

Abstract
Reproductive disease and fertility issues have dramatically increased in the human population over the last several decades, suggesting environmental impacts. Epigenetics provides a mechanistic link by which an organism can respond to environmental factors. Interestingly, environmentally induced epigenetic alterations in the germ line can promote aberrant gene expression and disease generationally. Environmentally induced epigenetic transgenerational inheritance is defined as germ-line transmission of altered epigenetic information between generations in the absence of continued environmental exposures. This form of nongenetic inheritance has been shown to directly influence fertility and reproductive disease. This review describes the studies in a variety of species that impact reproductive disease and abnormalities. Observations suggest serious attention be paid to the possibility that ancestral exposures to environmental insults promotes transgenerational inheritance of reproductive disease susceptibility. Environmentally induced epigenetic transgenerational inheritance appears to be an important contributing factor to reproductive disease in many organisms, including humans.
Keywords: developmental biology, environment, epigenetics, genomics
Introduction
Fertility issues have been increasing in human populations for decades. In men, there have been decreases in sperm count [1–4], increases in testicular cancer [5], and increases in genital abnormalities [6]. Fecundity rates in men and women in both developing and industrial countries have dramatically decreased in recent years [7, 8]. Although economics, social trends, and governmental policies certainly contribute to a decreased birthrate, attention must be paid to the role of environmental toxicants and other exposures in promoting reproductive disease [9–11]. Correlations have been made between environmental exposures and prostate and mammary disease [12, 13], semen quality [1, 14–16], reproductive developmental abnormalities [17–23], polycystic ovarian syndrome [24], and endometriosis [25, 26]. Therefore, the environment has a significant effect on fertility and reproductive disease etiology [27].
The majority of environmental factors and toxicants do not have the ability to promote DNA mutations [28]. Therefore, molecular mechanisms other than alterations in DNA sequence must mediate the ability of environmental factors to promote reproductive disease. Certainly, there are DNA mutations that are associated with specific disease. For example, polymorphisms in the FSH beta promoter result in low sperm counts [29], and fragile X mental retardation gene 1 CGG repeat abnormalities are associated with the oocyte follicle loss of primary ovarian insufficiency [30]. However, genome-wide association studies have generally shown that less than 2% of specific adult-onset diseased populations have a correlated DNA sequence mutation [31, 32]. In addition, the genetic background in human populations is essentially static, while increases in disease disorders and infertility are dramatically increasing [12]. Therefore, environmental exposures must act primarily through epigenetic mechanisms to promote reproductive disease [33, 34].
The term “epigenetics” was coined by Dr. Conrad Waddington, University of Edinburgh, in the 1940s to describe gene-environment interactions that could not be explained with classic genetics [35]. Using a more recent mechanistic definition, “epigenetics” is defined as molecular factors/processes around the DNA that regulate genome activity independent of DNA sequence and that are mitotically stable [33]. In the 1970s, the first epigenetic molecular mark was identified as being DNA methylation, in which a small chemical (methyl) group is attached to DNA at primarily the cytosine base in animals [36, 37]. In the 1990s, the histone proteins around which DNA is wrapped were also found to be chemically modified to alter gene expression. In the 2000s, noncoding RNA molecules were identified that can act as epigenetic factors [38]. The coiling, looping, and general structure of DNA, termed “chromatin structure,” is also an epigenetic factor [39]. Therefore, the currently known epigenetic molecular processes are DNA methylation, histone modifications, functional noncoding RNA, and chromatin structure [27]. All of these processes are a normal part of physiology and cell differentiation and are a part of the cellular machinery that regulates gene expression. Epigenetic processes are often the mechanistic link by which an organism can respond to its environment and change gene expression. Abnormal epigenetic mechanisms can result in alterations in gene expression patterns. These epigenetic abnormalities can lead to aberrant physiology and disease.
Epigenetic Changes Associated with Reproductive Disease
Previous studies have shown that many cases of reproductive disease are accompanied by changes in the epigenome. In males, the incidence of germ cell tumors is correlated with epigenetic disruption [40]. The sperm of infertile men has been shown to have abnormal DNA methylation, histone modifications, and an altered retention of histones in contrast to the usual replacement of histones with protamines [41–43]. In adult rat testes that show an increase in germ cell apoptosis, the isolated Sertoli cells showed changes in the DNA methylation at specific gene promoters as well as accompanying changes in their transcriptome [44]. Prostate tumor growth and invasiveness have been linked to abnormal expression of the polycomb repressor complex that induces changes in chromatin structure that are needed for the formation of heterochromatin [45].
In females, several reproductive diseases have also been associated with epigenetic abnormalities. The occurrence of endometriosis has been linked to the epigenetic deregulation of “endometriosis susceptibility” genes [46–48]. Women exposed in utero to diethylstilbesterol develop cervical malformations. These malformations are correlated with DNA methylation changes of Hox genes [49, 50]. Epigenetic changes may underlie some of the decrease in oocyte quality that occur as women age [51]. Embryo growth and development can be negatively impacted if the dietary methyl donors and cofactors required for normal DNA and histone methylation processes are deficient [52]. In rat ovaries that show a loss of oocytes, isolated granulosa cells showed changes in the DNA methylation at specific gene promoters as well as accompanying changes in their transcriptome [53]. Taken together, there is strong evidence that epigenetic abnormalities are linked to reproductive disease.
Epigenetic Transgenerational Inheritance
Environmentally induced epigenetic transgenerational inheritance is defined as the germ-line transmission of altered epigenetic information between generations in the absence of continued environmental exposures [10, 54]. These epigenetic germ-line alterations will subsequently affect gene expression and epigenetic programming patterns in somatic tissues [10, 44] (Fig. 1). The first mammalian example of an environmentally induced epigenetic transgenerational inheritance process was described in 2005 using an early developmental exposure to the endocrine disruptor vinclozolin [55]. Vinclozolin is an agricultural fungicide with antiandrogenic activity widely used in fruit and vegetable crops around the world [56]. Exposure of pregnant rats to vinclozolin produced increased apoptosis in spermatogenic cells, which was observed in each of the four generations after this initial exposure [55, 57, 58] (Fig. 1). The mechanism involved in the transgenerational transmission of these altered phenotypes was an induced alteration in the sperm epigenome that was observed three generations after the developmental exposure to vinclozolin [55, 59, 60].
