

Key Points
Nonpermissive T-cell epitope group mismatches can be predicted in silico for any HLA-DPB1 allele by functional distance scores.
In silico–predicted nonpermissive DPB1 mismatches are associated with mortality and GVHD after 8/8 matched HCT.
Abstract
In silico prediction of high-risk donor–recipient HLA mismatches after unrelated donor (UD) hematopoietic cell transplantation (HCT) is an attractive, yet elusive, objective. Nonpermissive T-cell epitope (TCE) group mismatches were defined by alloreactive T-cell cross-reactivity for 52/80 HLA-DPB1 alleles (TCE-X). More recently, a numerical functional distance (FD) scoring system for in silico prediction of TCE groups based on the median impact of exon 2–encoded amino acid polymorphism on T-cell alloreactivity was developed for all DPB1 alleles (TCE-FD), including the 28/80 common alleles not assigned by TCE-X. We compared clinical outcome associations of nonpermissive DPB1 mismatches defined by TCE-X or TCE-FD in 8/8 HLA-matched UD-HCT for acute leukemia, myelodysplastic syndrome, and chronic myelogenous leukemia between 1999 and 2011 (N = 2730). Concordance between the 2 models was 92.3%, with most differences arising from DPB1*06:01 and DPB1*19:01 being differently assigned by TCE-X and TCE-FD. In both models, nonpermissive mismatches were associated with reduced overall survival (hazard ratio [HR], 1.15, P < .006 and HR, 1.12, P < .03), increased transplant-related mortality (HR, 1.31, P < .001 and HR, 1.26, P < .001) as well as acute (HR, 1.16, P < .02 and HR, 1.22, P < .001) and chronic (HR, 1.20, P < .003 and HR, 1.22, P < .001) graft-versus-host disease (GVHD). We show that in silico prediction of nonpermissive DPB1 mismatches significantly associated with major transplant outcomes is feasible for any DPB1 allele with known exon 2 sequence based on experimentally elaborated FD scores. This proof-of-principle observation opens new avenues for developing HLA risk-prediction models in HCT and has practical implications for UD searches.
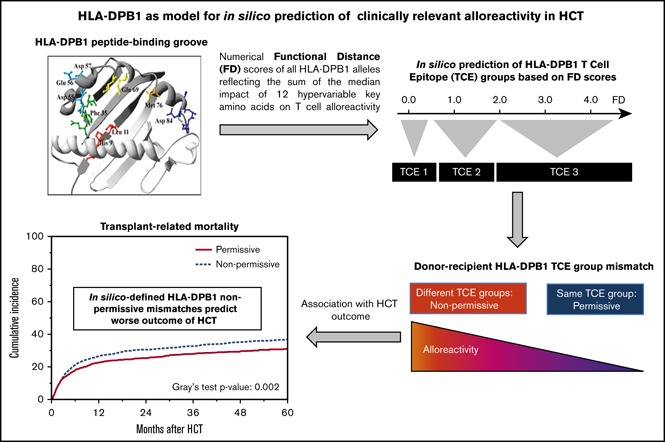
Visual Abstract
Introduction
Mismatches in the human leukocyte antigen (HLA) genes between donors and recipients mediate alloreactivity in hematopoietic cell transplantation (HCT) and contribute to the detrimental graft-versus-host disease (GVHD) and the beneficial graft-versus-leukemia effect.1,2 The search for mismatches that can be better tolerated than others, tilting the GVHD–graft-versus-leukemia balance toward the reduction in relapse rates with lower risks for transplant-related mortality (TRM), has been an intense area of research in HCT.3 However, our understanding of how to control and intelligently use alloreactivity is still incomplete, restricting its application in the clinical setting.
The exponential rate of discovery of new HLA alleles brought about by the advent of next-generation sequencing–based typing4 makes the in vitro functional testing of alloreactivity for all alleles unfeasible and underscores the need for functionally based in silico prediction models of nonpermissive mismatches applicable to all alleles. Considerable effort has been invested in developing algorithms able to predict high-risk mismatches in HCT. These models include predictions based on structural similarity of the mismatched HLAs,5-9 mismatched key amino acid residues,10,11 individual high- or low-risk mismatch combinations,12-17 and predicted indirectly recognized HLA epitopes.18,19 However, clinical outcome associations observed for these various predictive algorithms have emerged from mostly limited-sized retrospective studies that were not validated in independent cohorts in many instances,20-24 potentially exposing a lack of experimental evidence supporting the underlying hypotheses.
Successful translation of an experimentally proven hypothesis about T-cell alloreactivity into a clinically proven algorithm of nonpermissive mismatches has been achieved for the HLA-DPB1 (DPB1) locus.25 Functional assessment of alloreactive DPB1-specific T-cell cross-reactivity patterns led to the classification of 72 DPB1 alleles into 3 T-cell epitope (TCE) groups,26 and alloreactivity levels across TCE groups were shown to be higher than those between alleles within the same group.27 This differential alloreactivity was further dissected by pinpointing the weight of specific amino acid changes in the DPB1 peptide-binding groove responsible for these alloreactivity patterns.28 Moreover, poorly tolerated “nonpermissive” DPB1 mismatches have been associated with decreased survival rates in patients after HCT, while the remaining “permissive” DPB1 mismatches were associated with survival rates similar to those in patients receiving DPB1 allele matches,29,30 with the added value of reduced relapse risk for hematological malignancies. Despite these successes, the original TCE algorithm (TCE-X)26 was applicable only to those alleles for which functional evidence of cross-reactivity patterns had been obtained, and could not be extended to the remaining 903 DPB1 alleles identified to date.4 To address this limitation, we refined the TCE-X model by using the measurement of functional distance (FD) based on the median impact of amino acid polymorphism in DPB1’s peptide-binding groove to define the TCE groups, and extending it in silico to all known alleles (TCE-FD).28 In the present study, we sought to clinically validate the in silico assignment of TCE matching by TCE-FD and compare its performance with TCE-X in a large cohort of otherwise HLA-matched HCT from unrelated donors (UDs), providing a proof of principle that in silico–defined permissiveness of HLA mismatches can inform risk after HCT.
Materials and methods
Study population and clinical data
The study included 2730 patients diagnosed with acute myeloid leukemia, acute lymphoblastic leukemia, chronic myelogenous leukemia (CML), or myelodysplastic syndrome (MDS) who underwent a first myeloablative bone marrow or peripheral blood stem cell transplantation from a UD between 1999 and 2011.30,31 Details of the demographic, immunogenetic and clinical characteristics of patients and transplants are provided in Table 1. The clinical data were collected and stored by the Center for International Blood and Marrow Transplant Research, and all participants gave informed consent in agreement with the principles of the Declaration of Helsinki. The study was approved by the National Marrow Donor Program Institutional Review Board in conformity with the federal regulation regarding the protection of human research participants.
Table 1.
