

Abstract
The underlying mechanism and cellular responses of bacteria against toxic cadmium ions is still not fully understood. Herein, Escherichia coli TG1 expressing hexahistidine-green fluorescent protein (His6GFP) and cells expressing polyhistidine-fused to the outer membrane protein A (His-OmpA) were applied as models to investigate roles of cytoplasmic metal complexation and metal chelation at the surface membrane, respectively, upon exposure to cadmium stress. Two-dimensional gel electrophoresis (2-DE) and two-dimensional difference in gel electrophoresis (2D-DIGE) in conjunction with mass spectrometry-based protein identification had successfully revealed the low level expression of antioxidative enzymes and stress-responsive proteins such as manganese-superoxide dismutase (MnSOD; +1.65 fold), alkyl hydroperoxide reductase subunit C (AhpC; +1.03 fold) and DNA starvation/stationary phase protection protein (Dps; −1.02 fold) in cells expressing His6GFP in the presence of 0.2 mM cadmium ions. By contrarily, cadmium exposure led to the up-regulation of MnSOD of up to +7.20 and +3.08 fold in TG1-carrying pUC19 control plasmid and TG1 expressing native GFP, respectively, for defensive purposes against Cd-induced oxidative cell damage. Our findings strongly support the idea that complex formation between cadmium ions and His6GFP could prevent reactive oxygen species (ROS) caused by interaction between Cd2+ and electron transport chain. This coincided with the evidence that cells expressing His6GFP could maintain their growth pattern in a similar fashion as that of the control cells even in the presence of harmful cadmium. Interestingly, overexpression of either OmpA or His-OmpA in E. coli cells has also been proven to confer protection against cadmium toxicity as comparable to that observed in cells expressing His6GFP. Blockage of metal uptake as a consequence of anchored polyhistidine residues on surface membrane limited certain amount of cadmium ions in which some portion could pass through and exert their toxic effects to cells as observed by the increased expression of MnSOD of up to +9.91 and +3.31 fold in case of TG1 expressing only OmpA and His-OmpA, respectively. Plausible mechanisms of cellular responses and protein mapping in the presence of cadmium ions were discussed. Taken together, we propose that the intracellular complexation of cadmium ions by metal-binding regions provides more efficiency to cope with cadmium stress than the blockage of metal uptake at the surface membrane. Such findings provide insights into the molecular mechanism and cellular adaptation against cadmium toxicity in bacteria.
Keywords: Cadmium stress, Polyhistidine, Proteomics, Metal complexation, Outer membrane protein
Introduction
Cadmium ion is considered to be one of the harmful heavy metals that exerts its toxicities to all organisms including human, animals and microorganisms (Vallee & Ulmer, 1972). In microbes, the most common toxicity includes mutagenic effect, growth inhibition, physiological alterations and inhibition of enzymatic activities in metabolic pathway (Bischoff, 1982). Most of the microorganisms attempt to adapt themselves naturally from these metals by utilizing several cellular responses, e.g., reduction of metal-uptake process, sequestration of metal ions by biological macromolecules, transportation of metal ions out by efflux system, transformation of toxic metals by reductase enzymes, and metal sorption by biofilm and siderophores (Giovanella et al., 2017; Nies, 1999). Among these, active efflux has been thought to be the primary mechanism developed in prokaryotes to reduce intracellular cadmium ions. By contrast, intracellular complexation of the toxic cadmium ion by cadmium-binding components such as metallothioneins and phytochelatins is mainly used in eukaryotes (Nies, 1992).
In case of well-known bacteria e.g., Escherichia coli (E. coli), research findings during the past 50 years stated the toxic effects of cadmium ions and significant cellular responses and adaptation in various aspects as follows. (i) Cadmium toxicity: Cd2+ is readily taken up by E. coli cells by means of an active transport system with a Km of approximately 2.1 µM (Laddaga & Silver, 1985). Exposure to cadmium ion results in long lag phase of cell proliferation, reduction of cell survival and loss of their ability to form colony (Mitra, 1984; Mitra & Bernstein, 1978; Mitra et al., 1975). Explanations for those phenomena could be accounted as the breakage of single-strand DNA, inhibition of metalloenzyme (i.e., alkaline phosphatase) and aggregation of cytoplasmic materials (Mitra & Bernstein, 1978; Mitra et al., 1975). Other toxic effects including disturbation of the cell division as a consequence of improper Z-ring formation, metabolic dysfunction, and imbalance of oxidative pressure leading to growth retardation were reported (Hossain, Mallick & Mukherjee, 2012). Induction of oxidative damage by superoxide radical together with inability to express protective enzymes and to replace damaged proteins by de novo protein synthesis have currently been reported to be the main reason for growth stasis and cell death in Cd poisoning (Thomas & Benov, 2018). (ii) Cellular responses and adaptation: the efflux system not been widely accepted as the primary defense mechanism of E. coli to reduce the toxic effects of cadmium ions. Several reports stated that E. coli protected themselves against harmful cadmium mainly by synthesis of cytoplasmic cadmium-binding proteins to accumulate free Cd2+ intracellularly (Cohen, Bitan & Nitzan, 1991; Khazaeli & Mitra, 1981; Mitra, 1984). Such an accumulation was proposed to control the internal environment and most likely to resume normal metabolic function after the long lag phase (Khazaeli & Mitra, 1981). Another group of proteins, namely cadmium-induced proteins (CDPs), was also produced as a stress-responsive protein during cadmium exposure (Ferianc, Farewell & Nyström, 1998). Among the CDPs, YodA protein (a protein containing histidine-rich N-terminal sequence; HGHHSH) has been extensively studied. It was found that the YodA protein mainly localized in both cytoplasm and periplasm but was only tranlocated into a periplasmic space during cadmium stress (Puskárová et al., 2002). It is believed that the role of YodA protein might be to decrease the intracellular concentration of cadmium ions by its ability to bind heavy metal (Stojnev et al., 2007). Moreover, the YodA protein is hypothesized to be part of a cadmium-exporting transport protein due to its sequence similarity to the C-terminal domain of a metal-binding receptor of a member of bacterial ATP-binding cassette transporters (David et al., 2003). In recent studies, the YodA has also been recognized as the ZinT, a metal-binding protein involved in zinc homeostasis (Kershaw, Brown & Hobman, 2007). Such an involvement of the histidine-rich peptide in the reduction of cadmium toxicity as well as in cadmium translocation strongly supports a significant role of histidine residue in metal regulation in bacterial cells. It seems that E. coli naturally utilizes both the intracellular complexation and metal transporting system to cope with cadmium stress but the proportion of which is not well understood. Currently, regulation of the CDPs together with the other general stress responses was subsequently studied either at the transcriptional or translational levels by gene-array technology or two-dimensional gel electrophoresis (Brocklehurst & Morby, 2000; Ferianc, Farewell & Nyström, 1998; Isarankura-Na-Ayudhya et al., 2009; Lausova, Ferianc & Polek, 1999). Results revealed that some regulatory systems, e.g., OxyR, SoxRS, RpoH, UspA, GrpE, RecA, and YodA, participated in the cadmium stress (Ferianc, Farewell & Nyström, 1998; LaRossa, Smulski & Van Dyk, 1995; Lim et al., 2009; Puskárová et al., 2002; Shapiro & Keasling, 1996; Thomas & Benov, 2018; Van Dyk et al., 1995). These inducible regulators can be classified in various groups as general stress regulons (UspA, YodA), SOS regulons (RecA), oxidative stress regulons (OxyR, SoxRS), heat shock protein regulons (RpoH) and chaperones (GrpE) (Han & Lee, 2006). It can be speculated that there will be some interconnection between these regulons in the cadmium stress.
In parallel to those long-term studies, genetic engineering has extensively been applied to construct metal-binding proteins in bacteria and other organisms. The most popular target belongs to a group of cysteine-rich peptides known as metallothionein (MT) that is ubiquitously found in various organisms. The expression of MTs had successfully conferred metal toleration and/or intracellular metal accumulation against cadmium stress in many cases (He et al., 2014; Hou, Kim & Kim, 1988; Kille et al., 1990; Romeyer et al., 1988). The thiol group of cysteine residues acts as soft Lewis base that preferably binds to cadmium ions (served as soft Lewis acid). However, the difficulty of controlling their reduced states limits their maximum binding capability to cadmium ions (Mahnam et al., 2018). Moreover, the overexpression of proteins containing thiol-moiety is reported to alter the redox-regulating system and growth characteristics of E. coli (Kondo et al., 2000). Therefore, histidine residue (another amino acid known to coordinate with divalent ions) has widely been selected for cadmium binding and bioremediation purposes (Nair & Robinson, 2001; Patel et al., 2010; Samuelson et al., 2000; Sousa, Cebolla & De Lorenzo, 1996). The presence of histidine-rich peptides had conferred the engineered E. coli the capability to accumulate higher amounts of cadmium ions (∼11 fold) than that of the control (Sousa, Cebolla & De Lorenzo, 1996). Such findings explore the efficiency of applying metal-binding amino acids/peptides as tools not only to seek for their potential usages in bioremediation but also to study the molecular mechanism of metal toleration.