Fig. 1.
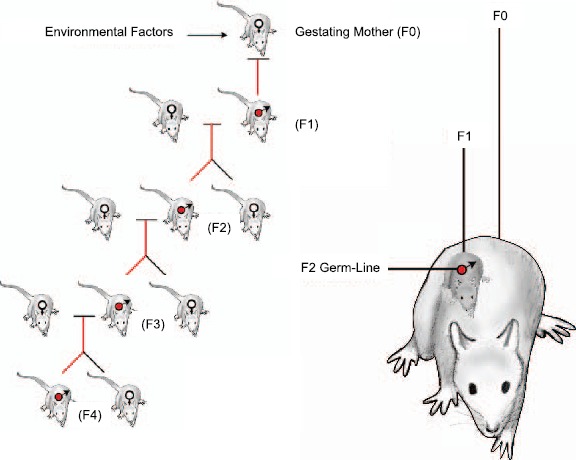
Environmentally induced epigenetic transgenerational inheritance through male germ line. Exposure of the F0 generation gestating female, F1 generation fetus, and germ line within the F1 generation fetus that will generate the F2 generation. Therefore, the F3 generation is the first transgenerational generation not directly exposed. Figure modified from Skinner [61].
In considering transgenerational phenomena, it is important to distinguish between direct exposure effects versus germ-line (sperm or egg)-mediated transgenerational events. When a gestating F0 generation female is exposed the F0 generation female, the F1 generation fetus and the germ cell (sperm or egg) that is inside the fetus and that will produce the F2 generation are all directly exposed (Fig.1). Any effects in the F0, F1, and F2 generations may be due to direct exposure toxicity or to environmentally induced epigenetic changes in the directly exposed cells. Examination of the F3 generation (great grand-offspring) is needed to determine if a transgenerational phenomenon has occurred since the F3 generation has had no direct exposure effects [61]. In contrast, in the event an adult male or nonpregnant female is exposed, the F0 generation adult and the germ cells that will generate the F1 generation are directly exposed such that examination of the F2 generation (grand-offspring) is required to demonstrate a transgenerational phenomenon [61].
In order for a transgenerational effect to occur, it is critical that the germ-line epigenome be altered to allow for transmission to future generations. There are several critical stages of germ cell development where dramatic epigenetic programming occurs [62, 63]. These periods in germ cell development and epigenetic programming represent critical windows of sensitivity to environmental factors [10, 27]. The first is when the stem cells (precursor cells) for the germ cells, called primordial germ cells, develop and migrate during the time of gonadal sex determination at the onset of testis and ovary development. The DNA methylation of those primordial germ cells is predominantly erased, and then subsequently remethylation is initiated during testis and ovary maturation (Fig. 2). The second period is when the sperm and egg come together at fertilization and the DNA contributed by the sperm and egg again are demethylated to create the embryonic stem cells [62, 63] (Fig. 2). This epigenetic programming allows the embryonic cells to develop pluripotency. Interestingly, when exposures to toxicants or abnormal nutrition occur during gonadal sex determination, the epigenetic programming or DNA methylation of the germ cell can become reprogrammed and transmit altered epigenetic information transgenerationally to subsequent generations [10, 55].
Fig. 2.
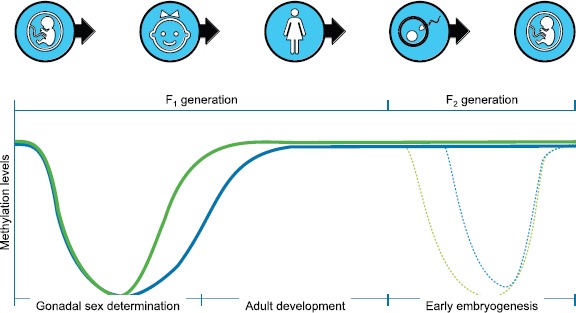
Epigenetic programming of DNA methylation levels during development during gonadal sex determination, fertilization, and embryonic development. The green line is the male and the blue line the female developmental pattern. Figure modified from Jirtle and Skinner [27].
Transgenerational Epigenetic Reproductive Disease
There are now several studies in mammals reporting the occurrence of environmentally induced transgenerational epigenetic inheritance of reproductive disease. Table 1 lists environmental exposures that can result in inheritance of epigenetic transgenerational abnormalities and highlights those that promote reproductive disease states. As shown, the majority of studies have associated epigenetic changes reported. In male rat testis, abnormalities observed included decreased sperm counts and impaired motility, which have been shown to be transgenerationally inherited after ancestral exposure to vinclozolin [55, 58] and DDT [64, 65]. In mice, similar transgenerational decreases in sperm production were seen after ancestral exposure to phthalates, which are components of plastics, cosmetics, and other products [66]. An increase in germ cell apoptosis was seen in rat testes transgenerationally after ancestral exposure to vinclozolin [55], DDT [64], and a jet fuel/hydrocarbon mixture [67]. In mice, transgenerational inheritance of testis germ cell apoptosis has also been reported after ancestral exposure to vinclozolin [59, 65]. Other testis abnormalities that have shown transgenerational epigenetic inheritance include an increased incidence of histologically detected seminiferous tubule atrophy, tubule vacuoles and germ cell agenesis, which occured after ancestral exposure of rats to vinclozolin [58], a mixture of the plasticizers bisphenol A (BPA) and phthalates [68], and the combination of the insecticide permethrin and the insect repellent DEET [69]. In mice, similar histologic seminiferous tubule defects were seen after ancestral exposure to phthalates [66] and the industrial environmental contaminant benzo[a]pyrene [70]. In rat testes that exhibit a transgenerational increase in germ cell apoptosis after ancestral vinclozolin exposure, isolated Sertoli cells show correlated alterations in both DNA methylation (epigenome) and RNA expression (transcriptome) [44].
