

Key Points
Human blood CD8+ T cells express distinct levels of TCF1, defining quiescent vs effector populations.
TCF1-hi cells proliferate and uniquely self-renew following T-cell receptor stimulation to produce both TCF1-hi and TCF1-low cells.
Abstract
Expression of the transcription factor T-cell factor 1 (TCF1) identifies antigen-experienced murine CD8+ T cells that retain potential for lymphoid recirculation and the ability to self-renew while producing more differentiated effector cells. We found that CD8+ T cells in the blood of both healthy and chronically infected humans expressed TCF1 at 3 distinct levels: high (TCF1-hi), intermediate (TCF1-int), and low (TCF1-lo). TCF1-hi cells could be found within both the naive and memory compartments and were characterized by relative quiescence and lack of immediate effector function. A substantial fraction of TCF1-int cells were found among memory cells, and TCF1-int cells exhibited robust immediate effector functions. TCF1-lo cells were most enriched in effector memory cells that expressed the senescence marker CD57. Following reactivation, TCF1-hi cells gave rise to TCF1-lo descendants while self-renewing the TCF1-hi progenitor. By contrast, reactivation of TCF1-lo cells produced more TCF1-lo cells without evidence of de-differentiating into TCF1-hi cells. Flow cytometric analyses of TCF1 expression from patient specimens may become a useful biomarker for adaptive immune function in response to vaccination, infection, autoimmunity, and cancer.
Visual Abstract
Introduction
Memory lymphocytes have the capacity to self-renew and maintain lifelong immunity to repeat pathogen exposure. Collapse of self-renewal is a hallmark of immune failure in chronic infection, cancer, and aging. Memory CD8+ T cells are organized in hierarchical subsets possessing progressively increased effector characteristics, including central memory (TCM), effector memory (TEM), and effector memory CD45RA positive (TEMRA). TCM cells share features with naive T cells, including circulation through secondary lymphoid organs and expression of CCR7 and CD62L. TCM cells retain a greater proliferative capacity than TEM cells but exhibit less function immediately after restimulation.1 TEMRA cells are thought to be terminally differentiated. While the identification of memory cell subsets by CCR7 and CD45RA expression provided insights into the organization of human immunological memory, these markers are heterogeneous and numerous subpopulations exist.
T-cell factor 1 (TCF1) is a critical regulator of T-cell development in mice.2,3 In acute infections, active progenitor cells that express TCF1 and have divided more than 4 times can produce TCF1-lo, irreversibly determined effector cells while self-renewing TCF1+ cells through asymmetric divisions.4,5 In persistent low-level infections, active, TCF1+, self-renewing progenitors continuously reseed the effector cell pool, presumably undergoing periodic replacement by recruitment of quiescent central memory cells.6,7 In chronic active infections, self-renewing TCF1+ progenitors replenish the effector cell pool and can be mobilized by immune checkpoint inhibitors.8-11 Expression of TCF1 in human CD8+ T-cell populations has not been fully explored.12 Here, we show that TCF1 expression marks subsets of memory CD8+ T cells in blood with increased self-renewal properties, which could substantially refine traditional CCR7/CD45RA-based classifications for immune profiling and lead to better understanding of human T-cell memory.
Study design
Human samples
Healthy adult human peripheral blood mononuclear cells were obtained from donor blood packs from the New York Blood Center. Donors with chronic hepatitis C virus infection were recruited under a Columbia University institutional review board–approved protocol (IRB-AAAP4004), and peripheral blood samples were obtained by venipuncture. All human participants gave written informed consent.
Flow cytometry
Samples were stained according to standard flow cytometry protocols. Antibodies used include CD3 (OKT3, eBioscience), CD4 (RPA-T4, BioLegend), CD8 (RPA-T8, Tonbo Biosciences), CD27 (O323, BioLegend), CCR7 (G043H7, BioLegend), CD107a (ebioH4A3, eBioscience), CD45RA (HI100, Tonbo Biosciences), CD127 (R34-34, Tonbo Biosciences), CD57 (TB01, eBioscience), Eomes (WD1928, eBioscience), LEF1 (C12A5, Cell Signaling Technology), TCF1 (C63D9, Cell Signaling Technology), T-bet (4B10, BioLegend), interferon γ (IFN-γ) (4S.B3, eBioscience), and granzyme B (GB11, BD Pharmingen). For cytokine staining, cells were stimulated with 50 ng/mL 12-O-tetradecanoylphorbol 13-acetate/1 μg/mL ionomycin for 4 hours in the presence of Golgi Plug (BD Biosciences).