Patient and transplant characteristics
| TCE-X | TCE-FD | |||
|---|---|---|---|---|
| Nonpermissive (n = 1166) | Permissive (n =1564) | Nonpermissive (n = 1279) | Permissive (n = 1451) | |
| Recipient age, n (%), y | ||||
| <19 | 213 (18) | 305 (19) | 250 (19) | 268 (18) |
| 20-59 | 901 (77) | 1167 (75) | 970 (76) | 1098 (76) |
| 60+ | 52 (5) | 92 (6) | 59 (5) | 85 (6) |
| Median (range), y | 39 (<1 to 66) | 38 (<1 to 66) | 38 (<1 to 66) | 39 (<1 to 66) |
| Recipient sex, n (%) | ||||
| Male | 649 (56) | 869 (56) | 701 (55) | 817 (56) |
| Female | 517 (44) | 695 (44) | 578 (45) | 634 (44) |
| Recipient race, n (%) | ||||
| White | 1088 (93) | 1435 (92) | 1191 (93) | 1332 (92) |
| African American | 27 (2) | 48 (3) | 29 (2) | 46 (3) |
| Asian | 15 (1) | 23 (1) | 15 (1) | 23 (2) |
| Pacific Islander | 21 (2) | 31 (2) | 24 (2) | 28 (2) |
| Native American | 2 (<1) | 5 (<1) | 3 (<1) | 4 (<1) |
| No data | 13 (1) | 22 (1) | 17 (1) | 18 (1) |
| Karnofsky score prior to HCT, n (%) | ||||
| <90 | 290 (25) | 374 (24) | 318 (25) | 346 (24) |
| ≥90 | 758 (65) | 1043 (67) | 836 (65) | 965 (66) |
| No data | 118 (10) | 147 (9) | 125 (10) | 140 (10) |
| Disease at HCT, n (%) | ||||
| AML | 585 (50) | 731 (47) | 623 (49) | 693 (48) |
| ALL | 308 (26) | 417 (26) | 344 (27) | 381 (26) |
| CML | 166 (14) | 245 (16) | 193 (15) | 218 (15) |
| MDS | 107 (10) | 171 (11) | 119 (9) | 159 (11) |
| Disease status at HCT, n (%) | ||||
| Early | 519 (45) | 703 (45) | 582 (46) | 640 (44) |
| Intermediate | 349 (30) | 417 (27) | 374 (29) | 392 (27) |
| Advanced | 298 (25) | 444 (28) | 323 (25) | 419 (29) |
| Graft type, n (%) | ||||
| Bone marrow | 572 (49) | 753 (48) | 626 (49) | 699 (48) |
| Peripheral blood | 594 (51) | 811 (52) | 653 (51) | 752 (52) |
| Donor age, n (%), y | ||||
| 18-32 | 567 (49) | 741 (47) | 622 (49) | 686 (47) |
| 33+ | 576 (49) | 790 (51) | 632 (49) | 734 (51) |
| No data | 23 (2) | 33 (2) | 25 (2) | 31 (2) |
| Median age (range), y | 33 (18-56) | 33 (18-57) | 33 (18-57) | 34 (18-57) |
| Donor/recipient sex match, n (%) | ||||
| Male/male | 488 (42) | 601 (38) | 523 (41) | 566 (39) |
| Male/female | 327 (28) | 423 (27) | 361 (28) | 389 (27) |
| Female/male | 161 (14) | 268 (17) | 178 (14) | 251 (17) |
| Female/female | 190 (16) | 271 (17) | 217 (17) | 244 (17) |
| No data | 0 | 1 (<1) | 0 | 1 (<1) |
| Donor/recipient CMV match, n (%) | ||||
| −/− | 381 (33) | 504 (32) | 429 (34) | 456 (31) |
| −/+ | 388 (33) | 549 (35) | 426 (33) | 511 (35) |
| +/− | 145 (12) | 178 (12) | 159 (12) | 164 (11) |
| +/+ | 232 (20) | 311 (20) | 246 (19) | 297 (21) |
| No data | 20 (2) | 22 (1) | 19 (2) | 23 (2) |
| Donor/recipient ABO match, n (%) | ||||
| Matched | 450 (39) | 646 (41) | 509 (40) | 587 (40) |
| Mismatched | 641 (55) | 809 (51) | 689 (53) | 761 (52) |
| No data | 75 (6) | 109 (8) | 81 (7) | 103 (8) |
| Total body irradiation, n (%) | ||||
| No | 505 (43) | 652 (42) | 546 (43) | 611 (42) |
| Yes | 649 (56) | 893 (57) | 720 (56) | 822 (57) |
| No data | 12 (1) | 19 (1) | 13 (1) | 18 (1) |
| In vivo T-cell depletion (ATG or alemtuzumab), n (%) | ||||
| No | 845 (72) | 1169 (75) | 919 (72) | 1095 (75) |
| Yes | 321 (28) | 395 (25) | 360 (28) | 356 (25) |
| DQB1 matching, n (%) | ||||
| Mismatch | 83 (7) | 143 (9) | 99 (8) | 127 (9) |
| Match | 1075 (92) | 1402 (90) | 1170 (91) | 1307 (90) |
| No data | 8 (<1) | 19 (1) | 10 (<1) | 17 (1) |
| Year of HCT, n (%) | ||||
| 1999-2006 | 754 (65) | 988 (63) | 823 (64) | 919 (63) |
| 2007-2011 | 412 (35) | 576 (37) | 456 (36) | 532 (37) |
| Median follow-up of survivors (range), mo | 61 (3-151) | 62 (3-150) | 61 (3-151) | 62 (3-150) |
Only DPB1- mismatched pairs that could be classified as nonpermissive or permissive by the TCE-X and the TCE-FD model are shown. GVHD prophylaxis was based on methotrexate, with or without cyclosporine A or other, or mycophenolate mofetil, cyclosporine A, or FK506 alone or with other.
ALL, acute lymphoblastic leukemia; AML, acute myeloid leukemia; ATG, antithymocyte globulin.
HLA typing and DPB1 matching
Only patient/donor pairs with second field resolution typing at the HLA loci A, B, C, DRB1, DQB1, and DPB1 were included. Patients and donors were 8/8 matched for HLA-A, -B, -C, and -DRB1 but were mismatched for DPB1. Pairs with mismatches at other major HLA loci were not included to avoid a confounding effect on clinical outcome. For definition of permissive and nonpermissive DPB1 mismatches, the TCE algorithm was applied as previously described.26 Briefly, DPB1 alleles were classified into 3 TCE groups according to the original TCE-X or the new TCE-FD model, as described in “ Results” (supplemental Table 1). Nonpermissive and permissive DPB1 mismatches were subsequently defined according to the TCE3 model,26,29 using the online DPB1 TCE Web tool from the IMGT/HLA database4 (https://www.ebi.ac.uk/ipd/imgt/hla/dpb.html) version 1 and version 2 for TCE-X and TCE-FD, respectively. No novel DPB1 alleles were identified in this cohort at this level of resolution.
