

SUMMARY
Most cellular stresses induce protein translation inhibition and stress granule formation. Here, using Drosophila S2 cells, we investigate the role of G3BP/Rasputin in this process. In contrast to arsenite treatment, where dephosphorylated Ser142 Rasputin is recruited to stress granules, we find that, upon amino acid starvation, only the phosphorylated Ser142 form is recruited. Furthermore, we identify Sec16, a component of the endoplasmic reticulum exit site, as a Rasputin interactor and stabilizer. Sec16 depletion results in Rasputin degradation and inhibition of stress granule formation. However, in the absence of Sec16, pharmacological stabilization of Rasputin is not enough to rescue the assembly of stress granules. This is because Sec16 specifically interacts with phosphorylated Ser142 Rasputin, the form required for stress granule formation upon amino acid starvation. Taken together, these results demonstrate that stress granule formation is fine-tuned by specific signaling cues that are unique to each stress. These results also expand the role of Sec16 as a stress response protein.
Graphical abstract

In Brief
Aguilera-Gomez et al. show that, although stress granules appear similar, the mode of their formation depends on the cellular stress. Contrary to arsenite treatment, the RNA-binding protein Rasputin needs to be phosphorylated and bound to the ERES scaffold Sec16 to lead to the formation of stress granules upon amino acid starvation.
INTRODUCTION
Stress granules are well-studied, cytoplasmic reversible, pro-survival stress assemblies where untranslated free RNAs (resulting from protein translation inhibition) are stored and protected together with RNA-binding proteins, translation initiation factors, and the 40S ribosomal subunits (Anderson and Kedersha, 2008; Protter and Parker, 2016). Stress granule formation has been best investigated in mammalian cells upon different type of stresses, including heat and oxidative stress (Anderson and Kedersha, 2002). This has led to the identification of a number of factors that are essential for their formation, such as the case for Tia-1 (Gilks et al., 2004; Kedersha et al., 1999) and Ras-GAP SH3 domain-binding protein (G3BP1/2, referred to as G3BP hereafter) (Tourriére et al., 2003).
G3BP was first identified in human cells through co-immunoprecipitation with the SH3 domain of RasGAP. However, it has an RNA recognition motif (RRM) toward the C terminus, suggesting that it binds mRNAs. In growing cells, G3BP is normally cytoplasmic; However, after stress induction (especially stress leading to eIF2α phosphorylation; McEwen et al., 2005), it is not only readily recruited to stress granules but also is necessary for their formation (Kedersha et al., 2016; Tourrie`re et al., 2003; White et al., 2007). Furthermore, G3BP drives stress granule formation when overexpressed in the absence of stress (Tourriére et al., 2003). Importantly, G3BP is phosphorylated on Ser149 during basal conditions, but it needs to be dephosphorylated to drive stress granule assembly triggered by arsenite treatment (Kedersha et al., 2016; Tourrie` re et al., 2003). Taken together, G3BP is critical for stress granule formation when its Ser149 is dephosphorylated and through its binding to Caprin and the 40S ribosomal subunit. Conversely, G3BP is inhibited when binding to peptidase USP10 (Kedersha et al., 2016).
G3BP also appears to have an important role in disease. First, viruses can exploit G3BP (reviewed in Tsai and Lloyd, 2014). However, G3BP also appears to slow down HIV replication by leading to the sequestration of viral mRNA (Cobos Jiménez et al., 2015). Second, G3BP is overexpressed in gastric cancer (Min et al., 2015) and bone and lung sarcomas (Somasekharan et al., 2015), where it is considered as a marker for poor survival. Strikingly, downregulation of G3BP in cells and in vivo reduces stress granule formation as expected but also tumor invasion and metastasis, showing a clear role for stress granule formation in cancer. Last, G3BP is a target of TDP-43 that is often mutated, mislocalized, and misaccumulated in amyotrophic lateral sclerosis (ALS) (Aulas et al., 2012).
Stress granules are also formed in Drosophila, for instance, upon heat stress, arsenite exposure (Farny et al., 2009), and amino acid starvation of Drosophila S2 cells (Zacharogianni et al., 2014). However, the mechanism behind their formation upon this latter stress is not completely understood. Interestingly, amino acid starvation leads to the formation of another recently described stress assembly, the Sec bodies that store and protect most of the COPII subunits and the endoplasmic reticulum exit site (ERES) component Sec16 (Zacharogianni et al., 2014). Importantly, Sec bodies and stress granules are independent structures that are formed at the same time frame of amino acid starvation.
Sec16 is a conserved peripheral membrane protein that tightly localizes and concentrates to the ERES (Connerly et al., 2005; Watson et al., 2006) via a domain that has been mapped to a small arginine-rich region upstream of the conserved central domain (Hughes et al., 2009; Ivan et al., 2008). It binds nearly all COPII subunits and controls at least two aspects of COPII-coated vesicle dynamics (Sprangers and Rabouille, 2015). Sec16 is essential for endoplasmic reticulum (ER) to Golgi transport, especially in Drosophila where the absence of Sec16 results in a severe inhibition of protein exit from the ER (Ivan et al., 2008).
Interestingly, Sec16 responds to nutrient stress (Zacharo-gianni et al., 2011, 2014). In this regard, we have recently shown that Sec16 is a key factor driving Sec body formation upon amino acid starvation that activates the ER-localized dPARP16. In turn, dPARP16 mono-ADP-ribosylates Sec16 on a conserved sequence close to its C terminus (Aguilera-Gomez et al., 2016), and Sec16 modification by dPARP16 is enough to elicit Sec body formation. This demonstrates that Sec16 is a stress response protein that plays an important role in the response to amino acid starvation.
Here we show that the phosphorylation state of the G3BP Drosophila ortholog Rasputin (Rin) is differentially required for the formation of stress granules that are formed upon arsenite treatment and amino acid starvation. Whereas stress granule formation upon arsenite treatment requires the non-phosphorylated form of Rin (as is the case for G3BP in mammalian cells), amino acid starvation requires the phosphorylated form. Furthermore, we show that Sec16 specifically interacts with phosphorylated Rin and mediates this differential requirement.
All together, these results provide a link, which to our knowledge has not been reported, between the protein transport from the ER and protein translation. It also explains the specific requirement of Sec16 for stress granule formation upon amino acid starvation, but not other stresses. Furthermore, it enlarges the scope of Sec16 function at the ER by identifying yet a new role in the response to amino acid starvation.
RESULTS
Amino Acid Starvation Induces the Formation of Bona Fide Stress Granules
Amino acid starvation (Incubation in KRB, see the Experimental Procedures and Figures S1A and S1B) induced the inhibition of protein translation (Figure 1A), the phosphorylation of eIF2α (Figure 1B) as strongly as sodium arsenite treatment, and the subsequent formation of stress granules. These were marked by three RNA-binding proteins, FMR1, Caprin, and Rin (Figures 1C and 1D) as well as Tral (Yang et al., 2006) and the initiation factor elF4E (Figure 1E). Stress granules began to form after 1 hr of starvation, with their number reaching a maximum after 3–4 hr (Figure 1F). They were clearly different from autophagosomes that also formed upon starvation (Figure S1C). Importantly, stress granule formation depended on the presence of ribosome-free mRNAs, as cycloheximide treatment completely blocked their formation in starved cells (Figures 1F and 1F’) and cycloheximide washout allowed their formation (data not shown). Conversely, puromycin led to the robust formation of stress granules (Figures 1G and 1G’). Last, they were completely reversible upon 60 min of stress relief (addition of full medium) (Figures 1H and 1H’). This suggests that the stress granules that were formed by amino acid starvation were canonical, as they had the same properties as those formed upon other stresses (Aulas et al., 2017; Farny et al., 2009).
Figure 1. Amino Acid Starvation Leads to the Formation of Canonical Stress Granules.