Herein, a proteomics profiling of E. coli TG1 upon exposure to sub-lethal dose of cadmium ions has been analyzed by using Two-dimensional gel electrophoresis (2-DE) and two-dimensional difference in gel electrophoresis (2D-DIGE) in conjunction with mass spectrometry-based protein identification. Engineered E. coli cells expressing metal-binding regions on the outer membrane or cytoplasm have been utilized in order to test whether controlling of metal transport across the membrane or intracellular metal complexation played major roles in modulating cellular adaptations against toxic cadmium ions. In Particular, the expression of green fluorescent protein carrying hexahistidine (His6GFP) or polyhistidine fused to outer membrane protein A (His-OmpA) have been used as models to mimic metal-binding regions located in the cytoplasm and on the surface membrane, respectively. It has previously been reported that the expression of chimeric His6GFP could modulate metal homeostasis and mobility inside E. coli cells (Isarankura-Na-Ayudhya et al., 2005). The rationale behind the expression of histidine-rich peptide in the cytoplasm is expected to chelate cadmium ions in order to prevent the sequelae of cadmium-induced oxidative cell damage via reactive oxygen species. The expression of His-OmpA has also been found to provide high binding affinity for metal ions at the surface of E. coli (Isarankura-Na-Ayudhya et al., 2005). The presence of polyhistidine on the cell surface is expected to trap cadmium ions and limit certain amount of cadmium to transport across the membrane. Then, the investigation of differentially-expressed proteins between these engineered E. coli and control cells has been conducted in order to gain a better understanding of the alternative ways that bacteria acclimatize themselves in toxic environments as compared to those observed in nature.
Materials and Methods
Bacterial strain and plasmids
E. coli strain TG1 (supE, hsdΔ5, thiΔ(lac-proAB), F’[traD36 proAB+ lacIq lacZΔM15]) was used as host cell. To express cytoplasmic metal-binding motifs, a plasmid designated as pHis6GFPuv (Isarankura-Na-Ayudhya et al., 2005) that encodes a chimeric green fluorescent protein carrying hexahistidine was transformed and further expressed in host cells. Cells harboring pUC19 and pGFPuv (Clontech Laboratories, Mountain View, CA, USA) were used as controls. In parallel, a plasmid namely pEVZn (Mejare, Ljung & Bulow, 1998) that codes for the outer membrane protein A-polyhistidine fusion protein was expressed in order to generate cells expressing surface metal-binding motifs. For comparison, cells harboring pEV208 (Mejare, Ljung & Bulow, 1998) was used to over-express the outer membrane protein.
Growth patterns of cells in the presence of sub-lethal dose of cadmium ions
All of the aforementioned cells with the exception of E. coli host were grown in 5 ml Luria-Bertani (LB) broth (10 g/L tryptone, 5 g/L NaCl and 5 g/L yeast extract, pH 7.2) that is supplemented with 100 mg/L ampicillin at 37 °C, 150 rpm for overnight. The cultivation of the TG1 host was performed in a similar manner but without the addition of ampicillin. Then, 50 µl of overnight cultures were transferred into 5 ml broth and grown until OD600 reached 0.5. Cells were adjusted to equal optical density at 0.05 in 5 ml LB broth supplemented with 100 mg/L ampicillin. Dose response assay was performed by addition of cadmium chloride stock solution to yield final concentrations of 0–1.0 mM. Cultures were further incubated at 37 °C, 150 rpm for 12 h. The growth rate was determined by monitoring the absorbance at 600 nm by spectrophotometer. It is noteworthy that the sub-lethal dose of 0.2 mM cadmium ions (exhibiting ∼50% growth inhibition of TG1 host) was selected as an effective dose for further experiments.
Two-dimensional gel electrophoresis (2-DE) of crude protein extracts
Overnight cultures of cells in 5 ml LB or LB/Amp were adjusted to OD600 of 1.0. One milliliter of cell suspension was inoculated into 50 ml LB or LB/Amp. Cells were grown at 37 °C, 150 rpm for 3 h prior to addition of CdCl2 to yield the final concentration of 0.2 mM. Cultivation was continued at 37 °C, 150 rpm for 15 h. Cells were collected by centrifugation at 4 °C, 6,000 rpm for 10 min. Crude protein extracts were prepared as previously described (Isarankura-Na-Ayudhya et al., 2010). The protein concentration was measured by Bradford’s method (Bio-Rad protein assay; Bio-Rad Laboratories, Hercules, CA, USA). These protein extracts were subjected to two-dimensional gel electrophoresis as follows. Three hundred micrograms of protein extract was mixed thoroughly with 250 µl of rehydration buffer (8 M urea, 4% CHAPS, 2 mM TBP, 0.001% bromphenol blue, 2.8 mg/ml dithiothreitol and 12 µl/ml destreak) containing 1% 3–10 IPG buffer and stored at room temperature for 10 min. Removal of insoluble material was further performed by spinning at 13,000 rpm for 10 min at 20 °C. The 13-cm IPG strips (pH range of 3–10) were placed in the IPGphor strip holder gel-side-down in rehydration solution containing sample proteins. These strips were then covered with mineral oil. Samples were run through steps of strip rehydration (30 V, 12 h) and isoelectric focusing (500 V for 1 h, 1,000 V for 1 h, and 8,000 V to reach 16,000 V h). The maximum current was maintained at 50 mA per strip. Once complete, the strip was equilibrated for two times (15 min each) in equilibration buffer (50 mM Tris pH 8.8, 6 M urea, 30% glycerol, 2% SDS, 0.03% bromphenol blue) supplemented with 65 mM DTT and 135 mM iodoacetamide. The separation of protein in the second dimension was performed using Hoefer™ DALT on 12.5% SDS-polyacrylamide gels. Proteins were separated under applied voltage of 250 V, 10 mA per strip at 20 °C for 30 min following by 250 V, 20 mA per strip at 20 °C for 2.5 h until the bromphenol blue dye front reached 0.5 cm from the bottom of the gel. Gels were further stained with colloidal Coomassie brilliant blue G for overnight. Excess dye was removed by rinsing several times with deionized distilled water. Gels were scanned with the Canoscan LiDE20 scanner (Canon, Huntington, NY, USA). Intensity of protein spots was analyzed by the ImageJ software tool (Schneider, Rasband & Eliceiri, 2012).
Protein labeling and Two-Dimensional Difference in Gel Electrophoresis (2D-DIGE)
Sample preparation
Protein extracts were cleaned-up by using the 2-D Clean-up kit (GE Healthcare, Chicago, IL, USA). Then, protein pellets were dissolved in a buffer containing 30 mM Tris–HCl, 7 M urea, 2 M thiourea, 4% (w/v) CHAPS, pH 8.5. Proteins were subsequently labeled with CyDye using GE Cydye DIGE Fluor (minimal dyes) Labeling Kit (GE Healthcare, Chicago, IL, USA) according to the manufacturer’s instruction. Then, 50 µg of protein from each sample was pooled together as the internal standard. In parallel, a total of 50 µg protein of each sample was randomly labeled with 400 pmol Cy3 or Cy5, and the internal standard was labeled with Cy2 for 30 min on ice. Samples were then rehydrated into 18 cm IPG strips (pH 3–10 NL) (GE Healthcare, Chicago, IL, USA) overnight in rehydration buffer (8 M urea, 4% CHAPS, 13 mM dithiothreitol, and 1% IPG buffer 3–10 NL). Isoelectric focusing was carried out using an IPGphor III apparatus (GE Healthcare, Chicago, IL, USA) according to the following procedures: 500 V for 500 V h, 1,000 V for 800 V h, and 10,000 V to reach 36,000 V h. Strips were further equilibrated for 15 min in equilibration buffer (75 mM Tris–HCl pH 8.8, 6 M urea, 30% glycerol, 2% SDS, 0.002% bromphenol blue) containing 1% DTT and then for 15 min with 2.5% iodoacetamide. Equilibrated IPG strips were transferred onto 12.5% SDS-polyacrylamide gels that had been pre-casted in low fluorescence glass plates. The second dimension separation was then carried out at 10 W/gel using Ettan Dalt six electrophoresis system (GE Healthcare, Chicago, IL , USA).
Gel image analysis
The 2D-DIGE gels were visualized using a Typhoon TRIO fluorescence scanner (GE Healthcare, Chicago, IL, USA) with excitation/emission at 532/580 nm (Cy3), 633/670 nm (Cy5) and 488/520 nm (Cy2). Scanning resolution used was 100 µm with photomultiplier of 550 V. Gel image and statistical analyses were performed using the DeCyder™ 2D Differential Analysis Software (DeCyder 2D version 7.2; DeCyder, Amersham, UK) by differential in-gel analysis (DIA) and biological variation analysis (BVA). Gels were subjected to silver staining for spot visualization and picking.
Peptide mass fingerprinting (PMF) analysis
Peptide mass fingerprinting (PMF) analysis of the protein separated by 2-DE was performed as previously described (Isarankura-Na-Ayudhya et al., 2010). The experiment was initiated by cutting spots of protein from gels and these spots were further transferred to 96-well microtitre plate and then soaked in 50% methanol and 5% acetic acid for overnight. In-gel digestion of protein was performed by the addition of sequencing grade of modified trypsin (Promega, Southampton, UK). Digested peptide fragments were extracted from gel segments on a Spot Handling Workstation (GE Healthcare, Chicago, IL, USA) using preset protocols from the manufacturer. Protein identification was further carried out using MALDI-TOF mass spectrometer (Model ReflexIV; Bruker Daltonics, Billerica, MA, USA) based on peptide fingerprint map. The tryptic digested peptide was mixed with a solution of 10 mg/ml α-cyano-4-hydroxycinnamic acid (LaserBio Labs, Sophia-Antipolis Cedex, France) in 66% acetonitrile and 0.1% trifluoroacetic acid (TFA) and further spotted onto a 96-well target plate. The acquisition of mass spectra was conducted in the positive ion reflector delayed extraction mode using approximately 200 laser shots. Creation of peak lists was performed using the XMASS software (Bruker Daltonics, FRG). These peaks were then queried using the MASCOT search engine (MatrixScience, http://www.matrixscience.com/) for protein identification. The reference database used in this study was NCBInr 20130802 (31350673 sequences; 10834990394 residues). The searching criteria were as follows: complete carbamidomethylation of cysteine and partial methionine oxidation; an initial mass tolerance of ±1.2 Da; the number of missed cleavage sites of up to 1. The accuracy of the experimental to theoretical pI as well as the molecular weight of proteins was carefully taken into consideration.