Table 1.
Environmental exposures that induce transgenerational reproductive disease phenotypes.
| Toxicants | Reproductive disease | References |
|---|---|---|
| Vinclozolin | Decreased sperm count, testis apoptosis, testis abnormalities,a prostate abnormalities, oocyte loss, ovarian cysts, altered mate selection. Epigenetic changes observed. | [51, 55–57, 63, 69, 78] |
| Methoxychlor | Ovarian cysts. Epigenetic changes observed. | [55, 75, 80] |
| TCDD/dioxin | Puberty onset, oocyte loss, ovarian cysts, fertility defect.b Epigenetic changes observed. | [72, 77, 78] |
| Plastics mixture (bisphenol-A, phthalate-DEHP, and DBP) | Testis abnormalities, puberty onset, oocyte loss, ovarian cysts. Epigenetic changes observed. | [67, 68] |
| Jet fuel (JP8) | Testis apoptosis, oocyte loss. Epigenetic changes observed. | [65, 74] |
| Permethrin and DEET | Testis abnormalities, puberty onset, oocyte loss, ovarian cysts. Epigenetic changes observed. | [69] |
| DDT | Decreased sperm count, testis apoptosis, ovarian cysts. Epigenetic changes observed. | [63, 64] |
| Bisphenol A | Decreased sperm count, fertility defect | [79, 89, 90] |
| Phthalates | Decreased sperm count, testis abnormalities, puberty onset, fertility defect | [66] |
| Tributyltin | [91] | |
| Benzo[a]pyrene | Testis abnormalities | [70] |
| Other types exposures | ||
| Folate (nutrition) | [92] | |
| High-fat diet (nutrition) | [93, 94] | |
| Caloric Restriction (nutrition) | [95–98] | |
| Temperature and drought (plant flowering and health) | Abnormal flowering, fertility defect. Epigenetic changes observed. | [99–102] |
| Stress (behavioral) | [103, 104] | |
| Smoking (health) | [105, 106] | |
| Alcohol (health) | [107] |
Includes seminiferous tubule atrophy, tubule vacuoles, and germ cell agenesis.
Fertility defect indicates reduced numbers of offspring.
Prostate gland abnormalities, including prostate epithelial hyperplasia and epithelial atrophy, have been transmitted transgenerationally in rats and mice after ancestral exposure to vinclozolin. This was accompanied by transgenerational changes in RNA expression in prostate epithelial cells [59, 71].
Ancestral exposure to environmental toxicants can induce transgenerational changes to the timing of the onset of puberty. In rats, exposure of pregnant F0 generation females to a mixture of BPA and phthalates [68], a combination of permethrin and DEET [69], or the industrial contaminant dioxin [72] induced changes in the time of puberty onset in the unexposed F3 generation. Similar transgenerational changes in the onset of puberty were seen in mice after ancestral exposure to phthalates [66].
A female reproductive disease that has shown transgenerational epigenetic inheritance is a decreased follicle pool associated with decreases in total oocytes per ovary [67]. Follicle loss such as this is a major component of the human disease condition primary ovarian insufficiency, which results in premature menopause [73]. Rats have shown transgenerational epigenetic inheritance of decreases in the follicle pool after ancestral exposures to jet fuel hydrocarbons [74], a mixture of BPA and phthalates [68], dioxin [72], permethrin/DEET [69], and vinclozolin [53]. Similarly, an increase in the incidence of ovarian cysts was transmitted to the F3 generation after ancestral exposure to the pesticide methoxychlor [75], DDT [64], a mixture of plasticizers [68], dioxin [72], permethrin/DEET [69], and vinclozolin [53]. In women, polycystic ovarian syndrome is a common problem resulting in infertility and is increasing in frequency [76]. In rat ovaries that exhibit a transgenerational decrease in their follicle pool and an increase in ovarian cysts after ancestral vinclozolin exposure, isolated granulosa cells show correlated alterations in both DNA methylation and RNA expression [53].
Several studies have documented a transgenerational decrease in fertility as measured by the number of offspring produced following ancestral exposure to environmental toxicants. Pregnant F0 generation mice exposed to dioxin had female progeny of the F1, F2, F3, and F4 generations with an increased incidence of preterm birth and decreased fertility [77]. Dioxin also was shown to decrease fertility transgenerationally in zebrafish [78]. Exposure of pregnant mice to phthalates also induced the transgenerational inheritance of reduced fertility [66]. Similarly, pregnant rats exposed perinatally to BPA gave birth to offspring with significantly impaired spermatogenesis and fertility, and there was transgenerational inheritance of this fertility defect in male progeny [79].
Ancestral exposure to toxicants can induce transgenerational inheritance of an altered mate preference that is associated with sexual selection. The exposure of F0 generation pregnant rats to vinclozolin resulted in the transgenerational F3 generation females showing an altered mate preference [80]. This demonstrates that transgenerational inheritance of epigenetically mediated changes could affect sexual selection and evolution [81].
Epigenetic transgenerational inheritance of disease susceptibility can be induced by ancestral exposure to several different environmental toxicants (Table 1). The question is raised whether different toxicants promote different epigenetic changes that are passed transgenerationally. A study in rats addressed this question by comparing the DNA methylation patterns in the sperm of the F3 generation progeny of F0 generation gestating females that had been exposed to either jet fuel hydrocarbons, dioxin, a plastics mixture, or a pesticide mixture [67]. Interestingly, the sites of changes in DNA methylation, known as differentially methylated regions (DMR), formed a unique pattern with each different ancestral toxicant exposure (Fig. 3). No transgenerational DMR was common to all the exposures. This suggests that DNA methylation patterns could be used as biomarkers to detect ancestral exposure to environmental toxicants. In the future, individuals so tested might then be able to predict susceptibility to later-life correlated disease. Further research is needed to confirm the accuracy of such biomarkers and determine what effect variables such as toxicant dose have on inherited epigenetic alterations.
Fig. 3.