Clinical outcomes
The primary study end point was overall survival (OS); secondary study end points were disease-free survival (DFS), TRM, relapse incidence (RI), chronic GVHD (cGVHD) and acute GVHD (aGVHD), defined as previously described.30
Statistical analysis
For evaluation of clinical results, the Kruskal-Wallis and χ2 tests were used to analyze differences for discrete or continuous factors, respectively, between the 2 different HLA-DPB1 matching groups (mismatched permissive or mismatched nonpermissive). Kaplan-Meier probabilities were calculated for OS and DFS. Estimated cumulative incidence was calculated for events with competing risks (ie, TRM, RI, and aGVHD). Comparisons of survival curves and cumulative incidence rates were done with the log-rank test and the Gray test, respectively. Multivariable regression analyses using the Cox proportional hazards model were generated for each HLA-DP matching algorithm independently as the main effect. Models were fit to determine which risk factors were related to a given outcome. All variables were tested for affirmation of the proportional hazards assumption. Factors violating the proportional hazards assumption were added as time-dependent covariates. A forward stepwise model-building procedure was used to select risk factors for each outcome with a threshold of P ≤ .01 for entering the models. Due to multiple comparisons, P < .01 was used to determine statistical significance for the main effect. All analyses were performed using SAS version 9.4 (SAS Institute, Inc.).
Results
Experimental and in silico prediction of nonpermissive DPB1 TCE group mismatches
The original classification of DPB1 alleles into 3 structurally different TCE groups (TCE-X) was based on the ability of T cells obtained from a patient with allograft rejection post-HCT alloreactive to DPB1*09:01 to cross-recognize other DPB1 alleles encoding similar epitopes (Figure 1A). This initial classification was limited to a set of 72 DPB1 alleles for which B lymphoblastoid cell lines were available for direct in vitro testing26,32 and includes only 52/80 DPB1 alleles that have been described to be common and well-documented (CWD) in catalogs from the American Society of Histocompatibility and Immunogenetics and the European Federation for Immunogenetics33,34 (supplemental Table 1).
Figure 1.

TCE group classification of DPB1 alleles by TCE-X or TCE-FD. (A) TCE classification of DPB1 alleles based on in vitro cross-reactivity patterns of alloreactive T cells nominally directed against DPB1*09:01.26 Alleles that were reproducibly cross-recognized by a panel of 5 DPB1*09:01-specific alloreactive T-cell effectors were grouped together as TCE group 1 (red), whereas DPB1 alleles recognized by only a part or none of these T cells were classified into TCE groups 2 (orange) and 3 (green), respectively.26 In total, 72 of the 975 DPB1 alleles known to date were experimentally tested for TCE-X (see Supplemental Table 1 for a comprehensive list of TCE group assignment for these alleles). These 72 DPB1 alleles classifiable by TCE-X represent only 7.4% of all known DPB1 alleles and account for only 65% of the CWD alleles for this locus.33,34 Shown here are 19 DPB1 alleles reported with a frequency ≥ 0.5% in in a large population of European descent,68 which together make up 98% of the total DPB1 allele frequency. Two alleles (DPB1*06:01 and *19:01) that changed classification from TCE3 in TCE-X to TCE2 in TCE-FD are indicated in bold. (B) TCE classification of any known DPB1 allele based on in silico prediction by FD scores. The latter were experimentally obtained by generating a panel of 12 site-directed single amino acid mutants of DPB1*09:01 and assessing the median strength of recognition for each mutant by alloreactive T cells relative to wild-type DPB1*09:01.28 Based on this, the FD amino acid (FDaa) scores were calculated for each mutant, with high scores representing a high functional impact and low scores representing a low functional impact of the mutation. For each DPB1 allele, the sum of FDaa scores of each of the 12 polymorphic residues analyzed was used to calculate the FDallele score. Experimental TCE classification by TCE-X was found to reflect the range of FDallele scores as follows: TCE1, FDallele < 0.6; TCE2, 0.6 < FDallele < 2.0; and TCE3, FDallele > 2.0.28 TCE classification of the 19 DPB1 from panel A is indicated with TCE1, TCE2, and TCE3 in red, orange, and green, respectively. Two alleles (DPB1*06:01 and *19:01) that changed classification from TCE3 in TCE-X to TCE2 in TCE-FD are indicated in bold.
In view of this and of the ever-increasing number of known DPB1 alleles (975 known variants to date),4 we developed an alternative model for DPB1 TCE group assignment by in silico prediction based on experimentally defined FD for each DPB1 allele (TCE-FD model). Briefly, FDaa scores for 12 key polymorphic amino acid residues encoded by exon 2 of DPB1*09:01 were determined as the median impact of naturally occurring substitutions at each of these residues on allorecognition of wild-type DPB1*09:01 by alloreactive T cells (Figure 1B). This allowed us to calculate the corresponding FDallele score as the sum of FDaa scores encoded by each allele’s exon 2 sequence. We found that FDallele score ranges faithfully reflected the TCE groups, with FDallele < 0.6 for TCE group 1, 0.6 < FDallele < 2.0 for TCE group 2, and FDallele > 2.0 for TCE group 3.28 Only 2 alleles fell short of this definition, namely DPB1*06:01 and DPB1*19:01, which had been assigned to TCE group 3 by TCE-X despite FDallele scores of 1.41 and 1.43, respectively. Further functional testing with a larger panel of T-cell effectors confirmed their classification into TCE group 2.28 Because all known DPB1 alleles have at least complete exon 2 sequence available, the TCE-FD model allows for in silico prediction of TCE assignment for all alleles at this locus. To confirm the clinical usefulness of this model, we tested its influence on HCT outcome, as explained below.
Study population and outcomes
The study cohort included 2730 patients with acute leukemia, CML, or MDS who received an 8/8 HLA-matched DPB1-mismatched UD HCT that could be classified as nonpermissive or permissive by the TCE-X and the TCE-FD model. Overall outcomes for the investigated clinical end points were OS 41%, DFS 37%, TRM 32%, RI 30%, severe (grades III-IV) aGVHD 19% at 100 days, and cGVHD 50% at 5 years. Median follow-up time was 61 months.
Prediction of nonpermissive or permissive DPB1 mismatches by TCE-X or TCE-FD
In 2697 of 2730 pairs, the DPB1 alleles of patient and donors were included in the 72 alleles assignable by TCE-X. Only in 33 of 2730 pairs (1.2%) was ≥1 DPB1 allele of patient and/or donor not part of these 72 alleles. These included 9 alleles: DPB1*18:01 (n = 16), DPB1*26:01 (n = 5), DPB1*27:01 (n = 4), DPB1*35:01 (n = 3), DPB1*85:01 (n = 2), and DPB1*21:01, *29:01, *30:01, and *130:01 (n = 1 each) (supplemental Table 1). In the TCE-X classification, these 9 alleles were assigned to TCE group 3 by default, whereas 4 of them were classified as TCE group 1 or 2 by TCE-FD (supplemental Table 1). This resulted in discordant assignment as permissive by TCE-X and nonpermissive by TCE-FD in only 5 cases. An additional 205 discordances in the same (N = 156) or the opposite (N = 49) direction were all due to the presence of DPB1*06:01 and/or *19:01, which were assigned to TCE groups 2 and 3 by TCE-FD and TCE-X, respectively (Table 2). Overall, 1118 and 1402 pairs were concordantly assigned as nonpermissive or permissive, respectively, by TCE-X and TCE-FD, for an overall concordance of 92.3%. TCE-X had slightly more permissive pairs (1564/2730, 57.3%) compared with TCE-FD (1451/2730, 53.2%) (Table 2).