Protein network analysis
Identified proteins were queried using the STRING software version 9.05 (http://string-db.org/) in order to create functional protein association networks. This database weights and integrates direct (physical) and indirect (functional) associations from various sources, e.g., genetic context, high-throughput experiments, co-expression and previous knowledge.
Statistical analysis
For the 2D-DIGE gel analysis, only the spots with changes having an abundance ratio of 1.5 fold and P values < 0.05 (Student’s t-test) were marked and selected for further protein identification. For the PMF analysis, a search result score greater than 71 was considered to be of significant difference (p < 0.05).
Results
Expression of green fluorescent protein of E. coli expressing cytoplasmic histidine-rich protein (chimeric His6GFP)
E. coli TG1 was applied as a host for the transformation of pUC19 (control plasmid), pGFPuv and pHis6GFPuv. Expression of the green fluorescent protein was confirmed by observing the bright greenish fluorescence of colonies of E. coli expressing native GFP or His6GFP under illumination with UV light (Figs. 1C and 1D). Meanwhile, no fluorescence was detected for TG1 host and TG1 carrying pUC19 (Figs. 1A and 1B).
Figure 1. Expression of GFP in tested cells as deduced from UV illumination.

Non-fluorescent colonies of TG1 host (A) and TG1 carrying pUC19 (B) as well as greenish fluorescent colonies of E. coli expressing native GFP (C) or His6GFP (D).
Effect of cadmium stress on E. coli expressing cytoplasmic histidine-rich protein (chimeric His6GFP)
In the absence of cadmium ions, it seems that the TG1 carrying a control plasmid (Fig. 2A) growed up in the LB broth faster than the others. A long lag phase (∼3.5–4 h) could be detected in the cells expressing native GFP (Fig. 2B). The growth pattern of cells expressing His6GFP (Fig. 2C) resembled those in between the TG1 carrying pUC19 (Fig. 2A) and the native GFP-expressing cells (Fig. 2B).
Figure 2. Effect of cadmium stress on E. coli expressing chimeric His6GFPuv, cells expressing native GFP and cells carrying control plasmid (pUC19).

Growth patterns of TG1/pUC19 (A), TG1/pGFPuv (B) and TG1/pHis6GFPuv (C) in the absence (opened symbol) or presence (closed symbol) of 0.2 mM cadmium ions.
In the presence of effective dose of cadmium, a growth arrest could be observed in cells expressing native GFP (Fig. 2B). However, this suppressing effect was more pronounced in cells carrying pUC19 (Fig. 2A). Interestingly, cells expressing the hexahistidine-GFP fusion protein displayed similar growth rates in the presence and absence of cadmium ions (Fig. 2C).
Proteomics profiling of E. coli expressing cytoplasmic histidine-rich protein (chimeric His6GFP)
Experimentation was initiated via the preparation of a reference map of the E. coli host proteome (Fig. 3). Approximately 200–250 spots of protein could be detected using a 13-cm IPG strip with pH range of 3–10. Some of these spots were picked up for further identification via peptide mass fingerprinting (http://www.matrixscience.com) as shown in Table 1. Most of the identified proteins pertained to energy metabolism, chaperones, heat shock proteins or stress proteins, transporters, protein synthesis machinery, and outer membrane proteins.
Figure 3. Reference map representing protein profiles of E. coli TG1.
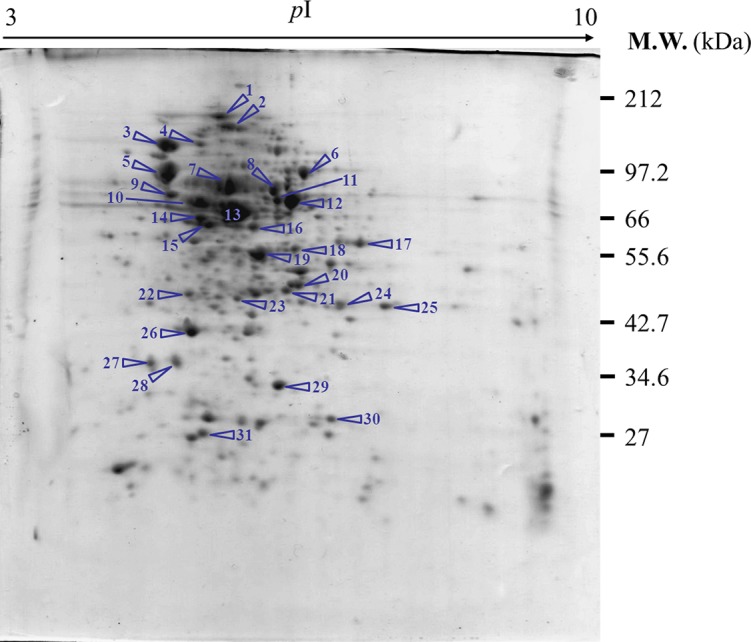
Numbers of protein spot represents identified proteins as presented in Table 1.
Table 1. Proteins of Escherichia coli TG1 host identified by mass spectrometry and peptide mass fingerprinting (PMF) analysis.
| Spot no. | Accession no. | Description | CalculatedpI value | Norminal mass(Mr) | Protein score | Sequence coverage(%) |
|---|---|---|---|---|---|---|
| 1 | gi∣485870954 | Aconitate hydratase 2 | 5.22 | 91,065 | 80 | 18 |
| 2 | gi∣486356887 | Chaperone ClpB | 5.37 | 95,712 | 76 | 31 |
| 3 | gi∣446438287 | Molecular chaperone DnaK | 4.83 | 69,096 | 115 | 46 |
| 4 | gi∣518680918 | Heat shock protein 90 | 5.09 | 71,360 | 84 | 27 |
| 5 | gi∣446651775 | Molecular chaperone GroEL | 4.85 | 57,494 | 89 | 47 |
| 6 | gi∣486650025 | Periplasmic oligopeptide-binding protein | 6.06 | 62,730 | 114 | 49 |
| 7 | gi∣485671395 | Glycerol kinase | 5.97 | 59,776 | 109 | 48 |
| 8 | gi∣1311039 | Chain A, Dipeptide binding protein | 5.75 | 57,599 | 79 | 35 |
| 9 | gi∣485778118 | ATP synthase F1, beta subunit | 4.90 | 50,341 | 125 | 41 |
| 10 | gi∣33383669 | Isocitrate dehydrogenase | 5.33 | 43,192 | 84 | 35 |
| 11 | gi∣485734442 | Tryptophanase | 5.80 | 40,109 | 81 | 54 |
| 12 | gi∣485734442 | Tryptophanase | 5.80 | 40,109 | 108 | 72 |
| 13 | gi∣445923422 | Elongation factor-Tu | 5.07 | 41,052 | 89 | 44 |
| 14 | gi∣485752584 | Phosphoglycerate kinase | 5.02 | 40,719 | 104 | 59 |
| 15 | gi∣485747310 | Glycerophosphodiester phosphodiesterase | 5.15 | 36,482 | 79 | 51 |
| 16 | gi∣485696065 | Fructose-bisphosphate aldolase | 5.52 | 39,337 | 88 | 41 |
| 17 | gi∣485665932 | Glyceraldehyde-3-phosphate dehydrogenase | 6.33 | 36,204 | 71 | 43 |
| 18 | gi∣9507742 | Outer membrane protein P | 5.91 | 35,477 | 88 | 42 |
| 19 | gi∣485723954 | Malate dehydrogenase | 5.62 | 32,532 | 72 | 48 |
| 20 | gi∣510898944 | D-ribose-binding periplasmic protein | 6.85 | 30,949 | 72 | 50 |
| 21 | gi∣486435677 | Uridine phosphorylase | 5.71 | 27,262 | 98 | 68 |
| 22 | gi∣485953066 | Translation elongation factor-Ts | 5.18 | 28,505 | 74 | 39 |
| 23 | gi∣487374764 | Purine nucleoside phosphorylase | 5.42 | 26,157 | 78 | 34 |
| 24 | gi∣485673269 | Glyceraldehyde-3-phosphate dehydrogenase | 6.32 | 31,252 | 74 | 36 |
| 25 | gi∣447012380 | Amino acid ABC transporter substrate-binding protein | 7.74 | 27,159 | 73 | 42 |
| 26 | gi∣170172436 | Alkyl hydroperoxide reductase C22 protein | 4.92 | 19,250 | 92 | 56 |
| 27 | gi∣487464714 | Glucose-specific phosphotransferase enzyme | 4.73 | 18,198 | 87 | 43 |
| 28 | gi∣446006533 | Lipid hydroperoxide peroxidase | 4.75 | 17,952 | 76 | 61 |
| 29 | gi∣485651470 | DNA starvation/stationary phase protection protein Dps | 5.72 | 18,711 | 85 | 69 |
| 30 | gi∣486164894 | 50S ribosomal protein L9 | 6.17 | 15,772 | 78 | 48 |
| 31 | gi∣446245719 | Universal stress protein A | 5.11 | 16,086 | 71 | 70 |
Further experiments were conducted to address the functional roles of cytoplasmic histidine-rich protein (His6GFP) on cellular adaptation against toxic cadmium ions. Therefore, protein expression profiles of E. coli expressing His6GFP grown in the absence or presence of 0.2 mM CdCl2 (Fig. 4C) were compared with cells harboring pUC19 and cells expressing native GFP (as represented in Figs. 4A–4B, respectively). The effects of cadmium ions on the expression of various proteins were scrutinized as follows. Aconitase, an enzyme implicated in the Tricarboxylic acid (TCA) cycle, was down-regulated and disappeared in cells expressing His6GFP. Translation elongation factor-Ts was up-regulated in all cases (at 2.12, 1.84 and 1.35 fold for TG1/pUC19, TG1/pGFPuv and TG1/pHis6GFPuv, respectively) (Table 2). Glycerol kinase, an enzyme involved in glycerol uptake and lipolysis, was up-regulated in cells expressing His6GFP (+1.92 fold) and native GFP (+1.73 fold). Oligopeptide-binding protein was down-regulated at −1.65- and −1.34 fold for control cells and cells expressing His6GFP while up-regulation was found in the case of GFP (+1.37 fold). The H+-transporting ATPase was up-regulated at 2.69 and 1.30 fold in case of control cells and cells expressing His6GFP, respectively. More importantly, the presence of cytoplasmic histidine residue due to the expression of His6GFP was found to play imperative roles in stress defense mechanisms. Manganese-superoxide dismutase (MnSOD), an oxidative scavenging enzyme, was found to be down-regulated in cells expressing His6GFP (−1.65 fold). Alkyl hydroperoxide reductase, a thiol-specific antioxidant protein, was up-regulated in the case of native GFP-expressing cells (+1.71 fold). This is in contrast to those observed in the case of His6GFP (−1.10 fold). For membrane proteins, our results revealed the down-regulation of OmpC precursor (−1.15, −1.31 and −1.56 fold) and OmpF (−1.18 fold and absence) in all cases in response to cadmium ions.