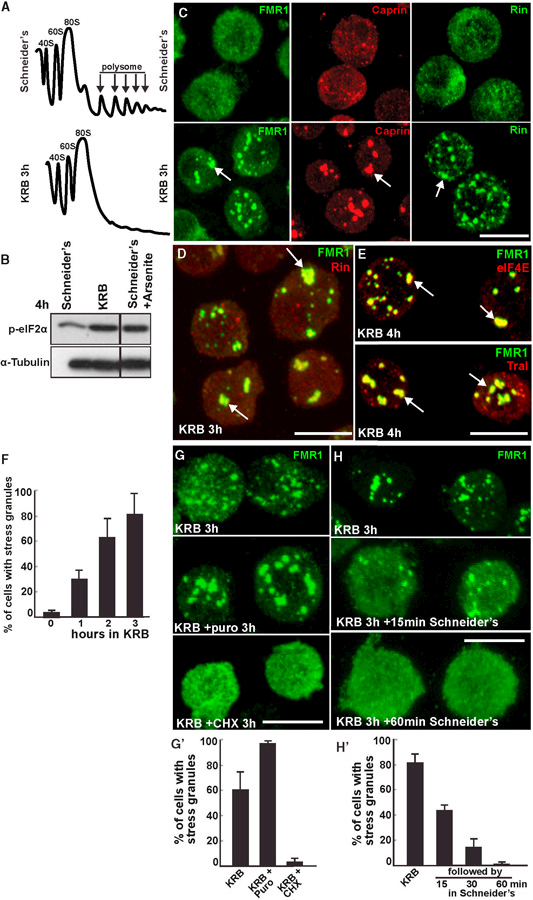
(A) Polysome profiles of Drosophila S2 cells grown in Schneider’s and incubated 4 hr in KRB.
(B) Western blot visualization of elF2a phosphorylation in growing, KRB- and arsenite-treated cells.
(C) Immunofluorescence (IF) visualization of endogenous FMR1, Caprin, and Rin in cells incubated in Schneider’s and KRB for 3 hr. Note that stress granules form (arrows).
(D) IF co-localization of FMR1 and Rin. Note that they perfectly co-localize in stress granules (arrows).
(E) IF co-localization of FMR1 and eIF4E and FMR1 and Tral in stress granules (arrows).
(F) Kinetics of stress granule formation (marked by FMR1) in S2 cells incubated in KRB over the indicated time.
(G) IF visualization of FMR1 upon KRB incubation, KRB supplemented with puromycin (puro) and cycloheximide (CHX), quantified in (G’). Note that stress granules do not form upon CHX incubation.
(H) IF visualization of FMR1 upon KRB incubation followed by 15 and 60 min reversion in Schneider’s, quantified in (H’). Note that stress granules dissolve rapidly upon stress relief.
Scale bars, 10 µm. Error bar, SEM.
PhosphoRin Is Required for Stress Granule Formation upon Amino Acid Starvation
As mentioned in the Introduction, Rasputin (Rin) is the Drosophila ortholog of G3BP and the molecular organization is largely conserved (Figure S2A). So are the amino acid sequences of the NFT2 and RRM domains (Figure S2C). Importantly, Ser149 of G3BP is conserved in Rin (Ser142) (Figure S2B). Furthermore, both proteins harbor a proline-rich intrinsically disordered domain and a glycine rich-domain at the C terminus (Kedersha et al., 2016). Interestingly, as G3BP, Rin overexpression in mammalian cells leads to the formation of stress granules in the absence of stress (Tourrie`re et al., 2003).
To test the role of Rin in stress granule formation in amino acid starved Drosophila S2 cells, we first depleted it. In the absence of Rin (Figure 2A’’), stress granules (marked by FMR1) did not form (Figures 2A and 2A’). We then investigated whether Rin overexpression drives stress granule formation in Drosophila S2 cells in the absence of stress. However, this was not the case (Figures 2B and 2B’), even after prolonged expression when the total level of Rin clearly increased (Figure 2B’’). Taken together, this suggests that Rin is necessary, but not sufficient, to drive the assembly of Drosophila stress granules.
Figure 2. Rin S142E, not S142A, Is Incorporated in Stress Granules upon Amino Acid Starvation.
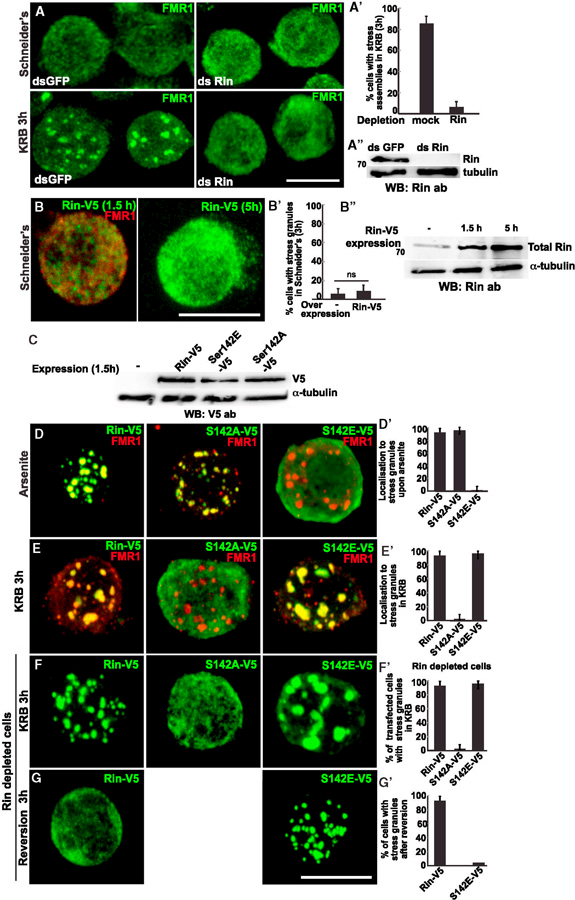
(A–A”) IF visualization of FMR1 in mock- (dsGFP) and Rin- (dsRin) depleted S2 cells (A and A” ) in growing conditions (Schneider’s) and upon amino acid starvation (KRB), quantified in (A’ ). Note that in Rin-depleted cells stress granules do not form.
(B) Note that Rin-V5 overexpression (1.5 and 5 hr) does not induce stress granule formation even though the level of total Rin measured by western blot (using the anti-Rin antibody) increases (B”), quantified in (B’).
(C) Western blot (using anti-V5 antibody) assessing the expression level of the V5-tagged Rin constructs used in (D)–(G).
(D, D’) IF visualization of FRM1 (red) and overexpressed Rin-V5, S142A-V5, and S142E-V5 upon arsenite treatment for 3 hr (D). Note that S142A-V5 is steadily recruited into stress granule, while S142E is not. Quantified in D’.
(E, E”) IF visualization of FRM1 (red) an-d overexpressed Rin-V5, S142A-V5, and S142E-V5 upon incubation in KRB for 3 hr (E). Note that S142E-V5 is steadily recruited into stress granule, while S142A is not. Quantified in E’.
(F–F’) IF visualization of Rin-V5, S142A-V5, and S142E-V5 in cells depleted of endogenous Rin incubated in KRB for 3 hr (F). Note that Rin-V5 and S142E-V5 expression rescues stress granule formation, whereas S142A expression does not. Quantified in F’.
(G–G’) IF visualization of Rin-V5 and S142E-V5 in Rin depleted cells after incubation in KRB for 3 hr followed by 3 hr incubation in Schneider’s (reversion) (G and quantified in G’ ). Note that Rin-V5 positive stress granules are completely reverted, whereas S142E positive stress granules are not.
Scale bars, 10 µm. Error bar, SEM.
As hinted at in the Introduction, S149A, but not S149E, G3BP rescues arsenite-triggered stress granule formation in G3BPdepleted mammalian cells (Tourrie`re et al., 2003; Kedersha et al., 2016). This shows that, upon arsenite treatment, the stress granule formation-competent form of G3BP is the dephosphorylated form. To test how Rin behaves in S2 cells, we generated phospho-mimetic S142E-V5 and non-phosphorylatable S142A-V5 Rin mutants, overexpressed them in wild-type S2 cells (Figure 2C), and tested their incorporation into stress granules upon arsenite treatment. Only S142A-V5 Rin was incorporated into stress granules, whereas S142E-V5 was completed excluded and remained cytoplasmic (Figures 2D and 2D’). In striking contrast, upon amino acid starvation, S142A-V5 remained cytosolic, whereas S142E-V5 Rin was readily incorporated into stress granules as efficiently as the wild-type Rin (Figures 2E and 2E’). We confirmed this result by expressing the Rin mutants in Rin-depleted cells. Both Rin-V5 and S142E-V5 expression rescued the formation of stress granules in amino acid-starved cells, whereas S142A did not (Figures 2F and 2F’). This shows that, contrary to arsenite treatment, phosphorylated Rin is the competent form for stress granule formation upon amino acid starvation.