Table 2.
Cross-tabulation between TCE-X and TCE-FD
| TCE-X | |||
|---|---|---|---|
| TCE-FD | Nonpermissive, n (%) | Permissive, n (%) | Total, N (%) |
| Nonpermissive | 1118 (40.9) | 161 (5.9) | 1279 (46.8) |
| Permissive | 49 (1.9) | 1402 (51.3) | 1451 (53.2) |
| Total | 1166 (42.8) | 1564 (57.2) | 2730 (100) |
Percentages refer to the total number of 2730 8/8 HLA-matched DPB1-mismatched donor–recipient pairs analyzed in this study. The 210 discordant assignments as TCE-X permissive and TCE-FD nonpermissive (n = 161) or vice versa (n = 49) were due to the presence of DPB1*06:01 and/or 19:01, which were classified as TCE group 2 and 3 by TCE-FD and TCE-X, respectively (n = 205) or to the presence of a DPB1 allele that was classified as TCE group 1 or 2 by TCE-FD but was attributed to TCE group 3 by TCE-X because of missing data (see Supplemental Table 1) (n = 5).
Clinical risk associations of nonpermissive DPB1 mismatches by TCE-X or TCE-FD
As previously reported,26,29-31,35,36 the cumulative incidence of TRM was significantly lower for HCT from nonpermissively DPB1-mismatched UDs according to TCE-X compared with permissively mismatched HCT (Figure 2A). The same was observed when nonpermissive and permissive DPB1 mismatches were defined according to the TCE-FD model (Figure 2B). The association of nonpermissive DPB1 mismatches according to the 2 models with all major outcome end points was analyzed using multivariable Cox regression models adjusted for the main clinical variables (Table 3). The significant association with TRM was confirmed for TCE-X and TCE-FD (hazard ratio [HR], 1.31; 95% confidence interval [CI], 1.14-1.50, P < .001 for TCE-X and HR, 1.26; 95% CI, 1.1-1.44, P < .001 for TCE-FD). In line with this, the probability of OS was lower for nonpermissive DPB1 mismatches defined by TCE-X and TCE-FD, although this was significant for TCE-X (HR, 1.15; 95% CI, 1.04-1.27, P < .006) but not for TCE-FD (HR, 1.12; 95% CI, 1.01-1.23, P < .03) (Table 3). Interestingly, in this cohort, nonpermissive DPB1 mismatches by TCE-X and TCE-FD were also associated with increased risks for aGVHD II-IV (HR, 1.16; 95% CI, 1.03-1.30; P < .02 for TCE-X and HR, 1.22; 95% CI, 1.09-1.37; P < .001 for TCE-FD) and cGVHD (HR, 1.20; 95% CI, 1.07-1.34; P < .003 for TCE-X and HR, 1.22; 95% CI, 1.09-1.36; P < .001 for TCE-FD) (Table 3).
Figure 2.

Cumulative incidence of TRM by nonpermissive or permissive DPB1 mismatches according to TCE-X or TCE-FD. Shown are probabilities of TRM for transplants stratified according to DPB1 nonpermissive or permissive mismatches by TCE-X (A) or TCE-FD (B). The numbers in each group were n = 1166, 1564, 1279, and 1451 for TCE-X nonpermissive, TCE-X permissive, TCE-FD nonpermissive, and TCE-FD permissive pairs, respectively.
Table 3.
Multivariable regression models for association between nonpermissive DPB1 mismatches and clinical outcome
| TCE-X nonpermissive (n = 1166) | TCE-FD nonpermissive (n = 1279) | |||
|---|---|---|---|---|
| HR (95% CI) | P | HR (95% CI) | P | |
| OS | 1.15 (1.04-1.27) | .005 | 1.12 (1.01-1.23) | .028 |
| DFS | 1.11 (1.00-1.22) | .044 | 1.07 (0.97-1.18) | .159 |
| TRM | 1.231 (1.14-1.50) | <.001 | 1.26 (1.1-1.44) | <.001 |
| Relapse | 0.93 (0.81-1.07) | .307 | 0.91 (0.79-1.05) | .194 |
| cGVHD | 1.20 (1.07-1.34) | .002 | 1.22 (1.09-1.36) | <.001 |
| aGVHD II-IV | 1.16 (1.03-1.30) | .014 | 1.22 (1.09-1.37) | <.001 |
| aGVHD III-IV | 1.08 (0.90-1.29) | .428 | 1.03 (0.86-1.24) | .759 |
Data are shown using permissive mismatches as reference (n = 1564 for TCE-X and n = 1451 for TCE-FD).
Because TCE-X and TCE-FD differ mainly in the assignment of DPB1*06:01 and 19:01 to different TCE groups, we analyzed multivariable outcome associations of the discordant groups separately, using the concordant permissive pairs as reference (supplemental Table 2). The HRs obtained for the 2 discordant subgroups mirrored the above-described findings for the overall cohort. However, probably due to the low number of discordant pairs, these associations were not significant for any of the end points studied with the exception of aGVHD II-IV, for which pairs considered permissive by TCE-X but deemed nonpermissive by TCE-FD showed significantly increased risks compared with the concordant permissive pairs (HR, 1.53; 95%, CI 1.22-1.93; P < .001) (supplemental Table 2).
We also tested an additional approach of identifying high-risk DPB1 mismatches based on the numerical difference in FD scores of DPB1 alleles between patients and donors, designated δ Functional Distance or dFD, as previously described.37 The cutoff threshold for high or low-risk dFD scores was set at 2.665 in our previous study.37 Here, we tested this cutoff and additionally the value 1.64, which is the mean of dFD scores observed in the present cohort (range, 0.00-9.40). As shown in supplemental Table 3, above-threshold dFD scores were significantly associated with TRM for both cutoff values in multivariable models. Moreover, dFD scores above the cutoff 1.64 were also significantly associated with OS and aGVHD II-IV, similar to TCE-defined nonpermissive mismatches. Based on these data, the dFD model is not superior to the TCE model in defining high-risk DPB1 mismatches in UD HCT.
Discussion
In this study we have performed a clinical validation of in silico–predicted nonpermissive DPB1 mismatches via extension of the refined TCE matching algorithm (TCE-FD) to all known DPB1 alleles. Matching concordance between the 2 TCE models was 92.3%, with most differences arising from alleles DPB1*06:01 and DPB1*19:01 having different TCE group assignments in TCE-X and TCE-FD. By studying the impact of DPB1 mismatches in a large cohort of otherwise HLA-matched HCT from UDs and comparing it with the previous limited algorithm (TCE-X), we show that in silico–predicted nonpermissive TCE-FD mismatches result in reduced OS and increased TRM, as well as high risks for aGVHD and cGVHD. The hazards for all clinical end points were very similar for the 2 models, likely reflecting the fact that the most common alleles in European populations share the same TCE classification in both. In line with this, analysis of 210 pairs with discordant assignments did not reveal any superiority of one of the models in predicting outcome. Therefore, the differential TCE group assignment of DPB1*06:01 and *19:01 appears not to have a strong impact on the predictive value of the model, as also shown by subgroup analyses of discordant pairs. On the other hand, the applicability of TCE-FD to all current and future DPB1 alleles is a major strength that renders it overall superior to TCE-X.
