initiation of NALT organogenesis is independent of the IL-7R, LTβR, and NIK signaling pathways but requires the Id2 gene and CD3- CD4+ CD45+ cells. Immunity 17, 31–40. [DOI] [PubMed] [Google Scholar]
- Fukuyama S, Nagatake T, Kim D-Y, Takamura K, Park EJ, Kaisho T, Tanaka N, Kurono Y, Kiyono H, 2006. Cutting edge: uniqueness of lymphoid chemokine requirement for the initiation and maturation of nasopharynx-associated lymphoid tissue organogenesis. J. Immunol 177, 4276–4280. [DOI] [PubMed] [Google Scholar]
- Fukuyama Y, Tokuhara D, Kataoka K, Gilbert RS, McGhee JR, Yuki Y, Kiyono H, Fujihashi K, 2012. Novel vaccine development strategies for inducing mucosal immunity. Expert Rev. Vaccines 11, 367–379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldstine SN, Manickavel V, Cohen N, 1975. Phylogeny of gut-associated lymphoid tissue. Am. Zool 15, 107–118. [Google Scholar]
- Harkema J, Plopper C, Hyde D, St George J, 1987. Regional differences in quantities of histochemically detectable mucosubstances in nasal, paranasal, and nasopharyngeal epithelium of the bonnet monkey. J. Histochem. Cytochem 35, 279–286. [DOI] [PubMed] [Google Scholar]
- Harmsen A, Kusser K, Hartson L, Tighe M, Sunshine MJ, Sedgwick JD, Choi Y, Littman DR, Randall TD, 2002. Cutting edge: organogenesis of nasal-associated lymphoid tissue (NALT) occurs independently of lymphotoxin-α (LTα) and retinoic acid receptor-related orphan recrptor-γ but the organization of NALT is LTα dependent. J. Immunol 168, 986–990. [DOI] [PubMed] [Google Scholar]
- Henriksson G, Helgeland L, Midtvedt T, Stierna P, Brandtzaeg P, 2004. Immune response to Mycoplasma pulmonis in nasal mucosa is modulated by the normal microbiota. Am. J. Respir. Cell Mol. Biol 31, 657–662. [DOI] [PubMed] [Google Scholar]
- Heritage P, Underdown B, Arsenault A, Snider D, McDermott M, 1997. Comparison of murine nasal-associated lymphoid tissue and Peyer’s patches. Am. J. Respir. Crit. Care Med 156, 1256. [DOI] [PubMed] [Google Scholar]
- Hiroi T, Iwatani K, Iijima H, Kodama S, Yanagita M, Kiyono H, 1998. Nasal immune system: distinctive Th0 and Th1/Th2 type environments in murine nasal-associated lymphoid tissues and nasal passage, respectively. Eur. J. Immunol 28, 3346–3353. [DOI] [PubMed] [Google Scholar]
- Ichinohe T, Pang IK, Kumamoto Y, Peaper DR, Ho JH, Murray TS, Iwasaki A, 2011. Microbiota regulates immune defense against respiratory tract influenza A virus infection. Proc. Natl. Acad. Sci. U. S. A 108, 5354–5359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeong K, Suzuki H, Nakayama H, Doi K, 2000. Ultrastructural study on the follicle-associated epithelium of nasal-associated lymphoid tissue in specific pathogen-free (SPF) and conventional environment-adapted (SPF-CV) rats. J. Anat 196, 443–451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang H, Yan M, Yu Q, Yang Q, 2013. Characteristics of aasal-associated Lymphoid tissue (NALT) and nasal absorption capacity in chicken. PLoS One 8, e84097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang H, Yan M, Yu Q, Yang Q, 2014. Characterization of Nasal cavity-associated lymphoid tissue in ducks. Anat. Rec 297, 916–924. [DOI] [PubMed] [Google Scholar]
- Kelemen G, 1947. The junction of the nasal cavity and the pharyngeal tube in the rat. Arch. Otolaryngol 45, 159–168. [DOI] [PubMed] [Google Scholar]