Interestingly, S142E-V5 overexpression (Table S1, line 7) led to a 1.7-fold increase in the size of stress granules when compared to Rin-V5 overexpression that resulted in the formation of smaller but more numerous stress granules (Table S1, line 5). This suggests that the dynamics of stress granules might be affected by the inability of Rin to be dephosphorylated. To test this, we assessed the reversibility of the Rin-V5- and S142E-V5-positive stress granules. We found that Rin-V5-positive stress granules are fully reversible, whereas the ones formed by S142E-V5 are not, even after 3 hr of full medium incubation (Figures 2G and 2G’; Table S1, lines 6 and 8). This shows that, upon amino acid starvation, stress granule formation requires the phosphorylated form of Rin, whereas stress granule reversion requires its dephosphorylation.
Taken together, these results show that the formation of stress granules upon both arsenite and amino acid starvation is modulated by different but specific signaling cues. This prompted us to look for factors that are required for stress granule formation specifically upon amino acid starvation, but not upon other stresses.
Sec16 Interacts with Rin
In addition to stress granule formation, amino acid starvation also triggers Sec body formation (Zacharogianni et al., 2014). Importantly, Sec bodies are exclusively formed upon amino acid starvation, not arsenite treatment. Furthermore, although stress granules and Sec bodies are distinct structures, they tend to form in close proximity to each other (Figures 3A–3A’’). In this respect, using immunoelectron microscopy, we found a small pool of Sec16 localized inside stress granules (asterisk in Figures 3B and 3B’, red circles), although, as reported before, Sec16 bulk is found in Sec bodies (Figure 3B, arrow) (Zacharogianni et al., 2014). This led us to investigate whether Sec16 plays a role in stress granule formation upon amino acid starvation.
Figure 3. Sec16 Interacts with Rin.

(A) IF visualization of FMR1 (green) and Sec16 (red) in cells in Schneider’s (A) and in KRB (A’). Note that Sec bodies (marked by Sec16) and stress granules form in close proximity to each other (A’’).
(B) Immuno-EM localization of Sec16 (10 nm gold) and Tral (15 nm gold) in ultrathin sections of S2 cells incubated in KRB for 4 hr (B and B’). Note that Sec16 is mostly localized to Sec bodies (arrow) but also populates (red circles) stress granules (asterisks, marked by Tral). Note the mitochondria surrounding stress granules.
(C) Pie chart representation of the 12 gene ontology groups representing the 149 interactors of endogenous Sec16 (see the Experimental Procedures and Table S2).
(D) Western blot visualization of Rin following immunoprecipitation of endogenous Sec16 from growing (Schneider’s) and amino acid-starved (KRB 3 hr) S2 cells.
(E–G and J) Confocal sections co-visualizing full-length Sec16-GFP-CAAX (E), DCter Sec16-GFP-CAAX (F), and Cter-GFP-CAAX (G) and endogenous Rin (red) upon amino acid starvation (KRB), quantified in (J).
(H, I, and K) Confocal sections co-visualizing Sec16-GFP-CAAX with Rin-V5 (red, H) and with DRRM Rin-V5 (red, I) upon amino acid starvation (KRB), quantified in (K). Note that Rin-V5 is efficiently recruited to the plasma membrane by Sec16-CAAX, whereas DRRM-V5 is not and also does not form stress granules.
Scale bars, 10 µm. Error bar, SD.
To begin to investigate this, we performed a mass spectrometry analysis of proteins co-immunoprecipitated with endogenous Sec16 from cells in growing conditions and upon amino acid starvation. 149 candidate interactors passing the selection criteria (see the Experimental Procedures; Table S2) were pulled down, 62% specifically from starved cells, 17% only from cells in growing conditions, and 21% from both conditions, such as Sec13 (Table S2). All candidates were grouped by 12 gene ontology (GO) terms, including one (RNA-binding proteins) that is functional for stress granule formation to which Rin belongs (Table S2; Figure 3C).
We confirmed that Sec16 binds to Rin using several approaches. First, Sec16 immunoprecipitation followed by western blot clearly showed that Sec16 bound Rin (Figure 3D) in a specific manner, as Caprin and FMR1 were not co-immunoprecipitated (Figure S3A). Second, their interaction was also suggested in physiological conditions where Rin was observed in very close proximity to Sec16, especially at the earlier time point of amino acid starvation (Figure S3B, S3B’). Third, we used an anchoraway strategy, whereby Sec16 tagged with the Ras CAAX motif resulted in its anchoring to the plasma membrane (Aguilera-Go-mez et al., 2016). Using full-length Sec16-CAAX, we demonstrated that endogenous Rin is very efficiently recruited to the plasma membrane (Figures 3E and 3J), confirming their interaction.
The C terminus part of Sec16 has been implicated in the stress response (Zacharogianni et al., 2011, 2014), and we tested whether it is also involved in Sec16/Rin interaction. To do this, we expressed Sec16-GFP-CAAX lacking its C terminus, but Rin was still recruited to the plasma membrane (Figures 3F and 3J). Accordingly, expression of the Sec16 Cter-CAAX did not recruit Rin (Figures 3G and3J), and Rin was able to form stress granules in the cytoplasm. The N terminus of Sec16 was also not required for Rin binding (Figure S3C; Figure 3J).
We then examined whether the RNA-binding domain (RRM) of Rin is required for Sec16 interaction, by generating a ΔRRM Rin mutant (Figure S2C) and expressing it together with Sec16-GFP-CAAX. This mutant was not recruited to the plasma membrane, suggesting that this domain is important for Sec16/Rin interaction (Figures 3H, 3I, and 3K). Importantly; ΔRRM Rin did not form stress granules. Furthermore, it had a dominant-negative effect as when expressed in wild-type cells; it prevented the formation of stress granules (Figure S3D). Taken together, these results show that Sec16 is a new Rin interactor.
Sec16, but Not Active Secretion, Is Specifically Required for Stress Granule Formation upon Amino Acid Starvation
We then examined whether Sec16 plays a role in stress granule formation specifically induced by amino acid starvation. We starved Sec16-depleted cells, and we found that stress granules are formed 75% less than in mock-depleted cells (Figure 4E), as monitored using Rin (Figure 4A), elF4E and Tral (Figure 4B), FMR1 (Figure 4C), and Caprin (data not shown). This indicates that Sec16 is a factor required in stress granule formation.
Figure 4. Sec16 Is Required for Stress Granule Formation Specifically upon Amino Acid Starvation.
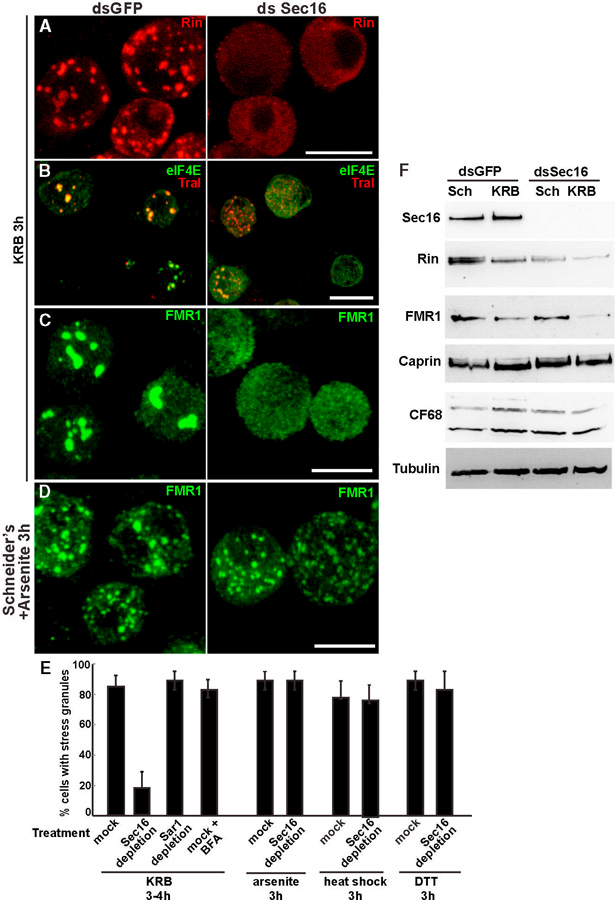
(A–C) IF visualization of endogenous Rin (A), Tral and eIF4E (B), and FMR1 (C) in mock- (dsGFP) and Sec16- (dsSec16) depleted cells upon 3 hr of incubation in KRB. Note that, in the absence of Sec16, stress granules do not form and Rin fluorescence is weak when compared to mock depleted cells.