T-cell differentiation assay
Cells from the indicated populations were fluorescence-activated cell sorter (FACS) sorted and labeled with cell proliferation dye (CTV, Thermo Fisher). 1 × 105 cells were plated per well in 96-well plates, coated with 1 μg/mL anti-CD3/CD28 antibodies and 1 μg/mL human Fc-ICAM (R&D Systems), and cultured in RPMI supplemented with 10% fetal bovine serum and 100 IU/mL interleukin-2 for 4 or 5 days.
Results and discussion
Hierarchical TCF1 expression by CD8+ T-cell subsets
Peripheral blood from healthy donors and patients chronically infected with hepatitis C virus was examined to determine the range of TCF1 expression across CD8+ T-cell subsets in different immunological states. For both healthy and chronically infected individuals, CD4+/CD8+ T-cell ratios and distributions of naive (CCR7+CD45RA+), TCM (CCR7+CD45RA−), TEM (CCR7−CD45RA−), and TEMRA (CCR7−CD45RA+) cells were similar (Figure 1A). Naive cells expressed the highest levels of TCF1 protein, whereas TCM expressed higher levels than TEM and TEMRA (Figure 1A). These results indicate that there is substantial heterogeneity in TCF1 expression by memory cells and that heightened TCF1 expression is characteristic of more quiescent cells.
Figure 1.

Memory CD8+T-cell heterogeneity associated with TCF1 expression. (A) Characterization of T-cell subsets in peripheral blood of healthy and chronically infected humans. CD4+/CD8+ T-cell ratios in donor subjects (top left). Identification of CD8+ T-cell subsets in peripheral blood: naive (CCR7+CD45RA+), TCM (CCR7+CD45RA−), TEM (CCR7−CD45RA−), and TEMRA (CCR7−CD45RA+) (bottom left). Distribution of naive and memory CD8+ T-cell subsets (top right). Subset frequencies were not significantly associated with infection status (2-way analysis of variance [ANOVA]). Steady-state expression (mean fluorescence intensity [MFI]) of TCF1 by CD8+ T cell subsets (bottom right). (B) Hierarchical expression of TCF1. Identification of TCF1-hi, TCF1-int, and TCF1-lo subsets within the bulk CD8+ T-cell compartment (left). Distribution of TCF1-subsets within traditional naive, TCM, TEM, and TEMRA populations (right). (C) Expression of transcription factors Eomes, LEF1, and T-bet by TCF1-hi, TCF1-int, and TCF1-lo populations in healthy donors. *P < .05. **P < .01. ***P < .001. Repeated measures 1-way ANOVA with Sidak correction. (D) Reciprocal expression of TCF1 and CD57 across CD8+ T-cell populations in healthy donors. Gate denotes frequency of the TCF1-lo CD57+ population. Results are representative of 5 independent donors. (E) Quiescent phenotype of TCF1-hi cells. Expression of CD127 (IL7-R), CD27 (costimulatory receptor), and CD57 (senescence marker) by TCF1 subsets in healthy donors. *P < .05; ***P < .001; n.s., not significant (repeated-measures 1-way ANOVA with Sidak correction). HCV, hepatitis C virus.
Examination of total CD8+ T cells revealed 3 distinct levels of TCF1 expression: high (TCF1-hi), intermediate (TCF1-int), and low (TCF1-lo) (Figure 1B). Expression of TCF1 was reciprocal to the transcription factor T-bet, which also occupied 3 distinct levels. Cells with TCF1-hi, TCF1-int, or TCF1-lo expression were represented at varying frequencies within each of the naive, TCM, TEM, and TEMRA compartments (Figure 1B). Similar distributions were detected in chronically infected patients, indicating preservation of hierarchical TCF1 expression despite effects of chronic infection on bystander immunity.13
Examination of other transcription factors governing T-cell differentiation revealed that TCF1-int cells had the highest expression of Eomes, which regulates memory cell formation in mice (Figure 1C).14 Lymphoid-enhancer-binding factor 1 (LEF1), which also regulates memory cell formation15 and is part of the same family of Wnt pathway factors as TCF1, was expressed coordinately with TCF1 (Figure 1C). T-bet expression, which is associated with effector differentiation, was expressed inversely to TCF1 (Figure 1C).