- Kiyono H, Fukuyama S, 2004. NALT-versus Peyer’s-patch-mediated mucosal immunity. Nat. Rev. Immunol 4, 699–710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koempel J, Solares C, Koltai P, 2006. The evolution of tonsil surgery and rethinking the surgical approach to obstructive sleep-disordered breathing in children. J. Laryngol. Otol 120, 993–1000. [DOI] [PubMed] [Google Scholar]
- Koni PA, Sacca R, Lawton P, Browning JL, Ruddle NH, Flavell RA, 1997. Distinct roles in lymphoid organogenesis for lymphotoxins α and β revealed in lymphotoxin β-deficient mice. Immunity 6, 491–500. [DOI] [PubMed] [Google Scholar]
- Krege J, Seth S, Hardtke S, Davalos-Misslitz ACM, Förster R, 2009. Antigen-dependent rescue of nose-associated lymphoid tissue (NALT) development independent of LTβR and CXCR5 signaling. Eur.J. Immunol 39, 2765–2778. [DOI] [PubMed] [Google Scholar]
- Kuper CF, Koornstra PJ, Hameleers DM, Biewenga J, Spit BJ, Duijvestijn AM, van Breda Vriesman PJ, Sminia T, 1992. The role of nasopharyngeal lymphoid tissue. Immunol. Today 13, 219–224. [DOI] [PubMed] [Google Scholar]
- LaPatra S, Kao S, Erhardt EB, Salinas I, 2015. Evaluation of dual nasal delivery of infectious hematopoietic necrosis virus and enteric red mouth vaccines in rainbow trout (Oncorhynchus mykiss). Vaccine 33, 771–776. [DOI] [PubMed] [Google Scholar]
- Lee H, Ruane D, Law K, Ho Y, Garg A, Rahman A, Esterházy D, Cheong C, Goljo E, Sikora A, 2015. Phenotype and function of nasal dendritic cells. Mucosal Immunol 8, 1083–1098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liang B, Hyland L, Hou S, 2001. Nasal-associated lymphoid tissue is a site of long-term virus-specific antibody production following respiratory virus infection of mice. J. Virol 75, 5416–5420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loo S, Chin K, 1974. Lymphoid tissue in the nasal mucosa of primates, with particular reference to intraepithelial lymphocytes. J. Anat 117, 249–259. [PMC free article] [PubMed] [Google Scholar]
- Lowrey L, Woodhams DC, Tacchi L, Salinas I, 2015. Topographical mapping of the rainbow trout (Oncorhynchus mykiss) microbiome reveals a diverse bacterial community in the skin with antifungal properties. Appl. Environ. Microbiol 81, 6915–6925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lycke N, 2012. Recent progress in mucosal vaccine development: potential and limitations. Nat. Rev. Immunol 12, 592–605. [DOI] [PubMed] [Google Scholar]
- Mair T, Batten E, Stokes C, Bourne F, 1987. The histological features of the immune system of the equine respiratory tract. J. Comp. Pathol 97, 575–586. [DOI] [PubMed] [Google Scholar]
- Mair T, Batten E, Stokes C, Bourne F, 1988. The distribution of mucosal lymphoid nodules in the equine respiratory tract. J. Comp. Pathol 99, 159–168. [DOI] [PubMed] [Google Scholar]
- Mestecky J, Lamm ME, Ogra PL, Strober W, Bienenstock J, McGhee JR, Mayer L, 2005. Mucosal Immunology. Academic Press, Amsterdam. [Google Scholar]
- Miyawaki S, Nakamura Y, Suzuka H, Koba M, Shibata Y, Yasumizu R, Ikehara S, 1994. A new mutation, aly, that induces a generalized lack of lymph nodes accompanied by immunodeficiency in mice. Eur. J. Immunol 24, 429–434. [DOI] [PubMed] [Google Scholar]
- Myers MA, 1928. A study of the tonsillar developments in the lingual region of anurans. J. Morphol 45, 399–439. [Google Scholar]
- Neutra MR, Kozlowski PA, 2006. Mucosal vaccines: the promise and the challenge. Nat. Rev. Immunol 6, 148–158. [DOI] [PubMed] [Google Scholar]