(D) IF visualization of endogenous FMR1 in mock- and Sec16-depleted cells upon arsenite treatment (3 hr). Note that stress granules form as efficiently in mock- and Sec16-depleted cells.
(E) Quantification of Sec16-depleted cells treated with KRB, arsenite, heat shock, and DTT, expressed in percentage of cell with stress granules.
(F) Western blot of Sec16, Rin, FMR1, Caprin, CF68, and tubulin in mock- and Sec16-depleted cells.
Scale bars, 10 µm. Error bars, SEM.
To investigate whether the Sec16 requirement is specific for stress granule formation upon amino acid starvation, we treated Sec16-depleted cells with arsenite (Figures 4D and 4E), heat stress, and ER stress (DTT) (Figure S4A; Figure 4E), and we monitored stress granule formation that was found to be as efficient as in mock-depleted cells. This shows that Sec16 is required for stress granule formation specifically upon amino acid starvation.
Last, we addressed whether the role of Sec16 in stress granule formation is linked to its role in protein exit from the ER by COPII-coated vesicles. To test this, we inhibited COPII vesicle formation by depleting Sar1, but this did not prevent stress granule formation upon amino acid starvation (Figure S4B; Figure 4E). Similarly, COPI-coated vesicle formation and retrograde transport from the Golgi to the ER were also not required, as stress granule formation was insensitive to incubation with brefeldin A (Figure S4C; Figure 4E). Taken together, transport through the early secretory pathway is not required for stress granule formation upon amino acid starvation, and the role for Sec16 in their formation lies somewhere else.
Sec16 Is Required for Rin Stability
We noticed that in Sec16-depleted cells the fluorescence level of Rin was low (Figure 4A). This was confirmed by western blot (Figure 4F), where, in the absence of Sec16, Rin level in amino acid-starved cells was reduced compared to control cells. This appears to be specific because, although FMR1 was partly affected, Caprin and CF68 (CG7185) were not. This result suggests that Sec16 depletion mimics Rin depletion, which, as we have shown above, results in a strong inhibition of stress granule formation (Figures 2A–2A’’).
There are at least three ways by which Sec16 can contribute to the maintenance of Rin protein level. It could stabilize its mRNA, it could promote its translation, and it could stabilize the Rin protein. To test the first hypothesis, we monitored Rin mRNA upon Sec16 depletion and found that it was not affected (Figure S5A). To test the latter hypothesis (whether Sec16 stabilizes Rin protein), we transfected mock- or Sec16-depleted cells with Rin-V5, and we treated them with or without the proteasome inhibitor MG132. This allowed us to trigger Rin expression acutely for a short period of time and monitor its level. If the reduced level of Rin observed upon Sec16 depletion was due to protein degradation, the addition of MG132 should rescue it.
We first confirmed that Rin-V5 level was also very low in Sec16-depleted cells (Figures 5A and 5B, compare lanes 1 and 2). In the presence of MG132, however, Rin-V5 level was partially restored (Figures 5A and 5B, compare lanes 3 and 4), showing that Sec16 stabilizes Rin at the protein level and protects it against degradation through the proteasome. Furthermore, whereas stress granule formation was inhibited in Sec16-depleted starved cells as reported above, MG132 treatment significantly rescued the formation of stress granules (also positive for FMR1; Figure S5B) in cells expressing Rin-V5 (Figure 5B, arrow; Figure 5E). This demonstrates that Sec16 stabilizes Rin protein that can then act as a driver for stress granule formation in amino acid-starved cells
Figure 5. Sec16 Prevents Rin Degradation by the Proteasome.
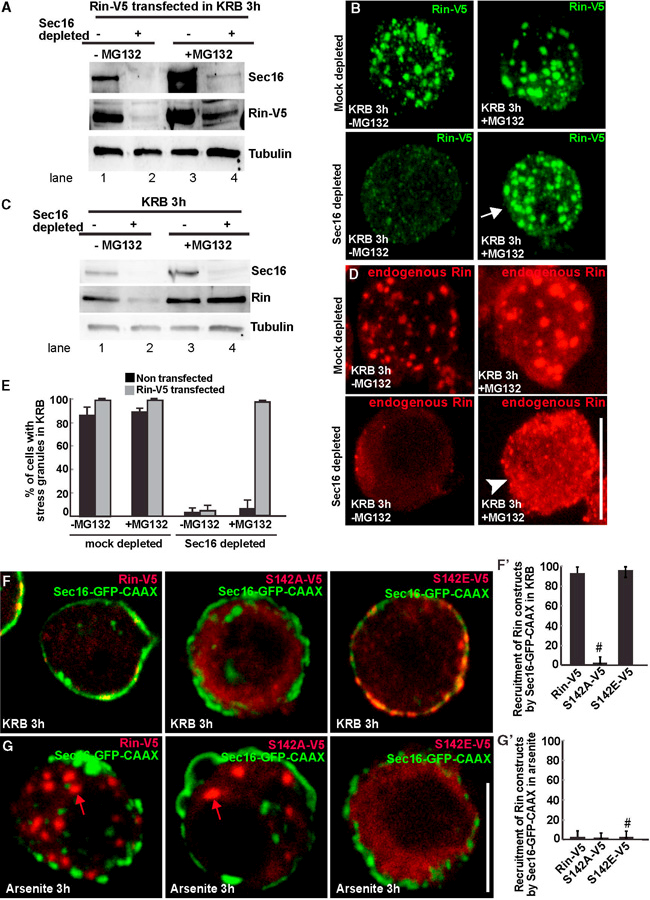
(A) Western blot visualization of Rin-V5 (using an anti-V5 antibody) transfected in mock- and Sec16-depleted cells incubated in Schneider’s and KRB (3 hr), supplemented or not by MG132. Note that MG132 incubation partially rescues Rin-V5 protein level.
(B) IF visualization of Rin-V5 transfected in mock- and Sec16-depleted cells in KRB, supplemented or not with MG132. Note that MG132 incubation completely rescues stress granule formation in Sec16-depleted cells.
(C) Western blot visualization of endogenous Rin in mock- and Sec16-depleted cells in KRB, supplemented or not with MG132. Note that MG132 incubation completely rescues endogenous Rin protein level.
(D) IF visualization of endogenous Rin in mock- and Sec16-depleted cells in KRB, supplemented or not with MG132. Note that MG132 incubation rescues endogenous Rin protein level (C), but it does not rescue stress granule formation.
(E) Quantification of stress granule formation in Rin-V5 transfected and non-transfected cells in mock- and Sec16-depleted cells incubated in KRB, supplemented or not with MG132, expressed in percentage of cell profiles with stress granules.
(F) Confocal section co-visualizing full-length Sec16-GFP-CAAX in cells expressing Rin-V5, S142A-V5, and S142E-V5 Rin mutants upon amino acid starvation (KRB) (F), quantified in F’ (expressed as percentage of Sec16-GFP-CAAX transfected cells). Note that both Rin-V5 and S142E-V5 steadily localize at the plasma membrane in cells expressing Sec16-GFP-CAAX, whereas S142A remains dispersed in the cytoplasm (marked by # in F’).
(G) Confocal section co-visualizing full-length Sec16-GFP-CAAX in cells expressing Rin-V5, S142A-V5, and S142E-V5 Rin mutants upon arsenite treatment (G), quantified in G’ (expressed as percentage of Sec16-GFP-CAAX transfected cells). Note that none of the Rin variants are recruited by Sec16-GFP-CAAX. Furthermore, in agreement with Figures 2E and 2F, Rin-V5 and S142A form stress granules (red arrows). In contrast, S142E remains diffuse in the cytoplasm (marked by # in G’).
Scale bars, 10 µm. Error bars, SD
Sec16 Interacts Specifically with PhosphoRin
However, although the level of endogenous Rin was completely recovered upon MG132 incubation of Sec16-depleted cells (Figure 5C), neither Rin (Figure 5D, arrowhead) nor FMR1 (Figure S5B, arrowhead) was recruited to stress granules, whose formation was not rescued (Figure 5E). This suggests that maintaining the level of endogenous Rin is not enough to form stress granules when Sec16 is absent.