We next assessed the surface marker phenotype of cells expressing different levels of TCF1. TCF1 expression was reciprocal to CD57, a marker of senescence and terminal differentiation (Figure 1D-E).16 Notably, the TEM and TEMRA compartments, conventionally regarded as more differentiated cells, contained readily detectable subpopulations of TCF1+ CD57− cells. Both TCF1-hi and TCF1-int cells exhibited high CD127 expression, associated with long-lived memory populations (Figure 1E).17 TCF1-hi cells also showed evidence of quiescence, including greater maintenance of the costimulatory receptor CD27 compared with TCF1-int and TCF1-lo cells (Figure 1E). Key markers associated with effector and memory formation in mice are thus differentially expressed by human CD8+ T cells and associated with memory cell heterogeneity.
Functional differences in CD8+ T cells with distinct TCF1 expression
To determine whether TCF1 expression is associated with functionally distinct memory cell populations, we assessed cytokine production and cytotoxicity. TCF1-int cells had the largest capacity for immediate cytokine production, producing greater levels of IFN-γ than TCF1-hi or TCF1-lo populations (Figure 2A). TCF1-lo cells expressed high levels of the cytotoxic granule component granzyme B, although they produced less IFN-γ and underwent less efficient degranulation than TCF1-int cells following stimulation (Figure 2A). Consistent with a quiescent and less differentiated phenotype, TCF1-hi cells had the least immediate effector function (Figure 2A).
Figure 2.

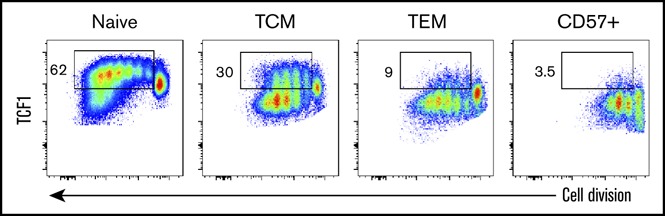
Self-renewal of TCF1-hi CD8+T cells. Distinct functional properties of TCF1 subsets. (A) Peripheral blood mononuclear cells stimulated with 12-O-tetradecanoylphorbol 13-acetate/ionomycin and analyzed for cytokine (IFN-γ) and cytolytic molecule (granzyme B) expression and degranulation (CD107a). *P < .05; **P < .01 (repeated-measures 1-way ANOVA with Sidak correction). (B) TCF1-hi cells self-renew following T-cell receptor (TCR) stimulation. The histogram on the left shows steady-state TCF1 expression by FACS-sorted CCR7+CD45RA+CD57− (naive), CCR7+CD45RA−CD57− (TCM), CCR7−CD45RA−CD57− (TEM), and CCR7−CD57+ populations. Data are representative of 5 separate donors. FACS-sorted naive, TCM, TEM, and CCR7−CD57+CD8+ T cells were TCR stimulated (right). Cell division vs TCF1 expression at day 4 is shown. Gate denotes TCF1-hi population. Quantification of TCF1-hi population frequency (bottom left). *P < .05; ****P < .0001 (1-way ANOVA with the Sidak multiple comparisons test). (C) Earlier loss of TCF1-hi cells from TCM following activation. Naive and TCM populations were sorted and stimulated as in panel B. Frequency of TCF1-hi cells was quantified on day 4 for naive and TCM starting populations at each division number. *P < .05 (Student t test with Holm-Sidak correction for multiple comparisons). (D) Stepwise differentiation of TCF1-hi and TCF1-lo populations following activation of naive CD8+ T cells. Gating of undivided (P1), TCF1-hi (P2), and TCF1-lo (P3) populations at day 4 of culture following TCR stimulation of naive cells (top left). The remaining panels show representative histograms of CD27, CCR7, CD127, T-bet, and Eomes expression by each population. Activated CD57+ cells were included as a positive control for Eomes induction. Plots are representative of 3 independent donors.
