

Abstract
There is an urgent need for strategies to combat dengue virus (DENV) infection; a major global threat. We reported that the cellular kinases AAK1 and GAK regulate intracellular trafficking of multiple viruses and that sunitinib and erlotinib, approved anticancer drugs with potent activity against these kinases, protect DENV-infected mice from mortality. Nevertheless, further characterization of the therapeutic potential and underlying mechanism of this approach is required prior to clinical evaluation. Here, we demonstrate that sunitinib/erlotinib combination achieves sustained suppression of systemic infection at approved dose in DENV-infected IFN-α/β and IFN-γ receptor–deficient mice. Nevertheless, treatment with these blood-brain barrier impermeable drugs delays, yet does not prevent, late-onset paralysis; a common manifestation in this immunodeficient mouse model but not in humans. Sunitinib and erlotinib treatment also demonstrates efficacy in human primary monocyte-derived dendritic cells. Additionally, DENV infection induces expression of AAK1 transcripts, but not GAK, via single-cell transcriptomics, and these kinases are important molecular targets underlying the anti-DENV effect of sunitinib and erlotinib. Lastly, sunitinib/erlotinib combination alters inflammatory cytokine responses in DENV-infected mice. These findings support feasibility of repurposing sunitinib/erlotinib combination as a host-targeted antiviral approach and contribute to understanding its mechanism of antiviral action.
Keywords: Dengue virus, Antivirals, Kinase inhibitors, Drug repurposing, Virus-host interactions
1. Introduction
Dengue virus (DENV) is a global threat estimated to infect 390 million people annually (Bhatt et al., 2013). The majority of individuals infected with any of the four DENV serotypes present with acute dengue fever. A fraction (~5–20%) of dengue patients will progress to severe dengue manifested by bleeding, shock, organ failure, and death. The greatest risk factor for severe dengue is secondary infection with a heterologous DENV serotype causing antibody-dependent enhancement (ADE) (Katzelnick et al., 2017; Wang et al., 2017). ADE has also been hindering the development of effective dengue vaccines, with recent data indicating increased dengue severity in vaccinated children (Halstead and Russell, 2016). Moreover, there are currently no approved therapeutics for the treatment of dengue.
The current “one drug-one bug” antiviral approach is not easily scalable to address the unmet clinical need posed by emerging viruses and is often associated with emergence of drug resistance (Bekerman and Einav, 2015). Host-targeted antivirals have a potential for broad-spectrum coverage and a higher barrier to resistance (Bekerman and Einav, 2015). The host-targeted approach is particularly attractive for the treatment of emerging viral infections lacking any treatment, given the opportunity to repurpose already approved, safe drugs that modulate specific host functions (Schor and Einav, 2018).
One cellular process that is usurped by multiple viruses is intracellular membrane traffic, which relies, in part, on the interactions between adaptor protein (AP) complexes and transmembrane cargo. The host cell kinases AP2-associated protein kinase 1 (AAK1) and cyclin G–associated kinase (GAK) regulate clathrin-associated adaptor-cargo trafficking in the endocytic and secretory pathways (Ricotta et al., 2002; Umeda et al., 2000; Zhang et al., 2005), in part by recruiting AP2 and AP1, respectively, to relevant membranes and enhancing their binding affinity for cargo (Ghosh and Kornfeld, 2003; Ricotta et al., 2002). We and others have reported roles for APs in the lifecycles of multiple unrelated viruses (Bekerman et al., 2017; Bhattacharyya et al., 2011; Neveu et al., 2012; Neveu et al., 2015; Ohno et al., 1997). Our work demonstrated that through AP1 and AP2 phosphorylation, AAK1 and GAK regulate early and late stages of the viral lifecycle, thereby representing “master regulators” of viral infection (Bekerman et al., 2017; Neveu et al., 2012; Neveu et al., 2015). Moreover, we reported that sunitinib and erlotinib, approved anticancer drugs with potent binding to AAK1 and GAK (dissociation constant [KD] of 11 and 3.1 nM, respectively (Karaman et al., 2008)), block entry and late stages in the lifecycle of the 4 DENV serotypes and multiple viruses in vitro (Bekerman et al., 2017; Kovackova et al., 2015; Neveu et al., 2012; Neveu et al., 2015). Treatment with combinations of sunitinib and erlotinib revealed synergistic inhibition of DENV2 with a high genetic barrier to resistance (Bekerman et al., 2017). In murine models of dengue and Ebola, 5-day treatment with the individual drugs provided partial or no protection, yet, consistent with our in vitro synergy results, sunitinib/erlotinib combinations significantly protected mice from morbidity and mortality (Bekerman et al., 2017). Nevertheless, DENV-infected mice were monitored for 10–12 days only and relatively high drug doses were used. To support planned efforts to study the safety and efficacy of sunitinib/erlotinib combinations in dengue patients and potentially in Ebola patients in future outbreaks (ClinicalTrials.gov NCT02380625), it is therefore important to examine efficacy in preventing disease relapse, efficacy of lower dosing regimens, and antiviral effect in primary human cells. Moreover, whereas our mechanistic studies indicated that inhibition of intracellular viral trafficking mediates the anti-DENV effect of sunitinib and erlotinib, other mechanisms of antiviral action were incompletely characterized. Here, we integrate in vitro, ex vivo and in vivo data to address these gaps in knowledge and provide further evidence for the therapeutic potential of this repurposed approach and insight into its mechanism of action (MOA).
2. Materials and Methods
2.1. Virus constructs.
DENV2 (New Guinea C strain) Renilla reporter plasmid used for ex vivo assays was a gift from Pei-Yong Shi (Xie et al., 2013; Zou et al., 2011). DENV 16681 plasmid (pD2IC-30P-NBX) used for transcriptomics was a gift from Claire Huang (CDC)(Huang et al., 2010). Mouse-adapted N124D/K128E DENV2 PL046 used for in vivo studies was a gift from Sujan Shresta (LIAI).
2.2. Cells.
Huh7 (Apath LLC), BHK-21 (ATCC), and T-REx 293 (ThermoFisher) cells were grown in DMEM (Mediatech) supplemented with 10% FBS (Omega Scientific), nonessential amino acids, 1% L-glutamine, and 1% penicillin-streptomycin (ThermoFisher Scientific) (5% CO2, 37°C). C6/36 cells were grown in Leibovitz’s L-15 media (CellGro) supplemented with 10% FBS and 1% HEPES (0% CO2, 28°C).