Given that Sec16 is specifically required for stress granule formation upon amino acid starvation, but not upon arsenite treatment (Figures 4C and 4D), and that the phosphorylated form of Rin is specifically recruited to stress granules upon amino acid starvation, we reasoned that Sec16 may specifically interact with Rin phosphorylated on Ser142. To test this, we anchored-away Sec16 to the plasma membrane using CAAX (as described in Figure 2), and we expressed the wild-type Rin-V5 and two Rin mutants S142A and S142E upon amino acid starvation. In agreement with our hypothesis, both Rin-V5 and S142E strongly interacted with Sec16, whereas S142A did not (Figures 5F and 5F’) and also did not form stress granules (as shown in Figures 2E and 2E’).
In striking contrast, none of the Rin forms were recruited to the plasma membrane by Sec16-GFP-CAAX upon arsenite stress (Figures 5G and 5G’). Instead, Rin-V5 and S142A-V5 formed stress granules (red arrows), whereas S142E remained diffuse in the cytoplasm (as shown in Figures 2D and 2D’).
Taken together, we show that the Sec16 requirement for stress granule formation upon amino acid starvation is mediated by its specific interaction with phosphoRin, the form that is incorporated into stress granules upon this type of stress.
DISCUSSION
A Specific Role for PhosphoRin in Stress Granule Formation upon Amino Acid Starvation
Stress granules are formed when protein translation initiation is inhibited by cellular stress that leads to elF2α phosphorylation and the accumulation of untranslated mRNAs (Anderson and Kedersha, 2006; Aulas et al., 2017; Kedersha et al., 2016). Stress granule components start to coalesce through protein-protein interactions mediated by proteins containing regions of low-complexity sequences and displaying multivalence interactions. This process is facilitated by the presence of accumulating free mRNAs (Molliex et al., 2015; Patel et al., 2015). Accordingly, incubation of stressed cells with cycloheximide, which locks ribosomes to the mRNA, blocks stress granule formation, whereas puromycin stimulates their formation. Stress granule formation is also driven by a number of critical factors, including G3BP. In all these respects, the cytoplasmic foci that are formed in Drosophila cells upon amino acid starvation and that are positive for RNA-binding proteins are bona fide stress granules, as they share many features of those found in mammalian cells after arsenite treatment, heat stress, and ER stress (Aulas et al., 2017). Furthermore, we show that Rin, the G3BP Drosophila ortholog, is an essential factor for amino acid starvation-driven stress granules.
However, stress granule formation displays a certain level of heterogeneity. First, some of the signaling cues inducing their formation appear to be different. For instance, although elF2α phosphorylation is required for arsenite-triggered stress granule formation in Drosophila cells, this phosphorylation is not necessary for their formation upon heat stress (Farny et al., 2009). Second, mammalian stress granules that are formed upon different stresses appear to have slightly different content, at least in HAP1 cells (Aulas et al., 2017). Third, stress granules display different material properties. Yeast stress granules possess a solid core made of components that exchange slowly and less dynamically (Buchan and Parker, 2009; Jain et al., 2016), whereas mammalian stress granules have liquid droplet properties (Molliex et al., 2015; Nott et al., 2015; Patel et al., 2015). This is also reflected by the fact that stress granules contain diverse proteomes (Jain et al., 2016).
Remarkably, here we show the phosphorylation status of Rin dictates its differential recruitment to stress granules formed upon arsenite treatment and amino acid starvation. Phosphorylated Rin (on Ser142) is instrumental for stress granule formation upon amino acid starvation, whereas arsenite-driven stress granules require dephosphorylated Rin (as for G3BP in mammalian cells). To the best of our knowledge, this is the first example that clearly demonstrates this differential usage. This further documents that stress granules are more complex and variable than previously anticipated and not just a temporal storage of stalled ribonucleoprotein particles (RNPs).
PhosphoRin Specifically Interacts with Sec16, a Key ERES Component
What determines the use for phosphoRin versus non-phosphoRin in stress granule formation upon different stresses is not fully understood, but the first clue is the discovery that the large hydrophilic ERES protein Sec16 specifically interacts with phosphoRin.
Sec16 adds to the lengthening list of factors modulating stress granule formation via their interaction with G3BP, such as Caprin (Kedersha et al., 2016; Solomon et al., 2007), TDP43 (Aulas et al., 2012), and Usp10 (Panas et al., 2015; Soncini et al., 2001; Kedersha et al., 2016). In addition, YB-1 promotes the translation of G3BP1 mRNA (Somasekharan et al., 2015). Sec16 depletion leads to a reduced level of total Rin, but the remaining Rin pool is enough to lead to stress granule formation upon arsenite treatment, heat stress, and ER stress.
Interestingly, Rin level is further reduced when Sec16- depleted cells are amino acid starved. Thus, Sec16 depletion upon amino acid starvation mimics Rin depletion, explaining the inhibition of stress granule formation upon this stress. This suggests that Sec16 protects Rin against proteasome degradation. It is very often the case that proteins in a complex stabilize each other and that when one partner is absent the other is degraded. However, Rin depletion does not affect Sec16 stability. Sec16 has, therefore, an active role in protecting Rin, perhaps by preventing Rin ubiquitination, a signal for degradation through the proteasome.
However, although proteasome inhibition restores Rin level in starved Sec16-depleted cells, this is not enough to recover stress granule formation. This suggests that Sec16 has an additional role. Given that Sec16 specifically interacts with phosphoRin, this interaction appears necessary not only to protect and stabilize phosphoRin but also to facilitate the role of phosphoRin in stress granule formation upon amino acid starvation. It is possible that the Sec16/phosphoRin complex is recruited to stress granules and/or that Sec16 allows Rin to bind another stress granule partner. In any case, this is strictly specific for amino acid starvation.
Indeed, phosphoRin is the form that is specifically required for stress granule formation upon amino acid starvation. Upon arsenite treatment, the dephosphorylated form of Rin is the form required for stress granule formation both in Drosophila (our results and in mammalian cells; Kedersha et al., 2016; Tourrie`re et al., 2003). This is mirrored by the role of Sec16 that is specifically required for stress granule formation upon amino acid starvation, but not upon heat stress, ER stress, and arsenite treatment. Accordingly, Sec16 does not interact with dephosphorylated Rin.
Does Sec16 have a role in Rin phosphorylation? Rin phosphorylation could take place in the cytoplasm independently of Sec16 that would then recognize phosphoRin and stabilize it. Alternatively, Sec16 could contribute to Rin phosphorylation by acting as a scaffold for the kinase required for Rin phosphorylation. This kinase, which we propose is likely to be (hyper-)activated by amino acid starvation, remains to be identified.
Sec16 Links the Inhibition of Protein Translation and Protein Transport
Why does amino acid starvation require this specific Sec16/ phosphoRin interaction? Why is an ERES component functionally linked to stress granule formation specifically upon this stress? It is likely that this specific interaction elicits the formation of unique stress granules, perhaps storing mRNAs encoding proteins key for survival upon starvation and fitness upon stress relief. In this respect, stress granules formed during amino acid starvation contain the P-body component Tral that is not found in those formed upon heat shock (Jevtov et al., 2015). Interestingly, Tral has been shown to bind mRNAs encoding COPII subunits (Wilhelm et al., 2005), and it is possible that other mRNAs encoding or secretory pathway components may be sequestered and protected inside these stress granules. This would reflect a type of multiplexing that has been observed in neurons (Carson et al., 2008; Smith et al., 2014). Conversely, as ER-translated mRNAs (possibly encoding secretory proteins required in stress recovery) are proposed to escape sequestration to stress granules (Unsworth et al., 2010), Sec16 interaction with stress granule components might restrict stress granule formation to specific sites away from these mRNAs.
Last, the enrichment of phosphoRin (and the presence of Sec16) in amino acid starvation-driven stress granules might change their material properties and, consequently, their dynamics. This is suggested by the large size of the S142E-positive stress granules and by their poor reversibility. Consequently, the exchange of components with the surrounding cytoplasm might be reduced in phosphoRin-based stress granules when compared to stress granules that depend on the dephosphorylated form of Rin (and G3BP). This is supported by fluorescence recovery after photobleaching (FRAP) experiments. G3BP-positive arsenite-driven stress granules show full recovery in less than 1 s (Patel et al., 2015), whereas the recovery of stress granules formed upon amino acid starvation is an order of magnitude slower (Zacharogianni et al., 2014). The biological relevance of this difference is, however, not fully understood.