Self-renewal of TCF1-hi CD8+ T-cell populations
In mice, silencing of TCF1 marks irreversible CD8+ effector T-cell differentiation and loss of progenitor cell self-renewal.4,5 Therefore, we assessed TCF1 expression dynamics following T-cell activation in populations that were TCF1-hi (naive or TCM), TCF1-int/lo (TEM), or TCF1-lo (CCR7−CD57+) (Figure 2B). When naive cells were stimulated, a TCF1-lo population emerged after several divisions, while a population of TCF1-hi cells was maintained among the proliferating cells at each division (Figure 2B). Activated TCM also maintained a population of TCF1-hi cells at each division. TCM cells, however, produced TCF1-lo progeny at earlier divisions and higher frequencies than naive cells (Figure 2B-C), in agreement with prior mouse experiments showing earlier and increased repression of TCF1 by memory T cells following pathogen rechallenge.4,5 In addition, the human TCM-phenotype (CCR7+CD45RA−) population contains a small proportion of cells with lower TCF1 expression than the naive samples (Figures 1B-D and 2B), which may further explain why TCM cell cultures yield more TCF1-lo progeny after stimulation.
TEM cells were predominantly TCF1-int before stimulation, whereas CCR7−CD57+ cells were mostly TCF1-lo (Figure 2B). TEM cells exhibited more proliferative capacity than CD57+ cells but lost TCF1 expression after the first division and did not maintain a TCF1-hi or TCF1-int population. Similarly, following activation and proliferation, CD57+ TCF1-lo cells produced more TCF1-lo descendants without evidence of de-differentiation18,19 into TCF1-hi progeny (Figure 2B). Prior to division, naive cells that had been activated for 4 days maintained high levels of CD27, CCR7, and CD127, as well as low levels of Eomes (Figures 2D). All proliferating cells downregulated CCR7, but proliferating TCF1-lo cells downregulated CD127 and CD27 to a greater extent than proliferating TCF1-hi cells (Figure 2D). Under these conditions, proliferating TCF1-hi and TCF1-lo cells had comparably high levels of T-bet and comparably low levels of Eomes expression (Figure 2D). Together, these results suggest that dividing TCF1-hi cells are more activated than quiescent naive and memory populations and that they function as an intermediate, self-renewing progenitor of more differentiated TCF1-lo cells.
Lineage transfer studies using TCF1 reporter mice during acute infections indicated that a substantial fraction of previously presumed memory precursor cells (KLRG1-lo) are actually TCF1-lo and incapable of re-expressing TCF1 under physiological conditions,5 consistent with our replating experiments using human CD8+ T cells (Figure 2B). Instead, active TCF1+ progenitor cells continually produce TCF1-lo effector cells when antigen is present.5-9 When antigen is cleared, TCF1+ progenitors, but not TCF1-lo effector cells, contribute to the quiescent, TCF1+ central memory cell pool.5 Profiling peripheral blood TCF1 expression, along with traditional markers of effector/memory cell differentiation, may become a useful biomarker for cancer immunotherapy, vaccination, and other immunologic conditions.19,20 Pharmacologic manipulation of T-cell self-renewal may become a novel strategy for combating infection and malignancy.
Acknowledgments
The authors are grateful to Nyanza Rothman for assistance and the anonymous blood donors and study participants who made this work possible.
This study was supported by the National Institutes of Health, National Institute of Allergy and Infectious Diseases (grants AI113365 and AI076458) (S.L.R.) and the Charles H. Revson Foundation.
Authorship
Contribution: R.K. performed experiments, analyzed data, and wrote the manuscript; A.M.M. coordinated patient recruitment and acquisition of blood samples; and S.L.R. analyzed data, wrote the manuscript, and supervised the project at all stages.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Steven L. Reiner, Department of Microbiology and Immunology and Department of Pediatrics, Roy and Diana Vagelos College of Physicians and Surgeons, Columbia University, 701 West 168th St, Hammer Building, Room 912, New York, NY 10032; e-mail: sr2978@cumc.columbia.edu.