2.3. Generation of MDDCs
(Rodriguez-Madoz et al., 2010). Buffy coats were obtained from the Stanford Blood Center. CD14+ cells were purified from by EasySep™ Human Monocyte Enrichment Kit without CD16 Depletion (Stemcell Technologies). Cells were seeded in 6-well plates (2×106 cells/well), stimulated with 500 U/ml G-CSF and 1,000 U/ml IL-4 (Pepro tech), and incubated at 37°C for 6 days prior to DENV infection (MOI 1).
2.4. Virus Production.
DENV RNA was transcribed in vitro by mMessage/mMachine (Ambion) kits and electroporated into BHK-21 cells. Supernatant was harvested at day 10 and titered by plaque assays on BHK-21 cells or used to inoculate C6/36 cells to amplify the virus. For mice studies, supernatant was concentrated 100-fold (50,000xg).
2.5. In vivo dengue studies.
AG B6 mice were generated by crossing C57BL/6.129S2-Ifnar1tm1Agt/Mmjax (N10) with C57BL/6.129S7-Ifngr1tm1Agt/J (N11). Age-matched, male and female mice at 8–10 weeks of age were used. Mouse adapted DENV2 (3×103 or 3×104 pfu produced in BHK-21 cells) was inoculated retro-orbitally under general and local anesthesia. Sunitinib (Selleckchem) and erlotinib (LC Laboratories) (both >95% pure by mass spectrometry) were dissolved in 10% Captisol. 100–200 μl of drugs or vehicle were administered intraperitoneally once daily starting at inoculation for 5 days. Mice were monitored twice daily. Moribund animals were euthanized by CO2 inhalation. Whole blood was obtained retro-orbitally under general and local anesthesia. Tissues were harvested following euthanasia.
2.6. Virus-inclusive single-cell RNA-Seq.
As described (Zanini et al., 2018), Huh7 cells were infected with DENV at time 0 at MOI 0, 1, or 10, then harvested at different time points, sorted and lysed into single wells. Whole genome mRNA and viral RNA (vRNA) were reverse transcribed and amplified from each cell. Cells were then screened for virus infection by qPCR. Libraries were made and sequenced on an illumina NextSeq 500 with a coverage of ~400,000 reads per cell.
2.7. Gain-of-function assays.
Doxycycline-inducible cell lines were established to overexpress AAK1 or GAK via the Flp-In™ recombination system (ThermoFisher)(Torres et al., 2009). T-REx293 cells with a pFRT/lacZeo site and pcDNA™6/TR were co-transfected with puromycin-resistant vector encoding Flp-In™ recombination target site and pOG44 plasmid containing Flp recombinase followed by puromycin selection. Eight hours post-induction with doxycycline, cells were treated with the drugs, infected with DENV (MOI=0.01) and incubated for 72 hours prior to luciferase and viability assays.
2.8. RNA extraction and quantification
of DENV, AAK1 and GAK RNA were performed as described(Bekerman et al., 2017).
2.9. Viability assays.
AlamarBlue® reagent (Invitrogen) was used according to manufacturer’s protocol. Fluorescence was detected at 560 nm on InfiniteM1000 plate reader (Tecan).
2.10. Luminex assay
(Stanford Human Immune Monitoring Center). Mouse 38-plex magnetic bead kits (eBioscience/Affymetrix) were added to a 96-well plate and washed in a Biotek ELx405 washer. Serum samples were added in duplicate and incubated at RT for 1 hour followed by overnight incubation at 4°C with shaking. Following washing, biotinylated detection antibody was added for 75 minutes at RT with shaking, followed by washing and addition of streptavidin-PE. After 30-minute incubation at RT and washing, reading buffer was added. Plates were read by Luminex 200. Custom assay control beads (Radix Biosolutions) were added to all wells.
2.11. Statistical analysis.
Data were analyzed with R or GraphPad Prism softwares. EC50s were measured by fitting data to a 3-parameter logistic curve. P values were calculated by one- or two-way ANOVA with various multiple comparisons test or by Mann-Whitney-Wilcoxon test. Survival curve p values were calculated by log-ranked Mantel-Cox test. Correlation coefficients of single-cell transcriptomic data were computed as Spearman’s rank correlation ⍴. Error bars are SD of 100 bootstraps over cells.
2.12. Study Approval.
Animal research was conducted under a protocol approved by Stanford Institutional Animal Care and Use Committee and its Institutional Biosafety Committee in compliance with the Animal Welfare Act and other federal regulations relating to experiments involving animals. The Stanford animal facility is AAALAC-accredited. Moribund mice were humanely euthanized on the basis of IACUC-approved criteria.
3. Results
3.1. Sunitinib/erlotinib combination provides sustainable protection of mice from severe dengue-associated mortality but not paralysis.
The IFN-α/β and IFN-γ receptor–deficient murine model of dengue on 129/Sv (AG-129) and C57BL/6 (AG-B6) genetic backgrounds is widely used for testing candidate antiviral compounds (Bekerman et al., 2017; Schul et al., 2007) (Chan et al., 2015). Depending on the DENV strain and inoculum, these mice either develop a systemic infection that mimics severe dengue or late-onset (day 9–30 postinfection) paralysis associated with DENV replication in the brain (Prestwood et al., 2008; Shresta et al., 2006). We previously reported that 5-day treatment with sunitinib/erlotinib combination reduced viral load in serum and tissues and early mortality in AG-B6 and AG-129 mice infected with DENV2 (Bekerman et al., 2017). These experiments were concluded on day 10–12 when the surviving treated animals regained full mobility and displayed weight gain for at least two consecutive days (Bekerman et al., 2017). Here, we sought to assess whether this protection persists longer. Upon inoculation of AG-B6 mice, we initiated once-daily drug administration at a dose shown to be protective (30 mg/kg of each drug) (Bekerman et al., 2017). 100% of vehicle-treated mice succumbed to infection on day 5–7 post-infection, whereas a 5-day intraperitoneal sunitinib/erlotinib treatment protected 70% of the mice until day 14, as reported (Figures 1A, B) (Bekerman et al., 2017). Nevertheless, between days 14 and 25 following initiation of treatment, the surviving mice developed paralysis accompanied by weight loss, and were therefore euthanized soon after the onset of paralysis. No paralysis was detected in uninfected mice treated with sunitinib/erlotinib combination, indicating that dengue infection rather than drug treatment mediates this phenotype (data not shown).
Figure 1. Sunitinib/erlotinib combination protects mice from severe dengue-associated mortality but not from paralysis in a high inoculum model.