Overall, the results presented here are in congruence with evidence of the link among protein translation, RNA metabolism, and the secretory pathway. Stress granules are formed in response to ER stress (McEwen et al., 2005; our data). P-bodies also localize in close proximity to the ER and increase in number in response to ER homeostasis perturbations and in Arf1 yeast mutant (Kilchert et al., 2010). Last, ER-resident proteins are shown to regulate P-body formation in yeast (Weidner et al., 2014).
Sec16, a Versatile Scaffold Protein in Basal and Stressed Conditions
Our results provide further evidences of the versatility of Sec16. In growing conditions, mammalian Sec16 exists as two isoforms that are both localized to the ERES (Watson et al., 2006; Bhatta-charyya and Glick, 2007) but have non-redundant functions in humans. Whereas Sec16A is classically required for the ER exit of proteins destined to the Golgi and the plasma membrane (Sprangers and Rabouille, 2015), Sec16B specializes in transport to peroxisomes (Budnik et al., 2011; Yonekawa et al., 2011). Furthermore, Sec16 exons have been shown to be alternatively spliced upon T cell activation (Martinez et al., 2012), and increased expression of the Sec16 isoform containing exon 29 leads to an increased number of ERESs and more efficient COPII transport in activated T cells (Wilhelmi et al., 2016). In this regard, Sec16 is also specifically phosphorylated by ERK2 upon serum stimulation in mammalian cells, leading to an increase in the number of ERESs and a larger secretory capacity (Farhan et al., 2010). Sec16 also interacts with LKKR2, albeit in a kinase activity-independent fashion (Cho et al., 2014), and with ULK (Atg1) in non-stressed conditions (Joo et al., 2016).
Sec16 also plays key roles in the response to stress, for instance, to ER stress where it appears to mediate the Golgi bypass of transmembrane proteins (Piao et al., 2017), but also to nutrient stress (Zacharogianni et al., 2011). Amino acid starvation is an interesting stress as it triggers the formation of two stress assemblies in the same time frame, both requiring Sec16 but in two different manners: The first, the MARylation of Sec16 on its C terminus by ER-localized dPARP16, is an event that is enough to trigger the formation of Sec bodies (Aguilera-Gomez et al., 2016). The second is the Sec16 interaction and stabilization of phosphoRin, leading to the formation of stress granules. Interestingly, neither of these is linked to the Sec16 role in protein exit from the ER or COPII-coated vesicle dynamics (Zacharogianni et al., 2014; this work).
Taken together, this demonstrates the versatility and capacity of the large scaffold protein Sec16 to regulate very diverse cellular processes, many of them pro-survival. Therefore, more Sec16 interactors need to be identified and studied.
EXPERIMENTAL PROCEDURES
Cell Culture, Amino Acid Starvation, Depletions, and Transfections
Drosophila S2 cells (mycoplasma free) were cultured in Schneider’s medium (Sigma) supplemented with 10% insect-tested fetal bovine serum at 26o C, as described previously (Kondylis and Rabouille, 2003; Kondylis et al., 2007). Amino acid starvation of cells for 3 or 4 hr was performed using Krebs Ringer’s Bicarbonate buffer (KRB: 10 mM D(+) Glucose, 0.5 mM MgCl2, 4.5 mM KCl, 121 mM NaCl, 0.7 mM Na2HPO4, 1.5 mM NaH2PO4, and 15 mM sodium bicarbonate) at pH 7.4 (Aguilera-Gomez et al., 2016; Zacharogianni et al., 2014). Note that the starvation buffer did not contain any dialysed fetal bovine serum (FBS), but the addition of this serum did not alter stress granule formation (see Figure S1A).
Wild-type Drosophila S2 cells were depleted by double-stranded (ds)RNAi, as previously described (Aguilera-Gomez et al., 2016). Cells were analyzed after incubation with dsRNAs for 5 days, typically leading to depletion in more than 90% of the cells.
Transient transfections of posttranslational modifications (PMT) constructs (see below) were performed using Effectene transfection reagent (301425, QIAGEN) according to the manufacturer’s instructions. Expression was induced 48 hr after transfection with 1 mM CuSO4 for 1.5 hr (Zacharogianni and Rabouille, 2013)
Antibodies
The antibodies used were as follows: Rabbit polyclonal anti-Sec16 (Ivan et al., 2008) 1:800 immunofluorescence (IF), 1:2,500 western blot (WB); Mouse monoclonal anti-V5 (Thermo Fisher Scientific 46-0705, 1:500 IF); Mouse monoclonal anti-V5 (Life Technologies R960, 1:500 IF, 1:2,000 WB followed by horseradish peroxidase (HRP)-conjugated sheep-anti-mouse HRP Na931, GE Healthcare, 1:2,000); Mouse monoclonal anti-FMR1 (RRID: AB_528251, DSHB supernatant clone 5A11, 1:800 IF, 1:2,000 WB); Rabbit anti-Caprin (1:500 IF, 1:2,000 WB; Reich and Papoulas, 2012); Rabbit polyclonal anti-phospho-eIF2α (S51) (Cell Signaling Technology 9721s, 1:1,000 WB); anti-Tral (gift from A. Nakamura, 1:200 IF); rat anti-eIF4E (gift from A. Na- kamura, 1:200 IF, with methanol fixation); Mouse monoclonal anti-a-tubulin (Sigma T5168, 1:100,000 WB); and Rabbit polyclonal anti-CF68 (CG7185, gift from Z. Dominski).
The Rabbit polyclonal anti-Rin is described in the Supplemental Experimental Procedures and Figure S2D and was used at 1:500.
PMT-DNA Constructs and dsRNAs
All the primers used for generating the DNA constructs and RNAi probes and the procedure are included in the Supplemental Experimental Procedures.
IF and Immunoelectron Microscopy
Drosophila S2 cells were plated on glass coverslips, treated as described, fixed in 4% paraformaldehyde (PFA) in PBS for 20 min, and processed for IF as previously described (Kondylis et al., 2007; Zacharogianni and Rabouille, 2013). Samples were viewed under a Leica SPE confocal microscope using a 633 oil lens and two to four times zoom; 14 to 20 planes were projected to capture the whole cell that is displayed unless indicated otherwise. Unless otherwise mentioned, profiles shown are projections of the whole cell profile.
The immunoelectron microscopy (immuno-EM) of FRM1, TRAL, and Sec16 was performed as described previously (Kondylis et al., 2007; van Donselaar et al., 2007) on ultrathin frozen section (70 nm) of S2 cells that had been amino acid starved for 3 hr.
Immunoprecipitation and Mass Spectrometry
For Sec16 immunoprecipitation, 200-300 million S2 cells were grown and starved in Krebs Ringer’s Bicarbonate buffer (KRB) for 4 hr. Cell lysate was prepared by incubating cells for 20 min on ice in lysis buffer (10% glycerol, 1% Triton X-100, 50 mM TrisHCl [pH 7.5], 150 mM NaCl, 50 mM NaF, 25 mM Na2-glycerophosphate, 1 mM Na2VO4, 5 mM EDTA, and tablet protease inhibitors tablet [Roche]). Protein A bead slurry was washed with lysis buffer and incubated for 1 hr at 4° C with 20 mg control IgG and anti Sec16 IgG. Subsequently, the beads where incubated for 2 hr at 4° C with cell lysate. After washing with lysis buffer, beads where boiled for 10 min in sample buffer followed by SDS-PAGE, and western blot was performed.
Mass spectrometry is described in the Supplemental Experimental Procedures. To be scored as Sec16-interacting proteins, candidates were required to be present in at least two experiments (of four biological replicates), with at least two peptides per experiment and experimental conditions (growing and/ or starvation); and (2) have an intensity of log2 (IgG:SEC16) R1 or, in cases were proteins were not found in IgG control, have an intensity >0. See Table S2 reporting the frequency of identification and the growth conditions. Both categories are color coded and sortable.
Heat Stress, Arsenite, and DTT Treatments
Heat stress was performed on 2 3 106 Drosophila S2 cells in an oven at 37° C (Thermo Electron) for 3 hr as described in Jevtov et al. (2015). Treatments with cycloheximide (10 μM) and puromycin (10 μg/mL), NaAsO2 (0.5 mM), and DTT (2.0 mM, stock in DMSO) were performed at 26° C for 3 hr.