Figure 4. Protein expression profiles of E. coli grown in the presence of 0.2 mM cadmium ions.
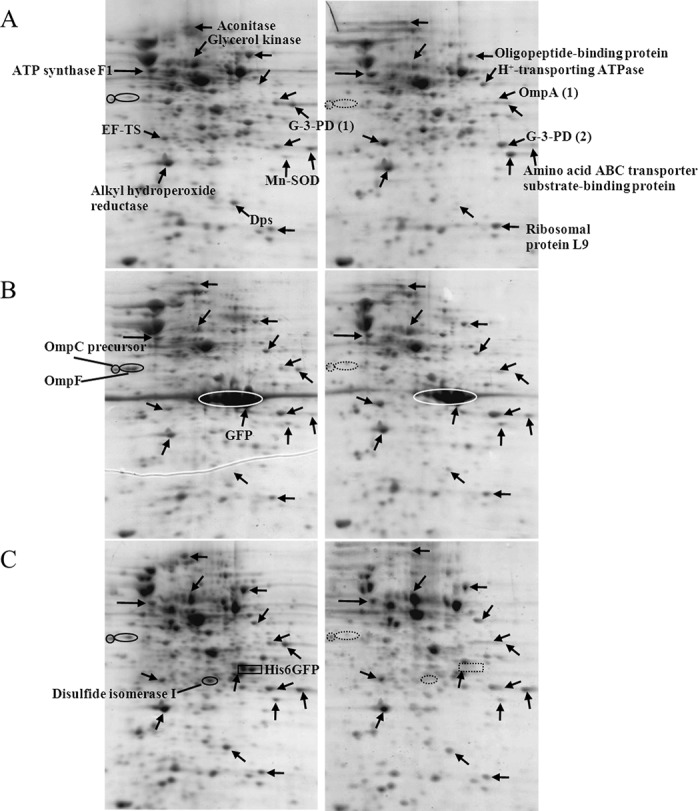
TG1/pUC19 (A), TG1/pGFPuv (B) and TG1/pHis6GFPuv (C).
Table 2. Changes of differentially expressed proteins of E. coli TG1 following exposure to cadmium.
| Protein | Up(+)/Down(−) regulation (fold) | ||||
|---|---|---|---|---|---|
| TG1 carrying pUC19 plasmid | TG1 expressing GFP | TG1 expressing His6GFP | TG1expressing OmpA | TG1 expressing His-OmpA | |
| Energy metabolism | |||||
| Glycolysis | |||||
| Glyceraldehyde-3-phosphate dehydrogenase (1) | −2.02 | −1.84 | Absence | +2.69 | Absence |
| Glyceraldehyde-3-phosphate dehydrogenase (2) | +1.75 | +1.65 | −1.06 | −2.29 | +1.63 |
| Tricarboxylic acid cycle | |||||
| Aconitase | −1.20 | No change | Absence | No change | Absence |
| Glycerol uptake and lipolysis | |||||
| Glycerol kinase | −1.60 | +1.73 | +1.92 | +1.43 | +1.33 |
| Protein biosynthesis machinery | |||||
| Translation elongation factor-Ts | +2.12 | +1.84 | +1.35 | −0.39 | +1.37 |
| Transporters | |||||
| Oligopeptide-binding protein | −1.65 | +1.37 | −1.34 | +1.22 | +1.21 |
| Amino acid ABC transporter substrate-binding protein | −1.42 | +1.37 | +1.29 | Not detectable | +1.24 |
| H+-transporting ATPase | +2.69 | −1.09 | +1.30 | −2.64 | No change |
| ATP synthase F1, beta subunit | +1.32 | +1.26 | +1.08 | +1.86 | −1.44 |
| Stress defense mechanism | |||||
| MnSOD | +2.20 | +1.29 | −1.65 | Not detectable | +2.08 |
| Alkyl hydroperoxide reductase | −1.15 | +1.71 | −1.10 | +1.52 | −1.01 |
| DNA starvation/stationary phase protection protein; Dps | −3.29 | Low amount | −1.35 | Low amount | −1.05 |
| Others | |||||
| Ribosomal protein L9 | +2.08 | +1.33 | −1.29 | +1.42 | +1.57 |
| GFP | Not detectable | −1.45 | Not detectable | Not detectable | Not detectable |
| His6GFP | Not detectable | Not detectable | −2.24 | Not detectable | Not detectable |
| Disulfide isomerase I | −1.71 | Not determined | −1.85 | Not determined | −1.36 |
| OmpA (1) | −1.68 | −1.26 | −1.40 | −1.35 | Absence |
| OmpC precursor | −1.15 | −1.31 | −1.56 | +1.32 | +2.11 |
| OmpF | −1.18 | Absence | Absence | No change | Absence |
Functional roles of histidine residue fused with outer membrane protein in E. coli
It has previously been proven that the presence of polyhistidine on the surface membrane of such engineered E. coli provided high binding affinity to zinc ions of up to 60 fold (Isarankura-Na-Ayudhya et al., 2005). Therefore, these cells were applied in the current study in order to reduce the cell permeability of cadmium ions. Figure 5 demonstrates the growth patterns of TG1/pEV208 and TG1/pEVZn in the presence and absence of cadmium ions. Our results revealed that the overexpression of the outer membrane protein of cells carrying plasmid pEV208 could protect the cell growth from the hazardous effect of cadmium ions. More importantly, the expression of polyhistidine-OmpA of cells harboring pEVZn could mediate a little bit higher degree of cell division than that of cells expressing the OmpA alone. Results from the proteomics analysis revealed that the enzyme glyceraldehyde-3-phosphate dehydrogenase form 1 was up-regulated in cells expressing OmpA (+2.69 fold) while the disapperance of this enzyme was observed in the case of cells expressing His-OmpA (Fig. 6 and Table 2). A diverse phenomenon was detected on the regulation of glyceraldehyde-3-phosphate dehydrogenase form 2 at −2.29 and +1.63 fold for TG1/OmpA and TG1/His-OmpA, respectively. Furthermore, H+-transporting ATPase was down-regulated in the case of the control (−2.64 fold) while no change was found in cells expressing His-OmpA. In addition, cadmium ions stimulated expression of MnSOD in cells expressing His-OmpA (+2.08 fold). It is noteworthy that an increased amount of the OmpC precursor was observed in cells expressing His-OmpA (+2.11 fold) as well as in cells expressing OmpA alone (+1.32 fold). Such a finding is in contrast to those observed in the case of cells expressing His6GFP and others as abovementioned.
Figure 5. Growth patterns of E. coli grown in the presence of 0.2 mM cadmium ions.

TG1/pEV208 (square) and TG1/pEVZn (diamond) in the absence (opened symbol) or presence (closed symbol) of 0.2 mM cadmium ions.
Figure 6. Protein expression profiles of E. coli grown in the presence of 0.2 mM cadmium ions.

TG1/pEV208 (A) and TG1/pEVZn (B).