(A and B) Weight loss (A) and mortality (B) of DENV-infected AG-B6 mice treated once daily for 5 days with sunitinib/erlotinib (SM/E) combination (30 mg/kg each) or vehicle (n=8 per treatment group). Administration was intraperitoneal and started at inoculation. In A, shown are means±SD. Dotted line in B represents first onset of paralysis. Mice were euthanized soon after the onset of paralysis. C. DENV titer in spleen of AG-B6 mice measured by plaque assay at various time points following once-daily administration of SM/E combination or vehicle for 5 days (n=4 per treatment group). Shown are means±SEM. D. DENV titer in brain, spleen, liver and serum measured by standard plaque assays in AG-B6 mice treated once daily for 5 days with SM/E or vehicle on day 11 postinfection (when the first mouse showed early signs of paralysis, n=6 per treatment group). Representative experiments out of two conducted are shown. Dotted lines represent limit of detection (100 PFU (C) and 10 PFU (D)). *P < 0.05, ***P < 0.001 relative to vehicle control by log-rank test (B) and two-way ANOVA with Bonferroni multiple comparisons test (C). ND, not detected; NA, not applicable.
To determine whether the observed paralysis resulted from relapsing systemic infection following treatment cessation, we measured viral titers in spleens of dengue-infected AG-B6 mice on various days postinfection by plaque assays. A 5-day once-daily treatment with sunitinib/erlotinib combination reduced the viral titers in the spleen by ~1 log relative to control mice on day 2 and 4 postinfection and effectively eradicated viral replication by day 6 (Figure 1C). Splenic viral titers did not increase on days 8 and 10. In contrast, high DENV titers were measured in the spleens of vehicle control mice until day 6, when all animals succumbed to infection. Next, we measured DENV titers in the brain, spleen, liver and serum of AG-B6 mice that recovered from severe dengue following a 5-day sunitinib/erlotinib treatment course. Samples were collected on day 11 when the first mouse displayed early signs of paralysis. DENV replication was detected in the brain of all these mice, yet it was not associated with systemic infection, as viral titers in spleen, liver and serum samples were undetectable (except for a single sample) (Figure 1D).
These results indicate that sunitinib/erlotinib treatment achieves sustained systemic suppression of DENV replication. Nevertheless, treatment does not prevent DENV replication in the brain and late-onset paralysis in this model. We predict that poor BBB permeability of sunitinib and erlotinib hinders their effect in the brain (Oberoi et al., 2013; Verheijen et al., 2017).
3.2. Sunitinib/erlotinib combination delays paralysis onset in a low-inoculum dengue mouse model.
To further study the effect of sunitinib/erlotinib treatment on paralysis, AG-B6 mice were infected at a 10-fold lower DENV inoculum. 100% of sunitinib/erlotinib-treated mice survived beyond day 8 (Figure 2A). In contrast to the high-inoculum model, only ~50% of the vehicle-treated mice succumbed to death prior to day 8, allowing for controlled monitoring of drug effect on paralysis onset and incidence (Figure 2A). All the surviving vehicle control mice developed paralysis on days 8–14 postinoculation (Figure 2B). Sunitinib/erlotinib treatment significantly delayed paralysis onset, and this effect was associated with reduced viral load in the brain relative to vehicle controls, as measured via qRT-PCR assays (Figures 2B, 2C). These data indicate that sunitinib/erlotinib treatment has a favorable, yet incomplete effect on paralysis in this low-inoculum model.
Figure 2. Sunitinib/erlotinib combination delays paralysis onset in a low-inoculum dengue model.
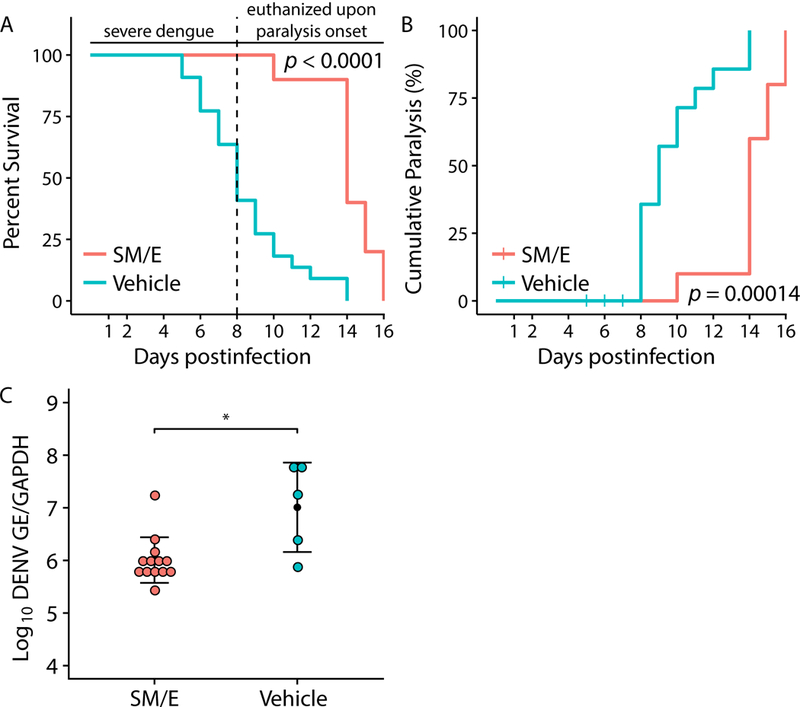
(A and B) Mortality (A) and cumulative percentage of paralysis (B) in AG-B6 mice inoculated with a low inoculum of DENV (3×103 PFU) and treated once daily for 5 days with sunitinib/erlotinib (SM/E) combination (30 mg/kg each) or vehicle (n = 10 per treatment group, per experiment). Administration was intraperitoneal and started at inoculation. Data pooled from two independent experiments is shown. Dotted line represents first onset of paralysis. Mice were euthanized soon after the onset of paralysis. C. DENV load in the brain of AG-B6 mice on day 8–12 postinoculation with a low DENV titer and once daily 5-day treatment with SM/E combination (n = 13) or vehicle (n = 5). *P < 0.05 relative to vehicle control by log-rank test (A, B) and nonparametric Mann-Whitney-Wilcoxon test (C). GE, genome equivalent; GAPDH, Glyceraldehyde 3-phosphate dehydrogenase.