Polysome Profiles
Polysome profiles were generated as described in Pereboom et al. (2011).
Quantification and Statistics
Two to five biological replicates were performed per experiment. For IF of depleted or treated cells, at least four fields per experiment were analyzed comprising at least 50 cells. For transfected cells, at least 30 cells were analyzed. Results are expressed as SD and SEM.
SUPPLEMENTAL INFORMATION
Supplemental Information includes Supplemental Experimental Procedures, five figures, and two tables and can be found with this article online at http://dx.doi.org/10.1016/j.celrep.2017.06.042.
Supplementary Material
Highlights.
Sec16 is specifically required for stress granule formation upon amino acid starvation
Sec16 specifically interacts with phosphorylated Drosophila G3BP (phosphoRin)
PhosphoRin is the form required for stress granule formation upon amino acid starvation
Dephosphorylated Rin is required upon arsenite treatment and does not interact with Sec16
ACKNOWLEDGMENTS
The polysomes profiles were generated by the group of Alyson McInnes at the Hubrecht Institute. Financial support for this paper was provided by the Hubrecht Institute of the KNAW and a grant from NWO (822-020-016 to C.R.). Work in the Behrends lab is supported by the Deutsche Forschungsgemein-schaft within the framework of EXC 1010 SyNergy and SFB1177. Work in the Gavis lab was supported by grant R01 GM061107 from the NIH.
Footnotes
AUTHOR CONTRIBUTIONS
Conceptualization, A.A.-G., M.Z., and C.R.; Methodology, A.A.-G., M.Z., M.v.O., H.G., R.G., T.V., and C.B.; Formal Analysis, A.A.-G., M.Z., C.R., and C.B.; Resources, K.S.S. and E.A.G.; Writing, Review & Editing, C.R. and A.A.-G.; Supervision, C.R., C.B., and E.A.G.
REFERENCES
- Aguilera-Gomez A, van Oorschot MM, Veenendaal T, and Rabouille C (2016). In vivo vizualisation of mono-ADP-ribosylation by dPARP16 upon amino-acid starvation. eLife 5, e21475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson P, and Kedersha N (2002). Stressful initiations. J. Cell Sci 115, 3227–3234. [DOI] [PubMed] [Google Scholar]
- Anderson P, and Kedersha N (2006). RNA granules. J. Cell Biol 172, 803–808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson P, and Kedersha N (2008). Stress granules: the Tao of RNA triage. Trends Biochem. Sci 33, 141–150. [DOI] [PubMed] [Google Scholar]
- Aulas A, Stabile S, and Vande Velde C (2012). Endogenous TDP-43, but not FUS, contributes to stress granule assembly via G3BP. Mol. Neurodegener 7, 54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aulas A, Fay MM, Lyons SM, Achorn CA, Kedersha N, Anderson P, and Ivanov P (2017). Stress-specific differences in assembly and composition of stress granules and related foci. J. Cell Sci 130, 927–937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhattacharyya D, and Glick BS (2007). Two mammalian Sec16 homologues have nonredundant functions in endoplasmic reticulum (ER) export and transitional ER organization. Mol. Biol. Cell 18, 839–849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buchan JR, and Parker R (2009). Eukaryotic stress granules: the ins and outs of translation. Mol. Cell 36, 932–941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Budnik A, Heesom KJ, and Stephens DJ (2011). Characterization of human Sec16B: indications of specialized, non-redundant functions. Sci. Rep 1, 77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carson JH, Gao Y, Tatavarty V, Levin MK, Korza G, Francone VP, Kosturko LD, Maggipinto MJ, and Barbarese E (2008). Multiplexed RNA trafficking in oligodendrocytes and neurons. Biochim. Biophys. Acta 1779, 453–458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cho HJ, Yu J, Xie C, Rudrabhatla P, Chen X, Wu J, Parisiadou L, Liu G, Sun L, Ma B, et al. (2014). Leucine-rich repeat kinase 2 regulates Sec16A at ER exit sites to allow ER-Golgi export. EMBO J 33, 2314–2331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cobos Jiménez V, Martinez FO, Booiman T, van Dort KA, van de Klundert MA, Gordon S, Geijtenbeek TB, and Kootstra NA (2015). G3BP1 restricts HIV-1 replication in macrophages and T-cells by sequestering viral RNA. Virology 486, 94–104. [DOI] [PubMed] [Google Scholar]
- Connerly PL, Esaki M, Montegna EA, Strongin DE, Levi S, Soderholm J, and Glick BS (2005). Sec16 is a determinant of transitional ER organization. Curr. Biol 15, 1439–1447. [DOI] [PubMed] [Google Scholar]
- Cox J, and Mann M (2008). MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nature biotechnology 26, 1367–1372. [DOI] [PubMed] [Google Scholar]
- Damgaard CK, and Lykke-Andersen J (2011). Translational coregulation of 5’TOP mRNAs by TIA-1 and TIAR. Genes & development 25, 2057–2068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farhan H, Wendeler MW, Mitrovic S, Fava E, Silberberg Y, Sharan R, Zerial M, and Hauri HP (2010). MAPK signaling to the early secretory pathway revealed by kinase/phosphatase functional screening. J. Cell Biol 189, 997–1011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farny NG, Kedersha NL, and Silver PA (2009). Metazoan stress granule assembly is mediated by P-eIF2alpha-dependent and -independent mechanisms. RNA 15, 1814–1821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilks N, Kedersha N, Ayodele M, Shen L, Stoecklin G, Dember LM, and Anderson P (2004). Stress granule assembly is mediated by prion-like aggregation of TIA-1. Mol. Biol. Cell 15, 5383–5398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hughes H, Budnik A, Schmidt K, Palmer KJ, Mantell J, Noakes C, Johnson A, Carter DA, Verkade P, Watson P, and Stephens DJ (2009). Organisation of human ER-exit sites: requirements for the localisation of Sec16 to transitional ER. J. Cell Sci 122, 2924–2934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ivan V, de Voer G, Xanthakis D, Spoorendonk KM, Kondylis V, and Rabouille C (2008). Drosophila Sec16 mediates the biogenesis of tER sites upstream of Sar1 through an arginine-rich motif. Mol. Biol. Cell 19, 4352–4365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ivan V, de Voer G, Xanthakis D, Spoorendonk KM, Kondylis V, and Rabouille C (2008). Drosophila Sec16 mediates the biogenesis of tER sites upstream of Sar1 through an arginine-rich motif. Molecular biology of the cell 19, 4352–4365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jain S, Wheeler JR, Walters RW, Agrawal A, Barsic A, and Parker R (2016). ATPase-Modulated Stress Granules Contain a Diverse Proteome and Substructure. Cell 164, 487–498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jevtov I, Zacharogianni M, van Oorschot MM, van Zadelhoff G, Aguilera- Gomez A, Vuillez I, Braakman I, Hafen E, Stocker H, and Rabouille C (2015). TORC2 mediates the heat stress response in Drosophila by promoting the formation of stress granules. J. Cell Sci 128, 2497–2508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joo JH, Wang B, Frankel E, Ge L, Xu L, Iyengar R, Li-Harms X, Wright C, Shaw TI, Lindsten T, et al. (2016). The Noncanonical Role of ULK/ATG1 in ER-to-Golgi Trafficking Is Essential for Cellular Homeostasis. Mol. Cell 62, 982. [DOI] [PubMed] [Google Scholar]
- Kedersha NL, Gupta M, Li W, Miller I, and Anderson P (1999). RNA-binding proteins TIA-1 and TIAR link the phosphorylation of eIF-2 alpha to the assembly of mammalian stress granules. J. Cell Biol 147, 1431–1442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kedersha N, Panas MD, Achorn CA, Lyons S, Tisdale S, Hickman T, Thomas M, Lieberman J, McInerney GM, Ivanov P, and Anderson P (2016). G3BP-Caprin1-USP10 complexes mediate stress granule condensation and associate with 40S subunits. J. Cell Biol 212, 845–860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kilchert C, Weidner J, Prescianotto-Baschong C, and Spang A (2010). Defects in the secretory pathway and high Ca2+ induce multiple P-bodies. Mol. Biol. Cell 21, 2624–2638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kondylis V, and Rabouille C (2003). A novel role for dp115 in the organization of tER sites in Drosophila. J. Cell Biol 162, 185–198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kondylis V, van Nispen tot Pannerden HE, Herpers B, Friggi-Grelin F, and Rabouille C (2007). The golgi comprises a paired stack that is separated at G2 by modulation of the actin cytoskeleton through Abi and Scar/WAVE. Dev. Cell 12, 901–915. [DOI] [PubMed] [Google Scholar]