Comparison of cadmium-induced protein alterations in engineered E. coli expressing cytoplasmic polyhistidine and polyhistidine- anchored surface membrane
Differentially expressed proteins of engineered E. coli upon exposure to cadmium ions were quantitatively identified as summarized in Table 2. For enzymes implicated in energy metabolism, glyceraldehyde-3-phosphate dehydrogenase form 1 was not detected in all cases except in cells expressing OmpA. The up-regulation of glyceraldehyde-3-phosphate dehydrogenase form 2 was found in TG1 carrying pUC19, cells expressing native GFP and cells expressing His-OmpA at 1.75, 1.65 and 1.63 fold, respectively. Aconitase was down-regulated in most cases. Meanwhile, the up-regulation of glycerol kinase was observed in the range of 1.3–1.9 fold in all cases except for the control. For the protein synthesis machinery, all cell types (with the exception of TG1 expressing OmpA) displayed approximately 1.3–2.1 fold increases. For transporters, oligopeptide-binding protein was decreased ∼1.3–1.6 fold only in the case of cells expressing His6GFP and control cells. Increased amounts of amino acid ABC transporter substrate-binding protein of 1.2–1.3 fold were found in TG1 expressing native GFP, His6GFP and His-OmpA while no change was observed in control cells. In regard to the H+-transporting ATPase, overexpressing of up to 2.7 fold was found in control cells. Up-regulation of approximately 1.9 fold of ATP synthase F1 (membrane-bound ATP synthase) was notified in cells expressing OmpA. For the stress defense mechanism, MnSOD is detected to be the most important antioxidative enzyme responsible for the toxic cadmium. Most cells enhanced the production of MnSOD in the range of 1.29–2.20 fold. Importantly, down-regulation of MnSOD (∼1.65 fold) was found only in cells expressing cytoplasmic polyhistidine. It should be noted that the up-regulation of AphC of up to 1.71 and 1.52 was found in TG1 expressing native GFP and cells expressing only outer membrane protein. For the ribosomal protein L9, up-regulation in the range of 1.3–2.1 fold was found in all cases with the exception of TG1 expressing His6GFP. Moreover, an increased expression of the OmpC precursor of approximately 1.3–2.1 fold as detected in cells expressing OmpA and His-OmpA. The disappearance of OmpF was observed in TG1 expressing native GFP, His6GFP and His-OmpA.
Confirmation of cadmium-induced protein alterations using two-dimensional difference in gel electrophoresis (2D-DIGE)
To further confirm the protective roles of metal complexation by histidine-rich peptides against cadmium stress, quantitative proteomics investigation using 2D-DIGE was employed as to provide high sensitivity and high discrimination power on protein expression. As shown in Fig. 7A, three different samples were individually labeled with three fluorescent dyes (Cy2, Cy3 and Cy5) and co-separated in one gel, which mediates spot matching and quantitation in a simpler and more accurate manner. An internal standard, as derived from a mixture of all samples pooled together and labeled with Cy2, was created to facilitate the normalization of each spot among all gels. This provides a reliable and reproducible detection by minimizing gel-to-gel variation and increasing experimental efficiency (Westermeier & Scheibe, 2008). Up- or down-regulation of proteins was compared via the detection of fluorescence intensities of Cy3 and Cy5 simultaneously at different wavelengths. Different expression of three key proteins (MnSOD, AhpC and Dps) involved in the stress defense mechanism was selected for further investigation (shown in Fig. 7B). Intracellular complexation of cadmium ions by His6GFP rendered cells to express minute amount of MnSOD (+1.65 fold) as compared to those of TG1 carrying pUC19 (+7.20 fold) and cells expressing native GFP (+3.08 fold). A similar observation was found with regards to the compensation of MnSOD expression by the His-OmpA (+3.31 fold) as compared to cells expressing only OmpA (+9.91 fold). The same phenomenon on the expression of AhpC in cells expressing native GFP and cells expressing OmpA was observed (Fig. 7B and Table 2). The high discriminative power of DIGE could quantify the up-regulation of Dps protein of up to 4.62, 1.82 and three fold in TG1 expressing native GFP, OmpA and His-OmpA, respectively. Moreover, the quantification of aconitase expression was obtained in a tangible manner. Down-regulation of aconitase of approximately 2.60 fold was detected in the case of TG1 expressing His-OmpA upon exposure to cadmium ions. Cadmium ions also induced the expression of ClpB of approximately 2.38 fold in TG1 expressing only OmpA. On the contrary, down-regulation of ClpB of up to 2.03 fold was observed when cells expressed polyhistidine on their surface membrane.
Figure 7. Quantitative proteomics analysis of proteins via 2D-DIGE.
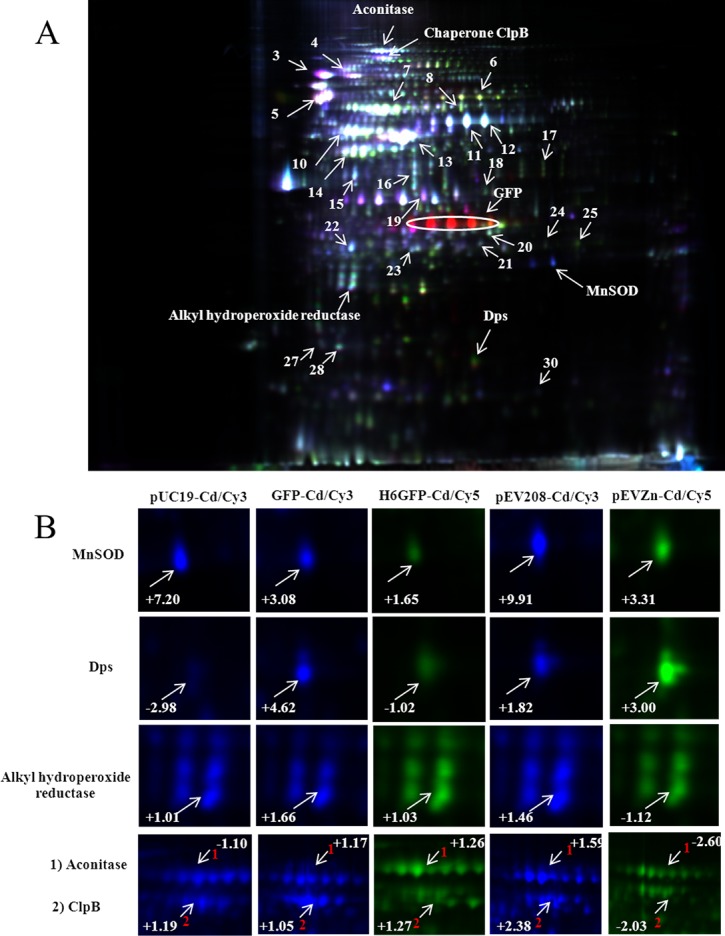
Two different samples were individually labeled with two kinds of fluorescent dyes (Cy3 and Cy5) and co-separated in one gel (A). An internal standard, derived from a mixture of all samples pooled together and labeled with Cy2, was created to facilitate the normalization of each spot among all gels. Protein spots with changes in abundance ratio of 1.5 fold and P values < 0.05 (Student’s t-test) were identified and compared between the gels (B).
Interconnection between proteins implicated in cadmium stress
Analysis using the STRING software provided insights on the biological inference of proteins implicated in cadmium stress. As shown in Fig. 8, most of the important proteins as identified by 2-DE (Figs. 4, 6, 7 and Table 2) were mapped onto the protein network. MnSOD encoded by the sodA gene is known to be regulated by the SoxRS regulon (Fig. 9). Even though, alkyl hydroperoxide reductase (coded by AhpC gene) is under the control of the OxyR regulon, however, these two enzymes have some cross-functional roles and have also the same connection point as mediated by the trxA gene (coding for thioredoxin). In addition, these enzymes also function together with the F0F1 ATP synthase (an enzyme that regulates proton transportation across the membrane and ATP synthesis/hydrolysis) linked via the trxA gene. MnSOD was also found to share connectivity with aconitase enzyme (coded by AcnA and AcnB). Particularly, aconitases serve as protective buffers against oxidative stress. AcnA enhances the stability of the sodA transcript whereas AcnB lowers its stability (Han & Lee, 2006). The OxyR also plays an important role on the regulation of DNA starvation/stationary phase protection protein (Dps). The Dps unspecifically binds and protects DNA from oxidative damage mediated by hydrogen peroxide (Han & Lee, 2006). Moreover, ClpB, which is known as a stress-induced multi-chaperone system, was also found to possess linkages with these proteins and enzymes. Interconnections between these proteins/enzymes provide new knowledge on the cellular responses of bacteria in the presence of cadmium stress.
Figure 8. Relationship between genes encoding proteins of E. coli involved in responses to cadmium stress as identified by the STRING software.

Figure 9. A proposed molecular mechanism of cellular responses and adaptation in the presence of toxic cadmium ions.

Responses against cadmium of E. coli cells expressing cytoplasmic His6GFP (A) and cells expressing His-OmpA (B).
Discussion
Herein, alterations of protein expression profiles of E. coli as consequences of cadmium stress have successfully been investigated using 2-DE and 2D-DIGE in conjunction with mass spectrometry-based protein identification. Differentially expressed proteins of TG1 expressing metal-binding regions in the cytoplasm and on the surface membrane provided insights into the underlying mechanisms of cellular adaptation against deleterious effects of cadmium ions (Fig. 9). In normal condition, Cd2+ is taken up by E. coli cells by an active transport system, in particular Mn2+ uptake system (Laddaga & Silver, 1985). Inside the cells, Cd2+ induces cell damage by generation of reactive oxygen species (ROS) through the electron transport chain (Pacheco et al., 2008; Thomas & Benov, 2018). The presence of superoxide radical () subsequently triggers the SoxRS regulon and results in the increased expression of antioxidative enzymes, mainly superoxide dismutase to protect the cells from cadmium stress (Geslin et al., 2001). In addition, the ROS also induces the OxyR and PerR regulons, which control the expression of catalase, peroxidases, alkyl hydroperoxide reductase enzymes. Toxic cadmium also stimulates the synthesis of cadmium-induced proteins (CDPs) such as DnaK, ClpB, RecA, UspA to form the cadmium stress stimulon (Ferianc, Farewell & Nyström, 1998; Han & Lee, 2006).