Several attempts to translate in vitro functional evidence of differential alloreactivity38,39 or in silico–predicted allorecognition patterns5,6,40 into clinically relevant HLA-matching algorithms have been carried out to inform risk assessment in HCT. Nevertheless, in silico immunogenicity predictions, as well as functional in vitro data, do not always correlate with each other or adequately inform clinical risk prediction, as previously shown by other investigators.20-24,41,42 In addition, approaches relying on predicted binding of peptides to HLA molecules are limited by the accuracy of these predictive tools and their algorithms.43 More successful strategies have been developed for transplantation of solid organs, for which prediction of the recognition of antibody-accessible epitopes present in donor HLAs and absent in the recipient’s HLAs44,45 is used to avoid humoral responses against the graft.7,46,47 Of note, however, the same algorithm failed to predict outcome in the HCT setting.48
With our approach based on the experimentally determined FD that arises from the specific weight of amino acid differences in DPB1’s peptide-binding groove between different alleles,28 we were able to extend the TCE algorithm to all 975 DPB1 alleles described to date,4 including 80 DPB1 alleles reported as CWD in the American Society of Histocompatibility Immunogenetics or the European Federation for Immunogenetics catalog,33,34 without the need for further in vitro testing. Importantly, although other approaches have tried to correlate the impact of amino acid substitutions at different positions of the HLA peptide-binding groove with alloreactivity patterns and HCT outcome,10-13,49 ours does not consider all substitutions equally; rather, it draws on direct experimental evidence on the differential impact of different types of amino acid substitutions at key positions. A further strength of our approach is that TCE grouping can now be applied prospectively to any new DPB1 allele discovered for which its exon 2 sequence is known. In the relatively ethnically conserved cohort under analysis in this study, DPB1 alleles assignable by TCE-FD, but not by TCE-X, were present in only 1.2% of pairs. However, the switch to next-generation sequencing–based HLA typing methods has significantly accelerated the discovery of new alleles in recent years,50,51 with 378 of the current 975 DPB1 alleles reported in the last 2 years alone.4 Moreover, HCT is also becoming increasingly feasible in emerging countries,52 further contributing to the likelihood of new alleles being discovered in new ethnic groups. Based on all of this, the usefulness of in silico prediction by TCE-FD is likely to increase over time. The TCE-X and TCE-FD matching algorithms are freely available through the IMGT/HLA database (www.ebi.ac.uk/ipd/imgt/hla).53 TCE-FD has also been incorporated into UD search tools provided to clinicians by stem cell donor registries in the United States (HapLogic; https://bethematch.org) and Germany (Optimatch; www.zkrd.de).54,55
In addition to the associations with mortality reported previously, both algorithms were also able to detect an association with acute and chronic GVHD in this cohort. This finding is concordant with a recent report underlining the relevant effect of DPB1 mismatching on GVHD risks in patients transplanted with UDs compared with sibling donors.56 In that same study, permissive and nonpermissive mismatching for DPB1 significantly decreased the risk for relapse or disease progression after UD HCT, whereas permissive DPB1 mismatches were associated with similar incidences of GVHD-related outcomes compared with sibling donors. These findings support again the concept of feasible57 intelligent DPB1 mismatching for specific leukemia immunotherapy in the context of allogeneic HCT, as proposed also by other investigators.58-61
In a previous single-center study, we found significant associations between the numerical difference in FD (dFD) and HCT outcome,37 which appeared to be even stronger compared with nonpermissive TCE mismatches. Although we did find significant associations of above-threshold dFD scores with OS, TRM, and aGVHD, when the mean dFD value was used as cutoff in the present study, these associations were not stronger than those observed with the TCE model of nonpermissive mismatches. These data suggest that dFD could be a surrogate for TCE, which remains superior because it has been most widely tested clinically and does not present the difficulty of defining a generally applicable cutoff value associated with the dFD model.
The TCE model of nonpermissive mismatches still has some limitations. In particular, all evidence for FD between DPB1 alleles is based on single-position site-directed mutagenesis of only 1 allele: DPB1*09:01. Moreover, our quantification of FD assumes an additive effect of the different amino acid substitutions appearing naturally together in other alleles. Future experiments examining FD derived from mutagenesis of other DPB1 alleles representative of all 3 TCE groups and studying the joint effect of several mutations in the molecule’s peptide-binding groove are warranted to further refine our in silico predictions. In addition, direct characterization of peptide repertoires and their overlap between TCE groups and alleles within each group are also being undertaken to complement the definition of FD. Our findings should also be revalidated in additional clinical cohorts, in particular in large cohorts homogenously receiving in vivo T-cell depletion by antithymocyte globulin or posttransplant cyclophosphamide. Interestingly, a recent report on a single-center study showed the validity of the TCE-FD algorithm in a cohort of patients treated with in vivo T-cell depletion by antithymocyte globulin,62 suggesting a predictive value of our new approach in that setting.
Recent reports have suggested an effect of differential 3′ untranslated region–controlled expression levels on permissiveness of DPB1 mismatches in the context of HCT.63,64 Moreover, another model of analysis of DPB1 mismatches developed for Japanese HCT recipients and based on the evolutionary relationship between DPB1 alleles defined by the region between exon 3 and the 3′UTR has also provided evidence of increased risks for aGVHD in transplant pairs with mismatches across the 2 evolutionary allele groups: DP2 and DP5.65 The TCE model and these 2 models correlate to some extent, and their mode of interaction is currently under investigation by us and other investigators.65-67
In conclusion, in this study we have demonstrated the feasibility and clinical relevance of in silico assignment of nonpermissive DPB1 mismatches conferring higher risks for complications after HCT. Nonpermissive TCE group mismatches can now be predicted in silico for any DPB1 allele by FD scores. This successful proof-of-principle experience of translation of direct FD to in silico prediction of clinically detrimental HLA mismatches in HCT should open new potential avenues for future development of risk prediction models including other loci of the HLA system.
Supplementary Material
The full-text version of this article contains a data supplement.