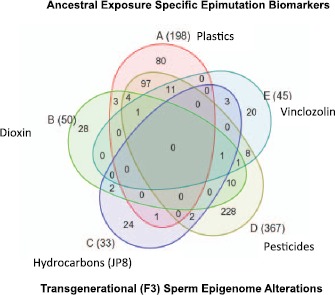
Ancestral exposure specific epimutation biomarkers. Transgenerational F3 generation sperm differential DNA methylation regions (epimutations) with the total listed next to exposure in brackets and Venn diagram showing overlap between the exposure epimutations. Figure modified from Manikkam et al. [67].
Biological Impacts and Mechanisms
Studies have demonstrated that epigenetic transgenerational inheritance of an increased susceptibility for reproductive disorders can occur after ancestral exposure to an environmental insult. Interestingly, transgenerational inheritance of many of the reproductive disorders described in this review may be induced by exposure to any one of several different environmental toxicants or factors (Table 1). For example, transgenerational inheritance of ovarian follicle loss in rats can be induced by ancestral exposure to jet fuel hydrocarbons, BPA, phthalates, dioxin, permethrin, DEET, and vinclozolin [53, 67]. This phenomenon of several toxicants inducing the same transgenerational reproductive disease occurs even though the epigenetic changes induced are different for each toxicant. Therefore, one would expect that different genes would have altered regulation for each toxicant. This phenomenon may be due to the systems biology of the reproductive tissue or organ system that is transgenerationally affected. If expression of enough of the genes in a vital signaling network is disrupted, then a particular disease manifests, regardless of the specific genes affected.
From Table 1, it may be noted that many of the environmental toxicants that can promote epigenetic transgenerational disease could be considered endocrine-disrupting compounds. Endocrine disruptors act to interfere with normal hormone signaling pathways and can have effects at very low doses due to nonmonotonic dose responses [82, 83]. However, it was found that the epigenetic transgenerational effect of vinclozolin in rats was not due to its antiandrogenic actions [65]. Therefore, toxicants may not need to be endocrine disruptors in order to induce epigenetic transgenerational changes.
It is important to remember that epigenetic mechanisms are a crucial part of normal biology and are some of the primary drivers of cell differentiation to produce all the different cell types in an organism [84, 85]. In addition, the epigenome is a normal mechanism by which organisms respond to changes in the environment by changing gene expression. Why, then, can environmental insults result in transgenerational inheritance of increased disease susceptibility, which is a maladaptive response? One possibility may be explained in terms of an environmental mismatch or the predictive adaptive response hypothesis [86–88]. In this hypothesis, an environmental stressor like famine may epigenetically promote an adaptive (thrifty) phenotype in progeny. If the current environment of those progeny has more-than-adequate nutrients, diseases like diabetes and obesity are promoted. Another possibility is that an environmental insult, such as exposure to a toxicant, may interfere with the normal molecular epigenetic machinery and result in stochastic and/or directed epigenetic changes that could be considered epimutations. If these epimutations occur in germ cells, then that can lead to transgenerational inheritance of a wider range of phenotypes in the progeny. Some of those phenotypes may be poorly adapted and develop disease, including reproductive disease. This would explain an increased disease susceptibility in organisms whose ancestors were exposed to environmental insults. However, the increased phenotypic variation may also result in some individuals who are better adapted to an altered environment, facilitating natural selection [81]. These two possibilities are not exclusive of each other, and future investigations will hopefully clarify further how environmental exposures relate to epigenetic transgenerational inheritance of reproductive disease.
Conclusions
Epigenetic and genetic processes always act in concert to regulate gene expression and physiology of an organism. Epigenetically mediated transgenerational inheritance is a normal physiological process. However, exposure to environmental insults, such as toxicants during critical developmental windows, may result in germ-line inheritance of epimutations and an increased susceptibility to reproductive disease in subsequent generations. In this review of studies in various experimental model species, the reports of epigenetic transgenerational inheritance of reproductive disease and abnormalities are described. These results suggest that serious attention be paid to the possibility that ancestral exposures to environmental insults has led to transgenerational inheritance of increased susceptibility to reproductive problems. This will be an important contributing factor to reproductive disease in most species, including humans. The use of epigenetic biomarkers may allow a diagnostic to be developed to determine the toxicant exposures that occurred in your ancestors, so actions can be taken to help mitigate possible disease onset. Observations suggest that much of the reproductive disease and infertility today may in part be due to ancestral environmental exposures through epigenetic transgenerational inheritance mechanisms.
Acknowledgment
We thank Ms. Heather Johnson for assistance in preparation of the manuscript.