This study proposes a detailed explanation on the adaptive mechanisms of the E. coli host in response to toxic cadmium ions. From our findings, it can be suggested that cadmium afforded the growth arrest of control cells (TG1 harboring pUC19) (Fig. 2A). In order to survive, E. coli cells adapted themselves in several ways as follows:
-
(i)
Up-regulation of H+-transporting ATPase and ATP synthase F1 for controlling proton or ions translocation as well as for ATP production.
-
(ii)
Reduction of OmpA form 1 together with minor changes of the OmpC precursor and OmpF were detected since these outer membrane proteins were reported to participate in the homeostasis of divalent metal ions (Cd2+, Zn2+ and Cu2+) (Egler et al., 2005; Faber, Egli & Harder, 1993).
-
(iii)
Down-regulation of oligopeptide-binding protein might also be potentiated in order to reduce the uptake of cadmium ions into cells. It is known that the oligopeptide-binding protein is a component of the oligopeptide permease and the binding protein-dependent transport system (Han & Lee, 2006). In E. coli, the oligopeptide-binding protein (coded by the OppA gene) mainly functions as a mediator for the recycling of cell wall peptides (Wu & Mandrand-Berthelot, 1995). The protein displays quite a broad range of substrate specificity in which it can transport any peptide of a given length with a wide variety of amino acid composition. In some circumstances, the expression of oligopeptide-binding protein in E. coli also increased the uptake of aminoglycoside antibiotics (Acosta et al., 2005). Deletion of the OppA gene resulted in a decreased sensitivity against the aminoglycoside (Kashiwagi et al., 1992). Moreover, it cannot be excluded that the decreased amount of the oligopeptide-binding protein might be a result of oxidative stress. Since it has been reported to be one of the major oxidatively-damaged protein targets of E. coli in the presence of oxidative stress and iron overload (Tamarit, Cabiscol & Ros, 1998).
-
(iv)
Once cadmium ions crossed the membranes, aconitase B was found to be markedly decreased and damaged due to its unusual FeS cluster (Helbig, Grosse & Nies, 2008). Cadmium ions could also trigger the SoxRS regulon and consequently leads to the increased expression of MnSOD (+7.20 fold), which is involved as the primary stress defense mechanism. Our finding was in good agreement with other studies at the transcriptional and translational levels. It has previously been reported that the adapted strain of TG1, which could tolerate cadmium ions, exhibited different transcriptional profiles from the sensitive strain (Brocklehurst & Morby, 2000). The intracellular generation of superoxide radical by cadmium (Thomas & Benov, 2018), nickel and cobalt exerted toxic effects toward E. coli in which the participation of sod genes was regulated for the protection against metal stress (Geslin et al., 2001). Increased production of the SOD enzyme was found in the E. coli strain BL21 and P4 in the presence of cadmium (Khan et al., 2017; Shen et al., 2012).
-
(v)
In parallel, proteins involved in energy metabolism, e.g., glyceraldehyde-3-phosphate dehydrogenase and glycerol kinase, were down-regulated in order to reduce the energy consumption as required for cell survival. Therefore, the reduction of the oligopeptide-binding protein (mentioned in iii) and amino acid ABC transporter substrate-binding protein was observed for regulating the transportation of amino acid and other carbon sources during energy crisis (Isarankura-Na-Ayudhya et al., 2009).
To further confirm the aforementioned mechanisms, GFP harboring hexahistidine (His6GFP) was included to mediate the intracellular cadmium complexation (Fig. 9). Interestingly, low level expression of MnSOD (+1.65 fold) and AhpC (+1.03 fold) was detected. On the contrary, the up-regulation of both antioxidant enzymes (+3.08- and +1.66 fold) was found in cells expressing only native GFP (Fig. 7B). Our findings indicated that the effective complexation of cadmium ions by histidine residue provided protective roles against toxic metals, possibly by inhibition of ROS production. It has been detected that the presence of His6GFP rendered the cells to accumulate cadmium ions intracellularly up to 8.00 ± 0.08 nmole/8 × 108 cells or ∼1.5 fold higher than those of cells expressing native GFP (5.35 ± 0.45 nmole/8 × 108 cells) and TG1 host (5.34 ± 0.7 nmole/8 × 108 cells). Such protection coincided with the recovery of growth characteristics of His6GFP-expressing cells in the presence of cadmium (Fig. 2C). However, it should be noted that the overexpression of native GFP somewhat protected the E. coli cells from toxic cadmium (Fig. 2B). Plausible explanations might be attributable to (i) the quenching ability and SOD-like activity of the native GFP to detoxify the superoxide radical (Bou-Abdallah, Chasteen & Lesser, 2006) and (ii) the increased transcription (Helbig, Grosse & Nies, 2008) and translation of alkyl hydroperoxide reductase (AhpC) (Table 2 and Fig. 7B) under the activation of OxyR. This enzyme belongs to a large family of thiol-specific antioxidant proteins found in both prokaryotes and eukaryotes. It is known to detoxify not only peroxides but also reactive oxygen-, nitrogen- and sulfur-species. Cloning of this enzyme from Anabaena spp. in E. coli rendered cells to be more resistant against CdCl2 of up to 4 mM (Mishra, Chaurasia & Rai, 2009). In other organisms, increased levels of alkyl hydroperoxide reductase protected the plant pathogenic bacterium namely Xanthomonas campestris from peroxides and cadmium stress (Banjerdkij, Vattanaviboon & Mongkolsuk, 2005). Mutant cells of Helicobacter pylori defective in AhpC were more sensitive to oxidative stress conditions (Wang et al., 2005). In Saccharomyces cerevisiae, disruption of the AHP1 gene rendered cells to be more susceptible to several kinds of metal ions (Nguyen-nhu & Knoops, 2002).
Next, the blockage of metal uptake as a consequence of anchored polyhistidine residues on surface membrane has also been proven to confer protection against cadmium toxicity in E. coli (Fig. 5). Such protective effect is comparable to that observed in cells expressing His6GFP (Fig. 2C). Results from proteomics analysis revealed differences of protein expression profiles among these two cases as follows. Up-regulation of MnSOD, glyceraldehyde-3-phosphate dehydrogenase form 2, ribosomal protein L9 and OmpC precursor was found in cells expressing His-OmpA. An important item of note is that down-regulation and no increased expression of ATP synthase F1 and H+-transporting ATPase were observed. This infers that the binding of cadmium ions was effectively potentiated at the surface membrane in which the regulation of ions-transporting system was not induced. However, a minute amount of cadmium ions could pass through the membrane portion and exert their toxic effects to cells as observed by the increased expression of MnSOD. Moreover, expression of the OmpC precursor (membrane protein porin) proportionally corresponded with information from transcriptional profiles of Cd-resistant strain of E. coli (Brocklehurst & Morby, 2000). Meanwhile, a mutant strain lacking the OmpC gene was proven to be sensitive to cadmium ions (Egler et al., 2005). In another case, the expression of OmpA alone could also protect E. coli cells from cadmium-induced growth arrest (Fig. 5). However, the induction of antioxidative scavenging enzyme consisting of AhpC and ATP synthase F1 was still required (Table 2 and Fig. 7B). These results were in contrast to those observed in cells expressing His6GFP (Table 2 and Fig. 7B). Therefore, all findings lend support to the notion that metal complexation by cytoplasmic metal-binding protein afforded more efficiency to cope with cadmium stress than the metal-binding affinity at the surface membrane (Fig. 9).
Conclusion
This study explores the underlying mechanism and cellular responses of bacteria against toxic cadmium ions. In particular, E. coli TG1 expressing His6GFP and His-OmpA were used as models for studying the roles of cytoplasmic metal complexation and metal chelation at the surface membrane, respectively, upon exposure to cadmium stress (Fig. 9). Results from 2-DE and 2D-DIGE together with mass spectrometry-based protein identification had revealed that the complexation of cadmium ions by His6GFP helped to reduce toxicity of cadmium-induced oxidative cell damage via initial inhibition of ROS production. Supportive evidences could be accounted mainly by the low expression level of antioxidative enzymes and stress responsive proteins under the regulations of SoxRS and OxyR as compared to those derived from the effect of His-OmpA. Thus, it can be rationalized that the complex formation between cadmium ions and histidine-rich peptides/proteins provides more efficiency to cope with cadmium stress than the blockage of metal uptake at the surface membrane. Such findings shed light on the molecular mechanism and cellular adaptation of bacteria against cadmium toxicity.
Supplemental Information
All raw data and raw images of Figures presented in this study are provided herein this Supplementary ZIP file.
Funding Statement
This research is financially supported in part by the Young Research Scholar grant (no. MRG 5480205) from the Thailand Research Fund to Patcharee Isarankura-Na-Ayudhya and the annual governmental grant of Mahidol University under the National Research Universities Initiative. There was no additional external funding received for this study. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Additional Information and Declarations
Competing Interests
The authors declare there are no competing interests.
Author Contributions
Patcharee Isarankura-Na-Ayudhya conceived and designed the experiments, performed the experiments, analyzed the data, contributed reagents/materials/analysis tools, prepared figures and/or tables, authored or reviewed drafts of the paper, approved the final draft.
Chadinee Thippakorn, Nipawan Bunmee and Suchitra Sawangnual performed the experiments, authored or reviewed drafts of the paper, approved the final draft.
Supitcha Pannengpetch performed the experiments, analyzed the data, prepared figures and/or tables, authored or reviewed drafts of the paper, approved the final draft.
Sittiruk Roytrakul performed the experiments, analyzed the data, contributed reagents/materials/analysis tools, authored or reviewed drafts of the paper, approved the final draft.
Chartchalerm Isarankura-Na-Ayudhya conceived and designed the experiments, analyzed the data, prepared figures and/or tables, authored or reviewed drafts of the paper, approved the final draft.