Acknowledgments
This work was supported by grants from the Deutsche José Carreras Leukämie Stiftung (DJCLS R 15/02) and from the Joseph Senker Stiftung (K.F.). The Center for International Blood and Marrow Transplant Research is supported by Public Health Service Grant/Cooperative Agreement 5U24-CA076518 among the National Institutes of Health National Cancer Institute, National Heart, Lung and Blood Institute, and National Institute of Allergy and Infectious Diseases; a Grant/Cooperative Agreement 5U10HL069294 between National Institutes of Health National Heart, Lung and Blood Institute and National Cancer Institute; Contract CHHSH250201200016C with Health Resources and Services Administration (Department of Health and Human Services); Grants N00014-15-1-0848 and N00014-16-1-2020 from the Office of Naval Research; and grants from Alexion; *Amgen, Inc.; an anonymous donation to the Medical College of Wisconsin; Astellas Pharma US; AstraZeneca; Be the Match Foundation; *Bluebird Bio, Inc.; *Bristol Myers Squibb Oncology; *Celgene Corporation; Cellular Dynamics International, Inc.; *Chimerix, Inc.; Fred Hutchinson Cancer Research Center; Gamida Cell Ltd.; Genentech, Inc.; Genzyme Corporation; *Gilead Sciences, Inc.; Health Research, Inc. Roswell Park Cancer Institute; HistoGenetics, Inc.; Incyte Corporation; Janssen Scientific Affairs, LLC; *Jazz Pharmaceuticals, Inc.; Jeff Gordon Children’s Foundation; The Leukemia & Lymphoma Society; Medac, GmbH; MedImmune; The Medical College of Wisconsin; *Merck & Co., Inc.; Mesoblast; MesoScale Diagnostics, Inc.; *Miltenyi Biotec, Inc.; National Marrow Donor Program; Neovii Biotech NA, Inc.; Novartis Pharmaceuticals Corporation; Onyx Pharmaceuticals; Optum Healthcare Solutions, Inc.; Otsuka America Pharmaceutical, Inc.; Otsuka Pharmaceutical Co, Ltd. – Japan; PCORI; Perkin Elmer, Inc.; Pfizer, Inc; *Sanofi US; *Seattle Genetics; *Spectrum Pharmaceuticals, Inc.; St. Baldrick’s Foundation; *Sunesis Pharmaceuticals, Inc.; Swedish Orphan Biovitrum, Inc.; Takeda Oncology; Telomere Diagnostics, Inc.; University of Minnesota; and *Wellpoint, Inc. (*corporate members).
The views expressed in this article do not reflect the official policy or position of the National Institutes of Health, the Department of the Navy, the Department of Defense, Health Resources and Services Administration or any other agency of the US Government.
Authorship
Contribution: P.C., K.F., S.R.S., and S.J.L. designed the study; K.W.A. and H.-L.W. performed statistical analyses; E.A.-B., P.C., and K.F. drafted the manuscript; and all authors participated in manuscript writing and review and provided final approval of the manuscript.
Conflict-of-interest disclosure: The authors declare no competing financial conflicts.
Correspondence: Katharina Fleischhauer, Institute for Experimental Cellular Therapy, Essen University Hospital, Hufelandstr 55, 45122 Essen, Germany; e-mail: katharina.fleischhauer@uk-essen.de.
References
- 1.Falkenburg JH, Jedema I. Allo-reactive T cells for the treatment of hematological malignancies. Mol Oncol. 2015;9(10):1894-1903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Fleischhauer K, Beelen DW. HLA mismatching as a strategy to reduce relapse after alternative donor transplantation. Semin Hematol. 2016;53(2):57-64. [DOI] [PubMed] [Google Scholar]
- 3.Negrin RS. Graft-versus-host disease versus graft-versus-leukemia. Hematology Am Soc Hematol Educ Program. 2015;2015:225-230. [DOI] [PubMed] [Google Scholar]
- 4.Robinson J, Halliwell JA, Hayhurst JD, Flicek P, Parham P, Marsh SG. The IPD and IMGT/HLA database: allele variant databases. Nucleic Acids Res. 2015;43(Database issue):D423-D431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Jöris MM, van Rood JJ, Roelen DL, Oudshoorn M, Claas FH. A proposed algorithm predictive for cytotoxic T cell alloreactivity. J Immunol. 2012;188(4):1868-1873. [DOI] [PubMed] [Google Scholar]
- 6.Elsner HA, DeLuca D, Strub J, Blasczyk R. HistoCheck: rating of HLA class I and II mismatches by an internet-based software tool. Bone Marrow Transplant. 2004;33(2):165-169. [DOI] [PubMed] [Google Scholar]
- 7.Kosmoliaptsis V, Bradley JA, Sharples LD, et al. Predicting the immunogenicity of human leukocyte antigen class I alloantigens using structural epitope analysis determined by HLAMatchmaker. Transplantation. 2008;85(12):1817-1825. [DOI] [PubMed] [Google Scholar]
- 8.Heemskerk MB, Roelen DL, Dankers MK, et al. Allogeneic MHC class I molecules with numerous sequence differences do not elicit a CTL response. Hum Immunol. 2005;66(9):969-976. [DOI] [PubMed] [Google Scholar]
- 9.Heemskerk MB, Cornelissen JJ, Roelen DL, et al. Highly diverged MHC class I mismatches are acceptable for haematopoietic stem cell transplantation. Bone Marrow Transplant. 2007;40(3):193-200. [DOI] [PubMed] [Google Scholar]
- 10.Ferrara GB, Bacigalupo A, Lamparelli T, et al. Bone marrow transplantation from unrelated donors: the impact of mismatches with substitutions at position 116 of the human leukocyte antigen class I heavy chain. Blood. 2001;98(10):3150-3155. [DOI] [PubMed] [Google Scholar]
- 11.Pidala J, Wang T, Haagenson M, et al. Amino acid substitution at peptide-binding pockets of HLA class I molecules increases risk of severe acute GVHD and mortality. Blood. 2013;122(22):3651-3658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kawase T, Matsuo K, Kashiwase K, et al. ; Japan Marrow Donor Program. HLA mismatch combinations associated with decreased risk of relapse: implications for the molecular mechanism. Blood. 2009;113(12):2851-2858. [DOI] [PubMed] [Google Scholar]
- 13.Kawase T, Morishima Y, Matsuo K, et al. ; Japan Marrow Donor Program. High-risk HLA allele mismatch combinations responsible for severe acute graft-versus-host disease and implication for its molecular mechanism. Blood. 2007;110(7):2235-2241. [DOI] [PubMed] [Google Scholar]
- 14.Yagasaki H, Kojima S, Yabe H, et al. ; Japan Marrow Donor Program. Acceptable HLA-mismatching in unrelated donor bone marrow transplantation for patients with acquired severe aplastic anemia. Blood. 2011;118(11):3186-3190. [DOI] [PubMed] [Google Scholar]
- 15.Fernandez-Viña MA, Wang T, Lee SJ, et al. Identification of a permissible HLA mismatch in hematopoietic stem cell transplantation. Blood. 2014;123(8):1270-1278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Passweg JR, Schanz U, Chalandon Y, et al. ; Swiss Blood Stem Cell Transplantation Group. High-resolution HLA matching in unrelated donor transplantation in Switzerland: differential impact of class I and class II mismatches may reflect selection of nonimmunogenic or weakly immunogenic DRB1/DQB1 disparities. Bone Marrow Transplant. 2015;50(9):1201-1205. [DOI] [PubMed] [Google Scholar]