References
- 1. Sharpe RM. Environmental/lifestyle effects on spermatogenesis. Philos Trans R Soc Lond B Biol Sci 2010; 365:1697–1712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Carlsen E, Giwercman A, Keiding N, Skakkebaek NE.. Evidence for decreasing quality of semen during past 50 years. Br Med J 1992; 305:609–613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Merzenich H, Zeeb H, Blettner M.. Decreasing sperm quality: a global problem? BMC Public Health 2010; 10:24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Swan SH, Elkin EP, Fenster L.. The question of declining sperm density revisited: an analysis of 101 studies published 1934–1996. Environ Health Perspect 2000; 108:961–966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Skakkebaek NE. Rajpert-De Meyts E, Jorgensen N, Main KM, Leffers H, Andersson AM, Juul A, Jensen TK, Toppari J. Testicular cancer trends as “whistle blowers” of testicular developmental problems in populations. Int J Androl 2007; 30:198–204. [DOI] [PubMed] [Google Scholar]
- 6. Main KM, Skakkebaek NE, Virtanen HE, Toppari J.. Genital anomalies in boys and the environment. Best Pract Res Clin Endocrinol Metab 2010; 24:279–289. [DOI] [PubMed] [Google Scholar]
- 7. Skakkebaek NE, Jorgensen N, Main KM. Rajpert-De Meyts E, Leffers H, Andersson AM, Juul A, Carlsen E, Mortensen GK, Jensen TK, Toppari J. Is human fecundity declining? Int J Androl 2006; 29:2–11. [DOI] [PubMed] [Google Scholar]
- 8. te Velde E, Burdorf A, Nieschlag E, Eijkemans R, Kremer JA, Roeleveld N, Habbema D.. Is human fecundity declining in Western countries? Hum Reprod 2010; 25:1348–1353. [DOI] [PubMed] [Google Scholar]
- 9. Guillette LJ Jr, Iguchi T.. Ecology. Life in a contaminated world. Science 2012; 337:1614–1615. [DOI] [PubMed] [Google Scholar]
- 10. Skinner MK, Manikkam M, Guerrero-Bosagna C.. Epigenetic transgenerational actions of environmental factors in disease etiology. Trends Endocrinol Metab 2010; 21:214–222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Pimentel D, Cooperstein S, Randell H, Filiberto D, Sorrentino S, Kaye B, Nicklin C, Yagi J, Brian J, O'Hern J, Habas A, Weinstein C.. Ecology of increasing diseases: population growth and environmental degradation. Hum Ecol 2007; 35:653–668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Global Burden of Disease Study C. Global, regional, and national incidence, prevalence, and years lived with disability for 301 acute and chronic diseases and injuries in 188 countries, 1990–2013: a systematic analysis for the Global Burden of Disease Study 2013.Lancet2015; 383:743–800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Devesa SS, Silverman DT, Young JL Jr, Pollack ES, Brown CC, Horm JW, Percy CL, Myers MH, McKay FW, Fraumeni JF Jr. Cancer incidence and mortality trends among whites in the United States, 1947–84. J Natl Cancer Inst 1987; 79:701–770. [PubMed] [Google Scholar]
- 14. Giwercman A, Giwercman YL.. Environmental factors and testicular function. Best Pract Res Clin Endocrinol Metab 2011; 25:391–402. [DOI] [PubMed] [Google Scholar]
- 15. Nordkap L, Joensen UN. Blomberg Jensen M, Jorgensen N. Regional differences and temporal trends in male reproductive health disorders: semen quality may be a sensitive marker of environmental exposures. Mol Cell Endocrinol 2012; 355:221–230. [DOI] [PubMed] [Google Scholar]
- 16. Ravnborg TL, Jensen TK, Andersson AM, Toppari J, Skakkebaek NE, Jorgensen N.. Prenatal and adult exposures to smoking are associated with adverse effects on reproductive hormones, semen quality, final height and body mass index. Hum Reprod 2011; 26:1000–1011. [DOI] [PubMed] [Google Scholar]
- 17. Damgaard IN, Skakkebaek NE, Toppari J, Virtanen HE, Shen H, Schramm KW, Petersen JH, Jensen TK, Main KM.. Persistent pesticides in human breast milk and cryptorchidism. Environ Health Perspect 2006; 114:1133–1138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Andersen HR, Schmidt IM, Grandjean P, Jensen TK, Budtz-Jorgensen E, Kjaerstad MB, Baelum J, Nielsen JB, Skakkebaek NE, Main KM.. Impaired reproductive development in sons of women occupationally exposed to pesticides during pregnancy. Environ Health Perspect 2008; 116:566–572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Swan SH, Main KM, Liu F, Stewart SL, Kruse RL, Calafat AM, Mao CS, Redmon JB, Ternand CL, Sullivan S, Teague JL.. Decrease in anogenital distance among male infants with prenatal phthalate exposure. Environ Health Perspect 2005; 113:1056–1061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Kristensen DM, Hass U, Lesne L, Lottrup G, Jacobsen PR, Desdoits-Lethimonier C, Boberg J, Petersen JH, Toppari J, Jensen TK, Brunak S, Skakkebaek NE et al. Intrauterine exposure to mild analgesics is a risk factor for development of male reproductive disorders in human and rat. Hum Reprod 2011; 26:235–244. [DOI] [PubMed] [Google Scholar]
- 21. Damgaard IN, Jensen TK, Petersen JH, Skakkebaek NE, Toppari J, Main KM.. Cryptorchidism and maternal alcohol consumption during pregnancy. Environ Health Perspect 2007; 115:272–277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Newbold RR. Lessons learned from perinatal exposure to diethylstilbestrol. Toxicol Appl Pharmacol 2004; 199:142–150. [DOI] [PubMed] [Google Scholar]
- 23. Kalfa N, Paris F, Soyer-Gobillard MO, Daures JP, Sultan C.. Prevalence of hypospadias in grandsons of women exposed to diethylstilbestrol during pregnancy: a multigenerational national cohort study. Fertil Steril 2011; 95:2574–2577. [DOI] [PubMed] [Google Scholar]
- 24. Franks S, McCarthy MI, Hardy K.. Development of polycystic ovary syndrome: involvement of genetic and environmental factors. Int J Androl 2006; 29:278–285;discussion 286–290. [DOI] [PubMed] [Google Scholar]
- 25. Upson K, Sathyanarayana S, Scholes D, Holt VL.. Early-life factors and endometriosis risk. Fertil Steril 2015; 104:964–971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Rier SE. The potential role of exposure to environmental toxicants in the pathophysiology of endometriosis. Ann N Y Acad Sci 2002; 955:201–212. [DOI] [PubMed] [Google Scholar]