Virapong Prachayasittikul analyzed the data, contributed reagents/materials/analysis tools, authored or reviewed drafts of the paper, approved the final draft.
Data Availability
The following information was supplied regarding data availability:
The raw data are provided in the Supplementary File.
References
- Acosta et al. (2005).Acosta MB, Ferreira RC, Ferreira LC, Costa SO. Intracellular polyamine pools, oligopeptide-binding protein A expression, and resistance to aminoglycosides in Escherichia coli. Memorias Do Instituto Oswaldo Cruz. 2005;100:789–793. doi: 10.1590/S0074-02762005000700020. [DOI] [PubMed] [Google Scholar]
- Banjerdkij, Vattanaviboon & Mongkolsuk (2005).Banjerdkij P, Vattanaviboon P, Mongkolsuk S. Exposure to cadmium elevates expression of genes in the OxyR and OhrR regulons and induces cross-resistance to peroxide killing treatment in Xanthomonas campestris. Applied and Environmental Microbiology. 2005;71:1843–1849. doi: 10.1128/AEM.71.4.1843-1849.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bischoff (1982).Bischoff B. Effects of cadmium on microorganisms. Ecotoxicology and Environmental Safety. 1982;6:157–165. doi: 10.1016/0147-6513(82)90003-3. [DOI] [Google Scholar]
- Bou-Abdallah, Chasteen & Lesser (2006).Bou-Abdallah F, Chasteen ND, Lesser MP. Quenching of superoxide radicals by green fluorescent protein. Biochimica Et Biophysica Acta. 2006;1760:1690–1695. doi: 10.1016/j.bbagen.2006.08.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brocklehurst & Morby (2000).Brocklehurst KR, Morby AP. Metal-ion tolerance in Escherichia coli: analysis of transcriptional profiles by gene-array technology. Microbiology. 2000;146:2277–2282. doi: 10.1099/00221287-146-9-2277. [DOI] [PubMed] [Google Scholar]
- Cohen, Bitan & Nitzan (1991).Cohen I, Bitan R, Nitzan Y. The effect of zinc and cadmium ions on Escherichia coli B. Microbios. 1991;68:157–168. [PubMed] [Google Scholar]
- David et al. (2003).David G, Blondeau K, Schiltz M, Penel S, Lewit-Bentley A. YodA from Escherichia coli is a metal-binding, lipocalin-like protein. Journal of Biological Chemistry. 2003;278:43728–43735. doi: 10.1074/jbc.M304484200. [DOI] [PubMed] [Google Scholar]
- Egler et al. (2005).Egler M, Grosse C, Grass G, Nies DH. Role of the extracytoplasmic function protein family sigma factor RpoE in metal resistance of Eschenchia coli. Journal of Bacteriology. 2005;187:2297–2307. doi: 10.1128/JB.187.7.2297-2307.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Faber, Egli & Harder (1993).Faber F, Egli T, Harder W. Transient repression of the synthesis of OmpF and aspartate transcarbamoylase in Escherichia coli K12 as a response to pollutant stress. FEMS Microbiology Letters. 1993;111:189–195. doi: 10.1111/j.1574-6968.1993.tb06384.x. [DOI] [PubMed] [Google Scholar]
- Ferianc, Farewell & Nyström (1998).Ferianc P, Farewell A, Nyström T. The cadmium-stress stimulon of Escherichia coli K-12. Microbiology. 1998;144:1045–1050. doi: 10.1099/00221287-144-4-1045. [DOI] [PubMed] [Google Scholar]
- Geslin et al. (2001).Geslin C, Llanos J, Prieur D, Jeanthon C. The manganese and iron superoxide dismutases protect Escherichia coli from heavy metal toxicity. Research in Microbiology. 2001;152:901–905. doi: 10.1016/S0923-2508(01)01273-6. [DOI] [PubMed] [Google Scholar]
- Giovanella et al. (2017).Giovanella P, Cabral L, Costa AP, De Oliveira Camargo FA, Gianello C, Bento FM. Metal resistance mechanisms in Gram-negative bacteria and their potential to remove Hg in the presence of other metals. Ecotoxicology and Environmental Safety. 2017;140:162–169. doi: 10.1016/j.ecoenv.2017.02.010. [DOI] [PubMed] [Google Scholar]
- Han & Lee (2006).Han MJ, Lee SY. The Escherichia coli proteome: past, present, and future prospects. Microbiology and Molecular Biology Reviews. 2006;70:362–439. doi: 10.1128/MMBR.00036-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He et al. (2014).He Y, Ma W, Li Y, Liu J, Jing W, Wang L. Expression of metallothionein of freshwater crab (Sinopotamon henanense) in Escherichia coli enhances tolerance and accumulation of zinc, copper and cadmium. Ecotoxicology. 2014;23:56–64. doi: 10.1007/s10646-013-1151-0. [DOI] [PubMed] [Google Scholar]
- Helbig, Grosse & Nies (2008).Helbig K, Grosse C, Nies DH. Cadmium toxicity in glutathione mutants of Escherichia coli. Journal of Bacteriology. 2008;190:5439–5454. doi: 10.1128/JB.00272-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hossain, Mallick & Mukherjee (2012).Hossain ST, Mallick I, Mukherjee SK. Cadmium toxicity in Escherichia coli: cell morphology, Z-ring formation and intracellular oxidative balance. Ecotoxicology and Environmental Safety. 2012;86:54–59. doi: 10.1016/j.ecoenv.2012.09.017. [DOI] [PubMed] [Google Scholar]
- Hou, Kim & Kim (1988).Hou YM, Kim R, Kim SH. Expression of the mouse metallothionein-I gene in Escherichia coli: increased tolerance to heavy metals. Biochimica Et Biophysica Acta—Gene Structure and Expression. 1988;951:230–234. doi: 10.1016/0167-4781(88)90045-0. [DOI] [PubMed] [Google Scholar]
- Isarankura-Na-Ayudhya et al. (2009).Isarankura-Na-Ayudhya P, Isarankura-Na-Ayudhya C, Treeratanapaiboon L, Kasikun K, Thipkeaw K, Prachayasittikul V. Proteomic profiling of Escherichia coli in response to heavy metals stress. European Journal of Scientific Research. 2009;25:679–688. [Google Scholar]
- Isarankura-Na-Ayudhya et al. (2010).Isarankura-Na-Ayudhya P, Isarankura-Na-Ayudhya C, Yainoy S, Thippakorn C, Singhagamol W, Polprachum W, Roytrakul S, Prachayasittikul V. Proteomic alterations of Escherichia coli by paraquat. EXCLI Journal. 2010;9:108–118. [PMC free article] [PubMed] [Google Scholar]
- Isarankura-Na-Ayudhya et al. (2005).Isarankura-Na-Ayudhya C, Suwanwong Y, Boonpangrak S, Kiatfuengfoo R, Prachayasittikul V. Co-expression of zinc binding motif and GFP as a cellular indicator of metal ions mobility. International Journal of Biological Sciences. 2005;1:146–151. doi: 10.7150/ijbs.1.146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kashiwagi et al. (1992).Kashiwagi K, Miyaji A, Ikeda S, Tobe T, Sasakawa C, Igarashi K. Increase of sensitivity to aminoglycoside antibiotics by polyamine-induced protein (oligopeptide-binding protein) in Escherichia coli. Journal of Bacteriology. 1992;174:4331–4337. doi: 10.1128/jb.174.13.4331-4337.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kershaw, Brown & Hobman (2007).Kershaw CJ, Brown NL, Hobman JL. Zinc dependence of zinT (yodA) mutants and binding of zinc, cadmium and mercury by ZinT. Biochemical and Biophysical Research Communications. 2007;364:66–71. doi: 10.1016/j.bbrc.2007.09.094. [DOI] [PubMed] [Google Scholar]
- Khan et al. (2017).Khan Z, Rehman A, Nisar MA, Zafar S, Zerr I. Biosorption behavior and proteomic analysis of Escherichia coli P4 under cadmium stress. Chemosphere. 2017;174:136–147. doi: 10.1016/j.chemosphere.2017.01.132. [DOI] [PubMed] [Google Scholar]
- Khazaeli & Mitra (1981).Khazaeli MB, Mitra RS. Cadmium-binding component in Escherichia coli during accommodation to low levels of this ion. Applied and Environmental Microbiology. 1981;41:46–50. doi: 10.1128/aem.41.1.46-50.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kille et al. (1990).Kille P, Stephens P, Cryer A, Kay J. The expression of a synthetic rainbow trout metallothionein gene in E. coli. Biochimica Et Biophysica Acta—Gene Structure and Expression. 1990;1048:178–186. doi: 10.1016/0167-4781(90)90054-6. [DOI] [PubMed] [Google Scholar]
- Kondo et al. (2000).Kondo A, Kohda J, Endo Y, Shiromizu T, Kurokawa Y, Nishihara K, Yanagi H, Yura T, Fukuda H. Improvement of productivity of active horseradish peroxidase in Escherichia coli by coexpression of Dsb proteins. Journal of Bioscience and Bioengineering. 2000;90:600–606. doi: 10.1016/S1389-1723(00)90003-3. [DOI] [PubMed] [Google Scholar]
- Laddaga & Silver (1985).Laddaga RA, Silver S. Cadmium uptake in Escherichia coli K-12. Journal of Bacteriology. 1985;162:1100–1105. doi: 10.1128/jb.162.3.1100-1105.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LaRossa, Smulski & Van Dyk (1995).LaRossa RA, Smulski DR, Van Dyk TK. Interaction of lead nitrate and cadmium chloride with Escherichia coli K-12 and Salmonella typhimurium global regulatory mutants. Journal of Industrial Microbiology. 1995;14:252–258. doi: 10.1007/BF01569936. [DOI] [PubMed] [Google Scholar]