- 17.Pasi A, Crocchiolo R, Bontempelli M, et al. The conundrum of HLA-DRB1*14:01/*14:54 and HLA-DRB3*02:01/*02:02 mismatches in unrelated hematopoietic SCT. Bone Marrow Transplant. 2011;46(7):916-922. [DOI] [PubMed] [Google Scholar]
- 18.Thus KA, de Hoop TA, de Weger RA, Bierings MB, Boelens JJ, Spierings E. Predicted indirectly recognizable HLA epitopes class I promote antileukemia responses after cord blood transplantation: indications for a potential novel donor selection tool. Biol Blood Marrow Transplant. 2016;22(1):170-173. [DOI] [PubMed] [Google Scholar]
- 19.Thus KA, Ruizendaal MT, de Hoop TA, et al. Refinement of the definition of permissible HLA-DPB1 mismatches with predicted indirectly recognizable HLA-DPB1 epitopes. Biol Blood Marrow Transplant. 2014;20(11):1705-1710. [DOI] [PubMed] [Google Scholar]
- 20.Askar M, Sobecks R, Morishima Y, et al. Predictions in the face of clinical reality: HistoCheck versus high-risk HLA allele mismatch combinations responsible for severe acute graft-versus-host disease. Biol Blood Marrow Transplant. 2011;17(9):1409-1415. [DOI] [PubMed] [Google Scholar]
- 21.Huo MR, Li D, Chang YJ, et al. Predicted indirectly recognizable HLA epitopes are not associated with clinical outcomes after haploidentical hematopoietic stem cell transplantation. Hum Immunol. 2018;79(2):117-121. [DOI] [PubMed] [Google Scholar]
- 22.Jöris MM, Lankester AC, von dem Borne PA, et al. Translating in vitro prediction of cytotoxic T cell alloreactivity to hematopoietic stem cell transplantation outcome. Transpl Immunol. 2014;30(2-3):59-64. [DOI] [PubMed] [Google Scholar]
- 23.Shaw BE, Barber LD, Madrigal JA, Cleaver S, Marsh SG. Scoring for HLA matching? A clinical test of HistoCheck. Bone Marrow Transplant. 2004;34(4):367-368, author reply 369. [DOI] [PubMed] [Google Scholar]
- 24.Spellman S, Klein J, Haagenson M, et al. Scoring HLA class I mismatches by HistoCheck does not predict clinical outcome in unrelated hematopoietic stem cell transplantation. Biol Blood Marrow Transplant. 2012;18(5):739-746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Fleischhauer K, Shaw BE. HLA-DP in unrelated hematopoietic cell transplantation revisited: challenges and opportunities. Blood. 2017;130(9):1089-1096. [DOI] [PubMed] [Google Scholar]
- 26.Zino E, Frumento G, Marktel S, et al. A T-cell epitope encoded by a subset of HLA-DPB1 alleles determines nonpermissive mismatches for hematologic stem cell transplantation. Blood. 2004;103(4):1417-1424. [DOI] [PubMed] [Google Scholar]
- 27.Sizzano F, Zito L, Crivello P, et al. Significantly higher frequencies of alloreactive CD4+ T cells responding to nonpermissive than to permissive HLA-DPB1 T-cell epitope disparities. Blood. 2010;116(11):1991-1992. [DOI] [PubMed] [Google Scholar]
- 28.Crivello P, Zito L, Sizzano F, et al. The impact of amino acid variability on alloreactivity defines a functional distance predictive of permissive HLA-DPB1 mismatches in hematopoietic stem cell transplantation. Biol Blood Marrow Transplant. 2015;21(2):233-241. [DOI] [PubMed] [Google Scholar]
- 29.Fleischhauer K, Shaw BE, Gooley T, et al. ; International Histocompatibility Working Group in Hematopoietic Cell Transplantation. Effect of T-cell-epitope matching at HLA-DPB1 in recipients of unrelated-donor haemopoietic-cell transplantation: a retrospective study. Lancet Oncol. 2012;13(4):366-374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Pidala J, Lee SJ, Ahn KW, et al. Nonpermissive HLA-DPB1 mismatch increases mortality after myeloablative unrelated allogeneic hematopoietic cell transplantation. Blood. 2014;124(16):2596-2606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Fleischhauer K, Ahn KW, Wang HL, et al. Directionality of non-permissive HLA-DPB1 T-cell epitope group mismatches does not improve clinical risk stratification in 8/8 matched unrelated donor hematopoietic cell transplantation. Bone Marrow Transplant. 2017;52(9):1280-1287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Zino E, Vago L, Di Terlizzi S, et al. Frequency and targeted detection of HLA-DPB1 T cell epitope disparities relevant in unrelated hematopoietic stem cell transplantation. Biol Blood Marrow Transplant. 2007;13(9):1031-1040. [DOI] [PubMed] [Google Scholar]
- 33.Mack SJ, Cano P, Hollenbach JA, et al. Common and well-documented HLA alleles: 2012 update to the CWD catalogue. Tissue Antigens. 2013;81(4):194-203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Sanchez-Mazas A, Nunes JM, Middleton D, et al. Common and well-documented HLA alleles over all of Europe and within European sub-regions: A catalogue from the European Federation for Immunogenetics. HLA. 2017;89(2):104-113. [DOI] [PubMed] [Google Scholar]
- 35.Crocchiolo R, Zino E, Vago L, et al. ; Italian Bone Marrow Donor Registry. Nonpermissive HLA-DPB1 disparity is a significant independent risk factor for mortality after unrelated hematopoietic stem cell transplantation. Blood. 2009;114(7):1437-1444. [DOI] [PubMed] [Google Scholar]
- 36.Fleischhauer K, Fernandez-Viña MA, Wang T, et al. Risk associations between HLA-DPB1 T-cell epitope matching and outcome of unrelated hematopoietic cell transplantation are independent of HLA-DPA1. Bone Marrow Transplant. 2014;49(9):1176-1183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Crivello P, Heinold A, Rebmann V, et al. Functional distance between recipient and donor HLA-DPB1 determines nonpermissive mismatches in unrelated HCT. Blood. 2016;128(1):120-129. [DOI] [PubMed] [Google Scholar]
- 38.Jeras M. The role of in vitro alloreactive T-cell functional tests in the selection of HLA matched and mismatched haematopoietic stem cell donors. Transpl Immunol. 2002;10(2-3):205-214. [DOI] [PubMed] [Google Scholar]
- 39.van der Meer A, Joosten I, Schattenberg AV, de Witte TJ, Allebes WA. Cytotoxic T-lymphocyte precursor frequency (CTLp-f) as a tool for distinguishing permissible from non-permissible class I mismatches in T-cell-depleted allogeneic bone marrow transplantation. Br J Haematol. 2000;111(2):685-694. [DOI] [PubMed] [Google Scholar]
- 40.Kosmoliaptsis V, Jöris MM, Mallon DH, et al. Physiochemical disparity of mismatched HLA class I alloantigens and risk of acute GVHD following HSCT. Bone Marrow Transplant. 2015;50(4):540-544. [DOI] [PubMed] [Google Scholar]