- 27. Jirtle RL, Skinner MK.. Environmental epigenomics and disease susceptibility. Nat Rev Genet 2007; 8:253–262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. McCarrey JR. The epigenome as a target for heritable environmental disruptions of cellular function. Mol Cell Endocrinol 2012; 354:9–15. [DOI] [PubMed] [Google Scholar]
- 29. Grigorova M, Punab M, Ausmees K, Laan M.. FSHB promoter polymorphism within evolutionary conserved element is associated with serum FSH level in men. Hum Reprod 2008; 23:2160–2166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Voorhuis M, Onland-Moret NC, Janse F. Ploos van Amstel HK, Goverde AJ, Lambalk CB, Laven JS, van der Schouw YT, Broekmans FJ, Fauser BC. The significance of fragile X mental retardation gene 1 CGG repeat sizes in the normal and intermediate range in women with primary ovarian insufficiency. Hum Reprod 2014; 29:1585–1593. [DOI] [PubMed] [Google Scholar]
- 31. Visscher PM, Brown MA, McCarthy MI, Yang J.. Five years of GWAS discovery. Am J Hum Genet 2012; 90:7–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Zhao H, Chen ZJ.. Genetic association studies in female reproduction: from candidate-gene approaches to genome-wide mapping. Mol Hum Reprod 2013; 19:644–654. [DOI] [PubMed] [Google Scholar]
- 33. Skinner MK. Environment, epigenetics and reproduction. Mol Cell Endocrinol 2014; 398:1–3. [DOI] [PubMed] [Google Scholar]
- 34. Crews D, Gillette R, Miller-Crews I, Gore AC, Skinner MK.. Nature, nurture and epigenetics. Mol Cell Endocrinol 2014; 398:42–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Waddington CH. The epigenotype. Endeavour 1942; 1:18–20. [Google Scholar]
- 36. Holliday R, Pugh JE.. DNA modification mechanisms and gene activity during development. Science 1975; 187:226–232. [PubMed] [Google Scholar]
- 37. Singer J, Roberts-Ems J, Riggs AD.. Methylation of mouse liver DNA studied by means of the restriction enzymes msp I and hpa II. Science 1979; 203:1019–1021. [DOI] [PubMed] [Google Scholar]
- 38. Kornfeld JW, Bruning JC.. Regulation of metabolism by long, non-coding RNAs. Front Genet 2014; 5:57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Yaniv M. Chromatin remodeling: from transcription to cancer. Cancer Genet 2014; 207:352–357. [DOI] [PubMed] [Google Scholar]
- 40. Oosterhuis JW, Looijenga LH.. Testicular germ-cell tumours in a broader perspective. Nat Rev Cancer 2005; 5:210–222. [DOI] [PubMed] [Google Scholar]
- 41. Rajender S, Avery K, Agarwal A.. Epigenetics, spermatogenesis and male infertility. Mutat Res 2011; 727:62–71. [DOI] [PubMed] [Google Scholar]
- 42. Hammoud SS, Nix DA, Hammoud AO, Gibson M, Cairns BR, Carrell DT.. Genome-wide analysis identifies changes in histone retention and epigenetic modifications at developmental and imprinted gene loci in the sperm of infertile men. Hum Reprod 2011; 26:2558–2569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Hammoud SS, Purwar J, Pflueger C, Cairns BR, Carrell DT.. Alterations in sperm DNA methylation patterns at imprinted loci in two classes of infertility. Fertil Steril 2010; 94:1728–1733. [DOI] [PubMed] [Google Scholar]
- 44. Guerrero-Bosagna C, Savenkova M, Haque MM, Sadler-Riggleman I, Skinner MK.. Environmentally induced epigenetic transgenerational inheritance of altered Sertoli cell transcriptome and epigenome: molecular etiology of male infertility. PLoS ONE 2013; 8:e59922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Nolan KD, Franco OE, Hance MW, Hayward SW, Isaacs JS.. Tumor-secreted Hsp90 subverts polycomb function to drive prostate tumor growth and invasion. J Biol Chem 2015; 290:8271–8282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Koike N, Higashiura Y, Akasaka J, Uekuri C, Ito F, Kobayashi H.. Epigenetic dysregulation of endometriosis susceptibility genes (review). Mol Med Rep 2015; 12:1611–1616. [DOI] [PubMed] [Google Scholar]
- 47. Izawa M, Taniguchi F, Terakawa N, Harada T.. Epigenetic aberration of gene expression in endometriosis. Front Biosci (Elite Ed) 2013; 5:900–910. [DOI] [PubMed] [Google Scholar]
- 48. Kobayashi H, Imanaka S, Nakamura H, Tsuji A.. Understanding the role of epigenomic, genomic and genetic alterations in the development of endometriosis (review). Mol Med Rep 2014; 9:1483–1505. [DOI] [PubMed] [Google Scholar]
- 49. Pistek VL, Furst RW, Kliem H, Bauersachs S, Meyer HH, Ulbrich SE.. HOXA10 mRNA expression and promoter DNA methylation in female pig offspring after in utero estradiol-17beta exposure. J Steroid Biochem Mol Biol 2013; 138:435–444. [DOI] [PubMed] [Google Scholar]
- 50. Bromer JG, Wu J, Zhou Y, Taylor HS.. Hypermethylation of homeobox A10 by in utero diethylstilbestrol exposure: an epigenetic mechanism for altered developmental programming. Endocrinology 2009; 150:3376–3382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Ge ZJ, Schatten H, Zhang CL, Sun QY.. Oocyte ageing and epigenetics. Reproduction 2015; 149:R103–R114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Xu J, Sinclair KD.. One-carbon metabolism and epigenetic regulation of embryo development. Reprod Fertil Dev 10.1071/RD14377. [DOI] [PubMed] [Google Scholar]
- 53. Nilsson E, Larsen G, Manikkam M, Guerrero-Bosagna C, Savenkova M, Skinner M.. Environmentally induced epigenetic transgenerational inheritance of ovarian disease. PLoS ONE 2012; 7:e36129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Skinner MK. Environmental epigenetic transgenerational inheritance and somatic epigenetic mitotic stability. Epigenetics 2011; 6:838–842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Anway MD, Cupp AS, Uzumcu M, Skinner MK.. Epigenetic transgenerational actions of endocrine disruptors and male fertility. Science 2005; 308:1466–1469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Wong C, Kelce WR, Sar M, Wilson EM.. Androgen receptor antagonist versus agonist activities of the fungicide vinclozolin relative to hydroxyflutamide. J Biol Chem 1995; 270:19998–20003. [DOI] [PubMed] [Google Scholar]