- Lausova, Ferianc & Polek (1999).Lausova A, Ferianc P, Polek B. The effect of different oxidative challenge on growth and stress protein induction in Escherichia coli. Biologia. 1999;54:649–660. [Google Scholar]
- Lim et al. (2009).Lim ES, Le TH, Lee SH, Kim YH, Min J. Stress responses analysis of As(III), Cd and Pb in Escherichia coli. Toxicology and Environmental Health Sciences. 2009;1:192–195. doi: 10.1007/BF03216485. [DOI] [Google Scholar]
- Mahnam et al. (2018).Mahnam K, Foruzandeh S, Mirakhorli N, Saffar B. Experimental and theoretical studies of cadmium ions absorption by a new reduced recombinant defensin. Journal of Biomolecular Structure & Dynamics. 2018;36:2004–2014. doi: 10.1080/07391102.2017.1340851. [DOI] [PubMed] [Google Scholar]
- Mejare, Ljung & Bulow (1998).Mejare M, Ljung S, Bulow L. Selection of cadmium specific hexapeptides and their expression as OmpA fusion proteins in Escherichia coli. Protein Engineering. 1998;11:489–494. doi: 10.1093/protein/11.6.489. [DOI] [PubMed] [Google Scholar]
- Mishra, Chaurasia & Rai (2009).Mishra Y, Chaurasia N, Rai LC. AhpC (alkyl hydroperoxide reductase) from Anabaena sp. PCC 7120 protects Escherichia coli from multiple abiotic stresses. Biochemical and Biophysical Research Communications. 2009;381:606–611. doi: 10.1016/j.bbrc.2009.02.100. [DOI] [PubMed] [Google Scholar]
- Mitra (1984).Mitra RS. Protein synthesis in Escherichia coli during recovery from exposure to low levels of Cd2+ Applied and Environmental Microbiology. 1984;47:1012–1016. doi: 10.1128/aem.47.5.1012-1016.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitra & Bernstein (1978).Mitra RS, Bernstein IA. Single strand breakage in DNA of Escherichia coli exposed to Cd2 + Journal of Bacteriology. 1978;133:75–80. doi: 10.1128/jb.133.1.75-80.1978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitra et al. (1975).Mitra RS, Gray RH, Chin B, Bernstein IA. Molecular mechanisms of accommodation in Escherichia coli to toxic levels of Cd2+ Journal of Bacteriology. 1975;121:1180–1188. doi: 10.1128/jb.121.3.1180-1188.1975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nair & Robinson (2001).Nair PS, Robinson WE. Cadmium binding to a histidine-rich glycoprotein from marine mussel blood plasma: potentiometric titration and equilibrium speciation modeling. Environmental Toxicology and Chemistry. 2001;20:1596–1604. doi: 10.1002/etc.5620200726. [DOI] [PubMed] [Google Scholar]
- Nguyen-nhu & Knoops (2002).Nguyen-nhu NT, Knoops B. Alkyl hydroperoxide reductase 1 protects Saccharomyces cerevisiae against metal ion toxicity and glutathione depletion. Toxicology Letters. 2002;135:219–228. doi: 10.1016/S0378-4274(02)00280-1. [DOI] [PubMed] [Google Scholar]
- Nies (1992).Nies DH. Resistance to cadmium, cobalt, zinc, and nickel in microbes. Plasmid. 1992;27:17–28. doi: 10.1016/0147-619X(92)90003-S. [DOI] [PubMed] [Google Scholar]
- Nies (1999).Nies DH. Microbial heavy-metal resistance. Applied Microbiology and Biotechnology. 1999;51:730–750. doi: 10.1007/s002530051457. [DOI] [PubMed] [Google Scholar]
- Pacheco et al. (2008).Pacheco CC, Passos JF, Castro AR, Moradas-Ferreira P, De Marco P. Role of respiration and glutathione in cadmium-induced oxidative stress in Escherichia coli K-12. Archives of Microbiology. 2008;189:271–278. doi: 10.1007/s00203-007-0316-8. [DOI] [PubMed] [Google Scholar]
- Patel et al. (2010).Patel J, Zhang Q, McKay RM, Vincent R, Xu Z. Genetic engineering of Caulobacter crescentus for removal of cadmium from water. Applied Biochemistry and Biotechnology. 2010;160:232–243. doi: 10.1007/s12010-009-8540-0. [DOI] [PubMed] [Google Scholar]
- Puskárová et al. (2002).Puskárová A, Ferianc P, Kormanec J, Homerová D, Farewell A, Nyström T. Regulation of yodA encoding a novel cadmium-induced protein in Escherichia coli. Microbiology. 2002;148:3801–3811. doi: 10.1099/00221287-148-12-3801. [DOI] [PubMed] [Google Scholar]
- Romeyer et al. (1988).Romeyer FM, Jacobs FA, Masson L, Hanna Z, Brousseau R. Bioaccumulation of heavy metals in Escherichia coli expressing an inducible synthetic human metallothionein gene. Journal of Biotechnology. 1988;8:207–220. doi: 10.1016/0168-1656(88)90003-X. [DOI] [Google Scholar]
- Samuelson et al. (2000).Samuelson P, Wernerus H, Svedberg M, Stahl S. Staphylococcal surface display of metal-binding polyhistidyl peptides. Applied and Environmental Microbiology. 2000;66:1243–1248. doi: 10.1128/AEM.66.3.1243-1248.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider, Rasband & Eliceiri (2012).Schneider CA, Rasband WS, Eliceiri KW. NIH Image to ImageJ: 25 years of image analysis. Nature Methods. 2012;9(7):671–675. doi: 10.1038/nmeth.2089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shapiro & Keasling (1996).Shapiro N, Keasling JD. The recA gene and cadmium toxicity in Escherichia coli K12. Microbios. 1996;86:23–26. [PubMed] [Google Scholar]
- Shen et al. (2012).Shen JJ, Qiao ZJ, Xing TJ, Zhang LP, Liang YL, Jin ZP, Yang GD, Wang R, Pei YX. Cadmium toxicity is alleviated by AtLCD and AtDCD in Escherichia coli. Journal of Applied Microbiology. 2012;113:1130–1138. doi: 10.1111/j.1365-2672.2012.05408.x. [DOI] [PubMed] [Google Scholar]
- Sousa, Cebolla & De Lorenzo (1996).Sousa C, Cebolla A, De Lorenzo V. Enhanced metalloadsorption of bacterial cells displaying poly-His peptides. Nature Biotechnology. 1996;14:1017–1020. doi: 10.1038/nbt0896-1017. [DOI] [PubMed] [Google Scholar]
- Stojnev et al. (2007).Stojnev T, Harichová J, Ferianc P, Nyström T. Function of a novel cadmium-induced YodA protein in Escherichia coli. Current Microbiology. 2007;55:99–104. doi: 10.1007/s00284-006-0516-5. [DOI] [PubMed] [Google Scholar]
- Tamarit, Cabiscol & Ros (1998).Tamarit J, Cabiscol E, Ros J. Identification of the major oxidatively damaged proteins in Escherichia coli cells exposed to oxidative stress. The Journal of Biological Chemistry. 1998;273:3027–3032. doi: 10.1074/jbc.273.5.3027. [DOI] [PubMed] [Google Scholar]
- Thomas & Benov (2018).Thomas M, Benov L. The Contribution of Superoxide Radical to Cadmium Toxicity in E. coli. Biological Trace Element Research. 2018;181:361–368. doi: 10.1007/s12011-017-1048-5. [DOI] [PubMed] [Google Scholar]
- Vallee & Ulmer (1972).Vallee BL, Ulmer DD. Biochemical effects of mercury, cadmium, and lead. Annual Review of Biochemistry. 1972;41:91–128. doi: 10.1146/annurev.bi.41.070172.000515. [DOI] [PubMed] [Google Scholar]
- Van Dyk et al. (1995).Van Dyk TK, Smulski DR, Reed TR, Belkin S, Vollmer AC, LaRossa RA. Responses to toxicants of an Escherichia coli strain carrying a uspA’::lux genetic fusion and an E. coli strain carrying a grpE’::lux fusion are similar. Applied and Environmental Microbiology. 1995;61:4124–4127. doi: 10.1128/aem.61.11.4124-4127.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang et al. (2005).Wang G, Conover RC, Olczak AA, Alamuri P, Johnson MK, Maier RJ. Oxidative stress defense mechanisms to counter iron-promoted DNA damage in Helicobacter pylori. Free Radical Research. 2005;39:1183–1191. doi: 10.1080/10715760500194018. [DOI] [PubMed] [Google Scholar]
- Westermeier & Scheibe (2008).Westermeier R, Scheibe B. Difference gel electrophoresis based on lys/cys tagging. Methods in Molecular Biology. 2008;424:73–85. doi: 10.1007/978-1-60327-064-9_7. [DOI] [PubMed] [Google Scholar]
- Wu & Mandrand-Berthelot (1995).Wu LF, Mandrand-Berthelot MA. A family of homologous substrate-binding proteins with a broad range of substrate specificity and dissimilar biological functions. Biochimie. 1995;77:744–750. doi: 10.1016/0300-9084(96)88192-2. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
All raw data and raw images of Figures presented in this study are provided herein this Supplementary ZIP file.
Data Availability Statement
The following information was supplied regarding data availability:
The raw data are provided in the Supplementary File.