- 41.Oudshoorn M, Doxiadis II, van den Berg-Loonen PM, Voorter CE, Verduyn W, Claas FH. Functional versus structural matching: can the CTLp test be replaced by HLA allele typing? Hum Immunol. 2002;63(3):176-184. [DOI] [PubMed] [Google Scholar]
- 42.Dankers MK, Heemskerk MB, Duquesnoy RJ, et al. HLAMatchmaker algorithm is not a suitable tool to predict the alloreactive cytotoxic T-lymphocyte response in vitro. Transplantation. 2004;78(1):165-167. [DOI] [PubMed] [Google Scholar]
- 43.Zhang L, Udaka K, Mamitsuka H, Zhu S. Toward more accurate pan-specific MHC-peptide binding prediction: a review of current methods and tools. Brief Bioinform. 2012;13(3):350-364. [DOI] [PubMed] [Google Scholar]
- 44.Duquesnoy RJ. HLAMMATCHMAKER: a molecularly based donor selection algorithm for highly alloimmunized patients. Transplant Proc. 2001;33(1-2):493-497. [DOI] [PubMed] [Google Scholar]
- 45.Duquesnoy RJ. HLAMatchmaker: a molecularly based algorithm for histocompatibility determination. I. Description of the algorithm. Hum Immunol. 2002;63(5):339-352. [DOI] [PubMed] [Google Scholar]
- 46.Dankers MK, Witvliet MD, Roelen DL, et al. The number of amino acid triplet differences between patient and donor is predictive for the antibody reactivity against mismatched human leukocyte antigens. Transplantation. 2004;77(8):1236-1239. [DOI] [PubMed] [Google Scholar]
- 47.Duquesnoy RJ. A structurally based approach to determine HLA compatibility at the humoral immune level. Hum Immunol. 2006;67(11):847-862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Duquesnoy R, Spellman S, Haagenson M, Wang T, Horowitz MM, Oudshoorn M. HLAMatchmaker-defined triplet matching is not associated with better survival rates of patients with class I HLA allele mismatched hematopoietic cell transplants from unrelated donors. Biol Blood Marrow Transplant. 2008;14(9):1064-1071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Marino SR, Lin S, Maiers M, et al. Identification by random forest method of HLA class I amino acid substitutions associated with lower survival at day 100 in unrelated donor hematopoietic cell transplantation. Bone Marrow Transplant. 2012;47(2):217-226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Lange V, Böhme I, Hofmann J, et al. Cost-efficient high-throughput HLA typing by MiSeq amplicon sequencing. BMC Genomics. 2014;15(1):63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Schöfl G, Lang K, Quenzel P, et al. 2.7 million samples genotyped for HLA by next generation sequencing: lessons learned. BMC Genomics. 2017;18(1):161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Niederwieser D, Baldomero H, Szer J, et al. Hematopoietic stem cell transplantation activity worldwide in 2012 and a SWOT analysis of the Worldwide Network for Blood and Marrow Transplantation Group including the global survey. Bone Marrow Transplant. 2016;51(6):778-785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Shaw BE, Robinson J, Fleischhauer K, Madrigal JA, Marsh SG. Translating the HLA-DPB1 T-cell epitope-matching algorithm into clinical practice. Bone Marrow Transplant. 2013;48(12):1510-1512. [DOI] [PubMed] [Google Scholar]
- 54.Bochtler W, Gragert L, Patel ZI, et al. A comparative reference study for the validation of HLA-matching algorithms in the search for allogeneic hematopoietic stem cell donors and cord blood units. HLA. 2016;87(6):439-448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Spellman SR, Eapen M, Logan BR, et al. ; Center for International Blood and Marrow Transplant Research. A perspective on the selection of unrelated donors and cord blood units for transplantation. Blood. 2012;120(2):259-265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Martin PJ, Levine DM, Storer BE, et al. Genome-wide minor histocompatibility matching as related to the risk of graft-versus-host disease. Blood. 2017;129(6):791-798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Tram K, Stritesky G, Wadsworth K, Ng J, Anasetti C, Dehn J. Identification of DPB1 permissive unrelated donors is highly likely. Biol Blood Marrow Transplant. 2017;23(1):81-86. [DOI] [PubMed] [Google Scholar]
- 58.Shaw BE, Gooley TA, Malkki M, et al. The importance of HLA-DPB1 in unrelated donor hematopoietic cell transplantation. Blood. 2007;110(13):4560-4566. [DOI] [PubMed] [Google Scholar]
- 59.Rutten CE, van Luxemburg-Heijs SA, Griffioen M, et al. HLA-DP as specific target for cellular immunotherapy in HLA class II-expressing B-cell leukemia. Leukemia. 2008;22(7):1387-1394. [DOI] [PubMed] [Google Scholar]
- 60.Herr W, Eichinger Y, Beshay J, et al. HLA-DPB1 mismatch alleles represent powerful leukemia rejection antigens in CD4 T-cell immunotherapy after allogeneic stem-cell transplantation. Leukemia. 2017;31(2):434-445. [DOI] [PubMed] [Google Scholar]
- 61.Yabe T, Azuma F, Kashiwase K, et al. ; Japanese Cord Blood Transplantation Histocompatibility Research Group. HLA-DPB1 mismatch induces a graft-versus-leukemia effect without severe acute GVHD after single-unit umbilical cord blood transplantation. Leukemia. 2018;32(1):168-175. [DOI] [PubMed] [Google Scholar]
- 62.Oran B, Saliba RM, Carmazzi Y, et al. Effect of nonpermissive HLA-DPB1 mismatches after unrelated allogeneic transplantation with in vivo T-cell depletion. Blood. 2018;131(11):1248-1257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Petersdorf EW, Malkki M, O’hUigin C, et al. High HLA-DP expression and graft-versus-host disease. N Engl J Med. 2015;373(7):599-609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Fleischhauer K. Immunogenetics of HLA-DP--a new view of permissible mismatches. N Engl J Med. 2015;373(7):669-672. [DOI] [PubMed] [Google Scholar]
- 65.Morishima S, Shiina T, Suzuki S, et al. ; Japan Marrow Donor Program. Evolutionary basis of HLA-DPB1 alleles affects acute GVHD in unrelated donor stem cell transplantation. Blood. 2018;131(7):808-817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Schöne B, Bergmann S, Lang K, et al. Predicting an HLA-DPB1 expression marker based on standard DPB1 genotyping: Linkage analysis of over 32,000 samples. Hum Immunol. 2018;79(1):20-27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Klasberg S, Lang K, Günther M, et al. Patterns of non-ARD variation in more than 300 full-length HLA-DPB1 alleles. Hum Immunol. 2018;S0198-8859(18)30156-3. [DOI] [PubMed] [Google Scholar]
- 68.Hollenbach JA, Madbouly A, Gragert L, et al. A combined DPA1∼DPB1 amino acid epitope is the primary unit of selection on the HLA-DP heterodimer. Immunogenetics. 2012;64(8):559-569. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.