- Ohshima K, Hiramatsu K, 2000. Distribution of T-cell subsets and immunoglobulin-containing cells in nasal-associated lymphoid tissue (NALT) of chickens. Histol. Histopathol 15, 713–720. [DOI] [PubMed] [Google Scholar]
- Pabst R, 2015. Mucosal vaccination by the intranasal route. Nose-associated lymphoid tissue (NALT) Structure, function and species differences. Vaccine 33, 4406–4413. [DOI] [PubMed] [Google Scholar]
- Peeters D, Day M, Farnir F, Moore P, Clercx C, 2005. Distribution of leucocyte subsets in the canine respiratory tract. J. Comp. Pathol 132, 261–272. [DOI] [PubMed] [Google Scholar]
- Perry M, Whyte A, 1998. Immunology of the tonsils. Immunol. Today 19, 414–421. [DOI] [PubMed] [Google Scholar]
- Plotkin S, 2014. History of vaccination. Proc. Natl. Acad. Sci. U. S. A 111, 12283–12287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Randall T, Mebius R, 2014. The development and function of mucosal lymphoid tissues: a balancing act with micro-organisms. Mucosal Immunol 7, 455–466. [DOI] [PubMed] [Google Scholar]
- Rangel-Moreno J, Moyron-Quiroz J, Kusser K, Hartson L, Nakano H, Randall TD, 2005. Role of CXC chemokine ligand 13, CC chemokine ligand (CCL) 19, and CCL21 in the organization and function of nasal-associated lymphoid tissue. J. Immunol 175, 4904–4913. [DOI] [PubMed] [Google Scholar]
- Rebelatto M, Mead C, HogenEsch H, 2000. Lymphocyte populations and adhesion molecule expression in bovine tonsils. Vet. Immunol. Immunopathol 73, 15–29. [DOI] [PubMed] [Google Scholar]
- Rodriguez-Monroy M, Moreno-Fierros L, 2010. Striking activation of NALT and nasal passages lymphocytes induced by intranasal immunization with Cry1Ac protoxin. Scand. J. Immunol 71, 159–168. [DOI] [PubMed] [Google Scholar]
- Rodríguez-Monroy MA, Rojas-Hernández S, Moreno-Fierros L, 2007. Phenotypi and functional differences between lymphocytes from NALT and nasal passages of mice. Scand. J. Immunol 65, 276–288. [DOI] [PubMed] [Google Scholar]
- Roland PS, Rosenfeld RM, Brooks LJ, Friedman NR, Jones J, Kim TW, Kuhar S, Mitchell RB, Seidman MD, Sheldon SH, 2011. Clinical practice guideline polysomnography for sleep-disordered breathing prior to tonsillectomy in children. OTO-HNS 145, S1–S15. [DOI] [PubMed] [Google Scholar]
- Salinas I, Zhang Y-A, Sunyer JO, 2011. Mucosal immunoglobulins and B cells of teleost fish. Dev. Comp. Immunol 35, 1346–1365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schuh J, Oliphant L, 1992. Development and immunophenotyping of the pharyngeal tonsil (adenoid) in cattle. J. Comp. Pathol 106, 229–241. [DOI] [PubMed] [Google Scholar]
- Shikina T, Hiroi T, Iwatani K, Jang MH, Fukuyama S, Tamura M, Kubo T, Ishikawa H, Kiyono H, 2004. IgA class switch occurs in the organized nasopharynx-and gut-associated lymphoid tissue, but not in the diffuse lamina propria of airways and gut. J. Immunol 172, 6259–6264. [DOI] [PubMed] [Google Scholar]
- Smith PD, MacDonald TT, Blumberg RS, 2013. Principles of Mucosal Immunology Garland Science, New York. [Google Scholar]
- Stanley A, Huntley J, Jeffrey M, Buxton D, 2001. Characterization of ovine nasal-associated lymphoid tissue and identification of M cells in the overlying follicle-associated epithelium. J. Comp. Pathol 125, 262–270. [DOI] [PubMed] [Google Scholar]