- Martinez NM, Pan Q, Cole BS, Yarosh CA, Babcock GA, Heyd F, Zhu W, Ajith S, Blencowe BJ, and Lynch KW (2012). Alternative splicing networks regulated by signaling in human T cells. RNA 18, 1029–1040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McEwen E, Kedersha N, Song B, Scheuner D, Gilks N, Han A, Chen JJ, Anderson P, and Kaufman RJ (2005). Heme-regulated inhibitor kinase-mediated phosphorylation of eukaryotic translation initiation factor 2 inhibits translation, induces stress granule formation, and mediates survival upon arsenite exposure. J. Biol. Chem 280, 16925–16933. [DOI] [PubMed] [Google Scholar]
- Min L, Ruan Y, Shen Z, Jia D, Wang X, Zhao J, Sun Y, and Gu J (2015). Overexpression of Ras-GTPase-activating protein SH3 domain-binding protein 1 correlates with poor prognosis in gastric cancer patients. Histopathology 67, 677–688. [DOI] [PubMed] [Google Scholar]
- Molliex A, Temirov J, Lee J, Coughlin M, Kanagaraj AP, Kim HJ, Mittag T, and Taylor JP (2015). Phase separation by low complexity domains promotes stress granule assembly and drives pathological fibrillization. Cell 163, 123–133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nott TJ, Petsalaki E, Farber P, Jervis D, Fussner E, Plochowietz A, Craggs TD, Bazett-Jones DP, Pawson T, Forman-Kay JD, and Baldwin AJ (2015). Phase transition of a disordered nuage protein generates environmentally responsive membraneless organelles. Mol. Cell 57, 936–947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panas MD, Schulte T, Thaa B, Sandalova T, Kedersha N, Achour A, and McInerney GM (2015). Viral and cellular proteins containing FGDF motifs bind G3BP to block stress granule formation. PLoS Pathog 11, e1004659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patel A, Lee HO, Jawerth L, Maharana S, Jahnel M, Hein MY, Stoynov S, Mahamid J, Saha S, Franzmann TM, et al. (2015). A Liquid-to-Solid Phase Transition of the ALS Protein FUS Accelerated by Disease Mutation. Cell 162, 1066–1077. [DOI] [PubMed] [Google Scholar]
- Pereboom TC, van Weele LJ, Bondt A, and MacInnes AW (2011). A zebrafish model of dyskeratosis congenita reveals hematopoietic stem cell formation failure resulting from ribosomal protein-mediated p53 stabilization. Blood 118, 5458–5465. [DOI] [PubMed] [Google Scholar]
- Piao H, Kim J, Noh SH, Kweon HS, Kim JY, and Lee MG (2017). Sec16A is critical for both conventional and unconventional secretion of CFTR. Sci. Rep 7, 39887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Protter DS, and Parker R (2016). Principles and Properties of Stress Granules. Trends Cell Biol 26, 668–679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reich J, and Papoulas O (2012). Caprin controls follicle stem cell fate in the Drosophila ovary. PLoS ONE 7, e35365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shevchenko A, Tomas H, Havlis J, Olsen JV, and Mann M (2006). In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nature protocols 1, 2856– 2860. [DOI] [PubMed] [Google Scholar]
- Smith R, Rathod RJ, Rajkumar S, and Kennedy D (2014). Nervous translation, do you get the message? A review of mRNPs, mRNA-protein interactions and translational control within cells of the nervous system. Cell. Mol. Life Sci 71, 3917–3937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Solomon S, Xu Y, Wang B, David MD, Schubert P, Kennedy D, and Schrader JW (2007). Distinct structural features of caprin-1 mediate its interaction with G3BP-1 and its induction of phosphorylation of eukaryotic translation initiation factor 2alpha, entry to cytoplasmic stress granules, and selective interaction with a subset of mRNAs. Mol. Cell. Biol 27, 2324–2342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Somasekharan SP, El-Naggar A, Leprivier G, Cheng H, Hajee S, Grunewald TG, Zhang F, Ng T, Delattre O, Evdokimova V, et al. (2015). YB-1 regulates stress granule formation and tumor progression by translationally activating G3BP1. J. Cell Biol 208, 913–929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soncini C, Berdo I, and Draetta G (2001). Ras-GAP SH3 domain binding protein (G3BP) is a modulator of USP10, a novel human ubiquitin specific protease. Oncogene 20, 3869–3879. [DOI] [PubMed] [Google Scholar]
- Sprangers J, and Rabouille C (2015). SEC16 in COPII coat dynamics at ER exit sites. Biochem. Soc. Trans 43, 97–103. [DOI] [PubMed] [Google Scholar]
- Tourrière H, Chebli K, Zekri L, Courselaud B, Blanchard JM, Bertrand E, and Tazi J (2003). The RasGAP-associated endoribonuclease G3BP assembles stress granules. J. Cell Biol 160, 823–831. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Tsai WC, and Lloyd RE (2014). Cytoplasmic RNA Granules and Viral Infection. Annu. Rev. Virol 1, 147–170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Unsworth H, Raguz S, Edwards HJ, Higgins CF, and Yagϋe E (2010). mRNA escape from stress granule sequestration is dictated by localization to the endoplasmic reticulum. FASEB J 24, 3370–3380. [DOI] [PubMed] [Google Scholar]
- van Donselaar E, Posthuma G, Zeuschner D, Humbel BM, and Slot JW (2007). Immunogold labeling of cryosections from high-pressure frozen cells. Traffic 8, 471–485. [DOI] [PubMed] [Google Scholar]
- Watson P, Townley AK, Koka P, Palmer KJ, and Stephens DJ (2006). Sec16 defines endoplasmic reticulum exit sites and is required for secretory cargo export in mammalian cells. Traffic 7, 1678–1687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weidner J, Wang C, Prescianotto-Baschong C, Estrada AF, and Spang A (2014). The polysome-associated proteins Scp160 and Bfr1 prevent P body formation under normal growth conditions. J. Cell Sci 127, 1992–2004. [DOI] [PubMed] [Google Scholar]
- White JP, Cardenas AM, Marissen WE, and Lloyd RE (2007). Inhibition of cytoplasmic mRNA stress granule formation by a viral proteinase. Cell Host Microbe 2, 295–305. [DOI] [PubMed] [Google Scholar]
- Wilhelm JE, Buszczak M, and Sayles S (2005). Efficient protein trafficking requires trailer hitch, a component of a ribonucleoprotein complex localized to the ER in Drosophila. Dev. Cell 9, 675–685. [DOI] [PubMed] [Google Scholar]
- Wilhelmi I, Kanski R, Neumann A, Herdt O, Hoff F, Jacob R, Preußner M, and Heyd F (2016). Sec16 alternative splicing dynamically controls COPII transport efficiency. Nat. Commun 7, 12347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang WH, Yu JH, Gulick T, Bloch KD, and Bloch DB (2006). RNA- associated protein 55 (RAP55) localizes to mRNA processing bodies and stress granules. RNA 12, 547–554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yonekawa S, Furuno A, Baba T, Fujiki Y, Ogasawara Y, Yamamoto A, Tagaya M, and Tani K (2011). Sec16B is involved in the endoplasmic retic- ulum export of the peroxisomal membrane biogenesis factor peroxin 16 (Pex16) in mammalian cells. Proc. Natl. Acad. Sci. USA 108, 12746–12751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zacharogianni M, and Rabouille C (2013). Trafficking along the secretory pathway in Drosophila cell line and tissues: a light and electron microscopy approach. Methods Cell Biol 118, 35–49. [DOI] [PubMed] [Google Scholar]
- Zacharogianni M, Kondylis V, Tang Y, Farhan H, Xanthakis D, Fuchs F, Boutros M, and Rabouille C (2011). ERK7 is a negative regulator of protein secretion in response to amino-acid starvation by modulating Sec16 membrane association. EMBO J 30, 3684–3700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zacharogianni M, Aguilera-Gomez A, Veenendaal T, Smout J, and Rabouille C (2014). A stress assembly that confers cell viability by preserving ERES com- ponents during amino-acid starvation. eLife 3, e04132. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.