3.3. Lower dose combination of sunitinib/erlotinib is effective in vivo.
Addition of erlotinib decreases sunitinib exposure by 40% (and total active compound exposure, i.e. sunitinib plus its active metabolite, by 20%) relative to sunitinib alone in humans (Blumenschein et al., 2012). We therefore hypothesized that reducing erlotinib dose may enable sunitinib dose reduction without impairing protection. We conducted dose-finding experiments revealing that 5-day once-daily treatment with sunitinib/erlotinib at 25/20 and 20/15 mg/kg reduced morbidity of DENV-infected mice, albeit the weight of mice treated with the two lower dose regimens plateaued at a lower level on day 7 and did not recover soon after (Figure 3A). Nevertheless, treatment with sunitinib/erlotinib at 25/20 and 20/15 mg/kg performed comparably to the 30/30 mg/kg combination in reducing mortality of DENV-infected mice (Figure 3B).. Sunitinib/erlotinib combination at 15/10 mg/kg exhibited a trend toward delayed mortality, albeit this effect did not meet statistical significance (Figures 3A, 3B). These findings provide evidence for protection with this drug combination at levels at or lower than the corresponding approved cancer dose.
Figure 3. Lower dose combinations of sunitinib/erlotinib are effective in vivo.

Weight loss (A) and mortality (B) of DENV-infected AG-B6 mice treated once daily for 5 days with sunitinib/erlotinib (SM/E) combination at the indicated doses (mg/kg) or vehicle (n=8 per treatment group, per experiment). Administration was by gavage and started at inoculation. Data pooled from three independent experiments is shown. In A, shown are means±SD. P by log-rank test are relative to vehicle control or between the various tested groups as indicated (B). N.S, non-significant.
3.4. Sunitinib and erlotinib inhibit DENV infection in human dendritic cells.
To further determine the feasibility of repurposing sunitinib and erlotinib as anti-DENV drugs, we studied their antiviral effect in primary human MDDCs; an established ex vivo model system for DENV (Rodriguez-Madoz et al., 2010). We measured a dose-dependent inhibition of DENV infection following a 3-day drug treatment with an EC50 of 1.962 μM for sunitinib and 4.07 μM for erlotinib by luciferase assays (Figure 4), similar to the effect of these drugs in human hepatoma (Huh7) cells (Bekerman et al., 2017). The concentration range yielding 30 (sunitinib) and 20 fold (erlotinib) reduction in viral infection showed minimal or no cytotoxicity as measured by alamarBlue assays with half-maximal cellular cytotoxicity (CC50) of >20 μM for sunitinib and >50 μM for erlotinib (Figure 4).
Figure 4. Ex vivo antiviral activity of sunitinib and erlotinib in human primary dendritic cells.

Cell viability (blue) and dose response of DENV infection (black) to sunitinib (left) and erlotinib (right) measured by alamarBlue assays and luciferase assays, respectively, 72 hours after infection of primary human MDDCs. Data are plotted relative to vehicle control. Shown is a representative experiment with cells from a single donor, out of 3 independent experiments conducted with cells derived from 3 donors, each with 5 biological replicates; shown are means ± SD.
3.5. AAK1, but not GAK, gene expression correlates with intracellular DENV abundance.
We previously demonstrated a requirement for AAK1 and GAK in DENV infection, thereby validating them as antiviral targets (Bekerman et al., 2017). Because viruses often modulate host gene expression (Geiss et al., 2000; Walsh and Mohr, 2011), we sought to determine the effect of DENV infection on AAK1 and GAK expression. DENV infection minimally increased the level of AAK1 and GAK transcripts in bulk Huh7 cell samples relative to uninfected cells, as measured via qRT-PCR at 24 and/or 48 hours postinfection (Figure 5A). To study this effect while avoiding the averaging process in these bulk samples, we used viscRNA-Seq (virus-inclusive single-cell RNA-Seq), an approach that we recently developed to sequence and quantify the transcriptome of single cells together with the viral RNA (vRNA) from the same cells (Zanini et al., 2018). We previously applied viscRNA-Seq to monitor DENV infection in Huh7 cells at various time points postinfection and discovered extreme heterogeneity in virus abundance and a subset of host genes (Zanini et al., 2018). Here, we directly computed Spearman’s rank correlation coefficient between intracellular virus abundance and transcript level for AAK1 and GAK. The expression of AAK1 significantly increased upon DENV infection at MOI of 1 and 10 in correlation with the intracellular virus abundance within single cells (⍴=0.26 and 0.22, respectively) (Figure 5B). To disentangle the effect of time since infection from the vRNA level within each cell, we computed the same correlation coefficient within single time points. The correlation of AAK1 with virus abundance slightly declined in the first 10 hours but then increased during the course of infection, with a ⍴ value of >0.4 at 50 hours postinfection, indicating a strong correlation (Figure 5C). In contrast, GAK level did not correlate with vRNA level in the same cells (⍴=−0.03˗−0.07) (Figures 5B, 5C).
Figure 5. AAK1, but not GAK, gene expression correlates with intracellular DENV abundance.

AAK1 and GAK transcript levels in bulk (A) and single cell (B-C) DENV infected (MOI=0 (control), 1, 10) Huh7 samples at various time points postinfection, measured via qRT-PCR (A) and virus-inclusive single-cell RNA-Seq (B-C). *P < 0.05, **P < 0.01 in A are relative to vehicle control normalized to TBP (TATA-binding protein) by two-way ANOVA with Tukey’s multiple comparisons test. B. Scatter plots depicting the correlation between vRNA content (viral reads, X axis) and AAK1/GAK gene expression (Y axis) during DENV infection. Each dot represents a single cell, color-coded based on the MOI. ρ represents Spearman correlation coefficients between abundances of dengue vRNA and of AAK1/GAK mRNA from the same cell. C. Correlation values measured for AAK1 (green) and GAK (orange) at various time points postinfection.
3.6. AAK1 and GAK are molecular targets underlying the antiviral effect of sunitinib and erlotinib.
Next, we conducted gain-of-function assays, to confirm that inhibition of AAK1 and GAK is a mechanism underlying the anti-DENV effect of these drugs. Following doxycycline-mediated induction of AAK1 or GAK expression, T-REx 293 cells were infected with a luciferase reporter DENV and treated with sunitinib or erlotinib for 72 hours prior to luciferase and viability assays. Doxycycline-induced AAK1 and GAK expression either partially or completely reversed the antiviral effect of sunitinib and erlotinib, respectively, relative to uninduced cells (Figure 6). These results validate AAK1 and GAK as key mediators of the anti-DENV effect of sunitinib and erlotinib.
Figure 6. AAK1 and GAK are molecular targets underlying the antiviral effect of sunitinib and erlotinib.