References
- 1.Mahnke YD, Brodie TM, Sallusto F, Roederer M, Lugli E. The who’s who of T-cell differentiation: human memory T-cell subsets. Eur J Immunol. 2013;43(11):2797-2809. [DOI] [PubMed] [Google Scholar]
- 2.Germar K, Dose M, Konstantinou T, et al. T-cell factor 1 is a gatekeeper for T-cell specification in response to Notch signaling. Proc Natl Acad Sci USA. 2011;108(50):20060-20065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Weber BN, Chi AW-S, Chavez A, et al. A critical role for TCF-1 in T-lineage specification and differentiation. Nature. 2011;476(7358):63-68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lin W-HW, Adams WC, Nish SA, et al. Asymmetric PI3K signaling driving developmental and regenerative cell fate bifurcation. Cell Reports. 2015;13(10):2203-2218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Lin WW, Nish SA, Yen B, et al. CD8+ T lymphocyte self-renewal during effector cell determination. Cell Reports. 2016;17(7):1773-1782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Chu HH, Chan SW, Gosling JP, et al. Continuous effector CD8(+) T cell production in a controlled persistent infection is sustained by a proliferative intermediate population. Immunity. 2016;45(1):159-171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Delpoux A, Michelini RH, Verma S, et al. Continuous activity of Foxo1 is required to prevent anergy and maintain the memory state of CD8+ T cells. J Exp Med. 2018;215(2):575-594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Im SJ, Hashimoto M, Gerner MY, et al. Defining CD8+ T cells that provide the proliferative burst after PD-1 therapy. Nature. 2016;537(7620):417-421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Utzschneider DT, Charmoy M, Chennupati V, et al. T cell factor 1-expressing memory-like CD8(+) T cells sustain the immune response to chronic viral infections. Immunity. 2016;45(2):415-427. [DOI] [PubMed] [Google Scholar]
- 10.Leong YA, Chen Y, Ong HS, et al. CXCR5(+) follicular cytotoxic T cells control viral infection in B cell follicles. Nat Immunol. 2016;17(10):1187-1196. [DOI] [PubMed] [Google Scholar]
- 11.Wu T, Ji Y, Moseman EA, et al. The TCF1-Bcl6 axis counteracts type I interferon to repress exhaustion and maintain T cell stemness. Sci Immunol. 2016;1(6):eaai8593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Willinger T, Freeman T, Herbert M, Hasegawa H, McMichael AJ, Callan MF. Human naive CD8 T cells down-regulate expression of the WNT pathway transcription factors lymphoid enhancer binding factor 1 and transcription factor 7 (T cell factor-1) following antigen encounter in vitro and in vivo. J Immunol. 2006;176(3):1439-1446. [DOI] [PubMed] [Google Scholar]
- 13.Stelekati E, Shin H, Doering TA, et al. Bystander chronic infection negatively impacts development of CD8(+) T cell memory. Immunity. 2014;40(5):801-813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Pearce EL, Mullen AC, Martins GA, et al. Control of effector CD8+ T cell function by the transcription factor Eomesodermin. Science. 2003;302(5647):1041-1043. [DOI] [PubMed] [Google Scholar]
- 15.Zhou X, Xue H-H. Cutting edge: generation of memory precursors and functional memory CD8+ T cells depends on T cell factor-1 and lymphoid enhancer-binding factor-1. J Immunol. 2012;189(6):2722-2726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Brenchley JM, Karandikar NJ, Betts MR, et al. Expression of CD57 defines replicative senescence and antigen-induced apoptotic death of CD8+ T cells. Blood. 2003;101(7):2711-2720. [DOI] [PubMed] [Google Scholar]
- 17.Mazzucchelli R, Durum SK. Interleukin-7 receptor expression: intelligent design. Nat Rev Immunol. 2007;7(2):144-154. [DOI] [PubMed] [Google Scholar]
- 18.Youngblood B, Hale JS, Kissick HT, et al. Effector CD8 T cells dedifferentiate into long-lived memory cells. Nature. 2017;552(7685):404-409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Akondy RS, Fitch M, Edupuganti S, et al. Origin and differentiation of human memory CD8 T cells after vaccination. Nature. 2017;552(7685):362-367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Huang AC, Postow MA, Orlowski RJ, et al. T-cell invigoration to tumour burden ratio associated with anti-PD-1 response. Nature. 2017;545(7652):60-65. [DOI] [PMC free article] [PubMed] [Google Scholar]