- 57. Anway MD, Leathers C, Skinner MK.. Endocrine disruptor vinclozolin induced epigenetic transgenerational adult-onset disease. Endocrinology 2006; 147:5515–5523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Anway MD, Memon MA, Uzumcu M, Skinner MK.. Transgenerational effect of the endocrine disruptor vinclozolin on male spermatogenesis. J Androl 2006; 27:868–879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Guerrero-Bosagna C, Covert T, Haque MM, Settles M, Nilsson EE, Anway MD, Skinner MK.. Epigenetic transgenerational inheritance of vinclozolin induced mouse adult onset disease and associated sperm epigenome biomarkers. Reprod Toxicol 2012; 34:694–707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Guerrero-Bosagna C, Settles M, Lucker B, Skinner M.. Epigenetic transgenerational actions of vinclozolin on promoter regions of the sperm epigenome. Plos One 2010; 5:e13100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Skinner MK. What is an epigenetic transgenerational phenotype? F3 or F2. Reprod Toxicol 2008; 25:2–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Feng S, Jacobsen SE, Reik W.. Epigenetic reprogramming in plant and animal development. Science 2010; 330:622–627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Messerschmidt DM, Knowles BB, Solter D.. DNA methylation dynamics during epigenetic reprogramming in the germline and preimplantation embryos. Genes Dev 2014; 28:812–828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Skinner MK, Manikkam M, Tracey R, Nilsson E, Haque MM, Guerrero-Bosagna C.. Ancestral dichlorodiphenyltrichloroethane (DDT) exposure promotes epigenetic transgenerational inheritance of obesity. BMC Med 2013; 11:228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Anway MD, Rekow SS, Skinner MK.. Comparative anti-androgenic actions of vinclozolin and flutamide on transgenerational adult onset disease and spermatogenesis. Reprod Toxicol 2008; 26:100–106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Doyle TJ, Bowman JL, Windell VL, McLean DJ, Kim KH.. Transgenerational effects of di-(2-ethylhexyl) phthalate on testicular germ cell associations and spermatogonial stem cells in mice. Biol Reprod 2013; 88:112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Manikkam M, Guerrero-Bosagna C, Tracey R, Haque MM, Skinner MK.. Transgenerational actions of environmental compounds on reproductive disease and identification of epigenetic biomarkers of ancestral exposures. PLoS ONE 2012; 7:e31901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Manikkam M, Tracey R, Guerrero-Bosagna C, Skinner M.. Plastics derived endocrine disruptors (BPA, DEHP and DBP) induce epigenetic transgenerational inheritance of adult-onset disease and sperm epimutations. PLoS ONE 2013; 8:e55387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Manikkam M, Tracey R, Guerrero-Bosagna C, Skinner M.. Pesticide and insect repellent mixture (permethrin and DEET) induces epigenetic transgenerational inheritance of disease and sperm epimutations. Reprod Toxicol 2012; 34:708–719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Mohamed el SA, Song WH, Oh SA, Park YJ, You YA, Lee S, Choi JY, Kim YJ, Jo I, Pang MG. The transgenerational impact of benzo(a)pyrene on murine male fertility. Hum Reprod 2010; 25:2427–2433. [DOI] [PubMed] [Google Scholar]
- 71. Anway MD, Skinner MK.. Transgenerational effects of the endocrine disruptor vinclozolin on the prostate transcriptome and adult onset disease. Prostate 2008; 68:517–529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Manikkam M, Tracey R, Guerrero-Bosagna C, Skinner MK.. Dioxin (TCDD) induces epigenetic transgenerational inheritance of adult onset disease and sperm epimutations. PLoS ONE 2012; 7:e46249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Qin Y, Jiao X, Simpson JL, Chen ZJ.. Genetics of primary ovarian insufficiency: new developments and opportunities. Hum Reprod Update 2015; 21:787–808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Tracey R, Manikkam M, Guerrero-Bosagna C, Skinner M.. Hydrocarbons (jet fuel JP-8) induce epigenetic transgenerational inheritance of obesity, reproductive disease and sperm epimutations. Reprod Toxicol 2013; 36:104–116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Manikkam M, Haque MM, Guerrero-Bosagna C, Nilsson E, Skinner M.. Pesticide methoxychlor promotes the epigenetic transgenerational inheritance of adult onset disease through the female germline. PLoS ONE 2014; 9:e102091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Barthelmess EK, Naz RK.. Polycystic ovary syndrome: current status and future perspective. Front Biosci (Elite Ed) 2014; 6:104–119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Bruner-Tran KL, Osteen KG.. Developmental exposure to TCDD reduces fertility and negatively affects pregnancy outcomes across multiple generations. Reprod Toxicol 2011; 31:344–350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Baker TR, King-Heiden TC, Peterson RE, Heideman W.. Dioxin induction of transgenerational inheritance of disease in zebrafish. Mol Cell Endocrinol 2014; 398:36–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Salian S, Doshi T, Vanage G.. Perinatal exposure of rats to Bisphenol A affects fertility of male offspring—an overview. Reprod Toxicol 2011; 31:359–362. [DOI] [PubMed] [Google Scholar]
- 80. Crews D, Gore AC, Hsu TS, Dangleben NL, Spinetta M, Schallert T, Anway MD, Skinner MK.. Transgenerational epigenetic imprints on mate preference. Proc Natl Acad Sci U S A 2007; 104:5942–5946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Skinner MK. Environmental epigenetics and a unified theory of the molecular aspects of evolution: a neo-Lamarckian concept that facilitates neo-Darwinian evolution. Genome Biol Evol 2015; 7:1296–1302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Beausoleil C, Ormsby JN, Gies A, Hass U, Heindel JJ, Holmer ML, Nielsen PJ, Munn S, Schoenfelder G.. Low dose effects and non-monotonic dose responses for endocrine active chemicals: science to practice workshop: workshop summary. Chemosphere 2013; 93:847–856. [DOI] [PubMed] [Google Scholar]