A. Expression of the indicated proteins in T-REx 293 cells induced with doxycycline to express AAK1 or GAK or uninduced control cells. B-C. DENV infection normalized to cellular viability in these cells via luciferase and alamarBlue assays, respectively, 72 hours following infection with a luciferase reporter DENV and treatment with sunitinib (B) or erlotinib (C). Shown is a representative experiment out of two conducted, each with 5 replicates. *P<0.05, **P<0.01 relative to induced or uninduced cells treated with DMSO by one-way ANOVA with Fisher’s least significant difference test. Means ± SD are shown.
3.7. Protection of dengue infected mice by sunitinib/erlotinib treatment is associated with altered cytokine responses.
To better understand the MOA underlying the in vivo anti-DENV activity of these drugs, we studied the effect of daily sunitinib/erlotinib treatment on the levels of 38 cytokines in the serum of dengue-infected AG-B6 mice and uninfected controls at 48 and 72 hours postinoculation via a Luminex assay. Sunitinib/erlotinib treatment did not alter cytokine levels in uninfected mice (Figure S1). As reported in this model (Costa et al., 2013; Soo et al., 2017), dengue infection increased the levels of multiple cytokines, including tumor necrosis factor (TNF)-α, interleukin (IL)-6, IL-10, IL-18, monocyte chemoattractant protein (MCP)-1, macrophage inflammatory protein (MIP)-1α, MIP-1β, and interferon (IFN)-γ (Figure S1). Sunitinib/erlotinib treatment of dengue-infected mice increased the levels of the innate immune response cytokines MIP1-β, MCP-1, and IL-22 as well as IL-31, a cytokine associated with cell-mediated immunity against microbial pathogens, at 48 and/or 72 hours following treatment (Figure 7). Additionally, sunitinib/erlotinib treatment significantly reduced the level of the proinflammatory cytokine IL-18 (Figures 7 and S1). These findings suggest that in addition to inhibiting intracellular viral trafficking (Bekerman et al., 2017), sunitinib/erlotinib treatment alters host cytokine responses.
Figure 7. Sunitinib/erlotinib combination alters inflammatory cytokines levels in the serum of dengue infected mice.

Cytokines levels measured by Luminex assay in AG-B6 mice 48 or 72 hours postinfection with DENV or in uninfected mice following once-daily administration of vehicle or sunitinib/erlotinib (SM/E) combination. Shown are box plots of the concentrations of the indicated cytokines. The box horizontal lines indicate the first, second (median) and third quartiles. Outliers above the upper whisker (Q3+1.5xIQR) or below the lower whisker (Q1–1.5xIQR) are labeled with black lines. n = 8 per infected treatment group and 2–3 per uninfected group treated with vehicle or sunitinib/erlotinib combination, respectively. *P < 0.05 and ***P < 0.001 relative to untreated infected mice by 2-way ANOVA with Tukey’s multiple comparisons test. IQR, interquartile range; NS, Non-Significant.
4. Discussion
We previously reported that sunitinib/erlotinib combination protects mice from DENV and EBOV infections, in part via inhibition of AAK1- and GAK-regulated intracellular viral trafficking (Bekerman et al., 2017). Nevertheless, better understanding of the therapeutic potential and MOA of this approach is warranted prior to advancing it into clinic. Here, we address these gaps and further demonstrate feasibility and biological rationale in repurposing sunitinib/erlotinib as anti-DENV drugs.
Our data indicate that the suppressive effect of 5-day treatment with sunitinib/erlotinib combination on systemic DENV replication in mice is sustainable. Treatment thus prevents progression to severe dengue rather than delays its onset. Notably, this treatment delays, but does not prevent the development of late-onset paralysis observed in this animal model. Poor BBB permeability may explain lack of viral control in the brain of infected animals (Oberoi et al., 2013; Verheijen et al., 2017). While neurological manifestations can complicate clinical dengue infection, they are primarily attributed to toxic and metabolic processes or post-infectious inflammation, and rarely to DENV replication in the brain (Solomon et al., 2000). The relevance of paralysis, commonly observed in dengue mouse models, to human infection is thus questionable (Prestwood et al., 2008; Shresta et al., 2006). Additionally, the immunosuppressed status of AG-B6 mice likely played a role in limiting viral clearance. These data therefore do not propose that drug candidates for the treatment of dengue should be BBB-permeable, but rather highlight the limitations of this model. Additionally, while treatment was administered intraperitoneally (Figures 1 and 2), we have previously reported that this combination treatment proved comparably efficacious with either intraperitoneal or oral administration (Bekerman et al., 2017).
Sunitinib and erlotinib are approved individually for the treatment of various cancers at a maximum once-daily oral dose of 37.5 mg and 150 mg, respectively. The 30 mg/kg intraperitoneal dose of sunitinib and erlotinib in mice was previously shown to yield serum drug concentrations comparable to those achieved in humans at the approved dose and is below the maximum tolerated dose in mice (Bekerman et al., 2017; Higgins et al., 2004; Mendel et al., 2003; Nair and Jacob, 2016). It was efficacious and similarly well tolerated in the dengue model (Bekerman et al., 2017). Nevertheless, per the FDA’s guidelines for drug dose conversion based on the body surface area (Nair and Jacob, 2016), 30 mg/kg sunitinib mouse dose is 3-fold higher than the approved human dose, while 30 mg/kg erlotinib dose is equivalent. Furthermore, because repurposing of approved anticancer drugs for an antiviral indication may require meeting more stringent safety criteria, demonstrating antiviral efficacy at the lowest possible dose is desirable. Our finding that oral administration of sunitinib/erlotinib combinations at 25/20 and 20/15 mg/kg are protective similarly to the previously reported 30/30 dose, suggests that finding a safe therapeutic dose lower than that approved in oncology may be feasible. It is possible that an increase in sunitinib metabolite, whose activity in other models is comparable to that of sunitinib, partially compensates for the concurrent decrease of sunitinib exposure upon erlotinib treatment (Bello et al., 2006; Blumenschein et al., 2012; Noé et al., 2016). Sunitinib/erlotinib combination has already been shown to be tolerated in ~1000 cancer patients, albeit with increased gastrointestinal adverse events (Blumenschein et al., 2012; Scagliotti et al., 2012). However, a shift from the long-term cancer treatment to acute dengue infection treatment should improve tolerance and minimize adverse effects.
Dendritic cells represent the primary target of DENV in humans. Moreover, primary cells better model human physiology and disease than immortalized cell lines (Schmid et al., 2014). Our finding that sunitinib and erlotinib treatment exhibits antiviral efficacy in MDDCs with minimal toxicity to host cells thus further supports the biological relevance of this approach.