- Sun Z, Unutmaz D, Zou Y-R, Sunshine MJ, Pierani A, Brenner-Morton S, Mebius RE, Littman DR, 2000. Requirement for RORγ in thymocyte survival and lymphoid organ development. Science 288, 2369–2373. [DOI] [PubMed] [Google Scholar]
- Tacchi L, Larragoite E, rin T, Munoz P, Amemiya C, hris T, Salinas I, 2015. African lungfish reveal the evolutionary origins of organized mucosal lymphoid tissue in vertebrates. Curr. Biol 25, 1–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tacchi L, Musharrafieh R, Larragoite ET, Crossey K, Erhardt EB, Martin SA, LaPatra SE, Salinas I, 2014. Nasal immunity is an ancient arm of the mucosal immune system of vertebrates. Nat. Commun 5, 6205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tamura S.-i., Iwasaki T, Thompson AH, Asanuma H, Chen Z, Suzuki Y, Aizawa C, Kurata T, 1998. Antibody-forming cells in the nasal-associated lymphoid tissue during primary influenza virus infection. J. Gen. Virol 79, 291–299. [DOI] [PubMed] [Google Scholar]
- Weese JS, Slifierz M, Jalali M, Friendship R, 2014. Evaluation of the nasal microbiota in slaughter-age pigs and the impact on nasal methicillin-resistant Staphylococcus aureus (MRSA) carriage. BMC Vet. Res 10, 69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu Z, Parra D, Gómez D, Salinas I, Zhang Y-A, von Gersdorff Jørgensen L, Heinecke RD, Buchmann K, LaPatra S, Sunyer JO, 2013. Teleost skin, an ancient mucosal surface that elicits gut-like immune responses. Proc. Natl. Acad. Sci. U. S. A 110, 13097–13102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan M, Pamp SJ, Fukuyama J, Hwang PH, Cho D-Y, Holmes S, Relman DA, 2013. Nasal microenvironments and interspecific interactions influence nasal microbiota complexity and S. aureus carriage. Cell Host Microbe 14, 631–640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yokota Y, Mansouri A, Mori S, Sugawara S, Adachi S, Nishikawa S-I, Gruss P, 1999. Development of peripheral lymphoid organs and natural killer cells depends on the helix-loop-helix inhibitor Id2. Nature 397, 702–706. [DOI] [PubMed] [Google Scholar]
- Younis RT, Lazar RH, 2002. History and current practice of tonsillectomy. Laryngoscope 112, 3–5. [DOI] [PubMed] [Google Scholar]
- Zardoya R, Meyer A, 1996. Evolutionary relationships of the coelacanth, lungfishes, and tetrapods based on the 28S ribosomal RNA gene. Proc. Natl. Acad. Sci. U. S. A 93, 5449–5454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y-A, Salinas I, Li J, Parra D, Bjork S, Xu Z, LaPatra SE, Bartholomew J, Sunyer JO, 2010. IgT, a primitive immunoglobulin class specialized in mucosal immunity. Nat. Immunol 11, 827–835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Z, Bashiruddin J, Doel C, Horsington J, Durand S, Alexandersen S, 2006. Cytokine and Toll-like receptor mRNAs in the nasal-associated lymphoid tissues of cattle during foot-and-mouth disease virus infection. J. Comp. Pathol 134, 56–62. [DOI] [PubMed] [Google Scholar]
- Zhao X, Sato A, Cruz CSD, Linehan M, Luegering A, Kucharzik T, Shirakawa A-K, Marquez G, Farber JM, Williams I, 2003. CCL9 is secreted by the follicle-associated epithelium and recruits dome region Peyer’s patch CD11b+ dendritic cells. J. Immunol 171, 2797–2803. [DOI] [PubMed] [Google Scholar]
- Zuercher AW, Coffin SE, Thurnheer MC, Fundova P, Cebra JJ, 2002. Nasal-associated lymphoid tissue is a mucosal inductive site for virus-specific humoral and cellular immune responses. J. Immunol 168, 1796–1803. [DOI] [PubMed] [Google Scholar]