- 83. Lagarde F, Beausoleil C, Belcher SM, Belzunces LP, Emond C, Guerbet M, Rousselle C.. Non-monotonic dose-response relationships and endocrine disruptors: a qualitative method of assessment. Environ Health 2015; 14:13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Cantone I, Fisher AG.. Epigenetic programming and reprogramming during development. Nat Struct Mol Biol 2013; 20:282–289. [DOI] [PubMed] [Google Scholar]
- 85. Ladewig J, Koch P, Brustle O.. Leveling Waddington: the emergence of direct programming and the loss of cell fate hierarchies. Nat Rev Mol Cell Biol 2013; 14:225–236. [DOI] [PubMed] [Google Scholar]
- 86. Bateson P, Gluckman P, Hanson M.. The biology of developmental plasticity and the predictive adaptive response hypothesis. J Physiol 2014; 592:2357–2368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Boekelheide K, Blumberg B, Chapin RE, Cote I, Graziano JH, Janesick A, Lane R, Lillycrop K, Myatt L, States JC, Thayer KA, Waalkes MP et al. Predicting later-life outcomes of early-life exposures. Environ Health Perspect 2012; 120:1353–1361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Raubenheimer D, Simpson SJ, Tait AH.. Match and mismatch: conservation physiology, nutritional ecology and the timescales of biological adaptation. Philos Trans R Soc Lond B Biol Sci 2012; 367:1628–1646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Salian S, Doshi T, Vanage G.. Impairment in protein expression profile of testicular steroid receptor coregulators in male rat offspring perinatally exposed to bisphenol A. Life Sci 2009; 85:11–18. [DOI] [PubMed] [Google Scholar]
- 90. Wolstenholme JT, Edwards M, Shetty SR, Gatewood JD, Taylor JA, Rissman EF, Connelly JJ.. Gestational exposure to bisphenol A produces transgenerational changes in behaviors and gene expression. Endocrinology 2012; 153:3828–3838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Chamorro-Garcia R, Sahu M, Abbey RJ, Laude J, Pham N, Blumberg B.. Transgenerational inheritance of increased fat depot size, stem cell reprogramming, and hepatic steatosis elicited by prenatal exposure to the obesogen tributyltin in mice. Environ Health Perspect 2013; 121:359–366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Padmanabhan N, Watson ED.. Lessons from the one-carbon metabolism: passing it along to the next generation. Reprod Biomed Online 2013; 27:637–643. [DOI] [PubMed] [Google Scholar]
- 93. Dunn GA, Morgan CP, Bale TL.. Sex-specificity in transgenerational epigenetic programming. Horm Behav 2011; 59:290–295. [DOI] [PubMed] [Google Scholar]
- 94. Burdge GC, Hoile SP, Uller T, Thomas NA, Gluckman PD, Hanson MA, Lillycrop KA.. Progressive, transgenerational changes in offspring phenotype and epigenotype following nutritional transition. PLoS ONE 2011; 6:e28282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95. Pembrey ME, Bygren LO, Kaati G, Edvinsson S, Northstone K, Sjostrom M, Golding J.. Sex-specific, male-line transgenerational responses in humans. Eur J Hum Genet 2006; 14:159–166. [DOI] [PubMed] [Google Scholar]
- 96. Veenendaal MV, Painter RC, de Rooij SR, Bossuyt PM, van der Post JA, Gluckman PD, Hanson MA, Roseboom TJ.. Transgenerational effects of prenatal exposure to the 1944–45 Dutch famine. BJOG 2013; 120:548–553. [DOI] [PubMed] [Google Scholar]
- 97. Burdge GC, Slater-Jefferies J, Torrens C, Phillips ES, Hanson MA, Lillycrop KA.. Dietary protein restriction of pregnant rats in the F0 generation induces altered methylation of hepatic gene promoters in the adult male offspring in the F1 and F2 generations. Br J Nutr 2007; 97:435–439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Painter RC, Osmond C, Gluckman P, Hanson M, Phillips DI, Roseboom TJ.. Transgenerational effects of prenatal exposure to the Dutch famine on neonatal adiposity and health in later life. BJOG 2008; 115:1243–1249. [DOI] [PubMed] [Google Scholar]
- 99. Song J, Irwin J, Dean C.. Remembering the prolonged cold of winter. Curr Biol 2013; 23:R807–R811. [DOI] [PubMed] [Google Scholar]
- 100. Norouzitallab P, Baruah K, Vandegehuchte M, Van Stappen G, Catania F, Vanden Bussche J, Vanhaecke L, Sorgeloos P, Bossier P.. Environmental heat stress induces epigenetic transgenerational inheritance of robustness in parthenogenetic Artemia model. FASEB J 2014; 28:3552–3563. [DOI] [PubMed] [Google Scholar]
- 101. Zheng X, Chen L, Li M, Lou Q, Xia H, Wang P, Li T, Liu H, Luo L.. Transgenerational variations in DNA methylation induced by drought stress in two rice varieties with distinguished difference to drought resistance. PLoS ONE 2013; 8:e80253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102. Suter L, Widmer A.. Environmental heat and salt stress induce transgenerational phenotypic changes in Arabidopsis thaliana. PLoS ONE 2013; 8:e60364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103. Dias BG, Ressler KJ.. Parental olfactory experience influences behavior and neural structure in subsequent generations. Nat Neurosci 2014; 17:89–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104. Gapp K, Jawaid A, Sarkies P, Bohacek J, Pelczar P, Prados J, Farinelli L, Miska E, Mansuy IM.. Implication of sperm RNAs in transgenerational inheritance of the effects of early trauma in mice. Nat Neurosci 2014; 17:667–669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105. Rehan VK, Liu J, Sakurai R, Torday JS.. Perinatal nicotine-induced transgenerational asthma. Am J Physiol Lung Cell Mol Physiol 2013; 305:L501–L507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106. Golding J, Northstone K, Gregory S, Miller LL, Pembrey M.. The anthropometry of children and adolescents may be influenced by the prenatal smoking habits of their grandmothers: a longitudinal cohort study. Am J Hum Biol 2014; 26:731–739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. Govorko D, Bekdash RA, Zhang C, Sarkar DK.. Male germline transmits fetal alcohol adverse effect on hypothalamic proopiomelanocortin gene across generations. Biol Psychiatry 2012; 72:378–388. [DOI] [PMC free article] [PubMed] [Google Scholar]