Our single-cell transcriptomic data reveals a strong correlation between AAK1 expression and intracellular virus abundance. Since the expression distribution in infected cells shifts beyond the tails of the expression distribution in uninfected cells, it is likely that the expression change is triggered by infection rather than representing a preexisting level setting permissive conditions for infection. We have recently reported that most human genes do not correlate with the intracellular vRNA (Zanini et al., 2018). AAK1 is thus among the small subset of genes, many of which are proviral factors (Zanini et al., 2018), that correlate with virus level in the same cells. This finding further validates AAK1 as an attractive antiviral target and proposes that like in cancer (Paul and Mukhopadhyay, 2004), targeting overexpressed proviral kinases in virally-infected cells may facilitate an effective means to suppress viral infection. Moreover, these data underscore the utility of the single-cell transcriptomic approach to characterize host response to viral infection at high resolution by combining temporal information, cell-to-cell variability, and high-quality expression data. While GAK gene level slightly increased in bulk samples, it did not correlate with vRNA in single cells. This may reflect our finding that mean values measured via bulk assays are inflated and tend to over-represent highly infected cells (Zanini et al., 2018). Alternatively, our single-cell coverage (~400,000 reads/cell) may be too low to detect smaller changes in transcript level.
Our gain-of-function assays support inhibition of AAK1 and GAK activity as an important mode of antiviral action of sunitinib and erlotinib. Nevertheless, we cannot exclude the possibility that other targets of sunitinib, such as AXL, KIT and RET, which may be essential for DENV infection (Bekerman et al., 2017), are involved in mediating this antiviral effect. Beyond suppression of AAK1- and GAK-mediated intracellular viral trafficking, our data suggest that alterations in cytokine responses may also contribute to the protective effect of sunitinib/erlotinib in dengue-infected mice. Some cytokines whose level is increased by sunitinib/erlotinib treatment are implicated in protecting from severe dengue or are associated with uncomplicated dengue. Elevated MIP-1β level is associated with NK cell activation and protection from development of severe dengue in patients (Azeredo et al., 2006; Bozza et al., 2008). MCP-1 levels are higher in mice inoculated with DENV isolates obtained from patients with dengue fever relative to severe dengue (Tuiskunen et al., 2011). IL-22 is protective in a dengue mouse model (Guabiraba et al., 2013) and its polymorphism is associated with hepatitis C virus clearance (Hennig et al., 2007). The increased level of the two chemokines MIP-1β and MCP-1 (Sokol and Luster, 2015) and of IL-22 (Wolk et al., 2004) suggests that sunitinib/erlotinib treatment may augment innate immune responses. The increase in IL-31, a cytokine associated with cell-mediated immunity (Hermanns, 2015) , suggests that treatment may also affect the adaptive immune system. Sunitinib/erlotinib treatment also significantly reduced the level of the proinflammatory cytokine IL-18, whose elevated level in severe dengue is thought to contribute to the pathogenesis of dengue hemorrhagic fever (Mustafa et al., 2001; Soo et al., 2017). This is an attractive feature of sunitinib/erlotinib treatment, as it suggests they may benefit patients not only by inhibiting viral infection, but also by altering cytokine responses in a way that shifts the balance from pathogenesis to viral clearance. It remains to be determined whether inhibition of AAK1 and GAK or other kinases mediates this altered cytokine responses. Alternatively, it is possible that these drugs’ effect on cytokine production results from reduced viral burden via their effect on viral replication. Since AG-B6 mice are immunodeficient, the relevance of these cytokine alterations to human dengue infection remains to be determined.
In summary, our study provides further evidence for the utility of two clinically approved compounds as potential therapies against DENV infection and novel insight into their MOA. This repurposed approach may provide additive and possibly synergistic effects in combination with other strategies being developed to combat dengue and other emerging viral infections.
Supplementary Material
Acknowledgments.
We thank Eva Harris and Karla Kirkegaard for assistance with establishing dengue tissue culture and mouse models. This work was supported by award number 1U19AI10966201 (CETR) from the National Institute of Allergy and Infectious Diseases (NIAID) to S.E., and grants from the Stanford SPARK program. S.P. was supported by the Child Health Research Institute, Lucile Packard Foundation for Children’s Health, as well as the Stanford CTSA (grant number UL1 TR000093). The opinions, interpretations, conclusions, and recommendations are those of the authors and are not necessarily endorsed by the U.S. Army or the other funders.
References
- Azeredo EL, De Oliveira-Pinto LM, Zagne SM, Cerqueira DIS, Nogueira RMR, Kubelka CF, 2006. NK cells, displaying early activation, cytotoxicity and adhesion molecules, are associated with mild dengue disease. Clin. Exp. Immunol. 143, 345–356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bekerman E, Einav S, 2015. Infectious disease. Combating emerging viral threats. Science 348, 282–283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bekerman E, Neveu G, Shulla A, Brannan J, Pu S-Y, Wang S, Xiao F, Barouch-Bentov R, Bakken RR, Mateo R, Govero J, Nagamine CM, Diamond MS, De Jonghe S, Herdewijn P, Dye JM, Randall G, Einav S, 2017. Anticancer kinase inhibitors impair intracellular viral trafficking and exert broad-spectrum antiviral effects. The Journal of clinical investigation 127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bello CL, Sherman L, Zhou J, Verkh L, Smeraglia J, Mount J, Klamerus KJ, 2006. Effect of food on the pharmacokinetics of sunitinib malate (SU11248), a multi-targeted receptor tyrosine kinase inhibitor: results from a phase I study in healthy subjects. Anticancer Drugs 17, 353–358. [DOI] [PubMed] [Google Scholar]
- Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, Moyes CL, Drake JM, Brownstein JS, Hoen AG, Sankoh O, Myers MF, George DB, Jaenisch T, Wint GRW, Simmons CP, Scott TW, Farrar JJ, Hay SI, 2013. The global distribution and burden of dengue. Nature 496, 504–507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhattacharyya S, Hope TJ, Young JA, 2011. Differential requirements for clathrin endocytic pathway components in cellular entry by Ebola and Marburg glycoprotein pseudovirions. Virology 419, 1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blumenschein GR Jr., Ciuleanu T, Robert F, Groen HJM, Usari T, Ruiz-Garcia A, Tye L, Chao RC, Juhasz E, 2012. Sunitinib Plus Erlotinib for the Treatment of Advanced/Metastatic Non-Small-Cell Lung Cancer: A Lead-In Study. Journal of Thoracic Oncology 7, 1406–1416. [DOI] [PubMed] [Google Scholar]
- Bozza FA, Cruz OG, Zagne SMO, Azeredo EL, Nogueira RMR, Assis EF, Bozza PT, Kubelka CF, 2008. Multiplex cytokine profile from dengue patients: MIP-1beta and IFN-gamma as predictive factors for severity. BMC Infect. Dis. 8, 86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan KW, Watanabe S, Kavishna R, Alonso S, Vasudevan SG, 2015. Animal models for studying dengue pathogenesis and therapy. Antiviral Res 123, 5–14. [DOI] [PubMed] [Google Scholar]
- Costa VV, Fagundes CT, Souza DG, Teixeira MM, 2013. Inflammatory and innate immune responses in dengue infection: protection versus disease induction. Am. J. Pathol. 182, 1950–1961. [DOI] [PubMed] [Google Scholar]
- Geiss GK, Bumgarner RE, An MC, Agy MB, van ‘t Wout A.l.B., Hammersmark E, Carter VS, Upchurch D, Mullins JI, Katze MG, 2000. Large-Scale Monitoring of Host Cell Gene Expression during HIV-1 Infection Using cDNA Microarrays. Virology 266, 8–16. [DOI] [PubMed] [Google Scholar]
- Ghosh P, Kornfeld S, 2003. AP-1 binding to sorting signals and release from clathrin-coated vesicles is regulated by phosphorylation. J Cell Biol 160, 699–708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guabiraba R, Besnard A-G, Marques RE, Maillet I, Fagundes CT, Conceição TM, Rust NM, Charreau S, Paris I, Lecron J-C, Renauld J-C, Quesniaux V, Da Poian AT, Arruda LB, Souza DG, Ryffel B, Teixeira MM, 2013. IL-22 modulates IL-17A production and controls inflammation and tissue damage in experimental dengue infection. Eur. J. Immunol. 43, 1529–1544. [DOI] [PubMed] [Google Scholar]
- Halstead SB, Russell PK, 2016. Protective and immunological behavior of chimeric yellow fever dengue vaccine. Vaccine 34, 1643–1647. [DOI] [PubMed] [Google Scholar]
- Hennig BJ, Frodsham AJ, Hellier S, Knapp S, Yee LJ, Wright M, Zhang L, Thomas HC, Thursz M, Hill AV, 2007. Influence of IL-10RA and IL-22 polymorphisms on outcome of hepatitis C virus infection. Liver Int. 27, 1134–1143. [DOI] [PubMed] [Google Scholar]
- Hermanns HM, 2015. Oncostatin M and interleukin-31: Cytokines, receptors, signal transduction and physiology. Cytokine Growth Factor Rev. 26, 545–558. [DOI] [PubMed] [Google Scholar]
- Higgins B, Kolinsky K, Smith M, Beck G, Rashed M, Adames V, Linn M, Wheeldon E, Gand L, Birnboeck H, Hoffmann G, 2004. Antitumor activity of erlotinib (OSI-774, Tarceva) alone or in combination in human non-small cell lung cancer tumor xenograft models. Anti-cancer drugs 15, 503–512. [DOI] [PubMed] [Google Scholar]
- Huang CYH, Butrapet S, Moss KJ, Childers T, Erb SM, Calvert AE, Silengo SJ, Kinney RM, Blair CD, Roehrig JT, 2010. The dengue virus type 2 envelope protein fusion peptide is essential for membrane fusion. Virology 396, 305–315. [DOI] [PubMed] [Google Scholar]
- Karaman MW, Herrgard S, Treiber DK, Gallant P, Atteridge CE, Campbell BT, Chan KW, Ciceri P, Davis MI, Edeen PT, Faraoni R, Floyd M, Hunt JP, Lockhart DJ, Milanov ZV, Morrison MJ, Pallares G, Patel HK, Pritchard S, Wodicka LM, Zarrinkar PP, 2008. A quantitative analysis of kinase inhibitor selectivity. Nat Biotech 26, 127–132. [DOI] [PubMed] [Google Scholar]
- Katzelnick LC, Gresh L, Halloran ME, Mercado JC, Kuan G, Gordon A, Balmaseda A, Harris E, 2017. Antibody-dependent enhancement of severe dengue disease in humans. Science 358, 929–932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kovackova S, Chang L, Bekerman E, Neveu G, Barouch-Bentov R, Chaikuad A, Heroven C, Sala M, De Jonghe S, Knapp S, Einav S, Herdewijn P, 2015. Selective Inhibitors of Cyclin G Associated Kinase (GAK) as Anti-Hepatitis C Agents. J Med Chem 58, 3393–3410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mendel DB, Laird AD, Xin X, Louie SG, Christensen JG, Li G, Schreck RE, Abrams TJ, Ngai TJ, Lee LB, Murray LJ, Carver J, Chan E, Moss KG, Haznedar JÖ, Sukbuntherng J, Blake RA, Sun L, Tang C, Miller T, Shirazian S, McMahon G, Cherrington JM, 2003. In Vivo Antitumor Activity of SU11248, a Novel Tyrosine Kinase Inhibitor Targeting Vascular Endothelial Growth Factor and Platelet-derived Growth Factor Receptors: Determination of a Pharmacokinetic/Pharmacodynamic Relationship. Clinical Cancer Research 9, 327–337. [PubMed] [Google Scholar]
- Mustafa AS, Elbishbishi EA, Agarwal R, Chaturvedi UC, 2001. Elevated levels of interleukin-13 and IL-18 in patients with dengue hemorrhagic fever. FEMS Immunol. Med. Microbiol. 30, 229–233. [DOI] [PubMed] [Google Scholar]
- Nair AB, Jacob S, 2016. A simple practice guide for dose conversion between animals and human. Journal of basic and clinical pharmacy 7, 27–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neveu G, Barouch-Bentov R, Ziv-Av A, Gerber D, Jacob Y, Einav S, 2012. Identification and Targeting of an Interaction between a Tyrosine Motif within Hepatitis C Virus Core Protein and AP2M1 Essential for Viral Assembly. PLoS pathogens 8, e1002845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neveu G, Ziv-Av A, Barouch-Bentov R, Berkerman E, Mulholland J, Einav S, 2015. AP-2-Associated Protein Kinase 1 and Cyclin G-Associated Kinase Regulate Hepatitis C Virus Entry and Are Potential Drug Targets. Journal of virology 89, 4387–4404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noé G, Bellesoeur A, Thomas-Schoemann A, Rangarajan S, Naji F, Puszkiel A, Huillard O, Saidu N, Golmard L, Alexandre J, Goldwasser F, Blanchet B, Vidal M, 2016. Clinical and kinomic analysis identifies peripheral blood mononuclear cells as a potential pharmacodynamic biomarker in metastatic renal cell carcinoma patients treated with sunitinib. Oncotarget 7, 67507–67520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oberoi RK, Mittapalli RK, Elmquist WF, 2013. Pharmacokinetic assessment of efflux transport in sunitinib distribution to the brain. J Pharmacol Exp Ther 347, 755–764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohno H, Aguilar RC, Fournier MC, Hennecke S, Cosson P, Bonifacino JS, 1997. Interaction of endocytic signals from the HIV-1 envelope glycoprotein complex with members of the adaptor medium chain family. Virology 238, 305–315. [DOI] [PubMed] [Google Scholar]
- Paul MK, Mukhopadhyay AK, 2004. Tyrosine kinase – Role and significance in Cancer. International journal of medical sciences 1, 101–115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prestwood TR, Prigozhin DM, Sharar KL, Zellweger RM, Shresta S, 2008. A mouse-passaged dengue virus strain with reduced affinity for heparan sulfate causes severe disease in mice by establishing increased systemic viral loads. J Virol 82, 8411–8421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ricotta D, Conner SD, Schmid SL, von Figura K, Honing S, 2002. Phosphorylation of the AP2 mu subunit by AAK1 mediates high affinity binding to membrane protein sorting signals. J Cell Biol 156, 791–795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodriguez-Madoz JR, Bernal-Rubio D, Kaminski D, Boyd K, Fernandez-Sesma A, 2010. Dengue virus inhibits the production of type I interferon in primary human dendritic cells. J Virol 84, 4845–4850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scagliotti GV, Krzakowski M, Szczesna A, Strausz J, Makhson A, Reck M, Wierzbicki RF, Albert I, Thomas M, Miziara JE, Papai ZS, Karaseva N, Thongprasert S, Portulas ED, von Pawel J, Zhang K, Selaru P, Tye L, Chao RC, Govindan R, 2012. Sunitinib plus erlotinib versus placebo plus erlotinib in patients with previously treated advanced non-small-cell lung cancer: a phase III trial. J Clin Oncol 30, 2070–2078. [DOI] [PubMed] [Google Scholar]
- Schmid MA, Diamond MS, Harris E, 2014. Dendritic cells in dengue virus infection: targets of virus replication and mediators of immunity. Front Immunol 5, 647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schor S, Einav S, 2018. Combating Intracellular Pathogens with Repurposed Host-Targeted Drugs. ACS Infect Dis. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schul W, Liu W, Xu HY, Flamand M, Vasudevan SG, 2007. A dengue fever viremia model in mice shows reduction in viral replication and suppression of the inflammatory response after treatment with antiviral drugs. J Infect Dis 195, 665–674. [DOI] [PubMed] [Google Scholar]
- Shresta S, Sharar KL, Prigozhin DM, Beatty PR, Harris E, 2006. Murine model for dengue virus-induced lethal disease with increased vascular permeability. J Virol 80, 10208–10217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sokol CL, Luster AD, 2015. The Chemokine System in Innate Immunity. Cold Spring Harb. Perspect. Biol. 7, a016303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Solomon T, Dung NM, Vaughn DW, Kneen R, Thao LTT, Raengsakulrach B, Loan HT, Day NPJ, Farrar J, Myint KSA, Warrell MJ, James WS, Nisalak A, White NJ, 2000. Neurological manifestations of dengue infection. The Lancet 355, 1053–1059. [DOI] [PubMed] [Google Scholar]
- Soo K-M, Khalid B, Ching S-M, Tham CL, Basir R, Chee H-Y, 2017. Meta-analysis of biomarkers for severe dengue infections. PeerJ 5, e3589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torres JZ, Miller JJ, Jackson PK, 2009. High-throughput generation of tagged stable cell lines for proteomic analysis. Proteomics 9, 2888–2891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tuiskunen A, Wahlström M, Bergström J, Buchy P, Leparc-Goffart I, Lundkvist A, 2011. Phenotypic characterization of patient dengue virus isolates in BALB/c mice differentiates dengue fever and dengue hemorrhagic fever from dengue shock syndrome. Virol. J. 8, 398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Umeda A, Meyerholz A, Ungewickell E, 2000. Identification of the universal cofactor (auxilin 2) in clathrin coat dissociation. Eur J Cell Biol 79, 336–342. [DOI] [PubMed] [Google Scholar]
- Verheijen RB, Yaqub MM, Sawicki E, van Tellingen O, Lammertsma AA, Nuijen B, Schellens JHM, Beijnen JH, Huitema ADR, Hendrikse NH, Steeghs N, 2017. Molecular Imaging of ABCB1/ABCG2 Inhibition at the Human Blood Brain Barrier using Elacridar and (11)C-Erlotinib PET. J Nucl Med. [DOI] [PubMed] [Google Scholar]
- Walsh D, Mohr I, 2011. Viral subversion of the host protein synthesis machinery. Nature Reviews Microbiology 9, 860–875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang TT, Sewatanon J, Memoli MJ, Wrammert J, Bournazos S, Bhaumik SK, Pinsky BA, Chokephaibulkit K, Onlamoon N, Pattanapanyasat K, Taubenberger JK, Ahmed R, Ravetch JV, 2017. IgG antibodies to dengue enhanced for FcgammaRIIIA binding determine disease severity. Science 355, 395–398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolk K, Kunz S, Witte E, Friedrich M, Asadullah K, Sabat R, 2004. IL-22 increases the innate immunity of tissues. Immunity 21, 241–254. [DOI] [PubMed] [Google Scholar]
- Xie X, Gayen S, Kang C, Yuan Z, Shi P-Y, 2013. Membrane Topology and Function of Dengue Virus NS2A Protein. Journal of virology 87, 4609–4622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zanini F, Pu SY, Bekerman E, Einav S, Quake SR, 2018. Single-cell transcriptional dynamics of flavivirus infection. Elife 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang CX, Engqvist-Goldstein AE, Carreno S, Owen DJ, Smythe E, Drubin DG, 2005. Multiple roles for cyclin G-associated kinase in clathrin-mediated sorting events. Traffic 6, 1103–1113. [DOI] [PubMed] [Google Scholar]
- Zou G, Xu HY, Qing M, Wang Q-Y, Shi P-Y, 2011. Development and characterization of a stable luciferase dengue virus for high-throughput screening. Antiviral research 91, 11–19. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.